
Abstract
Decellularized equine carotid arteries (dEAC) are potential alternatives to alloplastic vascular grafts although there are certain limitations in biocompatibility and immunogenicity. Here, dEAC were coated with the matricellular protein CCN1 and evaluated in vitro for its cytotoxic and angiogenic effects and in vivo for cellular repopulation, local biocompatibility, neovascularization, and immunogenicity in a sheep model. CCN1 coating resulted in nontoxic matrices not compromising viability of L929 fibroblasts and endothelial cells (ECs) assessed by WST-8 assay. Functionality of CCN1 was maintained as it induced typical changes in fibroblast morphology and MMP3 secretion. For in vivo testing, dEAC±CCN1 (n=3 each) and polytetrafluoroethylene (PTFE) protheses serving as controls (n=6) were implanted as cervical arteriovenous shunts. After 14 weeks, grafts were harvested and evaluated immunohistologically. PTFE grafts showed a patency rate of only 33% and lacked cellular repopulation. Both groups of bioartificial grafts were completely patent and repopulated with ECs and smooth muscle cells (SMCs). However, whereas dEAC contained only patch-like aggregates of SMCs and a partial luminal lining with ECs, CCN1-coated grafts showed multiple layers of SMCs and a complete endothelialization. Likewise, CCN1 coating reduced leukocyte infiltration and fibrosis and supported neovascularization. In addition, in a three-dimensional assay, CCN1 coating increased vascular tube formation in apposition to the matrix 1.6-fold. Graft-specific serum antibodies were increased by CCN1 up to 6 weeks after implantation (0.89±0.03 vs. 1.08±0.04), but were significantly reduced after 14 weeks (0.85±0.04 vs. 0.69±0.02). Likewise, restimulated lymphocyte proliferation was significantly lower after 14 weeks (1.78±0.09 vs. 1.32±0.09-fold of unstimulated). Thus, CCN1 coating of biological scaffolds improves local biocompatibility and accelerates scaffold remodeling by enhancing cellular repopulation and immunologic tolerance, making it a promising tool for generation of bioartificial vascular prostheses.
Introduction
Therefore, the search for alternatives circumventing these problems has led to the generation of tissue-engineered vascular grafts. These consist of synthetic (e.g., various polymers) or biological scaffolds (e.g., acellular vessels) and an autologous cellular compound.2–4 In particular, decellularized arteries can be regarded as highly promising vascular grafts as they combine suitable biomechanical properties, a geometry and architecture resembling the native vessel as well as the enormous potential to become integrated into the recipients organism by remodeling processes. 5 Although cellular in vivo repopulation of these vascular scaffolds tends to occur to a certain extend spontaneously, there have been reports that in vitro reseeding with autologous cells improves patency rates,6,7 reduces neointima formation 7 and local inflammatory response 8 leading to a significant improvement of the grafts' acceptance. These findings highlight the necessity for vascular grafts to be autologized before implantation. However, the technical prerequisites for in vitro reseeding of scaffolds are considerably demanding. Taking into account the often poor health status of patients requiring an implant, suitable autologous cell sources have to be identified and evaluated.4,9 Also, cell culture conditions in a bioreactor, including the evaluation of flow dynamics have to be established with the aim of reproducible quality of the cell components and the complete construct. 10
Because of the complexity of in vitro tissue-engineering techniques, in situ tissue engineering or so-called guided tissue regeneration has emerged. 11 This approach is defined as coating a scaffold with a homing factor, which subsequently enhances in vivo adhesion of circulating endothelial cells (ECs) or endothelial progenitor cells (EPCs) by interacting more or less specifically with membrane structures. The supported adhesion to a scaffold facilitates its repopulation and thus, shortens the period for which the unrepopulated surface is exposed to the blood stream. Among the molecules, which have been tested for in situ endothelialization, peptides or proteins displaying the RGD structure are considered to be most effective (as reviewed in 11 ). RGD is a common motif of extracellular matrix proteins binding as a ligand to integrins (αvβ3), which are expressed by ECs and EPCs and thus, serves as an anchor for cell adhesion.12,13 Basically, a similar mechanism has been described for CCN1, a protein of the CCN family of extracellular matrix-associated or matricellular proteins. 14 CCN1 interacts with αvβ3 integrins although it does not contain the canonical RGD sequence. Thus, CCN1 should facilitate the homing of EC- and EPC-like RGD peptides. Indeed, it has recently been shown that coating of cell culture dishes and of a biological decellularized scaffold with CCN1 enhanced adhesion of ECs. 15 Furthermore, CCN1 has been shown to have functional properties like induction of angiogenesis, 16 wound healing, 17 and modulation of inflammation, 18 making CCN1 a highly potential candidate to ameliorate integration and remodeling of vascular grafts.
To test the hypothesis that CCN1 indeed improves bioartificial vascular grafts, we coated decellularized equine carotid arteries (dEAC) with CCN1 and evaluated in vitro its functionality and its effect on the cell viability of fibroblasts and human umbilical vein endothelial cells (HUVECs) and in vivo its effect on scaffold repopulation, biocompatibility, and remodeling properties as well as on graft immunogenicity in a sheep model. Furthermore, angiogenic properties of CCN1-coated matrices were investigated by an in vitro three-dimensional (3D) tube formation assay. To our knowledge, this is the first time that CCN1 coating of a scaffold was comprehensively evaluated with respect to its functional properties for tissue integration and remodeling.
