
Abstract
Macrophages are phagocytic cells with great importance in guiding multiple stages of inflammation and tissue repair. By producing a large number of biologically active molecules, they can affect the behavior of other cells and events, such as the foreign body response and angiogenesis. Since protein adsorption to biomaterials is crucial for the inflammatory process, we addressed the ability of the pro-inflammatory molecule fibrinogen (Fg) to modulate macrophage behavior toward tissue repair/regeneration. For this purpose, we used chitosan (Ch) as a substrate for Fg adsorption. Freshly isolated human monocytes were seeded on Ch substrates alone or previously adsorbed with Fg, and allowed to differentiate into macrophages for 10 days. Cell adhesion and morphology, formation of foreign body giant cells (FBGC), and secretion of a total of 80 cytokines and growth factors were evaluated. Both substrates showed similar numbers of adherent macrophages along differentiation as compared with RGD-coated surfaces, which were used as positive controls. Fg did not potentiate FBGC formation. In addition, actin cytoskeleton staining revealed the presence of punctuate F-actin with more elongated and interconnecting cells on Ch substrates. Antibody array screening and quantification of inflammation- and wound-healing-related factors indicated an overall reduction in Ch-based substrates versus RGD-coated surfaces. At late times, most inflammatory agents were down-regulated in the presence of Fg, in contrast to growth factor production, which was stimulated by Fg. Importantly, on Ch+Fg substrates, fully differentiated macrophages produced significant amounts of macrophage inflammatory protein-1delta (MIP-1δ), platelet-derived growth factor-BB, bone morphogenetic protein (BMP)-5, and BMP-7 compared with Ch alone. In addition, other important factors involved in bone homeostasis and wound healing, such as growth hormone, transforming growth factor-β3, and insulin-like growth factor-binding proteins, as well as several angiogenic mediators, including endocrine gland-derived vascular endothelial factor, fibroblast growth factor-7, and placental growth factor, were significantly promoted by Fg. This work provides a new perspective on the inflammatory response in the context of bone repair/regeneration mediated by a pro-inflammatory protein (Fg) adsorbed onto a biomaterial (Ch) that does not otherwise exhibit osteogenic properties.
Introduction
M
Chitosan (Ch) is a nontoxic and biodegradable polysaccharide that is obtained by N-deacetylation of chitin, with anti-tumor, anti-fungal, and anti-microbial properties.10,11 It has been used in several biomedical applications, such as wound dressings, drug and gene delivery, and bone tissue repair.11–14 In general, Ch induces a minimal foreign body reaction with little or no fibrous encapsulation. 12 The composition of Ch, similar to extracellular matrix glycosaminoglycans, its gel-forming properties, easy chemical modification, and affinity to proteins, renders this polymer an interesting candidate for tissue-engineering applications.13–16 Previous studies suggest that Ch could accelerate wound healing by enhancing the functions of inflammatory and repairing cells, including macrophages.17–19 Other studies showed that the exposure of THP-1 human macrophage cell line to Ch-DNA nanoparticles did not induce the release of pro-inflammatory cytokines, 20 and water-soluble Ch was described as having an anti-inflammatory effect, being a strong immunomodulator of the alternative activation of macrophages to allergen stimulation. 21 In addition, we have recently documented that despite eliciting an early up-regulation of pro-inflammatory cytokines by macrophages, Ch is able to induce an anti-inflammatory time-dependent macrophage polarization. 22 Since the modulation of macrophage phenotype from pro- to anti-inflammatory is of key importance for tissue engineering, here we explored the potential of Ch in combination with the pro-inflammatory agent fibrinogen (Fg), to influence macrophage behavior, which is of relevance in tissue repair/regeneration.
Fg is a plasma glycoprotein that is involved in the maintenance of homeostasis and in several immune functions. Fg interacts with beta2 integrin receptors, such as αXβ2 (CD11b/CD18, Mac-1) and αXβ2 (CD11c/CD18), on monocytes/macrophages.23,24 Interactions through these adhesion molecules have been reported to induce monocyte/macrophage activation, regulating important activities such as phagocytosis, cell migration, apoptosis and cell death, and cytokine production.24,25 Preadsorption of Fg has been shown to have an important role in initiating inflammatory responses to implanted biomaterials and, in particular, in the regulation of the macrophage response.26–30 Fg is also crucial in wound healing and repair. Fg(−/−) mice form granulation tissue morphologically similar to control animals, but less mechanically stable, with a consequent wound instability. 31 In addition, Fg has been described as playing a key role in tissue repair at implant surfaces, 2 promoting new bone formation when conjugated with poly(ethylene glycol) and used as an hydrogel. 32 Of note, we have recently demonstrated that when adsorbed to Ch scaffolds, Fg leads to more bone formation in vivo, stimulates angiogenesis, and correlates with the systemic immune response. 33 Given the relevance of these findings and the impact of macrophages in inflammation and bone regeneration, 34 we broaden our research by exploring the potential of Fg to modulate macrophage behavior toward a pro-regenerative phenotype. This investigation contributes to further understanding macrophage-biomaterial interactions and, more importantly, to advance knowledge on the underestimated contribution of the inflammatory response to bone regeneration.
Materials and Methods
Ch purification and film preparation
Ch was purified as previously described, 35 and films were prepared as reported elsewhere. 22
Adsorption of human Fg and RGD peptide
Human Fg (Sigma-Aldrich) was reconstituted in Dulbecco's phosphate-buffered saline containing Ca2+ and Mg2+ (PBS++; Gibco), filtered, and stored at −80°C until further use. Where appropriate, Ch films were incubated for 2 h at room temperature (RT) with Fg solution at 100 μg/mL. Unadsorbed protein was washed out by rinsing the films twice with 0.5 mL PBS++. The amount of adsorbed Fg was quantified by protein radiolabeling with 125I and found to be 501±63 ng of protein/cm2. 36 The RGD peptide (Fibronectin-like Protein Polymer, F5022 from Sigma-Aldrich) was reconstituted according to the manufacturer's instructions and used at 25 μg/mL to adsorb (20 min at RT) glass coverslips on 24-well culture plates (Fisher Scientific). Before cell culture, wells were washed twice. RGD was used as a positive control surface for macrophage development and adhesion and FBGC formation, as previously reported.37–39 Adsorbed RGD is widely recognized for its adhesion-promoting capabilities in that it presents multiple copies of the RGD (arginine-glycine-aspartate) cell attachment sequence to integrin receptors, thus facilitating their engagement.
