
Abstract
Tendon injuries occur frequently in horses and have a poor capacity to regenerate, which leads to high re-injury rates. Equine embryo-derived stem cells (ESCs) survive in high numbers in the injured horse tendon and we hypothesized that they differentiate into tenocytes in vivo. Immunocytochemistry revealed that in the injured horse tendon ESCs express the tendon progenitor marker scleraxis and that there is a local upregulation of the transforming growth factor-β (TGF-β) at the injury site. The aim of this study was to determine if TGF-β signaling was able to drive tenocyte differentiation by ESCs. Exposure of differentiating ESCs to TGF-β in vitro produced an upregulation of scleraxis at the gene and protein level with the greatest effect being produced in the presence of TGF-β3. TGF-β3 treatment of differentiating ESCs also promotes a significant upregulation of other tendon-associated genes and proteins suggesting it can promote ESC differentiation into tenocytes. Our results demonstrate that equine ESCs can differentiate into a therapeutically relevant cell type and that TGF-β driven differentiation of ESCs may provide a model to study tendon development and better understand the transcriptional networks that are involved in equine tendon cell differentiation from the early embryonic stages.
Introduction
Autologous mesenchymal stromal cells (MSCs) have been used clinically in the horse since 2003 3 and some positive results have been reported experimentally with improvements in repair occurring at early time points in comparison to nonstem cell-treated controls. 4 Clinical results are also promising, 5 but no placebo-controlled clinical trial has been performed to date. For autologous cell therapy, every horse must undergo an invasive procedure to isolate a tissue sample from which the stem cells can be derived. The derivation of MSCs from the bone marrow requires that a bone marrow aspirate be taken, which has been associated with pneumopericardium. 6 It then takes 2–4 weeks to culture sufficient cells for therapeutic use, resulting in a delay between injury and the injection of the cells into the damaged tendon, which prevents the immediate treatment of acute injuries.
Equine embryo-derived stem cells (ESCs) have been derived and characterized previously.7,8 These cells can be grown extensively, while retaining ESC marker gene expression and the potential to differentiate in vitro into derivatives of the three germ layers. However, unlike their human and mouse equivalents, equine ESCs have not been shown to form teratomas following their injection into severe combined immunodeficient (SCID) mice. Due to their extensive expansion capacity and ability to undergo multilineage differentiation, the cells could provide a standardized off the shelf source of allogeneic cells for treating horse tendon injuries. Following their injection into surgically created regions of damage in the horse tendon, equine ESCs survive in high and constant numbers over a 90-day period and do not form any detectable tumors or elicit a detectable cell-mediated immune response. 9 It was therefore possible that the ESCs may have differentiated into tenocytes.
Determining tenocyte differentiation is complex due to a lack of specific markers. The bHLH transcription factor scleraxis has been shown to mark tendon progenitors in the developing mouse 10 and its overexpression in MSCs leads to tenocyte differentiation. 11 Scleraxis also has a nuclear localization making it possible to look for colocalization with the β-galactosidase reporter gene expressed by the injected ESCs. In light of the finding that scleraxis is upregulated in response to the transforming growth factor-β (TGF-β) 12 and that TGF-β has been reported to be upregulated after tendon injury,13–15 we hypothesized that equine ESCs can differentiate into tenocytes in response to TGF-β signaling.
Materials and Methods
Cell culture
Three lines of previously characterized ESCs7–9 were used in this study. ESCs were cultured on mitotically inactivated mouse embryonic fibroblasts at 37.5°C, 5% CO2 in the ESC medium (DMEM/F12 containing 15% fetal bovine serum, 2 mM L-glutamine, 1% nonessential amino acids, 1 mM sodium pyruvate, and 0.1 mM 2-mercaptoethanol (all from Invitrogen, Renfrewshire, United Kingdom) and 1000 units/mL of the leukemia inhibitory factor (LIF) (Sigma, Dorset, United Kingdom). ESCs were passaged mechanically every 5–7 days in the presence of Effectine (PAA laboratories, Somerset, United Kingdom). ESCs were used at passage 12–18.
Equine tenocytes were isolated from healthy equine tendon tissue at postmortem with institutional ethics approval from the Animal Health Trust Research Ethics Committee. Briefly, the tissue was cut into small pieces, and then incubated overnight in 1 mg/mL type I collagenase produced by the Clostridium histolyticum (Sigma) solution in tenocyte culture media (DMEM high glucose, 10% fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (all from Invitrogen)). The cells were pelleted by centrifugation and washed three times in cell culture media before culture. Equine tenocytes were passaged using 0.25% trypsin-EDTA (Sigma) every 3–4 days just before reaching confluency. Tenocytes were used at passage 2–5.
To induce differentiation, ESCs were passaged into conditions without feeder cells in ESC differentiation media (as for the ESC medium described above, but without LIF). Cells were allowed to attach for 5 days before the addition of 20 ng/mL TGF-β1, 2, or 3 (Peprotech, London, United Kingdom). This concentration was selected based on that used to induce scleraxis in mouse mesenchymal progenitor cells. 12 For tenocytes, 20 ng/mL TGF-β1, 2, or 3 was added 1 day after passaging in the standard tenocyte media described above. All cells were harvested after 1, 3, 7, or 14 days for RNA or immunocytochemistry described below. During this time, the media were changed every 2–3 days and fresh TGF-β added.
