
Abstract
Isolated islets can provide a source of tissue for research, transplantation, and drug discovery to develop therapies for diabetes. Empirical modeling of islet diffusion barriers demonstrated that only the outermost layers of cells were exposed to glucose and sufficient oxygen levels, resulting in core cell death. Islets under a diameter of 100 μm exhibited a lower diffusion barrier, superior survival rates, and improved functional properties. Utilizing these observations, we engineered optimal islets by dispersing them into single cells and reaggregating them over several days in a micromold. These custom-designed micromolds contained conical-shaped recesses that enhanced reaggregation of cells into a defined geometry. The engineered islets, or Kanslets, were all under 100 μm in diameter, and had the same general cellular composition as native islets. Kanslets continued to produce new insulin molecules and had microvilli on the islet surface, much like native islets. The engineered islets had a statistically higher viability (percent of live cells), and increased glucose diffusion compared to native islets. In addition, they remained responsive to varying glucose levels by secreting insulin. When transplanted into diabetic rats, engineered islets performed reduced random blood glucose to normal levels within 48 h. Optimally, engineering islets may be a suitable alternative to utilizing native, isolated islet tissue for a variety of applications. Reaggregating tissue in an optimized manner using our engineered micromold approach has immense impact for three-dimensional tissue production and its subsequent use in research, drug discovery, and the clinic.
Introduction
Isolating islets from the pancreas of cadaveric donors provide tissue that can be used for both transplantation and drug discovery. Significant strides have been made to offer pancreatic islet transplantation as a means to achieve insulin independence.2–6 With the advent of the Edmonton protocol, recipients were able to maintain normoglycemia for a full year after islet transplantation. 7 Unfortunately, regardless of these results, long-term success is limited and patients are subject to receiving multiple transplants to maintain insulin independence.8,9 With each transplant requiring islets from two to three donor pancreata and patients needing subsequent transplantations, a large burden is placed on the already limited donor availability, preventing the procedure from being accepted as a clinical standard and reaching the greater population.
Once isolated from their natural location within the pancreas, islets exhibit diminished survival and function, both in vitro studies and soon after transplantation.10–13 Within the pancreas, islets are immersed with their native blood supply. After isolation, diffusion becomes the primary means of oxygen, glucose, and nutrient transport into the core of isolated native islets. 13 Revasularization is essential for islet survival and function after transplantation, but this process can take several days or even weeks to occur.14–16 Diffusion barriers, particularly with respect to oxygen, affect the maintenance of cell viability and performance. The limited diffusion of oxygen into the core of a native islet can lead to reduced function, core cell death, and eventual islet loss. Studies have reported that the size of an islet has a profound effect on its survival and function.10,11,13,17 Several studies have demonstrated that small islets (diameter <125 μm) were superior large islets (diameter >150 μm) in vitro, and showed increased insulin independence when transplanted.10–13 Mathematical modeling of the diffusion of oxygen into the islet core demonstrated the absence of core cell death in islets with a radius less than 50 μm. 13 This was supported by stained images showing apoptotic and necrotic cells within the cores of large islets in culture as early as 24 h after isolation. Similarly, the direct glucose diffusion into islets demonstrated that glucose penetrated the core of small islets, while only the outermost layers of cells were penetrated in large islets. 13 The combination of these factors decreases islet survival and overall graft performance.
Engineering optimal islets provides a means to overcome the diffusion barriers affected by islet size limitations. Although dispersing islets into single cells would greatly decrease any diffusion barriers, it is well known that islet cells function better when clustered and are highly dependent on cell–cell communication and the paracrine interactions between the different cell types.18–28 For several decades, researchers have witnessed the spontaneous reaggregation of dispersed islet cells and have utilized this in reconstructing islet-like tissues.29–34 Traditionally, islet cells have been incubated in a culture medium and allowed to reaggregate into pseudoislets of a variety of shapes and sizes, often times resulting in pseudoislets much larger than 200 μm in diameter. The problem with this methodology is the high probability of regeneration of large cell clusters—reintroducing the diffusion barrier, inferior viability, and reduced function. Understanding these size limitations, engineering islets of a small size (<125 μm) will help overcome these issues.
Limited studies have been conducted aimed at engineering islets of a defined size, due to complications that may be inherent within their respective techniques. In 2007, islets of a standard size were engineered utilizing the hanging drop technique. 35 Although this technique allowed for the creation of uniform islet cell clusters, it required immense manual labor with individually placing drops and lacked the ability of scale up for mass production. Even with recent availability of hanging drop plates and the use of automated instrumentation, the technique is still low output and not an ideal solution. A tool that can allow controlled and optimized reaggregation of islet cell clusters can have a considerable impact on islet production for research and transplantation.