Monocyte isolation
Human peripheral monocytes and serum were isolated from the whole venous blood of healthy, unmedicated donors as described by McNally and Anderson. 40 Briefly, citrated blood (100–150 mL) was diluted 1:1 with PBS/5 mM EDTA (PBSE), layered onto Ficoll-Paque (GE Healthcare), and centrifuged (1700 rpm, 30 min, without brake). The resultant mononuclear cells were serially washed (two centrifugations at 1700 rpm and one at 1300 rpm, 10 min each) with PBSE and resuspended in 2–2.5 mL of PBSE. This cell suspension was layered onto two 10 mL columns of fetal bovine serum, and centrifuged (900 rpm, 9 min). Cells were then resuspended in PBSE, mixed with modified Percoll (7.29 mL Percoll [GE Healthcare]+1.8 mL dH2O+0.91 mL 1.5 M NaCl), and centrifuged (3300 rpm, 25 min, without brake). The resultant top cell layer (1 mL) was finally washed twice with cold Macrophage Serum-Free Medium (Macrophage-SFM; Gibco), and resuspended in cold Macrophage SFM for counting. Monocytes were maintained on ice until use to minimize aggregation.
Cell culture
After monocyte isolation, 5.0×105 cells were seeded per well in 24-well culture plates with 0.5 mL of Macrophage SFM supplemented with 20% of prefiltered autologous serum and allowed to adhere for 1.5 h. Nonadherent cells were removed by washing with warmed PBS++, and adherent monocytes were further cultured in fresh Macrophage SFM at 37°C, 5% CO2. At days 3, 7, and 10, cultures were either fixed or maintained in culture with 0.5 mL of Macrophage SFM supplemented with 5% heat-treated (1 h at 56°C), autologous serum. In parallel, the fusion-inducing cytokine interleukin (IL)-4 41 (15 ng/mL) (R&D Systems, reconstituted in 0.5% bovine serum albumin [low endotoxin; Sigma-Aldrich]), was added to the fresh media of selected cultures at days 3 and 7 to promote fusion. After 3, 7, and 10 days, supernatants were collected, nonadherent cells were removed by washing, and adherent cells were fixed with methanol or 3.7% v/v formaldehyde. Collected supernatants were combined, centrifuged at 2000 rpm for 10 min to remove nonadherent cells, and stored at −80°C.
Adherent cell density analysis
Fixed cells were stained with May-Grünwald for 5 min, rinsed twice with PBS++, incubated in Giemsa for 15 min, washed thrice with distilled water, and allowed to air-dry overnight. Macrophage adhesion and FBGC formation were simultaneously evaluated by counting the total number of adherent nuclei in representative microscopic fields per unit area in an inverted light microscope (Olympus). The percentage of nuclei within multinucleated cells (≥3 nuclei) was used to calculate the percentage of multinucleated cells.
Cell morphology analysis
Firstly, formaldehyde-fixed cells were treated for 1.5 h at 37°C with 100 μg/mL RNase A (EMD Biosciences, Inc.) to degrade cytoplasmic nucleic acids. Samples were then washed thrice with PBS++ for 5 min, and incubated (1 h, 37°C, in the dark) with rhodamine phalloidin (1/100; Molecular Probes), for F-actin staining, and YOYO-1 (1/10,000; Molecular Probes) to counterstain the nuclei. Finally, samples were washed thrice for 10 min, mounted with Gel/Mount, Biomeda, and kept at −20°C protected from the light until analysis by inverted fluorescence microscopy (Olympus IX71) using Olympus MicroSuite™ software.
Cytokine and growth factor screening and quantification
Macrophage-conditioned media were collected at days 3, 7, and 10, and pools from three to five donors were analyzed for each time point using an array system (Quantibody Human Cytokine Antibody Array 1000 kits; RayBiotech, Inc.) in accordance with the manufacturer's instructions. Slides were run on a laser scanner (GenePix 4000B Microarray Scanner) with the following settings: Excitation: 555 nm; Emission: 565 nm; Resolution: 10 μm.
Statistical analysis
Statistical analyses were performed using R version 2.15.2. 42 ANOVA analysis and Tukey's HSD tests with a 95% confidence interval were used to determine statistical significance between groups. When appropriated, data were analyzed using the nonparametric Kruskal–Wallis test followed by multiple-comparisons post-hoc test according to Siegel and Castellan. 43 The significant variation of soluble factors production was performed by a trend analysis. A p-value of <0.05 (two sided) was considered statistically significant. Data are presented as mean±standard deviation. Donor variability was accounted for using multiple human donors (n=5).
Results
Macrophage adhesion and FBGC formation
The effect of Fg adsorbed to Ch films on macrophage adhesion and fusion was evaluated. As shown in Figure 1, high cell density was observed on Ch-based substrates at day 3 of culture, with levels similar to those of the RGD positive control. At later time points, cell adhesion was equally supported on all three surfaces, despite showing a tendency to decrease on both Ch-based matrices. Moreover, the presence of Fg did not influence macrophage adhesion (Fig. 1). To explore the ability of Ch to further support macrophage adhesion and FBGC formation, the fusion promoting cytokine IL-4 was added to monocytes/macrophages at days 3 and 7, and cultures were analyzed at days 7 and 10. As expected, IL-4 induced a marked decrease in cell density on all surfaces, but at different stages of macrophage differentiation: while on RGD-coated surfaces, significant differences were found from day 3 to 7 (p<0.05), on Ch-based substrates, statistical significance was only observed later, from day 3 to 10 (p<0.05). No changes in macrophage adhesion were detected though from day 7 to 10 in any of the materials tested (Fig. 1).