Dual immunohistochemistry on tissue sections
Frozen tendon tissue sections, 14 μm thick, were prepared from tendons that had been injected with ESCs and collected 10–90 days postmortem as described in our previous study. 9 They had been fixed in 100% acetone for 10 min and stored at−20°C until used. For dual immunohistochemistry, the sections were incubated both separately and simultaneously with the primary antibodies at 4°C overnight before detection with fluorescently labeled secondary antibodies. All antibodies were used at optimized concentrations in phosphate-buffered saline (PBS). Primary antibodies included chicken anti-β-galactosidase 1:10,000 (Abcam, Cambridgeshire, United Kingdom), rabbit anti-β-galactosidase 1:1000 (Abcam), rabbit anti-scleraxis 1:100 (Abcam), and goat anti-scleraxis 1:200 (Santa Cruz, Heidelberg, Germany). Secondary antibodies included goat anti-chicken conjugated to FITC 1:100 (Jackson ImmunoResearch, Suffolk, United Kingdom), goat anti-rabbit conjugated to Texas Red 1:200 (Abcam), donkey anti-goat conjugated to FITC 1:200 (Abcam), and donkey anti-rabbit conjugated to Texas Red 1:200 (Abcam). All antibodies used were polyclonal IgGs, with the exception of the chicken anti-β-galactosidase, which is a polyclonal IgY. Combinations of chicken anti-β-galactosidase/rabbit anti-scleraxis and rabbit anti-β-galactosidase/goat anti-scleraxis were used to ensure that results were verified using two antibodies for each protein. Negative controls were carried out using isotype-matched sham primary antibodies and by omitting the primary antibodies. All sections were mounted with the Vectashield Hardset mounting medium containing DAPI (Vector Laboratories, Peterborough, United Kingdom).
Single immunohistochemistry on tissue sections
Tissue sections were prepared and stored as described above. For single immunohistochemistry, sections were incubated with the primary antibodies for 1 h at room temperature, which were then detected using the peroxidase ABC detection system (Vector Laboratories, Peterborough, United Kingdom) and DAB substrate (DakoCytomation, Ely, United Kingdom). The primary antibodies included rabbit anti-TGFβ 1:100 (Novus Biologicals, Cambridgeshire, United Kingdom) and rabbit anti-scleraxis 1:100 (Abcam). The TGFβ antibody used cross reacted with TGFβ1, 2, and 3.
Immunocytochemistry on cells
Cells to be used in immunocytochemistry were grown in 24-well plates on gelatin-coated (Sigma) coverslips. The cells were fixed in 3% paraformaldehyde in PBS for 20 min at room temperature and permeablized for 1 h with 0.1% triton-X-100 at room temperature. Primary antibody incubations were carried out overnight at 4°C before detection with the fluorescently labeled secondary antibody. Primary antibodies included rabbit anti-scleraxis 1:100 (Abcam), rabbit anti-TGF-β receptor I 1:100, rabbit anti-TGF-β receptor II 1:100 (both Santa Cruz), rabbit anti-tenascin C 1:100 (Abcam), rabbit anti-tenomodulin 1:100 (Santa Cruz), mouse anti-collagen I 1:100 (Abcam), rabbit anti-COMP 1:500 (kindly provided by Professor Roger Smith, Royal Veterinary College, United Kingdom), and rabbit anti-thrombospondin 4 1:100 (Santa Cruz). Secondary antibodies included goat anti-rabbit Texas Red 1:200 (Abcam), goat anti-mouse FITC 1:200 (Abcam), and goat anti-rabbit FITC 1:100 (Sigma). Immunocytochemistry was performed on three lines of cells and representative data are shown.
Western blot
To confirm that the antibodies could recognize the equine proteins, Western blots were performed using rabbit anti-scleraxis (Abcam; 1:500), rabbit anti-TGF-β (Novus Biologicals, Cambridgeshire, United Kingdom; 1:250), rabbit anti-tenascin C (Abcam; 1:1000), rabbit anti-tenomodulin (Santa Cruz, 1:500), mouse anti-collagen I (Abcam, 1:3000), and rabbit anti-thrombospondin 4 (Santa Cruz, 1:1500). Whole cell extract was isolated from homogenized healthy equine tendon tissue (taken at postmortem from horses euthanized for reasons unrelated to this study) by three rounds of freeze–thaw in an extraction buffer (20 mM Hepes pH7.9, 450 mM NaCl, 0.4 mM EDTA, 25% glycerol, 1 mM PMSF) and supernatants were collected by centrifugation. Twenty micrograms of a denatured protein was run on sodium dodecyl sulfate polyacrylamide gels and transferred to a polyvinylidene fluoride membrane with the exception of collagen I. For collagen I, 20 μg of the nondenatured protein was run under nonreducing conditions as the antibody only recognizes the native protein. Immunoreactivity was detected using the electrochemiluminescence plus detection system (Amersham, Buckinghamshire, United Kingdom). The mouse anti-β actin antibody (Abcam, 1:1000) was used as a positive control.