In this study, we focused on limiting the diameter during reaggregation of islet cells to that of small islets (<125 μm). We engineer optimal islets, or Kanslets, in vitro by reaggregating dispersed islet cells on a glass mold containing microrecesses of a defined geometry. Use of this tool resulted in the production of uniform, reproducible Kanslets. We assessed several characteristics of the Kanslets and compared their function to that of isolated native islets. Finally, we transplanted the engineered Kanslets into diabetic rats and compared the response to matched animals receiving native islets. In summary, we have developed a tool and a high-output technique for the uniform production of reaggregated islets of an optimal geometry. Kanslets can provide more viable tissue for use in research and transplantation.
Materials and Methods
Design and fabrication of micromolds
Glass substrates, precut in a disc shape were ordered from Precision Glass and Optics. The glass substrates were cleaned using acid and base piranha solutions and dried at 200°C to ensure the surface was free of moisture. Glass micromolds were fabricated through a multistep process that included thin-film deposition, photolithography, and wet etching techniques. Briefly, one surface of the glass substrates was sputtered with a layer of chromium (Lesker Thin Film Deposition System). Positive photoresist (AZ1518) was spin-cast onto the chromium surface and prebaked at 100°C for 2 min. A transparency mask template was created containing the defined geometry and layout of wells to be etched using computer-assisted design (CAD) in AutoCAD software (Autodesk) and high-resolution transparency masks were printed. The photoresist-coated discs were exposed to UV-light through the transparency mask for 4 s. Patterned glass was then postbaked at 100°C for 10 min, and then immersed in a developer (AZ 300 MIF Developer) to render the image. The chromium layer was subsequently etched (CR7S Chromium Etchant). Glass was washed with water and dried with nitrogen.
To etch the pattern into the surface of the glass, the disc was wet etched by immersion in a buffered oxide etch solution containing a 14:20:66 ratio of HNO3 to HF to H2O, respectively. A profilometer (TencorAlphastep 200) was used periodically to measure the etched surface and adjustments were made accordingly. The photoresist and chromium layers were removed to reveal the etched micromold.
To contain the cell culture on top of the glass micromold, a polydimethylsiloxane (base and cross linker; Dow Corning) ring was cast to surround the glass micromold disc. All micromold components were cleaned and steam sterilized before any cell culture use or experimentation.
Islet isolation, dispersion, and reaggregation
Pancreatic islets were isolated and dispersed using our previously published procedures.11,13,36 Briefly, rats were anesthetized by intraperitoneal injection of a mixture of ketamine and xylazine. After the peritoneal cavity was exposed, the pancreatic main duct to the duodenum was clamped, cannulated in situ via the common bile duct, and distended with cold collagenase (CLS1, 450 units/mL; Worthington). After excision, the pancreas was incubated for 20–30 min with gentle tumbling in a 37°C incubator. The contents of the tube were washed, passed through a 100-μm mesh screen, and sedimented in a refrigerated centrifuge. The pellet was mixed with Histopaque (density=1.1085) and centrifuged. The islets, collected from the gradient, were sedimented and washed over a sterile 40-μm mesh cell strainer. Islets were placed into a modified Dulbecco's modified Eagle medium (DMEM)/F-12 medium supplemented with 10% fetal bovine serum (FBS), 1% antibiotic/antimycotic and allowed to recover overnight in an incubator at 37°C and 5% CO2.
Isolated islets were dispersed into single-cell suspensions. Islets were washed twice with a calcium- and magnesium-free Hank's balanced salt solution (cmf-HBSS) before addition of a digestion medium consisting of cmf-HBSS supplemented with 4.8 mM HEPES and papain (5 units/mL; Worthington). Suspensions were incubated on a rotator at 37°C for 20 min. Islets were dispersed by trituration using a pipette until the cell suspension primarily contained single cells. The cells were then washed to remove residual papain and transferred to a customized DMEM:F12-based, serum-free Kanslet aggregate culture medium. Occasionally, a sample was taken and cell counts and yield were determined using a hemocytometer.
The single-cell suspension was then carefully plated onto sterile glass micromolds. Within several minutes, cells began to settle into the recesses of the micromold and were in close proximity to each other allowing cell–cell readhesion. Micromolds were incubated for 6–7 days at 37°C and 5% CO2. Cells reaggregated, while limiting the size to no bigger than the recess allowed. The aggregate culture medium was changed every 24–48 h until reaggregated islets were formed. Kanslets were removed by simply washing the micromold several times with the culture medium until islets dislodged and were aspirated with a pipette.