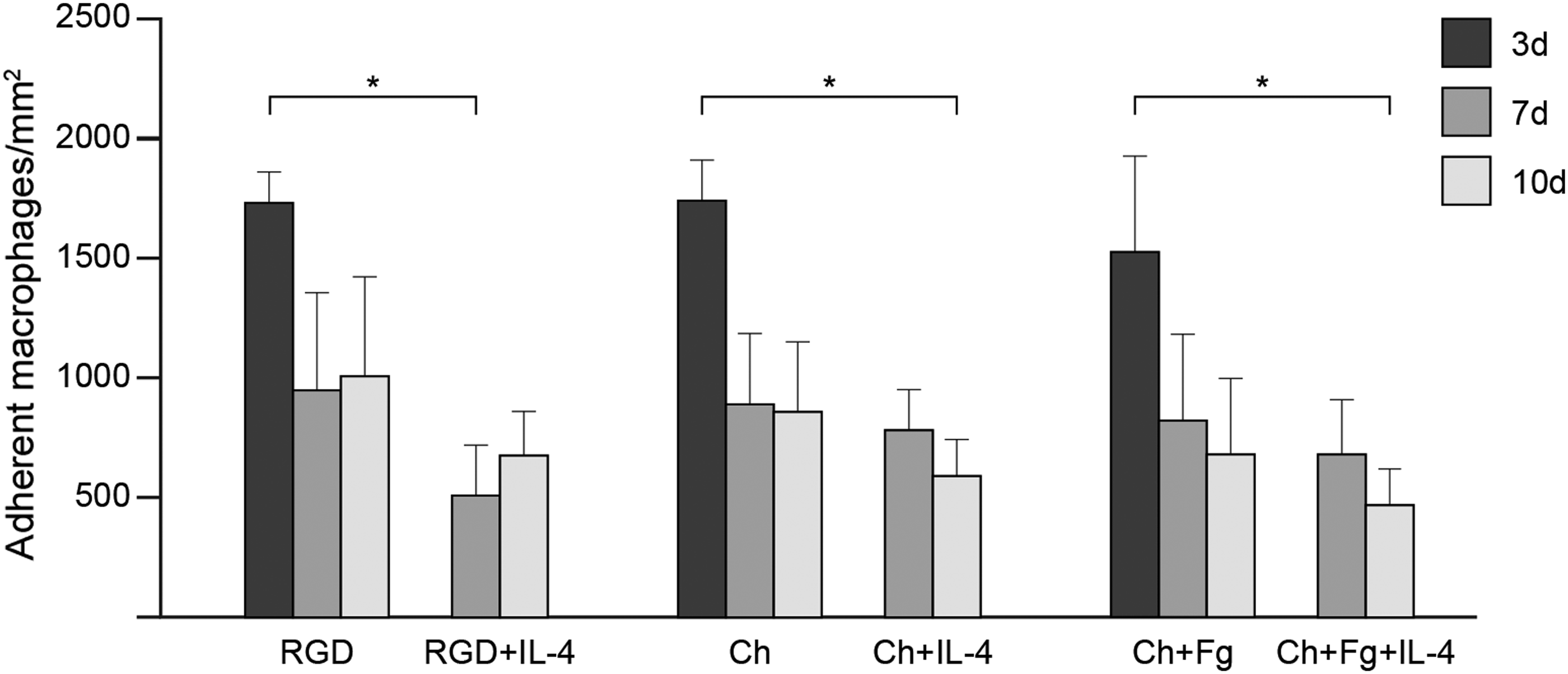
In vitro human monocyte/macrophage adhesion to chitosan (Ch) films. Human monocytes were cultured on Ch films and Ch films with adsorbed human fibrinogen (Fg). Interleukin (IL)-4-induction of macrophage fusion was performed at days 3 and 7. RGD-modified glass was used as a control. Cultures were fixed and stained with May–Grünwald/Giemsa at days 3, 7, and 10 and cells were counted. Results represent the mean of adherent monocytes/macrophages per area (mm2)±standard deviation, n=3–5 different monocyte donors. Asterisks indicate statistically significant difference (*p<0.05) at each respective time point.
The formation of FBGC on Ch films and the influence of Fg on this process were investigated next. For this purpose, percent fusion, that is, percentage of nuclei within multinucleated cells (cells with three or more nuclei), was determined at different time points (Fig. 2). On unmodified Ch films, macrophage fusion increased significantly (p<0.05) from day 3 to 10, similar to RGD control surfaces. In contrast, Fg coating had no effect on macrophage fusion throughout the culture period. Addition of IL-4 significantly potentiated the formation of FBGC from day 3 to 7 on Ch-based surfaces (p<0.05; Fig. 2). However, from day 7 to 10, no changes in macrophage fusion were noted in Ch and Ch+Fg with or without IL-4, as opposed to RGD where a trend for greater FBGC formation was observed (Fig. 2).
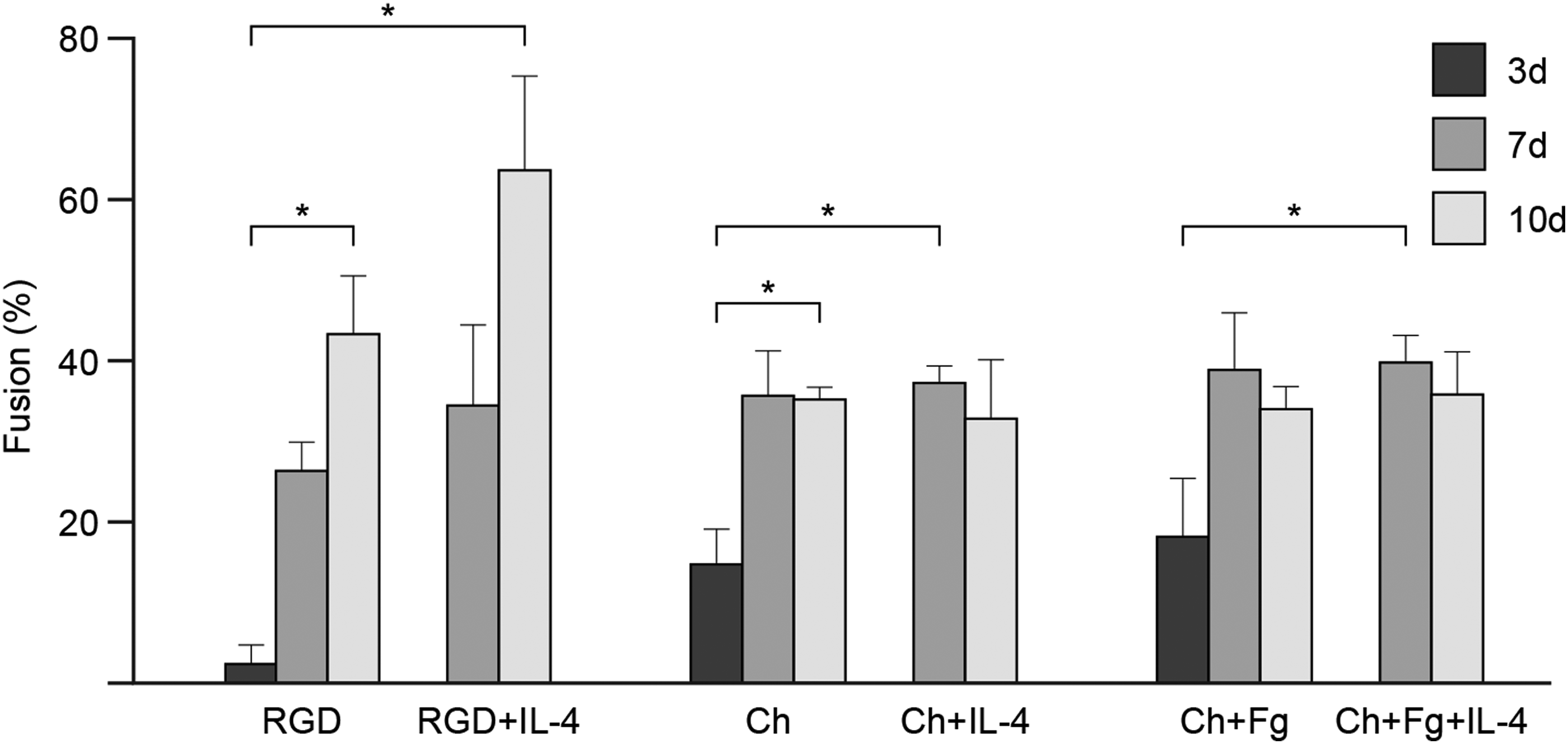
FBGC formation: % fusion of macrophages on Ch films. Human monocytes were cultured on Ch films and Ch films with adsorbed human Fg. IL-4 induction of macrophage fusion was performed at days 3 and 7. RGD-modified glass was used as a control. Cultures were fixed and stained with May–Grünwald/Giemsa at days 3, 7, and 10, and percent macrophage fusion was determined by counting the nuclei within multinucleated cells (cells with three or more nuclei). Results represent mean% fusion±standard deviation, n=3–5 different monocyte donors. Asterisks indicate statistically significant differences (*p<0.05) at each respective time point.