RNA extraction and cDNA synthesis
RNA was extracted using Tri-reagent (Sigma) and treated with Ambion DNA-free (Life Technologies, Paisley, United Kingdom) according to the manufacturer's instructions. cDNA was made from 2 μg of RNA using Moloney murine leukemia virus reverse transcriptase (Promega, Hampshire, United Kingdom) and oligo(dT) and random hexamers as primers (both Promega). Two microliter aliquots of cDNA were used in quantitative polymerase chain reaction (qPCR)
Quantitative PCR
Primers were designed using primer3 (http://frodo.wi.mit.edu/primer3) and mfold (http://mfold.rna.albany.edu/?q=mfold/DNA-Folding-Form) programs to obtain amplicons with a melting temperature (Tm) of 58°C–62°C, devoid a secondary structure at Tm 60°C and with an amplicon size of 50–150 bp. Primer sequences are available on request. qPCR was carried out using SYBR Green containing supermix (Bioline, London, United Kingdom) on a Quantica machine (Techne, Staffordshire, United Kingdom). All PCR reactions were performed in duplicate. PCR cycle parameters were 95°C 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 15 s, and 72°C for 15 s. At the end of the program, the temperature was reduced to 65°C, and then gradually increased by 1°C increments up to 95°C to produce a melt curve. Gene expression was normalized to 18 s rRNA gene expression levels, which did not change between treatments (data not shown), using the 2−ΔΔCt method. 16 A student's t-test was used to determine statistically significant fold changes in gene expression between the control and treated groups at each time point. qPCR was performed on three lines of ESCs and tenocytes with two technical repeats carried out for each cell line.
Results
Equine ESCs in vivo express the tendon progenitor marker scleraxis
We have previously demonstrated that equine ESCs injected into the damaged horse tendon survive in high and constant numbers for at least 90 days. 9 In this study, we used dual immunohistochemistry for the β-galactosidase reporter gene and the transcription factor scleraxis (Fig. 1). This demonstrated that all of the β-galactosidase-positive cells coexpressed scleraxis up to 90 days postinjection indicating that all of the injected ESCs have turned on scleraxis. However, not all scleraxis-positive cells were β-galactosidase-positive indicating that there is a population of endogenous scleraxis-positive cells, which do not arise from the ESCs and appear after an injury (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