Viability
Viability of Kanslets was assayed by placing them in phosphate-buffered saline (PBS) in an Attofluor Chamber. Apoptotic and necrotic fluorophores (Propidium Iodide and YO-PRO-1; Vybrant Apoptosis Assay Kit #4, Molecular Probes, Invitrogen) were added and incubated at room temperature for 15–30 min. After staining, confocal images were collected through the center of the islet/Kanslet. Samples were imaged on an Olympus FluoView 300 confocal microscope housed at the Diabetes Research Lab. The area of positively stained cells was measured and divided by the total islet area to determine the percent viability. All images were collected within 60 min of initial exposure to the fluorophores.
A hemocytometer was used to estimate the cell number before reaggregation of single, dispersed islet cells. The islet diameter was measured using FluoView software (Olympus Corporation) and size distribution was calculated.
Electron microscopy
Native islets and Kanslets were handpicked and placed in 2% glutaraldehyde in 0.1 M sodium cacodylate solution and stored at 4°C until imaging. Samples were rinsed twice in 0.1 M sodium cacodylate buffer for 10 min before postfixation in 1% osmium tetroxide for 1 h. Rinsing with distilled water was followed by a graded ethanol dehydration (30%, 70%, 80%, 95%, 100%) for 10 min each. Samples were rinsed twice in propylene oxide for 15 min before being infiltrated in a mixture of propylene oxide and Embed 812 resin (Electron Microscopy Sciences) overnight. BEEM embedding capsules were used to embed the samples in fresh resin before curing overnight in a 70°C oven. Thin sections, 80 nm in diameter, were cut using a Leica UCT ultramicrotome and placed on 300 mesh thin bar grids. Contrast was applied to the sections by adding uranyl acetate followed by Sato's lead stain. Images of islets were captured from random sections using a J.E.O.L JEM 1400 transmission electron microscope.
Glucose diffusion and uptake
Glucose diffusion and uptake experiments were completed using a confocal microscope as described previously. 13 A PBS solution containing 0.5 mM 2-[N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino]-2-deoxy-D glucose (2-NBDG) (Molecular Probes, Invitrogen) was perifused into a chamber containing Kanslets. Images were captured every 2–5 s upon commencing 2-NBDG perifusion and subsequently decreased after several minutes for the remainder of the experiment. The rate of glucose was measured from the time the outer cells became fluorescent to when the core cells became fluorescent. The percentage of 2-NBDG uptake was calculated by measuring the area of fluorescence within the islet that was higher than the background and dividing by the total islet area.
Immunofluorescence and cellular composition
Kanslet samples were fixed in 4% paraformaldehyde in 0.1 M PBS (pH 7.4). Cellular composition of Kanslets was determined by triple-staining islets for insulin, glucagon, and somatostatin. The following primary antibodies were used: anti-insulin (Abcam or Santa Cruz Biotechnology), anti-glucagon (Abcam), and anti-somatostatin (Abcam). Corresponding secondary antibody conjugates included: Cy2 (Jackson ImmunoResearch Laboratories, Inc.), Alexa 647, and Alexa 555 (Molecular Probes, Invitrogen).
Images were obtained using a Nikon C1Si or a C1Plus confocal microscope. Images were acquired at 10×–100× and analyzed using FluoView or Adobe Photoshop software. To determine the cell composition, the proportion of immunolabeled cells (alpha-, beta-, and delta-cells) was evaluated by counting the number of individual visible cells and dividing them by the total number of cells per islet within the confocal image. Kanslet cellular composition was compared to native islets. Proinsulin (an insulin precursor molecule) (Abcam) and insulin were stained to visualize production of new insulin molecules.
Static insulin secretion (static incubation)
Kanslets were equilibrated overnight in the DMEM/F-12 medium containing 5 mM glucose and 10% FBS (low-glucose medium). Islets were handpicked using a micropipette and a known quantity of islet equivalents (IEQs) were distributed in 24-well plates. Kanslets were subject to low glucose (5 mM), high glucose (17.5 mM), or high glucose with KCl (30 mM). After 60 min of static incubation at 37°C and 5% CO2, conditioned media samples were collected and frozen at −80°C. The insulin concentration was later quantified using an insulin ELISA kit (Alpco).