Morphological features of macrophages and FBGC
In addition to evaluating macrophage adhesion and fusion, the influence of substrate composition on macrophage morphological development was also investigated. Figure 3 depicts representative images of monocytes/macrophages cultured on the different surfaces after 3, 7, and 10 days. During monocyte differentiation into macrophages, adherent cells became larger in size. By day 7, many multinucleated cells could be seen on all substrates (Fig. 3A-b, e, h). F-actin appeared diffused around the nuclei and delineated cell boundaries on all substrates. Cells seeded on Ch (Fig. 3A-e, f) and Ch+Fg films (Fig. 3A-h, i) showed an irregular shape, frequently forming inter-cellular connections through long cytoplasmic filaments. Furthermore, filopodia were visible on Ch (Fig. 3A-e, f) and Ch+Fg films (Fig. 3A-h, i). No differences were observed on Ch+Fg films as compared with Ch alone.
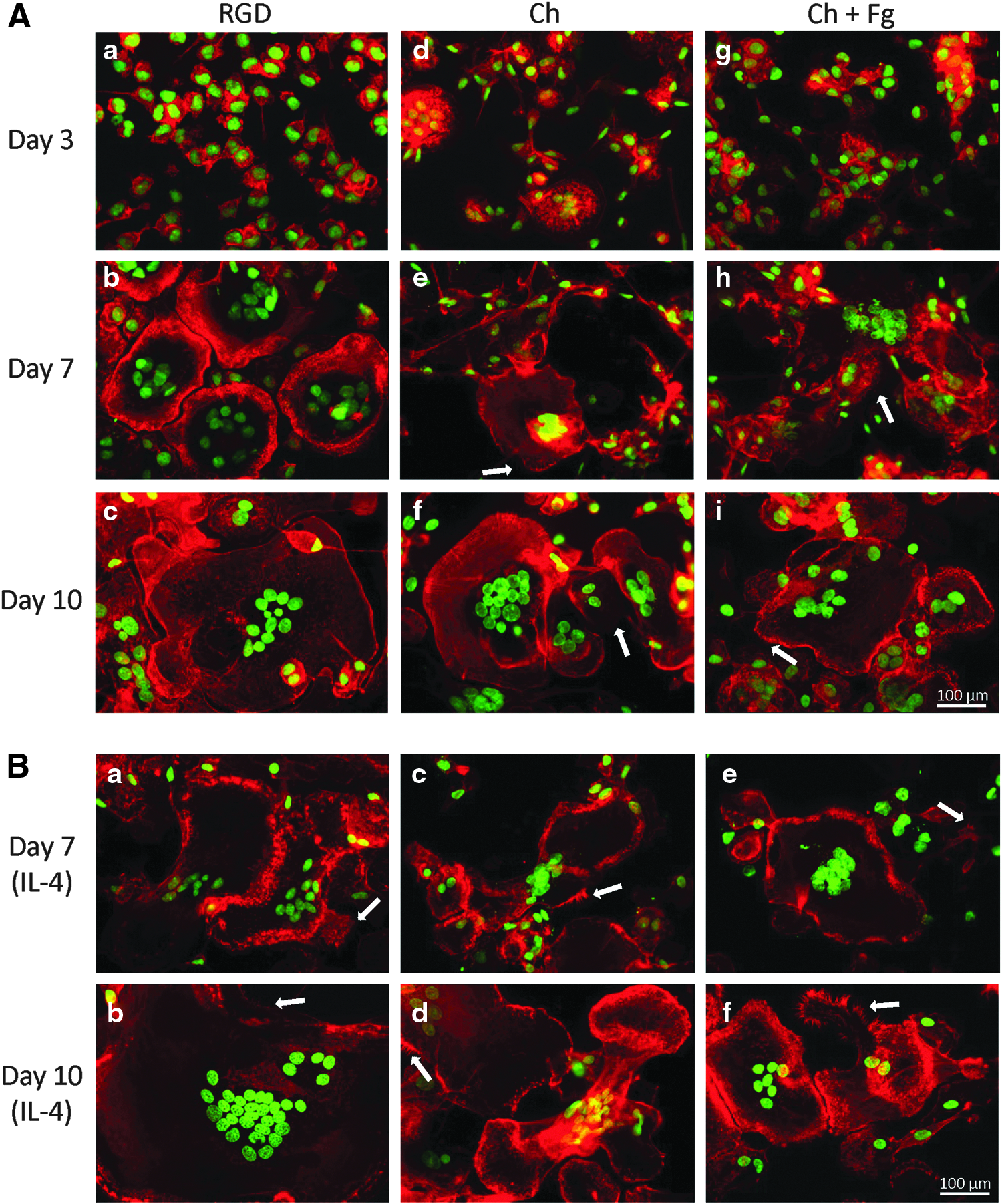
Monocyte/macrophage morphology on Ch films.