Materials and Methods
Decellularization of equine carotid arteries
EAC were obtained from a local slaughter house and stored in a cold 0.9% NaCl+1% penicillin/streptomycin solution until further processing. The adjacent soft tissue was carefully removed before the decellularization process was initiated. Briefly, EAC pieces of 12-cm length were disinfected with 70% ethanol, rinsed with 0.9% NaCl, and were then transferred to the decellularization solution containing 0.5% sodium dodecyl sulfate and 0.5% sodium deoxycholate (both from Roth, Karlsruhe, Germany) for 36–42 h. After intense washing with water (three cycles for 15 min) and 0.9% NaCl (eight cycles for 12 h), EAC were treated with endonuclease (125 U/mL; Merck) for 20–26 h at 37°C. The resulting dEAC were stored in the 0.9% NaCl+1% penicillin/streptomycin solution at 4°C until further use.
To confirm acellularity of the matrices, the DNA content was quantified by Quant-iT Picogreen Assay (Invitrogen) according to the manufacturer's instructions. Sterility was tested by incubation of 5 vol. of the last washing solution with 1 vol. 6×caso media (Roth) for 14 days at 37°C.
Sterile matrices with a DNA content <1.0 ng/mg tissue were considered to be acellular and suitable for implantation.
Graft coating
For coating with CCN1 (recombinant human CCN1; R&D Systems), dEAC were filled with 100 ng/mL CCN1 in EBM-2 media (Lonza), placed in a 50-mL tube, covered with the same solution, and incubated at 37°C for 24 h. CCN1-coated EAC were rinsed three times with 0.9% NaCl and stored at 4°C for maximal 7 days until implantation.
Effect of CCN1 on culture of L-929 cells
L-929 mouse fibroblasts were cultivated in 24-well plates (2500 cells/well) in the RPMI medium containing 10% fetal calf serum (Biochrom) and 1% glutamine (Invitrogen) at 37°C and 5% CO2. Immediately, after plating, 100 ng/mL CCN1 was added to the media. Controls remained untreated. After 5 days, cells were photographed by light microscopy (eclipse TE300; Nikon).
In vitro seeding of CCN1-coated dEAC
2 cm×2-cm pieces of noncoated and CCN1-coated dEAC (n=3 each) were fixed to six-well cell culture dishes by a fibrin-based glue (tissucol-kit 1,0 immuno; Baxter) according to the manufacturer's instructions and covered with the RPMI medium containing 10% fetal calf serum (Biochrom) and 1% glutamine (Invitrogen). L-929 mouse fibroblasts were seeded to each well (1.7×105 cells) and incubated at 37°C and 5% CO2 for 5 days. Before epifluorescent microscopy, cells were loaded with 1 mg/mL calcein-acetoxymethyl ester (Invitrogen) and living cells growing on the matrices were visualized by green fluorescence (BX40 with AxioCam MRc camera; Olympus).
Cytotoxicity of CCN1-coated dEAC
1.5 cm2 of noncoated and CCN1-coated dEAC (n=3 each) were cut into equally sized pieces and extracted in 0.5 mL RPMI with 1% glutamine or vascular basal media (VBM; ATCC-LCG standards) for 72 h at 37°C under vigorous shaking. Media without EAC shaken under the same conditions served as controls. RPMI extracts were supplemented with 10% FCS and VBM extracts with the growth supplements provided by ATCC. About 100 μL of the extracts were added undiluted to L-929 fibroblasts (2500 cells/well) or HUVECs (ATCC-LCG standards; 5000 cells/well) seeded in 96-well plates the day before. After 24 h, 10 μL WST-8 (Promokine) was added to each well and absorption at 450 nm was measured after 2 h in a microtiter plate reader.
MMP-3 quantification in cell culture supernatants
1×1 cm-pieces of noncoated and CCN1-coated dEAC (n=3 each in triplicate) were fixed to 24-well cell culture dishes by a fibrin-based glue (tissucol-kit 1,0 immuno; Baxter) according to the manufacturer's instructions and covered with the RPMI medium containing 10% fetal calf serum (Biochrom) and 1% glutamine (Invitrogen). L-929 mouse fibroblasts were cultured for 24 h in RPMI with 0.5% bovine serum albumin before being seeded to each well (105 cells) in 500 μL RPMI with 0.5% albumin. Supernatants were collected after 24 h, lyophilized, reconstituted in 60 μL water, and analyzed for MMP-3 by a sandwich ELISA (Quantikine mouse MMP-3; R&D systems) according to the manufacturer's instructions.
Three-dimensional vascular tube formation assay
HUVECs (ATCC-LCG standards) were used from P3-P6 and grown with the vascular cell basal medium (ATCC-LCG standards), including one endothelial cell growth kit BBE (ATCC-LCG standards) per 500 mL and an additional 0.1% penicillin/streptomycin (PAA).