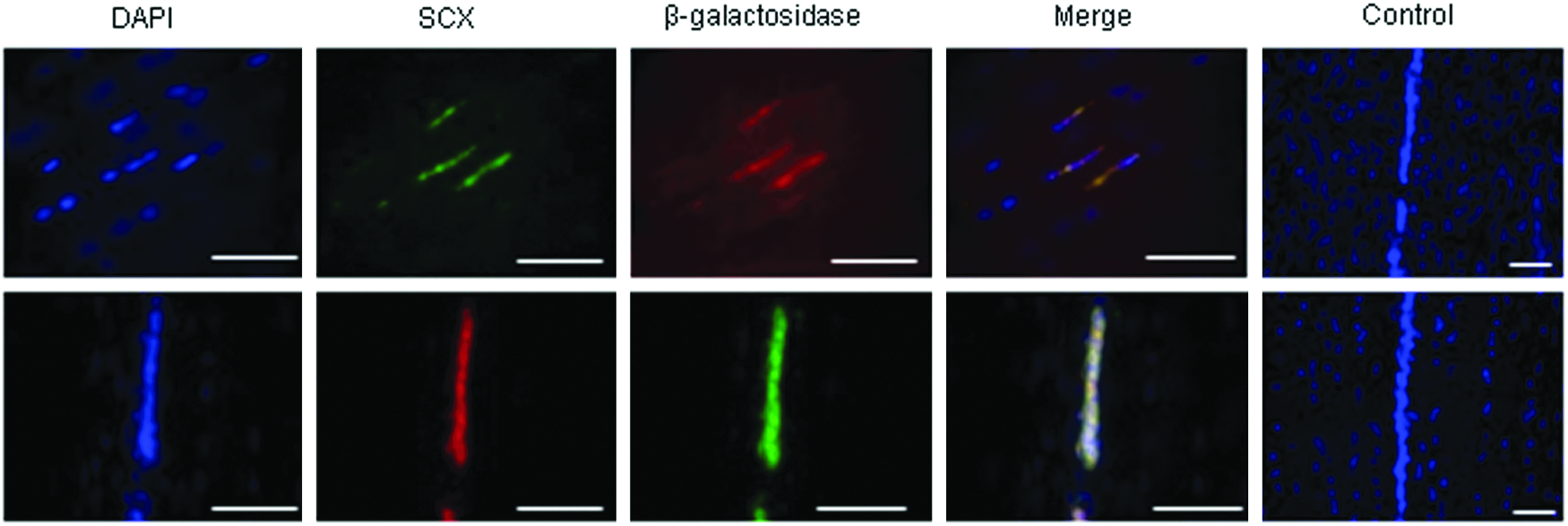
Dual immunofluorescence demonstrates that in the injured horse tendon, β-galactosidase-labeled embryo-derived stem cells (ESCs) coexpress the tendon progenitor marker scleraxis (SCX). DAPI staining of the nuclei is shown in blue. Scale bar=200 μm. In this figure, the top line shows goat anti-scleraxis and rabbit anti-β-galactosidase detected with donkey anti-goat FITC and donkey anti-rabbit Texas Red. The bottom line shows chicken anti-β-galactosidase and rabbit anti-scleraxis detected with goat anti-chicken FITC and goat anti-rabbit Texas Red. Merged images from the negative controls are shown in the end panels. Color images available online at www.liebertpub.com/tea
Scleraxis and TGF-β are both upregulated in the injured horse tendon
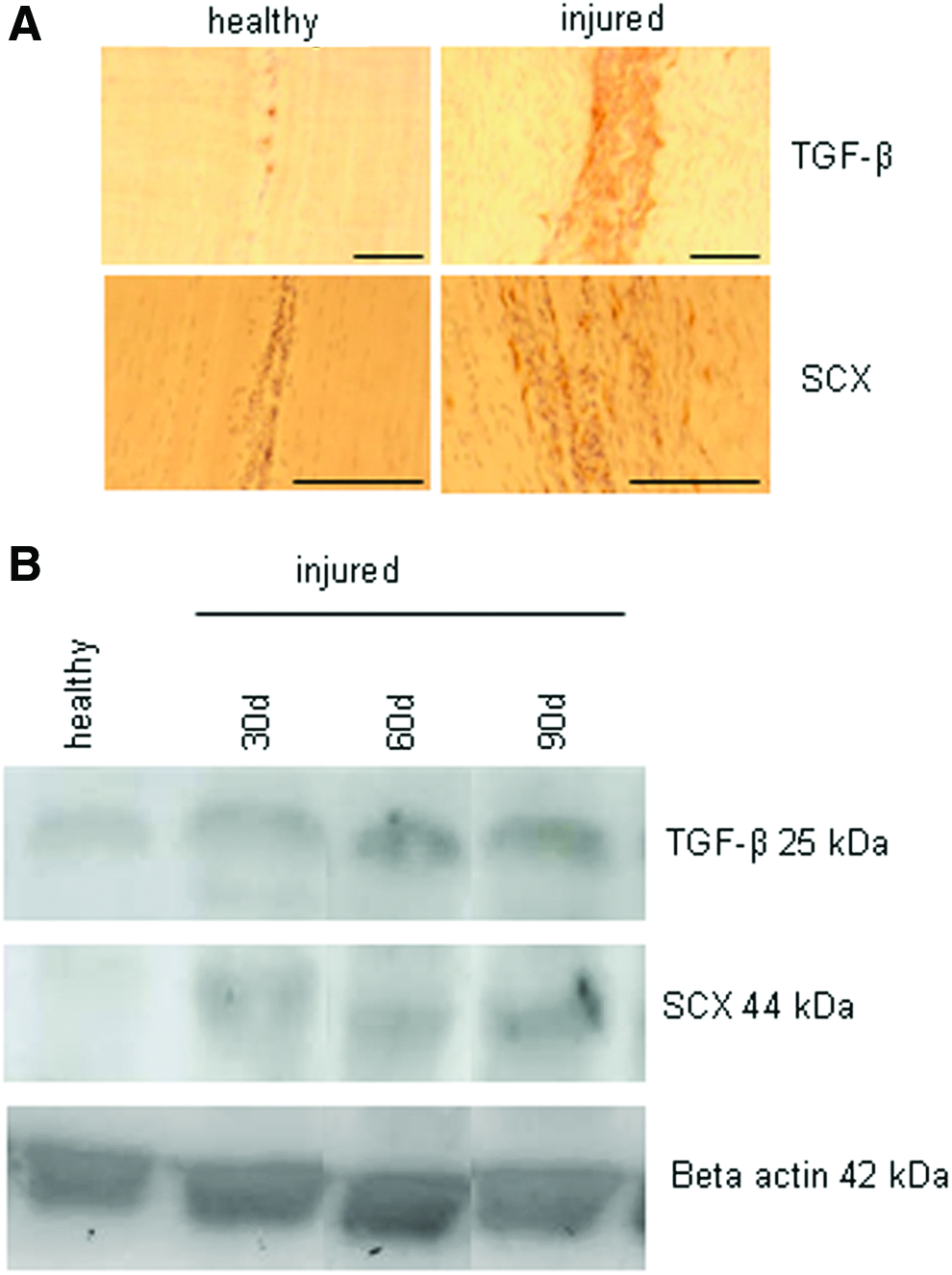
Immunohistochemistry for scleraxis in the healthy adult horse tendon produced little staining. However, in the damaged adult tendon, scleraxis-positive cells were present in and around the site of damage (Fig. 2A). This occurred whether or not stem cells had been injected at the site confirming that a population of scleraxis-positive endogenous cells is present following tendon tissue damage. Western blot analysis confirmed that scleraxis levels were increased in the damaged tendon relative to healthy tendon at all time points analyzed (Fig. 2B). Results for TGF-β were similar to those found for scleraxis. In the healthy tendon, only very low levels of staining were seen for TGF-β and positive cells were confined to vessels. In contrast, in the damaged tendon, there was a dramatic increase in TGF-β, which was localized to the site of damage (Fig. 2A). This result was also confirmed by Western blotting, which demonstrated increased amounts of TGF-β 30, 60, and 90d after injury (Fig. 2B). This study was limited by the lack of available isoform-specific TGF-β antibodies, which would work on equine tissue. The TGF-β antibody used in these experiments cross reacts with all three isoforms of equine TGF-β and so we are unable to determine which isoforms are being upregulated.