Dynamic insulin secretion (perifusion incubation)
Known quantities of native islets and Kanslets were placed into chambers of a perifusion system at a constant flow rate (250–300 μL/min) at 37°C for 30 min. After a 30-min preincubation at a basal glucose level (3 mM), the samples were then subjected to three testing conditions during a 3-h incubation period: 60 min of basal condition (3 mM glucose), then 60 min of high glucose (20 mM), followed by 60 min of basal condition (3 mM glucose). During the perifusion, samples of the conditioned medium were collected from the output fraction every 10 min. Samples were frozen at −80°C and later analyzed for insulin content using ELISA.
Transplantation
Islet transplantation was completed using our published protocols.11,13 Diabetes was induced in Sprague-Dawley rats by intraperitoneal injection of streptozotocin (65 mg/kg body weight). Rats were monitored for 1 week to ensure that they were diabetic, with blood glucose levels well above 350 mg/dL. They were injected with long-acting Lantus insulin if blood glucose levels were too high. Animals were then divided randomly into three groups: diabetic control (sham procedure), 1500–2000 IEQ native islet transplantation, or the same volume of Kanslets. On the day of transplantation, rats were anesthetized with 45 mg/kg pentobarbital. The incision site was shaved and cleaned with betadine scrub, and an incision made in the body wall on the left flank. The kidney was exposed through the wound and a small incision made into the kidney capsule. Using a small-bore pipette, native islets or Kanslets were placed under the capsule. The kidney was returned to its original position, the muscle was sutured, and the skin closed with wound clips. Novolog insulin (0.55 units/kg) injections were given twice daily post-transplant until transplants provided the recipient with necessary insulin. Blood glucose levels were checked via blood drawn from the tail each morning at approximately the same time.
Statistics
Measurements were performed in duplicate or more, with results calculated as averages with standard error. Statistical significance was evaluated using a Student's t-test with significance defined as p<0.05.
Results
Size and morphology
Visualization of the reaggregated Kanslets indicates that they are below 100 μm in diameter, as shown in Figure 1A. These islets generally measured less than 100 μm with an average diameter of 40.85±0.39 μm. In contrast, reaggregated islet cells that were not constrained by the micromolds often reaggregated in an unconstrained manner forming mega-islets measuring more than 200 μm in diameter (Fig. 1B). A size histogram of Kanslets compared to native rat islets is provided in Figure 1C.