HUVECs were placed in 3D vascular tube formation assays as described recently 19 following a modified protocol of Saunders and colleagues.20,21 Briefly, HUVECs (5×106 cells/mL) were suspended in 1.75 mg/mL of rat tail collagen type I (4°C) (Invitrogen). dEAC were coated with varying concentrations of human recombinant CCN1 (0, 1, and 10 μg/mL) at 37°C for 24 h and cut into equally sized pieces. Pieces were then embedded into the tube formation assay (n=7–10). Afterward, gels were transferred to 37°C,where polymerization occurs and HUVECs together with matrices were allowed to form 3D networks in the presence of VEGF 165 (40 ng/mL) and FGF2 (40 ng/mL) (Peprotech) 22 in 96-well plates. The addition of a medium denoted the starting point of the assay. Cultures were monitored every 24 h for tube formation surrounding the embedded matrix and having direct contact. Toluidine blue staining of gels for visualization by light microscopy (eclipse TE300; Nikon) was performed as described previously. 21
Implantation protocol
All surgical procedures were carried out at the Animal Research Facility of the Hannover Medical School and according to human standard techniques. For in vivo testing, noncoated (n=3) and CCN1-coated dEAC (n=3) of 100-mm length and a diameter of 8–10 mm were implanted in eve Schwarzkopf sheep. ePTFE grafts (expanded polytetrafluoroethylene; GORE; n=6) were implanted in another 6 sheep and served as control. Grafts were inserted subcutaneously as right cervical AV fistulae, connecting the carotid artery and the external jugular vein. Anastomoses (ca. 2.5×larger than the graft diameter) were performed in the standard end-to-side running suture technique using polypropylene 6-0 (Ethicon). A single bolus of 100 U heparin per kg bodyweight was administered i.v. before clamping. Thereafter, no further anticoagulants were administered. All animal experiments were performed under general anesthesia (induced with 30 mg per kg ketamine, and maintained with an i.v. bolus injection of 2 mg/kg propofol). For pain control, animals received 4 mg/kg carprofen (Pfizer) intraoperatively and on days 1 and 3 thereafter, if necessary. For explantation, animals were sacrificed in deep anesthesia as described above followed by harvest of the grafts. The study protocol was approved by the local authorities (Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit). All sheep received care in compliance with the guide for the care and use of Laboratory Animals (Washington DC: National Academy Press; 1996 www.nap.edu/catalog/5140.html).
Observations
Animals were followed up for 14 weeks. Graft patency was evaluated clinically by auscultation, immediately after implantation, after 1, 2, 6, and 8 weeks and before explantation. Serum samples for antibody determination were taken before implantation, after 1, 2, and 6 weeks and at explantation after 14 weeks.
Examination of the explanted grafts
At explantation, grafts were harvested for histological and immunohistochemical evaluation.
Specimen processing and histopathology
The obtained grafts were dissected and tissue samples were taken from the arterial and venous anastomoses as well as from the middle part of the grafts (Fig. 1). One half of the samples were formaldehyde fixed, embedded in paraffin, and sections were stained by hematoxylin and eosin (H&E), Elastica von Gieson (EvG), or von Kossa (vKo). The other half of the samples were cryofixed.
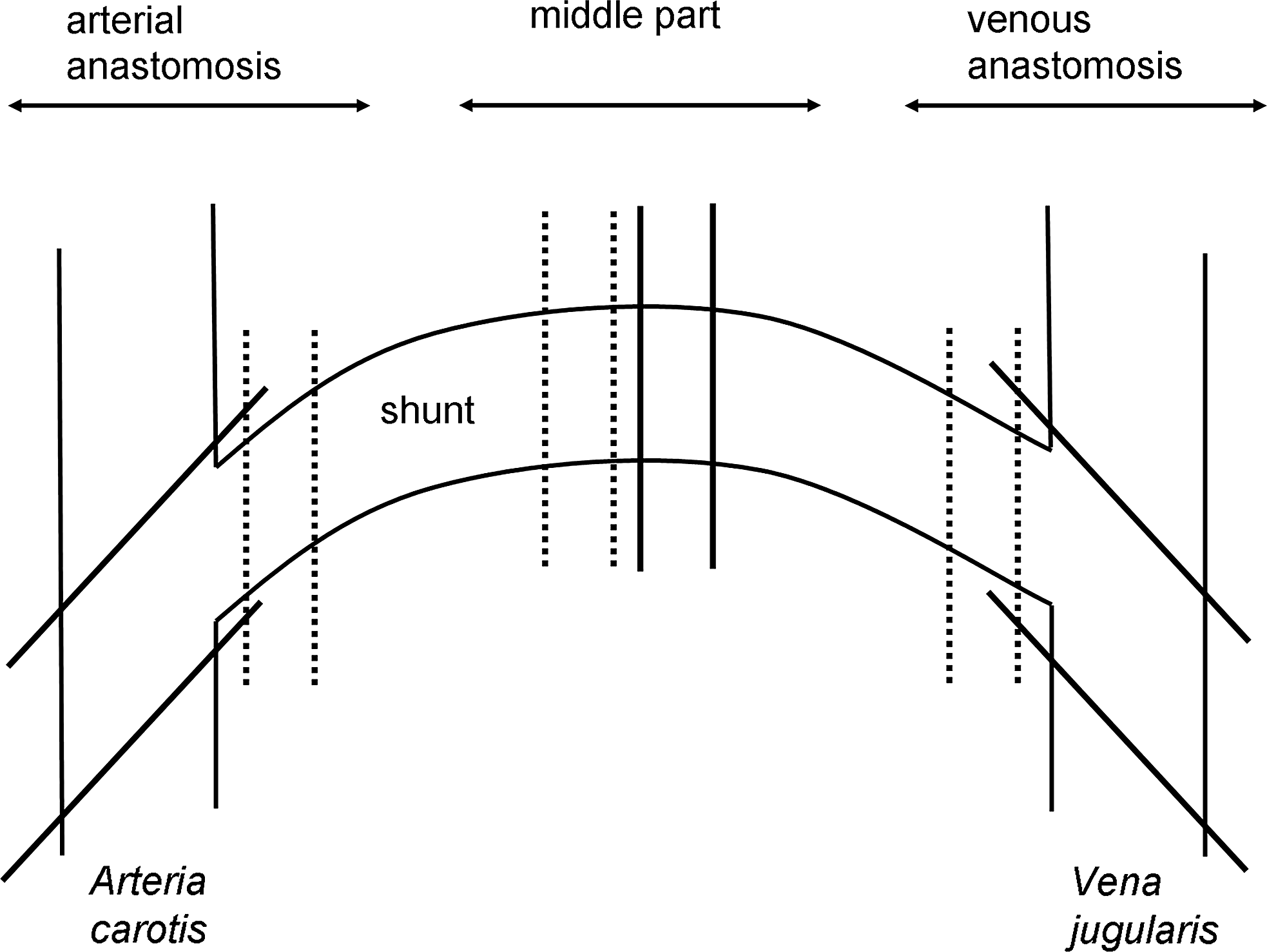
Dissection of specimen from the explanted grafts. After explantation, samples for histopathological evaluation (solid lines) and immunohistochemistry (dotted lines) were taken from the indicated regions of each graft.