With IL-4, more elongated FBGC were formed, with punctuate F-actin and the filopodia visible on all substrates (Fig. 3B). Nevertheless, in the presence of IL-4, RGD-coated surfaces (Fig. 3B-a, b) induced formation of larger cells than Ch films with (Fig. 3B-e, f) or without Fg (Fig. 3B-c, d). Again, no differences were seen between Ch and Ch+Fg.
Cytokine and growth factor secretion profile
Supernatants from macrophage cultures were collected at days 3, 7, and 10, and screened using quantitative antibody arrays for the presence of 40 inflammation-related soluble mediators and 40 growth factors. Data were normalized based on the adherent cell population, and concentrations were determined as the amount of cytokine/growth factor produced per cell. To better illustrate the effect of substrate on macrophage cytokine/growth factor profiles, results were plotted as color gradient tables, where each shade represents a range of concentrations and factors are organized into functional categories; for example, pro- and anti-inflammatory cytokines, chemokines, and so on. (Figs. 4 and 5). Individual concentrations measured over time are presented in Supplementary Figure S1 (Cytokines, Chemokines) and S2 (Growth factors). Statistical analysis is shown in Supplemmentary Tables S1 and S2 (Supplementary Data are available online at www.liebertpub.com/tea). Macrophage differentiation on RGD surfaces resulted in an overall greater production of soluble factors than on Ch-based matrices (Fig. 4). Still, despite the reduced activation of adherent macrophages on Ch-based films, macrophage inflammatory protein-1 alpha (MIP-1α) and tissue inhibitor of metalloproteinase (TIMP) 1 and 2 displayed high responses at all three time points (Fig. 4). In addition, elevated levels of inter-cellular adhesion molecule-1 (ICAM-1) were already observed at day 3 in the presence of Fg, which continued to increase until day 10. Furthermore, moderate amounts of tumor necrosis factor (TNF) receptor I and II were detected on Ch and Ch+Fg. On the other hand, lower levels of pro-inflammatory cytokines were produced by Fg-stimulated macrophages versus those cultured on unmodified-Ch films. This is particularly evident for IL-6 and IL-15 and chemokine RANTES, which were greatly up-regulated on Ch at day 10 but not on Ch+Fg films (Fig. 4). When comparing macrophage secretory profiles on the three substrates, differences were statistically significant for all inflammation-related factors between RGD and Ch-based films, and for many proteins between Ch and Ch+Fg, namely IL-1β, IL-6, TNF-α, MIP-1β, MIP-1δ, RANTES, and TIMPs 1 and 2.

Color gradient representation of ranges of cytokine levels released by macrophages cultured on Ch films. Macrophages were cultured on Ch films or Ch films with adsorbed Fg for 10 days, and supernatants were collected at days 3, 7, and 10. Pools of culture supernatants from three to five donors were analyzed for each time point. RGD-modified glass was used as a positive control. Supernatants were analyzed using protein antibody arrays. Each color represents a range of concentrations, and functional categories are indicated on the left. Statistical significance is indicated on the right, as follows: † indicates statistical significance (p<0.05) between RGD and Ch. # indicates statistical significance (p<0.05) between RGD and Ch+Fg. ¥ indicates statistical significance (p<0.05) between Ch and Ch+Fg. Color images available online at www.liebertpub.com/tea

Color gradient representation of ranges of growth factor levels released by macrophages cultured on Ch films. Macrophages were cultured on Ch films or Ch films with adsorbed Fg for 10 days, and supernatants were collected at days 3, 7, and 10. Pools of culture supernatants from three to five donors were analyzed for each time point. RGD-modified glass was used as a positive control. Supernatants were analyzed using protein antibody arrays. Each color represents a range of concentrations, and functional categories are indicated on the left. Statistical significance is indicated on the right, as follows: † indicates statistical significance (p<0.05) between RGD and Ch. # indicates statistical significance (p<0.05) between RGD and Ch+Fg. ¥ indicates statistical significance (p<0.05) between Ch and Ch+Fg. Color images available online at www.liebertpub.com/tea
Data regarding growth factor release are presented in Figure 5. As for inflammation-related factors, RGD surfaces potentiated significantly macrophage production of growth factors relative to Ch-based films. Nonetheless, high levels of bone morphogenetic proteins (BMP) 5 and 7, particularly on Ch+Fg films, are noted at times as early as day 3. The same applies, albeit to a lesser extent, macrophage chemotactic factor receptor (MCF R). Of note, Fg accelerated the release of factors that are important in bone and wound-healing processes (BMP-5, BMP-7, growth differentiation factor (GDF) 15, growth hormone (GH), and heparin-binding epidermal growth factor-like growth factor (HB-EGF)), which are increased at earlier time points on Ch+Fg in comparison to unmodified Ch films. However, statistical significance between Ch+Fg and Ch was only found for GDF-15.
To further understand how Ch films with or without Fg would affect macrophage activation, the ratio between the released factors produced after 3, 7, and 10 days of differentiation on Ch and Ch+Fg substrates was calculated. Results are depicted in Figures 6 (inflammation-related factors) and 7 (growth factors). At day 3, most pro-inflammatory molecules were down-regulated in the presence of Fg relative to Ch alone (Fig. 6, top panel). Regarding chemokine secretion at day 3, monokine induced by interferon-gamma (MIG) and MIP-1δ levels significantly increased on Ch+Fg films. In addition, greater production of ICAM-1, interleukin-6 soluble receptor (IL-6sR), platelet-derived growth factor-BB (PDGF-BB), TIMP-2, and TNF RI was noted with Fg. At later time points, that is, days 7 and 10, an overall decrease in the Ch+Fg/Ch ratio was observed (Fig. 6, middle and lower panels). Importantly, this was not due to a reduction in cytokine release with Fg, which increased until day 10, but rather to a higher production rate on unmodified Ch films from day 3 to 7 and 10 (Supplementary Fig. S1). Strikingly, however, at day 10, significantly higher levels of MIP-1δ (Fig. 6, lower panel) and a trend for greater ICAM-I and PDGF-BB production on Ch+Fg relative to uncoated Ch films were observed.