Equine ESCs and adult tenocytes in vitro express receptors for TGF-β
Undifferentiated ESCs and cultured adult tenocytes express both TGF-β receptor I and receptor II suggesting that they could respond to TGF-β signaling. ESCs that undergo spontaneous differentiation for 7 days also retain the expression of both receptors. Although this is a mixed population of cells it suggests that ESC progeny are also capable of responding to TGF-β signaling (Fig. 3).
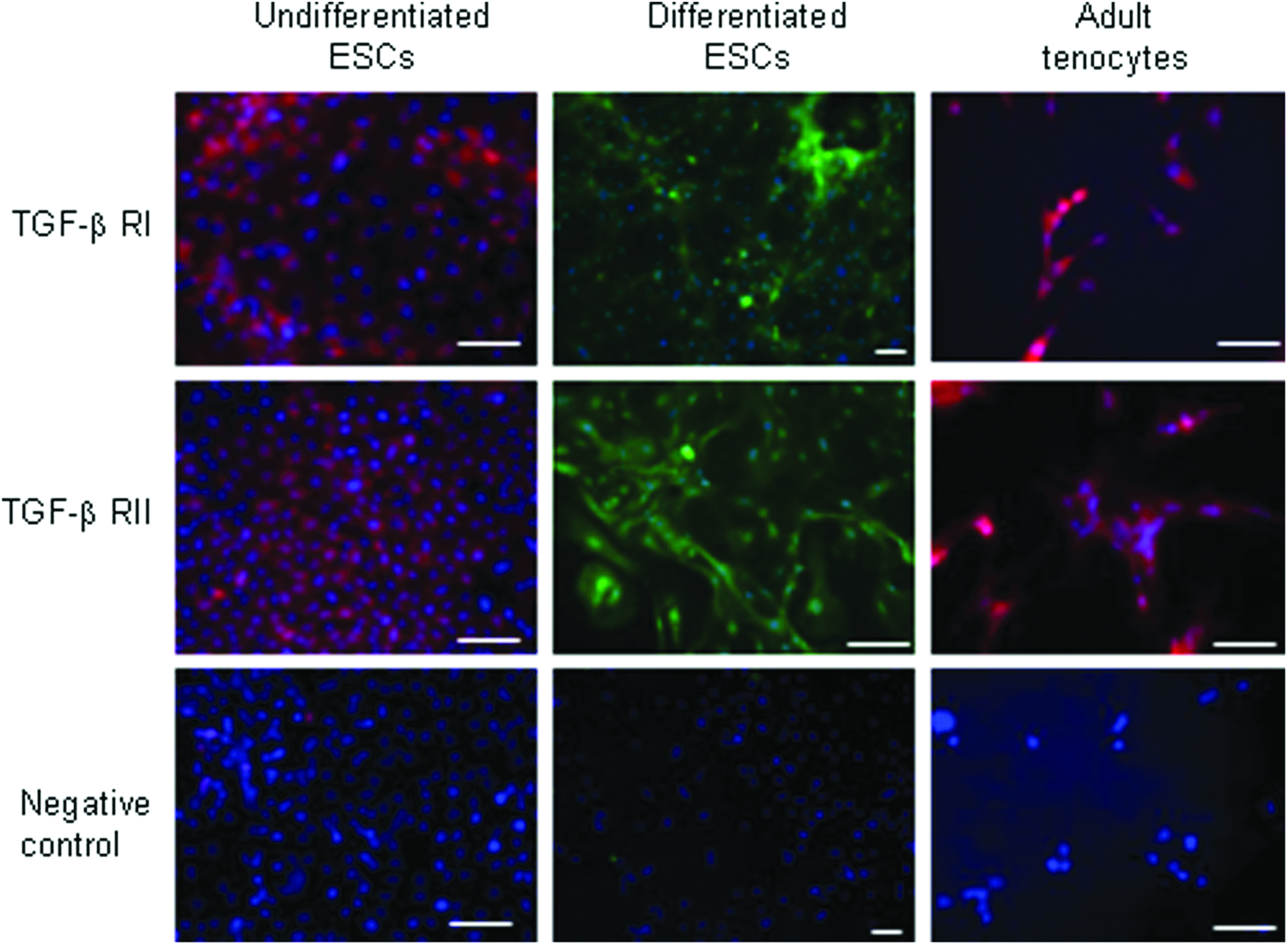
Immunocytochemistry demonstrates that undifferentiated ESCs, ESCs which have undergone spontaneous differentiation for 7 days, and adult tenocytes express TGF-β receptors I and II. DAPI staining of the nuclei is shown in blue. Scale bar=40 μm. Data shown are representative of testing on three cell lines. Color images available online at www.liebertpub.com/tea
In vitro exposure of equine ESCs to TGF-β upregulates scleraxis expression
Undifferentiated ESCs do not express scleraxis. When the ESCs were allowed to spontaneously differentiate for 72 h through the removal of LIF and feeders, scleraxis was expressed in a small proportion of the cells. After 72 h of differentiation in the presence of TGF-β, there was an upregulation in the intensity of scleraxis staining and the number of cells that are positively stained. All three isoforms of TGF-β appeared to produce this upregulation, but the greatest effect was observed with TGF-β3 (Fig. 4). This isoform was therefore selected for further investigation.