Histological examinations were performed by a histopathologist in a blinded manner on H&E-stained sections. Typical histopathological changes in comparison to healthy, native carotid tissue were identified as fibrosis, leukocyte infiltration, endothelialization, calcification, neovascularization, and artherosclerosis and the severity of each deviation was evaluated based on a 4-step score system (0=no delimitable changes; 1=minimal; 2=moderate; 3=strong/severe), which was designed according to standardized histopathological approaches. EvG and vKo stainings were analyzed qualitatively.
Immunohistochemistry
Immunofluorescent staining was performed for the evaluation of endothelial and smooth muscle cell (SMC) reseeding in vivo by specific staining of cryosections with anti-CD31 (Santa Cruz biotechnology) and anti-alpha smooth muscle actin (aSMA) antibodies (Acris Antibodies GmbH). Briefly, cryosections were fixed with ice-cold acetone and blocked with 0.1% bovine serum albumin and 10% fetal calf serum in phosphate-buffered saline (PBS). The CD31 antibody was diluted 1:750 and the aSMA antibody was diluted 1:500 in the blocking buffer. Secondary antibodies (Alexa 555, Alexa 488; Invitrogen) were diluted 1:250 in the blocking buffer. Reseeding was assessed by confocal laser scanning microscopy (inverted-2, Leica) and quantified by counting specific signals on 6 slices from three regions (the middle part, venous, and arterial anastomosis) of each explanted graft. ECs were quantified by counting cells with green stained cell membranes forming the endothelial lining based on the number of DAPI-stained nuclei. SMC repopulation was evaluated by assessing specifically stained cells with red cell bodies indicating stained smooth muscle actin by a four-level score (0=no observations; 1=minimal repopulation; 2=moderate repopulation; 3=strong repopulation).
Detection of graft-specific antibody formation by Western blot analysis
Antibodies specific for dEAC were detected qualitatively by Western blot analysis using extracts of dEAC for blotting and sheep serum taken 6 weeks after implantation as the primary antibody. dEAC were homogenized and extracted as described previously. 23 Briefly, 600 mg matrix was submitted to bead mill homogenization and ultrasonification before extraction by 5% sodium dodecyl sulfate (SDS) and 5% sodium desoxycholate. The protein content was determined by Pierce BCA assay (Thermo fisher Scientific).
For Western blot analysis, 10, 20, and 30 μg of protein of EAC extracts were separated by 10% SDS-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes by tank blotting over night at 4°C. Membranes were probed by serum of implanted sheep taken 6 weeks after implantation in a dilution of 1:3000 and peroxidase-coupled anti-sheep secondary antibodies (1:10,000). Bound antibodies were visualized with ECL plus system (GE healthcare).
Relative quantitation of graft-specific antibodies by ELISA
For quantitation of graft-specific antibodies, enzyme-linked immunosorbent assays with extracted proteins from dEAC as solid-phase antigen were performed. dEAC were homogenized as described above by 1 mL of PBS. The protein content was determined by Pierce BCA assay (Thermo fisher Scientific).
Ninety six-well plates (Maxisorb C; Nunc-Thermo Fisher Scientific) were coated overnight at 4°C with 10 μg/mL aqueous extract of dEAC in a 0.1M carbonate/bicarbonate buffer at pH 9.6. After blocking, sheep serum diluted 1:1000 in PBS was incubated on the plates for 4 h at RT. After washing with PBS 0.05% Tween-20, a peroxidase-coupled secondary antibody was incubated (1:80.000 in PBS) for 1 h at RT. The TMB substrate (Biochrom) was added and stopped after color development with 2 M H2SO4. The optical density was measured at 450 nm within 20 min. As the positive control and internal standard, mouse plasma from immunized mice described previously 23 was incubated in a dilution of 1:1000 and detected by an anti-mouse peroxidase-coupled antibody. Absorptions from the sheep serum samples were normalized to the absorption received by the mouse serum to correct for interassay variations. ELISA was validated by calculating the inter- and intra-assay variability from four independent experiments using the serum taken at explantation from a noncoated dEAC-implanted sheep.