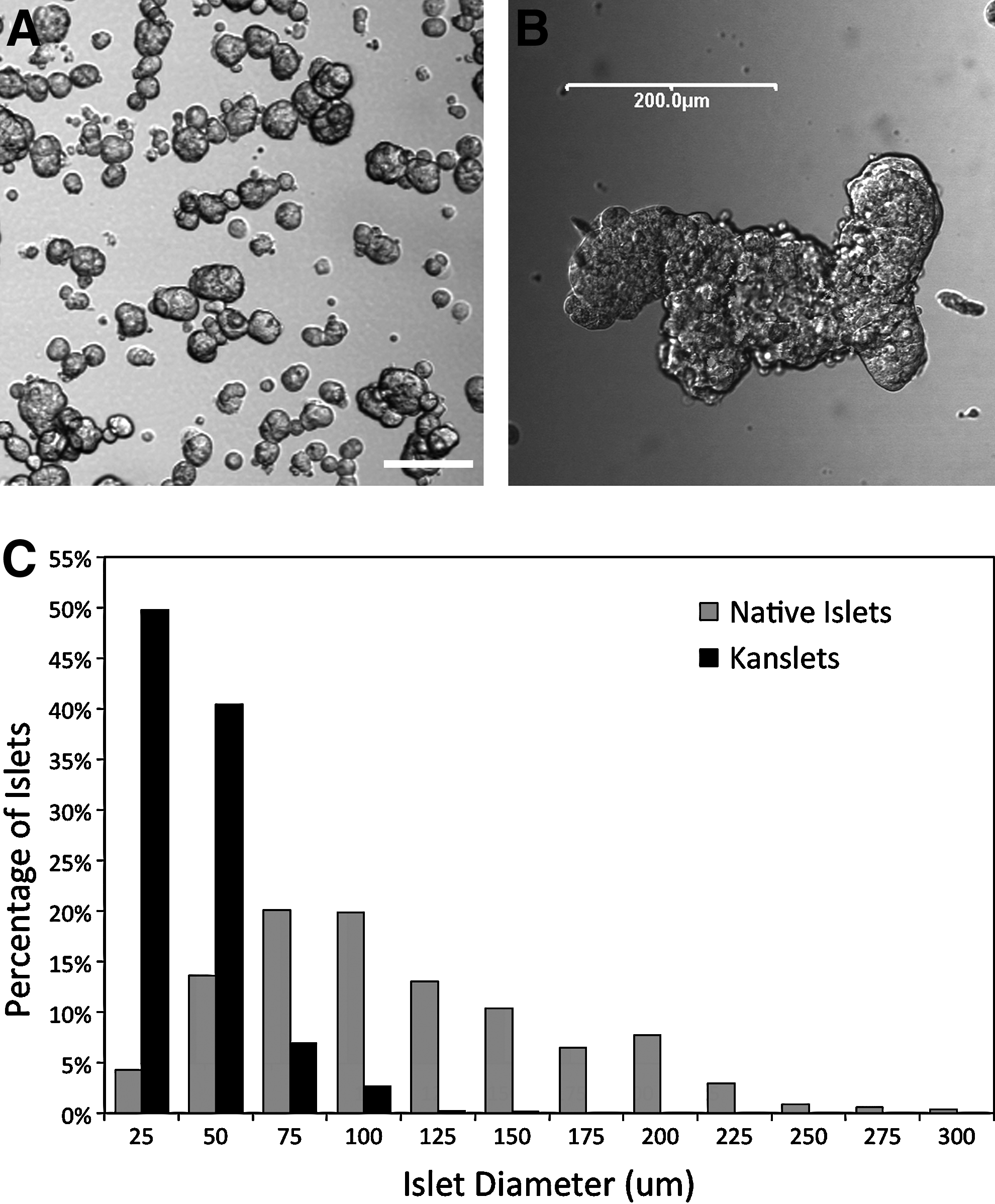
Uniformity of size.
It is important that the Kanslets return to native islet morphology after formation, including the appearance of microvilli covering the surface of the islet. Figure 2A and B compare the islet/Kanslet general morphology illustrating the similarity. Electron micrographs highlight the microvilli that cover the surface of a native islet (Fig. 2C). The microvilli begin to appear on the surface of the Kanslet within 10 days after reaggregation and removal from the micromold (Fig. 2D).
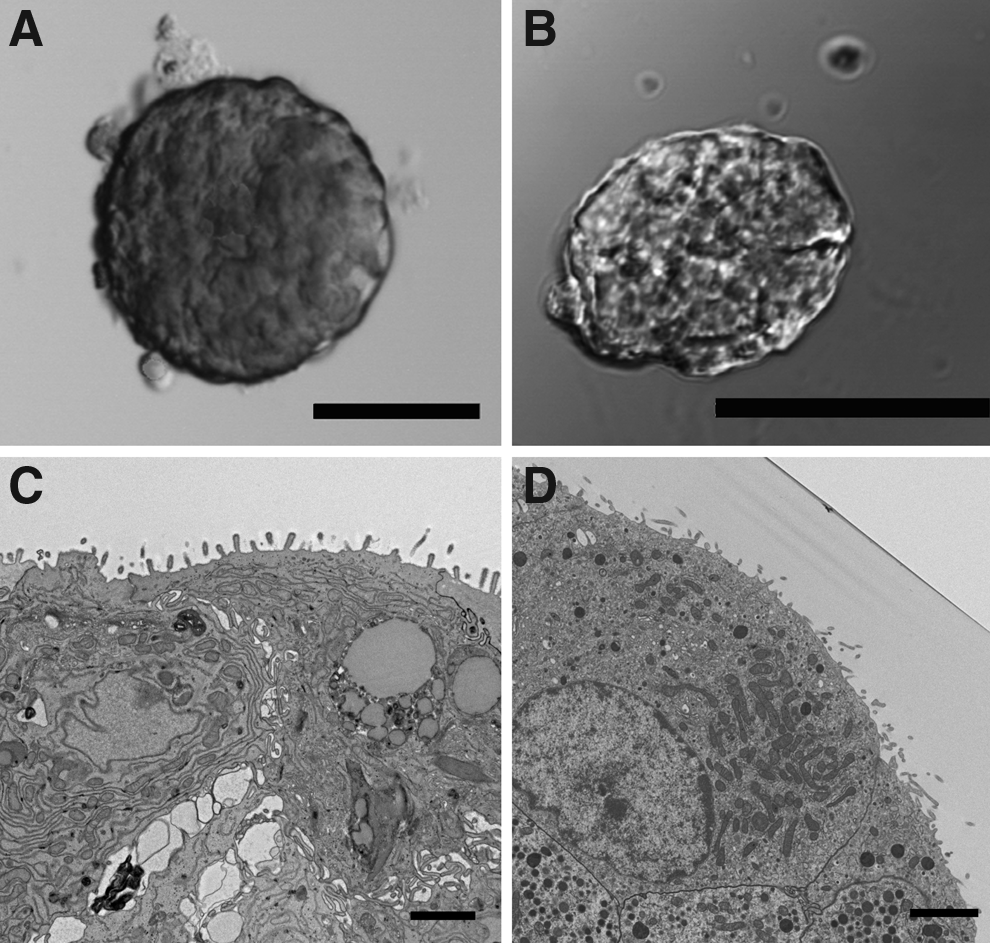
Morphological similarity between native islets and Kanslets. Native small islet
Viability
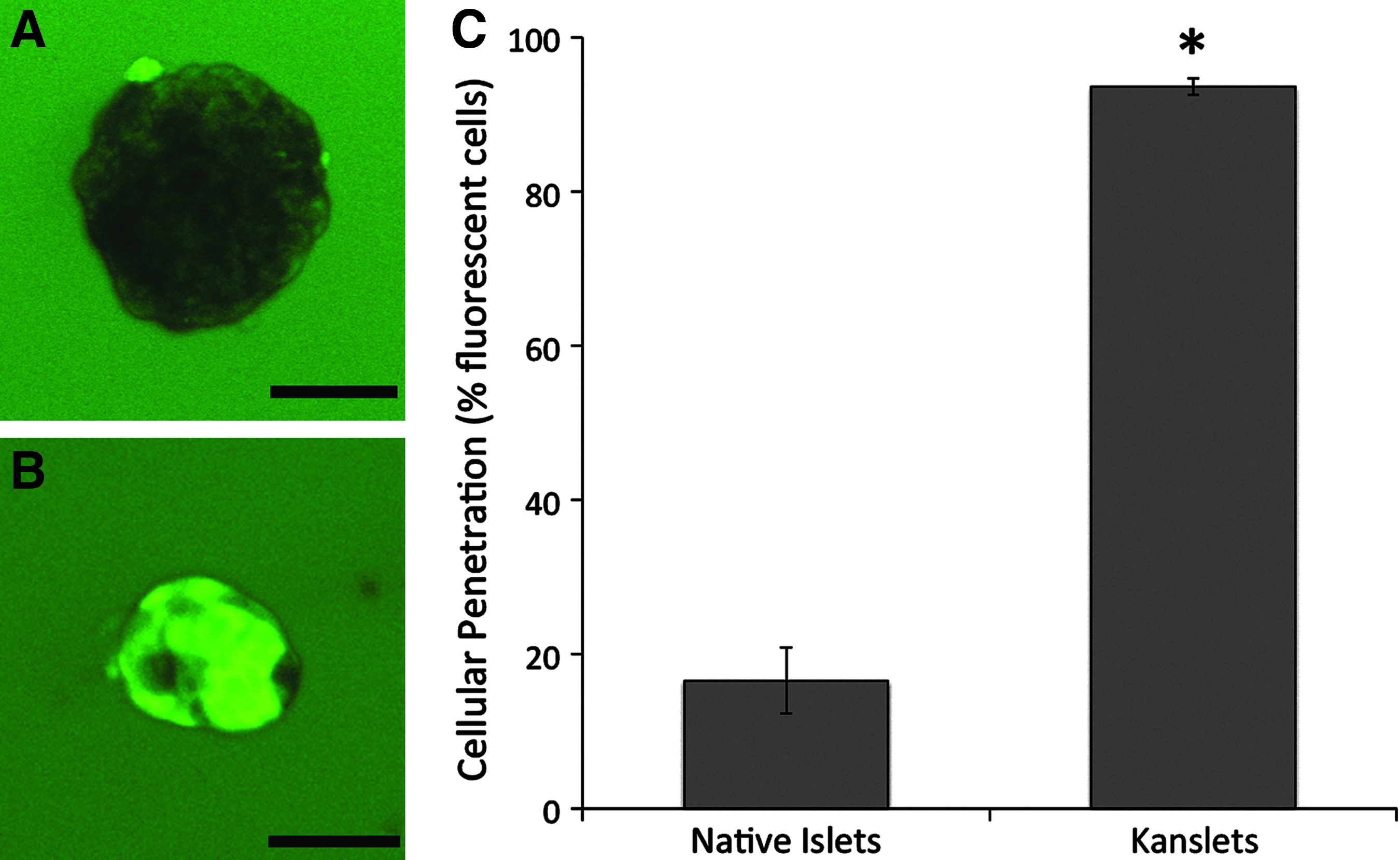
To determine the percentage of viable cells within Kanslets, a viability assay was performed on day 5 (after reaggregation of islet cells). Very few Kanslets exhibited any dead cells, with some exhibiting a few random dead cells. For the most part, dead cells were attached to the surface of the Kanslets, and visibly were not incorporated under the islet capsule (Fig. 3A). On average, Kanslets presented viability over 99.7%±0.1%, in comparison to 72.4%±2.8% viability in the native islets (Fig. 3B).