Immunocytochemistry for scleraxis shows that no protein is detected in undifferentiated ESCs. However, following 72 h of spontaneous differentiation, low-intensity staining for scleraxis is detected. The number and intensity of positively stained cells is increased in the presence of TGF-β with the largest effect produced by TGF-β3. DAPI staining of the nuclei is shown in blue. Scale bar=40 μm. Data shown are representative of testing on three cell lines. Color images available online at www.liebertpub.com/tea
ESCs differentiating in the presence of TGF-β3 upregulate the expression of other tendon-associated genes
Exposure of differentiating ESCs to TGF-β3 also resulted in an increase in the expression of the tendon-associated genes tenascin-C (TNC), collagen I (COL1A1), and cartilage oligomeric matrix protein (COMP) over different time points. Tenomodulin (TNMD) only showed a difference in its expression after 7 days. At this time, no TNMD expression was detected in ESCs differentiating in the absence of TGF-β, but TNMD expression was detected in ESCs differentiating in the presence of TGF-β3 (i.e., TNMD is induced at this time point by TGF-β3). However, expression of thrombospondin-4 (THBS4) was not detected at any time point (Fig. 5A). Of all the genes, SCX and TNC show the earliest detectable change following TGF-β3 exposure with a significant increase being detected after 24 h of differentiation between ESCs differentiating in the presence compared to the absence of TGF-β3. TNMD, COL1A1, and COMP tended not to show differences in expression in TGF-β3-treated versus -untreated ESCs until longer periods of differentiation were studied (3–7 days). Interestingly, by 14 days of differentiation, there was little difference in gene expression between treated and untreated ESCs.
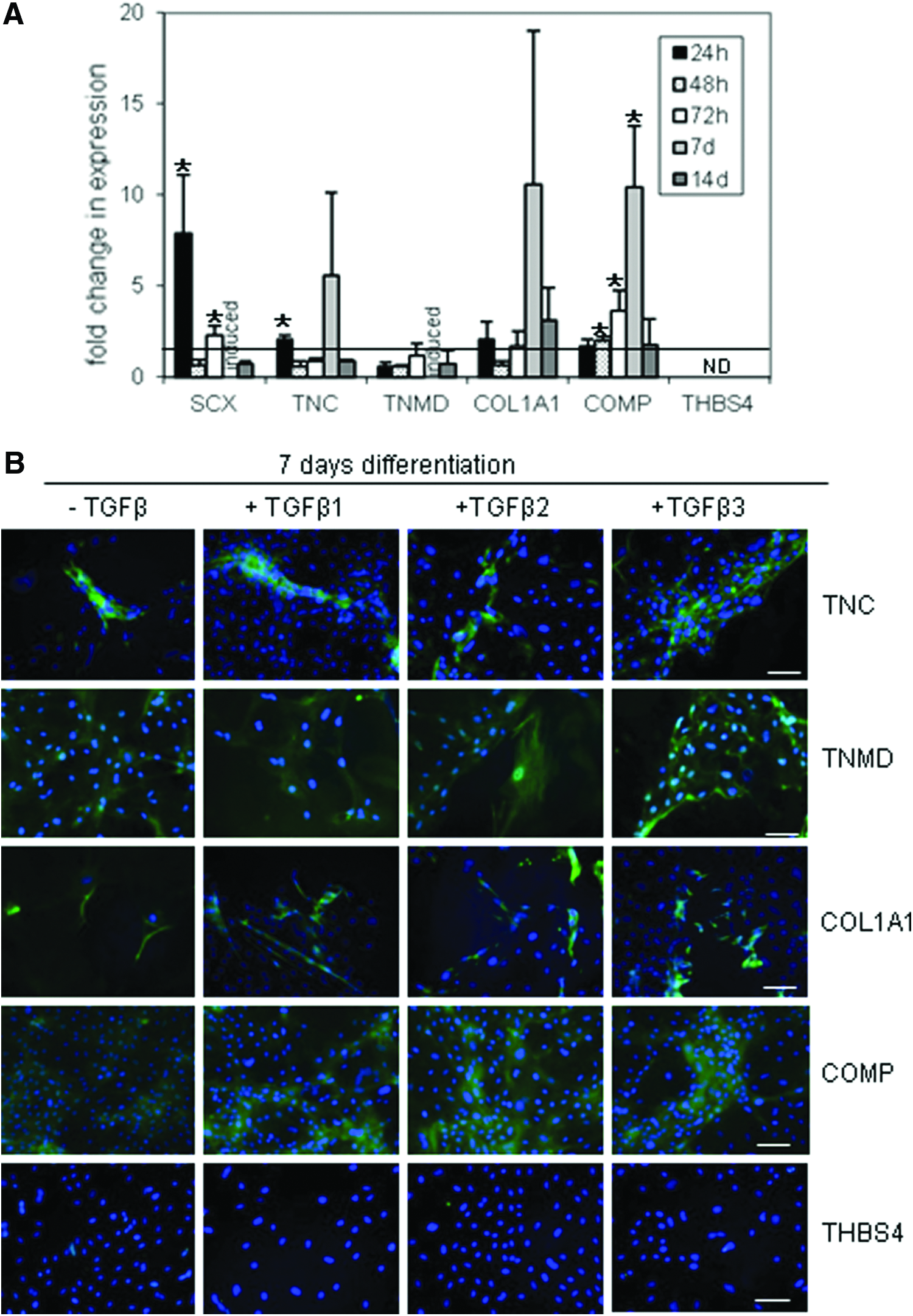
ESCs differentiating in the presence of TGF-β upregulate the expression of tendon-associated proteins
Immunocytochemistry for the proteins confirmed the results of the gene expression analysis with an increase in the intensity of staining and/or the number of positively stained cells observed for TNC, TNMD, COL1A1, and COMP following 7 days of exposure to TGF-β3 (Fig. 5B). Although TNMD gene expression is undetectable in cell cultures without TGF-β after 7 days of differentiation, the protein is detected at this time point. This suggests that we are detecting protein translated from mRNA, which is produced at the earlier time points and that the temporal expression of the proteins and RNA is different. However, the temporal expression of the proteins was not performed in this study. To determine if these effects are specific for TGF-β3, we also performed immunocytochemistry on ESCs allowed to differentiate in the presence of TGF-β1 and 2. All three isoforms of TGF-β produced similar effects after 7 days of differentiation suggesting that they are all able to promote tenocyte differentiation.