Peripheral blood mononuclear cells isolation and proliferation assay
At explantation, 2×9 mL of blood was taken in lithium-heparin monovettes (Sarstedt) from sheep, which received noncoated and CCN1-coated dEAC. Peripheral blood mononuclear cells (PBMCs) were isolated by standard density centrifugation using the Ficoll-Paque Plus separation solution (GE healthcare). Interface PBMCs were harvested, washed three times with PBS, and cryoconserved in RPMI media containing 1% glutamine, 10% ovine serum (PAA), and 10% DMSO (AppliChem). Before proliferation assay, PBMCs were thawed, resuspended in RPMI with 1% glutamine and 10% FCS, and rested at 37°C. After 24 h, cells were counted and seeded to round bottom 96-well plates in a concentration of 1×105 cells/well. After 24 h, cells were stimulated with aqueous extracts from dEAC prepared as described above in a final concentration of 10 μg/mL for 5 days. Cells were labeled with 10 nCi/well [methyl 3 H]-thymidin (Hartmann Analytic) for 24 h and stored at −20°C. Incorporated radioactivity was quantified using a beta-counter (Perkin-Elmer).
Statistics
Statistical calculations were done by Graphpad Prism 5.04 (GraphPad Software) statistic functions. Mean and standard error of the mean were calculated for all continuous values obtained in this study. Gaussian's normal distribution was tested by the d'Agostino&Pearsons omnibus normality test. Two group comparisons were performed by the Student's t-test for normal distributed data; more than two groups were analyzed by one-way ANOVA. For nonparametric data, a comparison between two groups was performed by the Mann–Whitney U test and between more than two groups by the Kruskal–Wallis test. Differences were considered significant at p<0.05. Significance levels were given as follows:*p<0.05; **p<0.01; ***p<0.001; #p<0.0001. An overview of the data obtained and statistical tests used is given in the Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/tea).
Results
In vitro seeding of L929 fibroblasts on CCN1-coated dEAC
As coating with CCN1 has not been described for decellularized equine arteries yet, first the maintenance of CCN1 on the matrix after the coating procedure and a possible cytotoxic effect on cells were tested. For this purpose, L929 fibroblasts were seeded onto CCN1- and noncoated matrices. In parallel, L929 on culture dishes were treated with CCN1, which was added to the media. L929 fibroblasts represent the cell line recommended by the European and International Standard for biological evaluation of medical devices (EN ISO 10993-5:2009: Tests for in vitro cytotoxicity) and to assess the in vitro biocompatibility of medical devices. Furthermore, Chen et al. 24 described morphological changes of fibroblasts by CCN1 easily to identify by light microscopy. Thus, fibroblasts seem to be a suitable tool to check both, the presence and functionality of CCN1 coating on the matrix as well as its biocompatibility. After 24 h of culturing, fibroblasts on CCN1-coated surfaces showed morphological changes toward spindle-like cell bodies with long extensions (Fig. 2B, D) on both, cell culture dishes and decellularized matrices. Cells on culture dishes or dEAC matrices without CCN1 (Fig. 2A, C) remained unchanged. Furthermore, viability of the cells was not compromised as no dead cells were observed and calcein staining was clearly visible indicating that calcein-AM was cleaved by intracellular esterases of metabolic active cells deliberating the fluorescent cleavage product. To confirm that CCN1 was functional and nontoxic when coated onto dEAC, two further assays were performed. As described in, 24 CCN1 induces the secretion of matrixmetalloproteases in fibroblasts. Indeed, a 2.3-fold concentration of MMP-3 was found in cell culture supernatants of L-929 cells, which have been cultivated on CCN1-coated matrices for 24 h (Fig. 3A). Moreover, extracts of CCN1-coated and noncoated matrices were incubated on the L-929 fibroblast before viability assessment by WST-8 assay. As observed before, 25 extracts of noncoated matrices slightly reduced viability of L-929 cells when compared to the media control. Extracts of CCN1-coated matrices, however, increased cell viability by 1.8-fold, which was highly significant (Fig. 3B). The same effect was observed when ECs were treated with the respective extracts. Thus, the coating of dEAC with CCN1 was sufficient and effective and led to nontoxic matrices for implantation.