Kanslet viability.
Glucose diffusion and uptake
To compare the diffusion of glucose within Kanslets to the previously published work on native islets, we used a fluorescent glucose analog, 2-NBDG. Islets were exposed to 0.5 mM 2-NBDG though perifusion and monitored under a confocal microscope. Glucose diffused into the intracellular spaces almost as immediately as glucose was apparent in the viewing field. Figure 4A shows the lack of glucose diffusion into a native islet compared to full penetration in a Kanslet (Fig. 4B). In fact, the Kanslet showed a superior glucose uptake with diffusion rates that were beyond our ability to resolve with an imaging scan speed of 2 scans/s. As soon as the glucose-containing media came in contact with the Kanslet, it entered the core. When 2-NBDG fluorescence was analyzed, it was clear that a higher percentage of Kanslet cells were permeated with 2-NBDG compared to native islets (Fig. 4C).
Diffusion barrier of Kanslets.
Cellular composition
Immunofluorescence staining was conducted to ensure that Kanslets were similar in cellular composition to their native counterpart. Kanslets, triple-stained for insulin, glucagon, and somatostatin, indicate that the three major endocrine cell types were present in our engineered constructs (Fig. 5A). The distribution of these cell types across the islet appears to be in a random order. In general, the composition was not radically different from native islets, even though Kanslets consisted of a significantly larger portion of beta-cells compared to native islets (Fig. 5B).

Cellular composition of Kanslets.
The production of new insulin molecules was identified by staining for the insulin precursor molecule, proinsulin. The presence of proinsulin in islets indicates the synthesis of new insulin molecules (Fig. 6A). Kanslets also demonstrated the presence of proinsulin levels even 7 days after reaggregation in a pattern similar to native islets (Fig. 6B).

Proinsulin production in Kanslets. Proinsulin (red) is present along with insulin (green) in both isolated native islets
Glucose-stimulated insulin secretion
In static incubation studies, Kanslets were subjected to low- and high-glucose concentrations in static culture. Kanslets responded to glucose as expected with increased insulin secretion as the glucose level increased (Fig. 7A). Kanslets subjected to high glucose resulted in a significant increase in insulin output, double that of the basal level (p<0.05). Exposure to high glucose and potassium (membrane-depolarizing agent) further increased this output to four times the insulin secreted at a basal level (p<0.001). This was also significantly higher than the output at high glucose alone (p<0.001).

Insulin secretion from Kanslets.
Perifusion experiments were completed to test the insulin secretion over time (dynamic secretion) in response to varying glucose levels. Kanslets were subjected to basal glucose, then high glucose, and again basal glucose over a 3-h period. Insulin secretion remained stable in response to basal glucose, and resulted with a spike in insulin secretion immediately upon exposure to a high-glucose concentration. Insulin secretion more than doubled in response to the presence of high glucose (Fig. 7B). These values were normalized by volume (IEQs) and by cell number, 37 and showed little difference between calculation methods.
Transplantation
Streptozotocin-induced diabetic rats received transplantation of 1500–2000 IEQ of either native islets or Kanslets under the kidney capsule or a sham procedure (control). Post-transplant nonfasting blood glucose values were monitored each morning following the transplants. With transplants of either native islets or Kanslets, diabetes was reversed within 48 h (Fig. 8). Both groups continued to maintain low nonfasting blood glucose levels over the 2-week period, which were not statistically different (except for day 7). Specifically, all average values for the Kanslet group were below 250 mg/dL. In contrast, rats receiving native islets had some readings over 300 mg/dL after the transplant. The sham diabetic group failed to reach normoglycemic levels for any of the measurements taken, and was significantly higher than either transplant group for each day following the procedure.

Transplantation of native islets or Kanslets lowered random blood glucose. Native islets and Kanslets were transplanted into streptozotocin-induced diabetic Sprague-Dawley rats (three rats per group). Rats receiving the same volume of either native islets (filled circles) or Kanslets (open circles) had significantly lower nonfasting blood glucose levels for the 2-week period following transplant. Rats receiving Kanslets had blood glucose levels <250 mg/dL on average for each day after the transplant. Control diabetic rats that did not receive transplants continued to be hyperglycemic throughout the experiment. Day 0 values were obtained on the morning before the procedure.