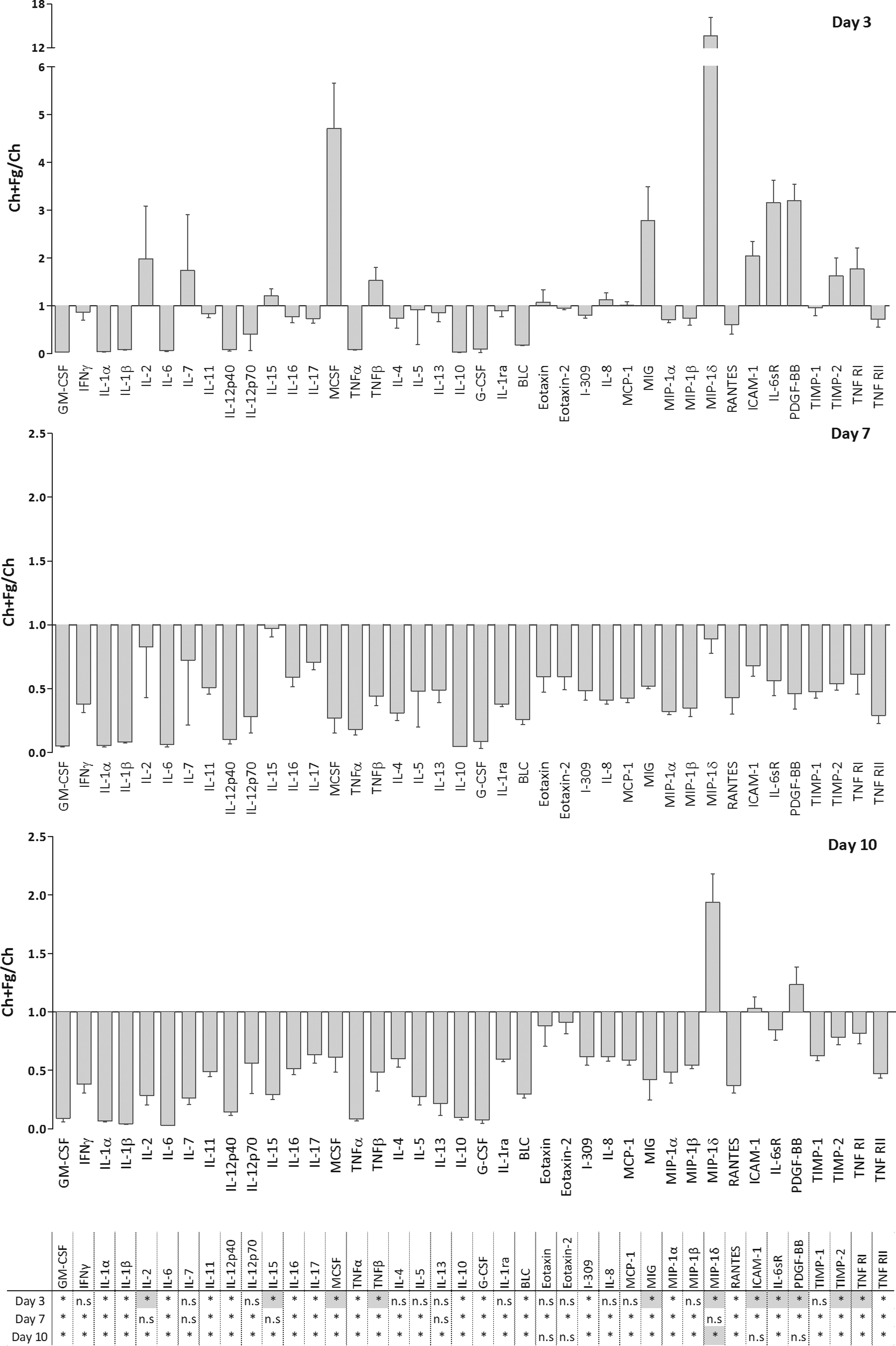
Ratio of macrophage-released cytokines after culture on Ch films with or without Fg over time. Macrophages were cultured on Ch films or Ch films with adsorbed Fg. Cells were cultured for 10 days, and supernatants were collected at days 3, 7, and 10. Supernatants were analyzed using antibody arrays. The ratio between macrophage-released cytokines on Ch+Fg and Ch was calculated for each time point as indicated. Asterisks indicate statistically significant difference (*p<0.05) between proteins up- (gray squares) or down-regulated (white squares) on Ch+Fg versus Ch. n.s, not significant.
As for growth factor secretion, a different scenario was detected. In this case, at day 3, Fg promoted production of many proteins, especially BMP-5, BMP-7, GDF-15, amphiregulin (AR), and insulin (Fig. 7 upper panel, Supplementary Fig. S2). Of note, elevated levels of both BMP-5 and BMP-7 were maintained at later time points on Fg-modified substrates (Fig. 7 middle and lower panels). Still, statistical significance for BMP-5, BMP-7, and insulin could not be detected at all time points due to the variability of protein levels increase observed for these proteins. In addition, other factors involved in angiogenesis, wound healing, and bone homeostasis, such as endocrine gland-derived vascular endothelial growth factor (EG-VEGF), GH, fibroblast growth factor (FGF)-7, and transforming growth factor (TGF)-β3, among others, were also significantly increased on Ch+Fg at days 7 and 10, although to a lower extent than BMPs (Fig. 7 middle and lower panels). Lastly, at day 10, Fg augmented the release of most members of the Insulin-like growth factor-binding proteins (IGFBP) family, which are important regulators of bone metabolism (Fig. 7, lower panel).