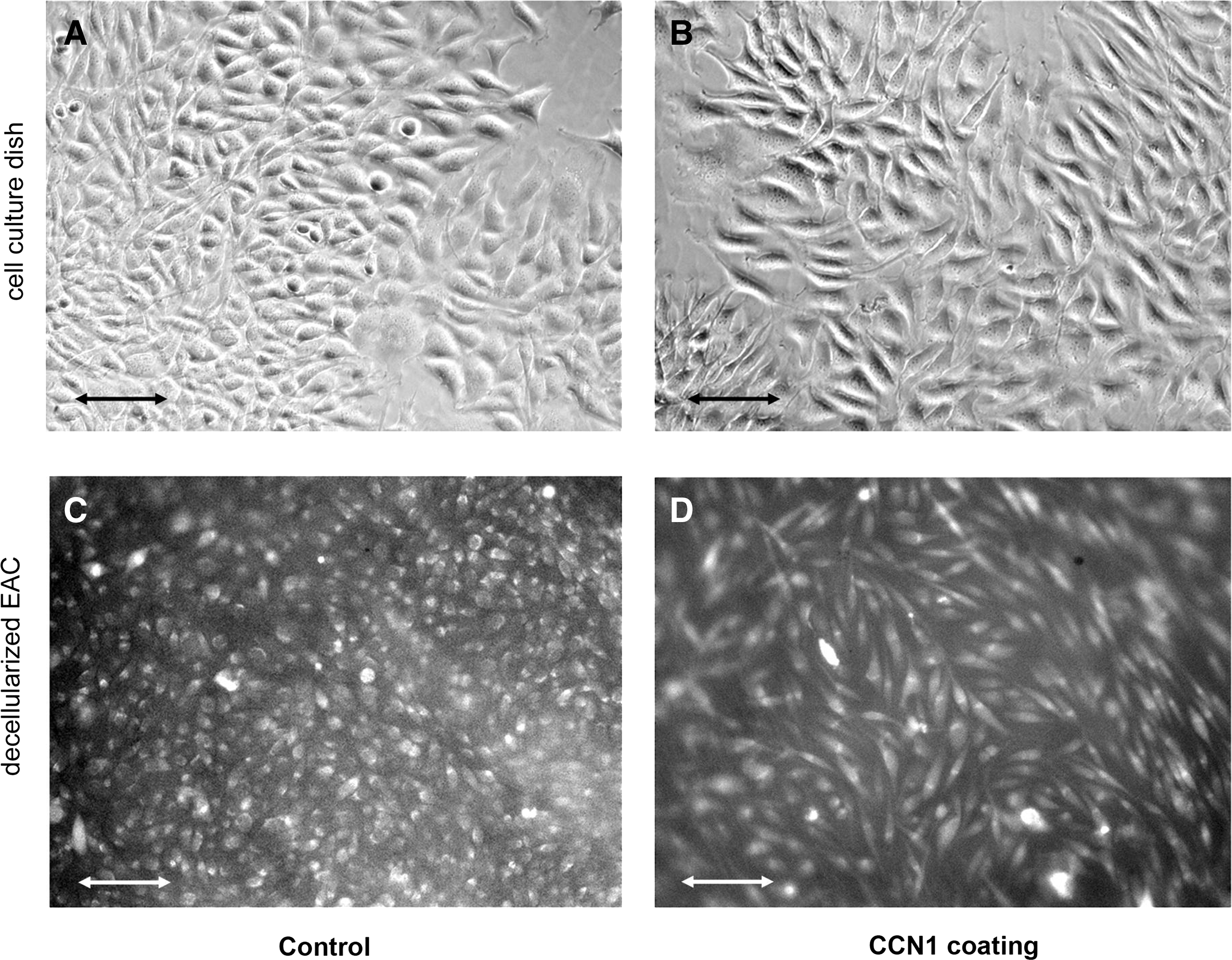
The effect of CCN1 on fibroblast morphology. L929 fibroblasts were cultured on cell culture dishes in the absence or presence of CCN1

Evaluation of functionality and cytotoxicity of CCN1 coating.
In vivo evaluation of alloplastic and bioartificial grafts with or without CCN1 coating
Patency
dEAC coated with or without CCN1 were implanted into sheep as a cervical arteriovenous shunt and followed up for 14 weeks. The control group received an alloplastic vascular prosthesis of ePTFE. During the follow-up, patency rates for the alloplastic material decreased steadily to 33.3% (Table 1), whereas the bioartificial grafts appeared to be patent throughout the observation period.
Patency was checked at the indicated time points by ausculation and after explantation macroscopically.
PTFE, polytetrafluoroethylene; dEAC, decellularized equine carotid arteries; n.d., not determined.
Cellular repopulation by immunohistochemistry
After explantation, the grafts were evaluated for their repopulation with ECs by CD31 staining (green) and SMCs by aSMA staining (red), respectively (Fig. 4A, B, D, E, G, H). The native ovine carotid artery showed ECs lining the luminal surface and SMCs forming typical stacks indicating the specificity of the antibodies chosen (Fig. 4A). PTFE shunts presented with strong formation of a neointima, justifying the poor patency rates described above (Fig. 4A, B). The neointima was partially endothelialized on the luminal face, but lacked completely SMC infiltration. In contrast, bioartificial dEAC grafts revealed the formation of a so-called neomedia infiltrated by both cell types resembling the tunica media of native arteries. In dEAC, without CCN1-coating, SMCs were scattered across the neomedia without clear organization (Fig. 4E) leaving some parts completely cell free (Fig. 4D). Also, the endothelial lining did not seem to cover the neomedia completely (Fig. 4E). When dEAC were coated with CCN1, the graft repopulation pattern differed considerably from that without coating. The formation of a neomedia was more pronounced as it showed SMCs stacked compactly in numerous layers (Fig. 4G, H). The neomedia was lined by a complete layer of ECs, (Fig. 4H) forming a border very similar to native arteries (Fig. 4A).