The COMP antibody used has previously been demonstrated to be specific for the equine protein. 17 To confirm the cross-species reactivity and specificity of the other antibodies, Western blots were performed on equine tendon tissue (Supplementary Fig. S2).
TGF-β treatment of adult tenocytes does not upregulate the expression of tendon-associated genes or proteins
Although adult equine tenocytes express receptors for TGF-β, exposure to TGF-β3 does not produce any statistically significant increases in expression of any of the tendon-associated genes studied at any time point (Fig. 6A). Correspondingly, no changes in protein expression are observed after 7 days of exposure to any isoform of TGF-β (Fig. 6B). Interestingly, TNMD and THBS4 proteins are detected in the absence of mRNA detection, suggesting that these proteins may have a low turnover and different rates of degradation to their corresponding mRNAs. 18

Discussion
The transcription factor scleraxis is required for the normal development of mouse tendons. 19 It is expressed in developing mouse tendons from embryonic day (E) 3.5 20 to E14.5.10,21 Recently, it has been demonstrated in a mouse model of patellar tendon injury that scleraxis mRNA levels are upregulated 4 weeks after injury with levels returning to normal after 12 weeks. 22 However, analysis of scleraxis mRNA levels in horse tendons with an acute (<6 weeks) injury showed no increase compared to healthy tendon controls. 23 In our study, we used immunohistochemistry and Western blot to demonstrate that the healthy horse tendon has very low levels of the scleraxis protein, but levels are increased after an injury. The increase is mostly localized around the lesion site and is sustained for at least 90 days in our tendon injury model. To our knowledge, this is the first report on scleraxis protein levels in response to injury and the data suggest that scleraxis has a role in tendon healing in horses.
We have previously shown that equine ESCs injected into the damaged tendon survived in high and constant numbers over the 90-day time period studied. 9 We therefore hypothesized that they were undergoing tenocyte differentiation. In this study, we have demonstrated that the majority of the ESCs express the scleraxis protein after 10, 30, 60, and 90 days following their injection, although there are large numbers of scleraxis-positive endogenous cells, which appear after an injury. The identity and source of the scleraxis-positive endogenous cells is currently unknown. However, as our results demonstrate that mature tendon cells do not upregulate scleraxis expression following TGF-β exposure, we hypothesize that the endogenous scleraxis-positive cells represent tendon progenitor cells, which are recruited to the site of injury, rather than local mature tendon cells which upregulate scleraxis themselves. Further work is required to test this hypothesis.
TGF-β2/3 signaling has been shown to be essential for tendon formation and can bring about the upregulation of scleraxis expression in mouse progenitor cells. 12 In this study, we demonstrated that TGF-β was increased in our injury model. A previous study in horses demonstrated that TGF-β1 was upregulated following an injury, but the authors did not assess the other isoforms. 13 However, all three isoforms of TGF-β are expressed in injured human and rat tendons14,15 and so it seems likely that all three isoforms are also upregulated in the injured horse tendon. Due to the lack of available antibodies, we are not able to confirm this. The precise roles that the different isoforms play during tendon healing and development are not clear. As TGF-β3 has been shown to be required for tendon formation and it produced the greatest effect on scleraxis expression by ESCs, we focused on its effects in this study. TGF-β binding requires the presence of TGF-β receptors RI and RII and we confirmed that in vitro ESCs and adult tenocytes express both TGF-β RI and RII suggesting they would be able to respond to TGF-β signaling.
Equine ESCs must be maintained in an undifferentiated state by the presence of feeder cells and LIF7,8 and in these conditions, the cells do not express detectable levels of the scleraxis protein or RNA (data not shown for RNA). Following spontaneous differentiation induced by the removal of LIF and feeder cells, scleraxis is induced in a small proportion of the cells. However, when cultured in the presence of TGF-β, there is an increase in the intensity of scleraxis staining and in the number of positively stained cells. This is reflected at the mRNA level, where 24 h of TGF-β exposure produces an eightfold increase in SCX expression compared to cells that are spontaneously differentiating in the absence of TGF-β.