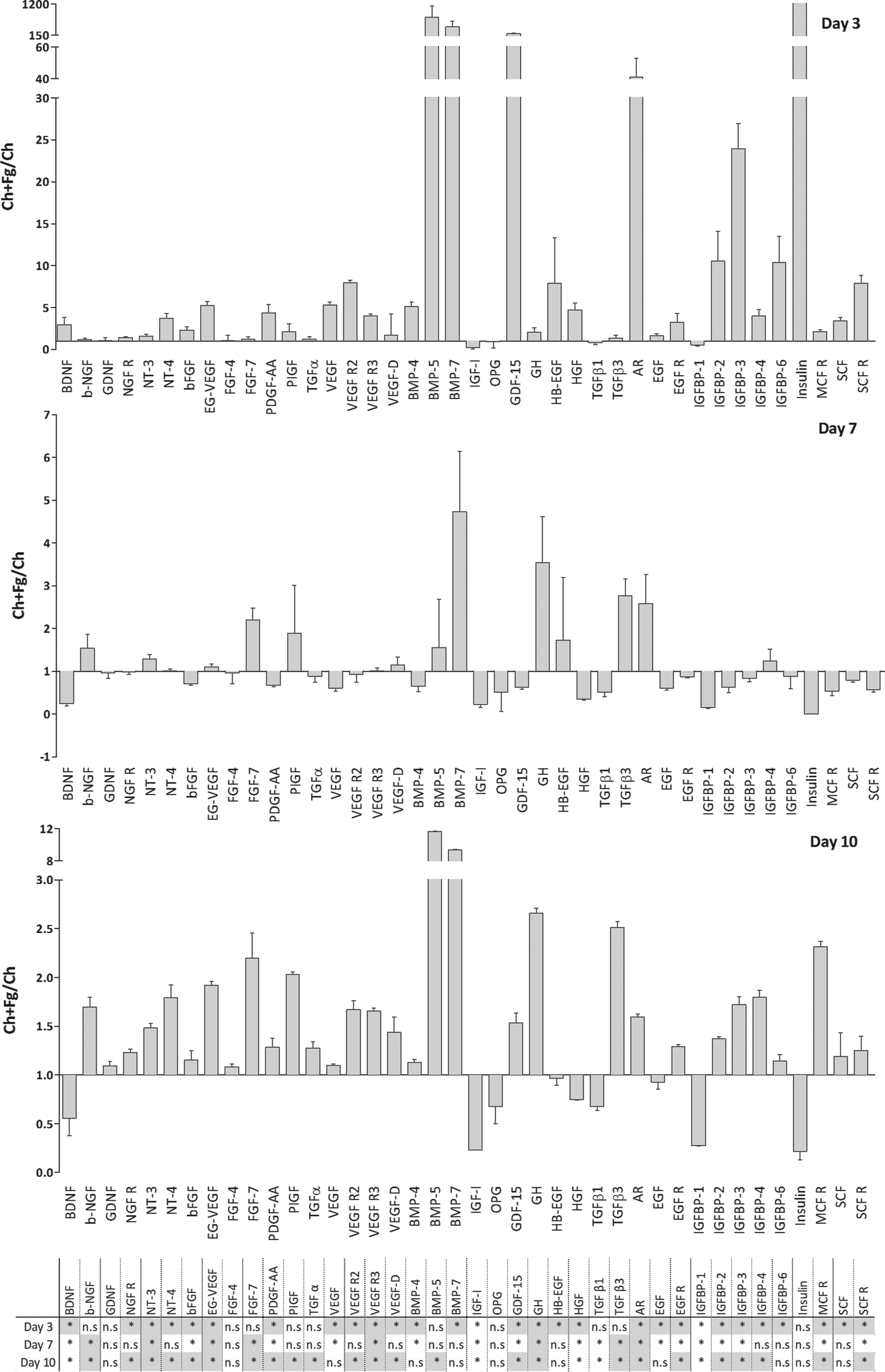
Ratio of macrophage-released growth factors after culture on Ch films with or without Fg over time. Macrophages were cultured on Ch films or Ch films with adsorbed Fg. Cells were cultured for 10 days, and supernatants were collected at days 3, 7, and 10. Supernatants were analyzed using antibody arrays. The ratio between macrophage-released cytokines on Ch+Fg and Ch was calculated for each time point as indicated. Asterisks indicate statistically significant differences (*p<0.05) between proteins up- (gray squares) and down-regulated (white squares) on Ch+Fg versus Ch.
Discussion
Macrophages are described as playing disparate functions in response to biomaterial implantation. 2 Here, we have investigated the influence of Ch, modified or not with a pro-inflammatory molecule, on monocyte/macrophage adhesion and development, FBGC formation, and cytokine/growth factor production. Particular interest has been given to secretion of soluble mediators related to bone biology, as contradictory roles have been suggested for macrophages and FBGC in bone regeneration. Some studies indicate that these cell types are responsible for chronic inflammation and failure of bone formation, 44 whereas others argue that macrophages participate in biomaterial biodegradation and produce osteoinductive and osteogenic cytokines in an osseous wound-healing environment.1,45 Considering that monocyte/macrophage responses are influenced by protein coatings on implant surfaces, 4 and given the relevance of Fg in the inflammatory reaction26,29 and bone formation, 32 we have adsorbed Fg on Ch films and evaluated the ability of this pro-inflammatory protein to modulate macrophage behavior. Importantly, we have recently reported improved bone formation in vivo using Fg-modified Ch scaffolds, which correlated with the systemic immune response. 33
Cell adhesion (Fig. 1) and FBGC formation (Fig. 2) showed similar patterns on modified or unmodified Ch substrates. In addition, cell morphology (Fig. 3) was identical on Ch and Ch+Fg surfaces, with cells adopting a more irregular and elongated shape than in control conditions. The absence of a Fg-mediated effect on these cellular responses is not completely unexpected, as preadsorption of Fg has been suggested to variably affect monocyte and macrophage adhesion and activation. 46 McNally et al. observed lower monocyte/macrophage adhesion and no IL-4-induced FBGC formation on standard cell culture polystyrene preadsorbed with Fg when compared with RGD-coated substrates at all time points studied (0, 3 and 7 days). 37 On the other hand, others have found enhanced macrophage adhesion on different surfaces preadsorbed with Fg, relative to corresponding uncoated surfaces, 2 h and 1 day after cell seeding. 47 In addition, Yun et al. showed that Fg preadsorbed onto poly(tetrafluoroethylene/hexafluoropropylene) (FEP) copolymers induced higher monocyte adhesion on surfaces containing carboxyl groups compared with an FEP surface with only amine groups. 27 Moreover, preadsorption of Fg onto three distinct polymers, previously coated with poly(2-methacryloyloxyethyl phosphorylcholine [MCP]-co-n-butyl methacrylate [BMA]), affected macrophage adhesion differently, according to the substrate used. Still, the number of adherent cells diminished in comparison to uncoated polymers and TCPS. 46 Our data suggest that, when combined with Ch, Fg does not affect macrophage density, morphological features, or fusion. These findings further corroborate a biomaterial-dependent effect of Fg on macrophage behavior and highlight the importance of protein coating to modulate cellular responses.