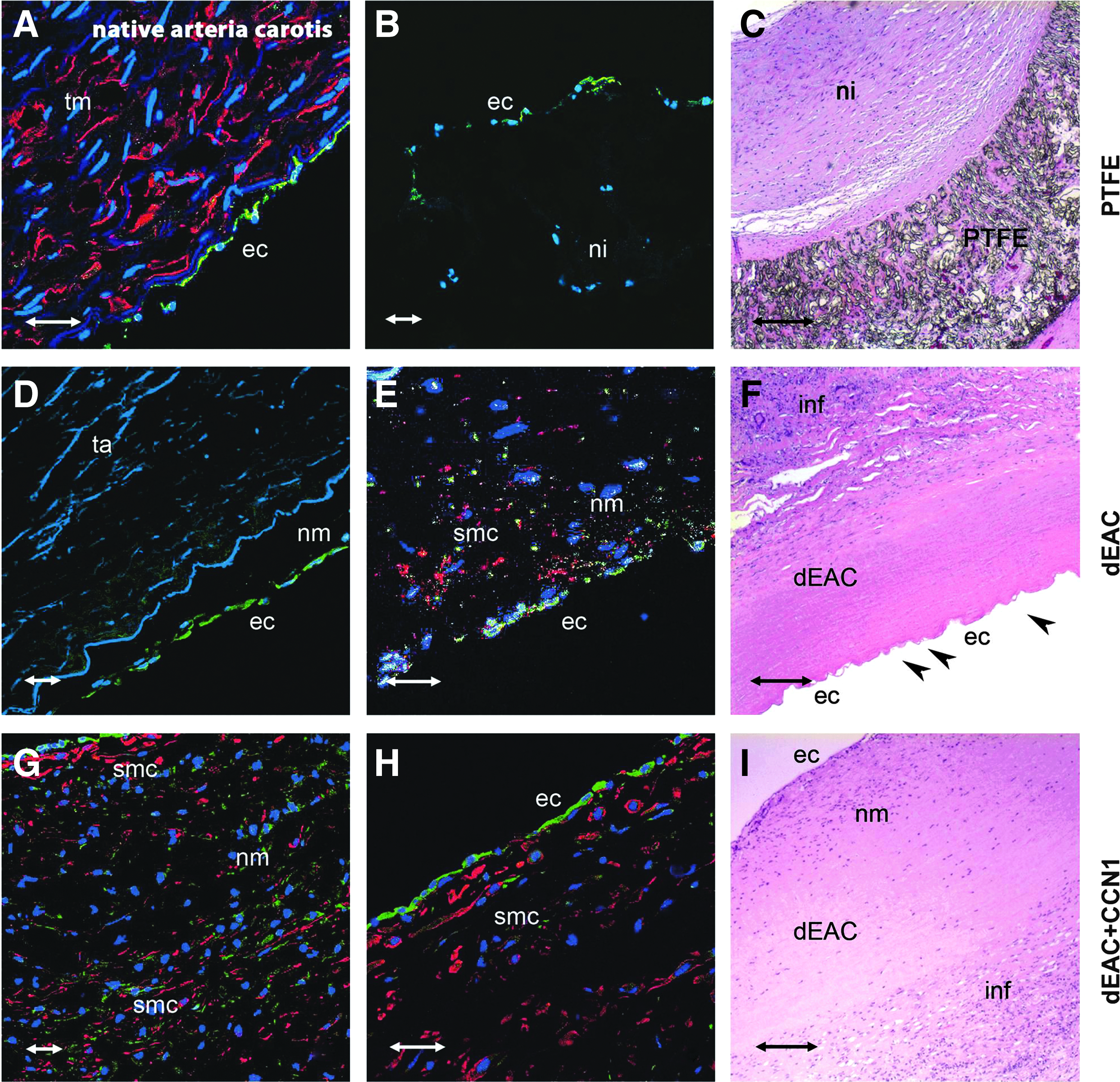
Cellular repopulation and histopathology of alloplastic (PTFE) and bioartificial (noncoated or CCN1-coated dEAC; dEAC and dEAC+CCN1) vascular grafts after implantation in sheep. After 14 weeks, grafts were explanted and specimens were taken according to Fig. 1. Cryosections were analyzed for reseeding with endothelial cells (ECs) by staining with the anti-CD31 antibody (green) and smooth muscle cells (SMCs) by staining with the anti-aSMA antibody (red), respectively. Nuclei were stained by DAPI. Positive control: ovine arteria carotis
Histopathology by H&E staining
In parallel to immunohistochemistry, histopathological parameters were assessed (Fig. 4C, F, J). H&E staining of the respective explanted grafts revealed the formation of a neointima on the luminal side of PTFE grafts, clearly separated from the alloplastic material (Fig. 4C). Both groups of bioartificial grafts showed the formation of a neomedia in close proximity to the fibers of the decellularized scaffold (Fig. 4F, J). Furthermore, in these grafts, infiltration by leukocytes (mainly macrophages and lymphocytes) was observed, which was moderate in the noncoated group (Fig. 4F) and seemed to be reduced in the CCN1-coated group (Fig. 4J).
Evaluation of endothelial and SMC repopulation
Matrix repopulation with cells staining positive for CD31 and aSMA was quantified from three regions as depicted in Figure 1 of each explanted graft (Fig. 5). PTFE grafts were endothelialized only in the middle part of the graft, whereas the anastomoses regions were not positive for CD31-specific signals. Noncoated dEAC showed an increase of endothelialization, with 1.5-fold for the middle part (not significant) and 5.3- and 6.1-fold for the anastomoses (p<0.0001). CCN1 coating, however, increased the number of ECs on the grafts luminal surface to 2.3-fold of PTFE in the middle part and 9.3- resp. 9.8-fold at the anastomoses, which was significantly more than in noncoated grafts (p<0.05 for the middle part and <0.01 for the anastomoses). Similarly, repopulation with aSMA-positive cells was higher in noncoated dEAC than in PTFE grafts (4.8-fold of PTFE in the middle part and 8.8- and 13.1-fold in the anastomoses, p<0.0001). As before, CCN1-coated grafts showed a higher repopulation with aSMA-positive cells (7.5-fold of PTFE repopulation for the middle part and 11.7- and 19.2-fold for the anastomoses), compared to noncoated grafts (p<0.05). Comparing the effect of CCN1 in the regions, endothelialization was increased by CCN1 by 1.86-, 1.51-, and 1.53-fold (arterial anastomosis, middle part, venous anastomosis) and SMC repopulation by 1.46-, 1.58-, and 1.32-fold, indicating an increase in repopulation not restricted to a certain region.