Scleraxis overexpression has been shown to promote the differentiation of MSCs to tendon progenitors. 11 Scleraxis is a transcription factor and has previously been demonstrated to upregulate the expression of TNMD 24 and COL1A1. 25 It is not yet known if these genes are direct targets of scleraxis, but scleraxis can directly bind to the COL1A1 promoter. 26 In our study, after 7 days of differentiation, COL1A1 showed a large increase in its expression (although not statistically significant) and TNMD gene expression was induced (i.e., expression was only detected in the TGF-β3-treated cultures and not the control cultures at this time point). We also demonstrated that TGF-β3 promotes the relative upregulation of TNC and COMP genes in differentiating ESCs. These effects occur after as little as 24–48 h of TGF-β3 treatment, although larger increases are seen after 7 days for both genes. This suggests that scleraxis may be one of the earliest genes to respond to TGF-β3 signaling and it then drives the expression of the other tendon genes. However, future experiments to modulate scleraxis levels in the presence or absence of TGF-β are required to determine if TGF-β is promoting tenocyte differentiation directly through scleraxis. After 14 days of differentiation, there was little difference in gene expression between TGF-β3-treated and -nontreated ESCs. This suggests that while TGF-β3 enhances the rate of tendon differentiation by ESCs, some differentiation to this lineage occurs when the cells are allowed to spontaneously differentiate.
There is some variability in our gene expression results that was present between both biological (i.e., different lines of ESCs) and technical repeats (i.e., the same line of ESCs used in different experimental setups.) This seemed to be predominantly due to the variation in expression in the control cultures of ESCs, which were allowed to differentiate spontaneously in the absence of TGF-β. Our data demonstrate that some differentiation to the tenocyte lineage occurs when ESCs are allowed to spontaneously differentiate but, as previously reported, 7 a mixed cell population in observed with ectodermal and endodermal cell types also being present. The ESC lines are very similar in their basic stem cell properties such as marker gene expression and differentiation capacity,7,8 however, there may exist more subtle differences between lines as has been shown for equine mesenchymal stem cells isolated from different tissues and different individual horses. 27
Interestingly, we observed no induction of thrombospondin-4 in response to TGF-β treatment of the ESCs. Along with tenomodulin, thrombospondin-4 is thought to be the most specific marker of mature tendon 28 and it may be that longer time periods are required to see its induction. However, we do not believe that the lack of induction reflects any limitation with the differentiation potential of the ESCs. Although THBS4 is expressed in adult tendons, its gene expression is lost in cultured tenocytes ( 28 and Fig. 6A). Therefore, it seems likely that we will not detect an induction of THBS4 mRNA expression in two-dimensional (2D) differentiation assays using ESCs. Three-dimensional assays are currently being performed to provide an improved model of the in vivo situation, where cells are also being exposed to mechanical forces, which may well impact on TGF-β signaling 29 and scleraxis expression. 30
Equine tenocytes cultured from healthy equine tendon tissue do express TGF-β receptors, but do not significantly upregulate the expression of any of the tendon-associated genes studied following treatment with TGF-β3. In addition, no changes in protein expression are observed after 7 days of exposure to TGF-β1, 2, or 3. However, it is possible that TGF-β1 and/or 2 can result in changes in cultured adult tendon cell gene expression and that TGF-β may affect protein levels at other time points, which were not included in our study. The lack of detectable responses to TGF-β by adult tenocytes does not appear to be due to a higher baseline of gene expression as the expression levels of all genes except COMP and COL1A1 are comparable between the cultured tenocytes and differentiating ESCs. The differentiating ESCs and tenocytes are cultured in different basal media and we cannot rule out the possibility that this might have affected the results. Likewise, the protein data were performed on cells growing on gelatin-coated coverslips to allow adherence, whereas the cells used in the gene expression data were cultured on regular tissue culture plastic. In contrast to our results, previous reports have shown that TGF-β exposure can enhance COL1A1 expression in cultured tenocytes.14,31,32 However, increasing endogenous TGF-β in cultured tenocytes does not result in increased collagen production. 33 The reasons for these discrepancies are not clear and highlight the need for better understanding of the role of TGF-β in the adult tendon.
In this study, we have demonstrated that TGF-β signaling may play a role in promoting stem cell differentiation to tenocytes. Although the expression of scleraxis in ESCs injected into the injured horse tendon does not prove a full commitment to a tenocyte fate, 12 our in vitro studies reveal that equine ESCs can upregulate the expression of multiple tendon-associated genes in response to TGF-β signaling suggesting that they can undergo tenocyte differentiation. Further work is currently being performed to determine if these differentiated cells are functional tenocytes.
We have shown that, in contrast to ESCs, adult tendon cells do not respond to TGF-β signaling in 2D culture. We propose that the endogenous scleraxis-positive cells, which appear after an injury, may therefore represent tendon progenitor cells, which are recruited to the injury site. As the adult tendon undergoes poor regeneration, the functionality or numbers of these endogenous tendon cells may be limiting regeneration. By supplementing the damaged tendon with a population of ESCs that are able to differentiate into tenocytes, we may be able to enhance adult tendon regeneration. Our results therefore support further in vivo studies into the efficacy of equine ESCs on tendon regeneration.
Together, this work demonstrates that equine ESCs are capable of differentiating into a therapeutically relevant cell type for the treatment of horse tendon injuries and also that they may provide a model to study tendon development and better understand the transcriptional networks that are involved in equine tendon cell differentiation from the early embryonic stages.
Footnotes
Acknowledgments
The authors would like to thank Professor Roger Smith (Royal Veterinary College) for kindly providing the COMP antibody, Ms Emma Goodfellow (Animal Health Trust) for her assistance with collecting tendon tissue samples, and the Paul Mellon Foundation for funding.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.