Following activation and depending on the signals they sense from the external milieu, macrophages release a variety of soluble mediators that will govern their function and interactions with other cells. The surface onto which this cell population adheres has been shown to influence such secretion.6,27,46 Our antibody array analysis revealed greater production of soluble factors by macrophages differentiated on RGD compared with those cultured on Ch-based films. Consistent with this, enhanced secretion of matrix metaloproteinases (MMPs) and TIMPs by RGD-differentiated macrophages, as well as of more cytokine production by RGD-adherent dendritic cells, has been previously described.48,49 We 22 and others 21 have recently reported that Ch is able to elicit an anti-inflammatory macrophage polarization. From a tissue-engineering perspective, induction of an anti-inflammatory pro-regenerative macrophage phenotype is of key relevance, as reparative macrophages are crucial in tissue remodeling after inflammation, guiding the host response to implanted biomaterials. Thus, the claim that Ch may be an adequate material for tissue-engineering applications, such as in bone repair,11,50 may be related to its ability to modulate the inflammatory response. Moreover, besides supporting attachment and proliferation of osteoblastic cells,16,51 Ch also drives macrophage production of soluble mediators involved in bone physiology. These include insulin and IGFBPs, which were detected in this investigation at high levels at all time points. Insulin enhances osteogenesis 52 and accelerates bone healing, 53 and IGFBPs are important regulators of bone metabolism. 54 In addition, increased amounts of ICAM-1, TIMP-1, TIMP-2, and TGF-β1 were also produced by fully differentiated macrophages (i.e., at day 10) cultured on Ch films. ICAM-1 is an important adhesion molecule involved in leukocyte extravasation to inflamed areas,55,56 but an active role for the membrane and soluble forms of the protein in osteoclast activity has also been reported. 57 As for TIMP-1 and TIMP-2, the high levels detected are in line with Ch-induced MMP9 secretion, 22 as TIMPs regulate MMP activity, and corroborate previous research work on MMPs/TIMPs production by biomaterial-adherent macrophages and FBCG. 48 Importantly, MMPs/TIMPs mediate many biological processes, namely inflammation, wound healing, and bone remodeling.48,58 Regarding the pluripotent cytokine TGF-β1, it is required for bone formation59,60 and fracture repair, 61 and plays a crucial role in bone turnover. 62 In future in vivo studies, it would be interesting to investigate cytokine production at Ch-scaffold implant sites.
Interestingly, macrophage activation on biomaterials does not correlate with macrophage adhesion or rates of fusion.6,63,64 Consistent with this, Ch+Fg films triggered a distinct cell activation, as assessed by the amount of soluble factors, than Ch films without adsorbed protein, despite supporting equivalent numbers of adherent and fusing macrophages. Previous studies have demonstrated that preadsorption of Fg affected macrophage release of pro-inflammatory cytokines, in a surface- and time-dependent manner. 46 In this work, we have observed that the majority of pro-inflammatory proteins is less abundant and produced at a lower rate when Fg is preadsorbed onto Ch. Of note, TNF-α, INFγ, and IL-17 were significantly down-regulated in the presence of Fg. Interestingly, no detectable amounts of these cytokines in the plasma of animals implanted with Fg-adsorbed Ch scaffolds were previously found. 33 On the other hand, after 10 days of macrophage differentiation on Fg-coated Ch films, despite a gradual increase over time (Fig. 5 and Supplementary Fig. S2), there was no significant up-regulation of TGF-β1 relative to Ch alone, as seen in vivo. 33 This discrepancy may be related to the different experimental conditions (in vitro vs. in vivo) and time of analysis (10 days vs. 2 months) used in this investigation and in our previous in vivo study with Fg-coated Ch. 33 Nonetheless, high levels of important bone-related mediators detected on Ch are also maintained in Ch+Fg films. Of note, MIP-1δ and BMP-5 are significantly up-regulated on Ch+Fg substrates, and BMP-7 and PDGF-BB followed the same trend. MIP proteins play a major role in the recruitment of immune cells to sites of injury. 65 In addition, MIP-1δ was recently described as an osteoclast-stimulating factor, specifically by potentiating the recruitment of osteoclast precursors and enhancing RANKL-induced osteoclast differentiation and activity. 65 Regarding BMPs, these members of the TGF-β superfamily of growth factors are involved in the development of different organs and tissues. 66 In bone, BMPs promote differentiation of mesenchymal cells into osteoblasts.66,67 BMP-5 was described to significantly enhance osteoclastogenesis when combined with BMP-2. However, combination of BMP-2, BMP-5, and BMP-6 stimulated osteoblasts but not osteoclastogenesis. 68 Similar to other BMPs, BMP-7- has osteogenic activity and has been shown to be up-regulated during bone healing.66,69 PDGF-BB is a potent chemotactic and mitogenic factor for osteoblasts and osteoblast lineage cells, and it up-regulates the expression of VEGF. 70 Moreover, it has been implicated in the formation of new bone in several in vivo studies.71–76 Interestingly, fully differentiated macrophages cultured on Fg also evidenced enhanced secretion of angiogenic-associated proteins. This suggests that when triggered by Fg, macrophages influence different key steps, are able to modulate the inflammatory response, and also contribute to new bone formation and angiogenesis, both of which were stimulated with Fg-adsorbed Ch scaffolds in vivo. 33 Furthermore, Fg has also been shown to improve recruitment of mesenchymal stem/stromal cells by NK cells 36 and to promote resorption of Ch by osteoclasts. 77 Taken together, these findings support the view that biomaterials combining Ch and Fg may constitute a complementary strategy for bone tissue engineering, by modulating the behavior of different cell populations and influencing diverse events, ranging from inflammation to biomaterial resorption.
Conclusions
In this work, we analyzed how Ch films modified or not with Fg affected monocyte/macrophage behavior in vitro. No differences in cell density, morphological features, and fusion were observed between Ch and Ch+Fg. Still, relative to RGD positive control, Ch substrates induced more elongated and interconnecting cells. Importantly, at day 10, Fg promoted significant up-regulation of soluble mediators involved in bone homeostasis, wound healing, and angiogenesis, such as MIP-1δ, BMP-5, GH, TGF-β3, and EG-VEGF. BMP-7 and PDGF-BB followed the same trend. Taken together, these findings suggest that Fg adsorbed on Ch films modulates macrophage response toward a bone remodeling/regenerative phenotype.
Footnotes
Acknowledgments
The authors would like to thank Pieter Faber (Cleveland Clinic Genomics Core, Cleveland, Ohio, USA) for help with the microarray reading. This work was financed by FEDER funds through the Programa Operacional Factores de Competitividade – COMPETE and Programa Operacional Regional do Norte (ON.2 - O Novo Norte), in the framework of the project “NORTE-07-0124-FEDER-000005 – Project on Biomedical Engineering for Regenerative Therapies and Cancer”, under Quadro de Referência Estratégico Nacional (QREN), and by Portuguese funds through FCT – Fundação para a Ciência e a Tecnologia in the framework of the project PTDC/SAU-BEB/099954/2008. J.M. and M.I.O. are grateful to FCT for awarding them a doctoral (SFRH/BD/37582/2007) and a post-doctoral (SFRH/BPD/37090/2007) fellowships, respectively. Partial support for the study was also provided by a grant from the NIH-NIBIB, #EB000282.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.