Discussion
In this study, we utilized a novel method for the uniform production of engineered islets optimized to reduce diffusion barriers. Kanslets provide a feasible alternative to using native islet tissue in research and transplantation. The process minimizes the diffusion barriers necessary for transport of key nutrients, including oxygen and glucose, allowing for increased viability and long-term performance. Uniform, engineered islets were produced by dissociating islets into single cells and utilizing a micromold to optimally reaggregate the cells. This uniformity of size was limited to a maximum diameter of 125 μm, because the critical oxygen concentrations resulted in core cell death above this diameter as previously described.11,13 By limiting the diameter of this engineered islet tissue, we were able to increase the viability to nearly 100%; a significant improvement over previously published data on native islets. 11 It is likely that the engineering process aids in the elimination of nonviable or dead cells, as islet cells reaggregated to form new clusters.
Further, experiments with diffusion of glucose molecules showed a reduction in, or lack of, a glucose diffusion barrier as Kanslets were nearly instantaneously infiltrated with the fluorescent tag. This is in contrast to the published diffusion rate of 2.8 μm/min into native large islets, indicating an extremely slow penetration. 13 Previous experiments with glucose diffusion into native islets indicated that even after 1-h of incubation, fluorescent glucose penetrated only the outermost layers of a large islet, while diffusing with an increased efficiency into small islets. 13 Our results replicated previously published data, while using a much smaller concentration of labeled glucose (0.5 mM 2-NBDG). 13 Our results show that the diffusion barrier to glucose was markedly reduced and support the necessity to produce islets less than 100 μm in diameter.
The engineering process allowed for successful production of islets that contained alpha-, beta-, and delta-cells, and was similar in composition to their native islet counterparts. It is unknown why there is a slight increase in beta-cells in Kanslets and may be due to the engineering process. It is possible that during islet isolation from the pancreas, a portion of the outer islet cells, which are predominantly alpha- and delta-cells may be lost. This is not necessarily a negative outcome as this restructured composition may be more ideal compared to native islets for both research and transplantation. Allowing the islet cells to self-aggregate into an islet may allow for cell–cell communication and signaling that leads to an increased incorporation of beta-cells. Given that native islets contain blood vessels, there is a great probability that insulin will be secreted close to a vessel. In the Kanslets, diffusion is the primary method of insulin release into the media. Thus, having an abundance of beta-cells on the surface increases the chances of glucose detection and corresponding insulin secretion. The positive staining for proinsulin indicates that even after the reaggregation engineering process, Kanslets are still capable of producing new insulin molecules.
Given the similarities in islet composition, it is not surprising that Kanslets responded to changes in glucose levels as expected. Kanslets secreted more insulin when exposed to high-glucose levels over the basal level. This increase could be witnessed in static and dynamic experimental conditions, with insulin levels returning to normal after high-glucose stimulation ceased in dynamic experiments.
Kanslets were able to provide a transplantable solution that matches the effect of native islets. As expected, there was a reduction in blood glucose levels in diabetic rats that received the Kanslet transplant. More notably, these levels were similar to those produced by the native islet transplant group. In this study, rats were transplanted with approximately the same amount of islet tissue. Although our findings show similar results between treatments, it is uncertain at this time whether Kanslet transplants are more potent or outperform native islets with a lower transplanted tissue volume. Kanslets do provide an alternative solution to conventional islet transplantation. Subsequent studies will be necessary to determine the dosage comparison between native islets and Kanslets, and the marginal mass required of Kanslets required to achieve normoglycemia.
With previous studies validating the superiority of small islets to large islets, there was a shift to preferentially utilizing small islets for both research and transplantation.10–12 However, this preference leaves significant numbers of large islet tissue unused. The driving force behind our process was to recreate small islets from native islet cells and limit islet size during the process. This has implications both in research and in clinical transplants. The use of this highly viable tissue in transplantation may increase overall success rates. In research, the production of uniform islet tissue allows for less error between islet preparations and a higher reproducibility of data. Utilizing this technology for research and transplantation makes significant strides toward developing the next generation of treatments for diabetes.
Footnotes
Acknowledgments
We wish to thank Barbara Fegley and Tiffany Schwasinger-Schmidt for assistance with electron microscopy. We also wish to thank Dr. Karen Kover for assistance with islet transplantation. This work is supported by the Emilie Rosebud Diabetes Research Foundation (L.S.B.) and the Institute for Advancing Medical Innovation (L.S.B. and K.R.).
Disclosure Statement
The authors do not have any financial or commercial interest to disclose at this time.