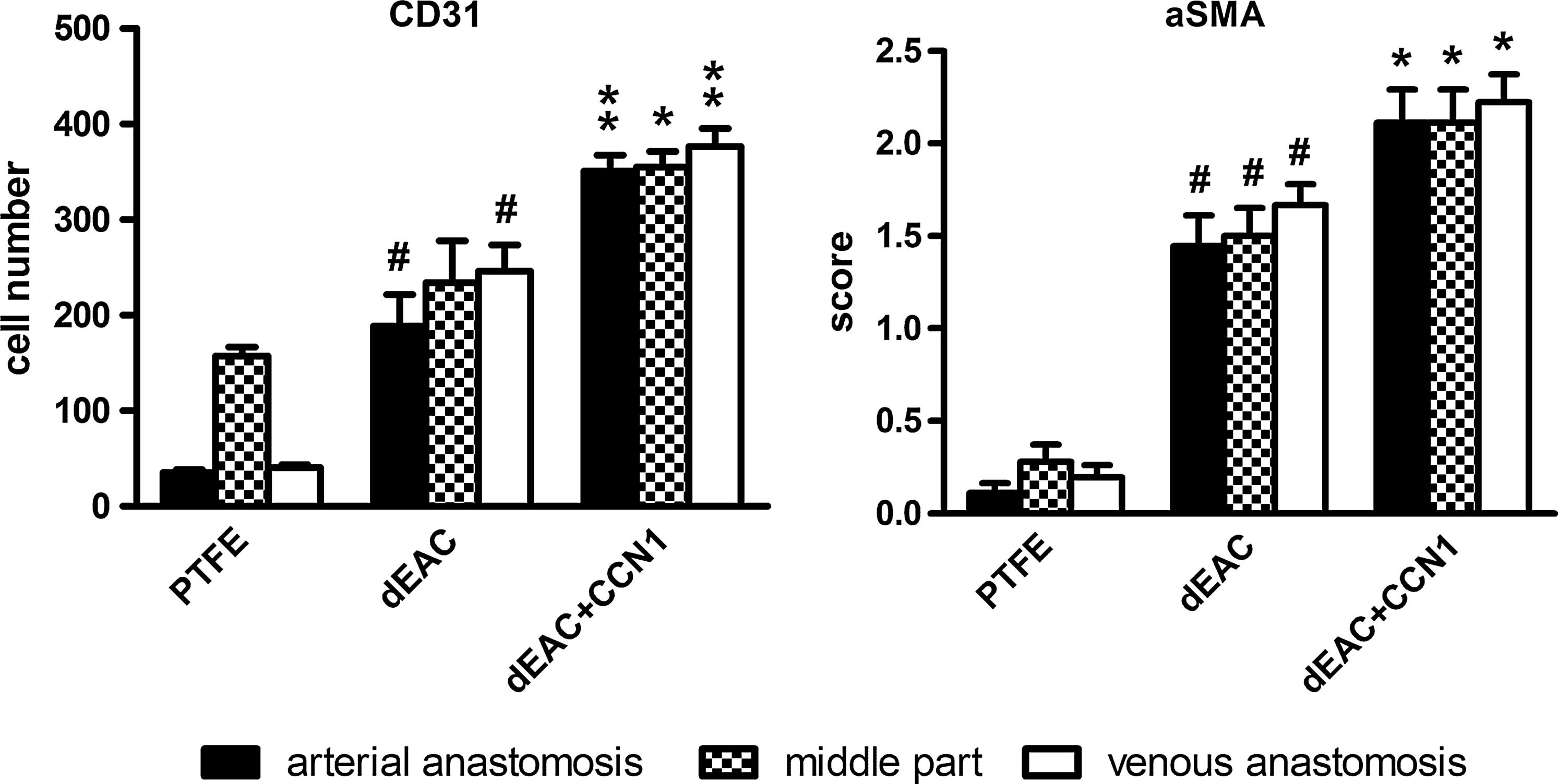
Evaluation of the repopulation of alloplastic (PTFE) and bioartifical (noncoated and CCN1-coated dEAC; dEAC and dEAC+CCN1) after implantation for 14 weeks in sheep. CD31-positive signals (endothelial cells) were counted and aSMA-positive signals (smooth muscle cells) were evaluated in a four-level score (0=no observation; 1=minor; 2=moderate; 3=strong) on cryosections from the indicated regions and expressed as means for each region. Shown are means±SEM of 18 cryosections from 3 regions of 6 (PTFE) or 3 (dEAC and dEAC+CCN1) animals. #p<0.0001 of dEAC versus the PTFE group, *p<0.05 and **p<0.01 of dEA+CCN1 versus dEAC calculated by the nonparametric Mann–Whitney U-Test.
Evaluation of histopathologic parameters on H&E, EvG, and vKo stainings
In addition to leukocyte infiltration (Fig. 4C, F, J), several other parameters were assessed semiquantitatively by a four-level score from the H&E stainings (Fig. 6). Leukocyte infiltration as an indicator for local inflammation was increased 4.6-fold in dEAC compared to PTFE grafts (p<0.001). CCN1 coating reduced this increase to only 1.7-fold of PTFE (p<0.01 vs. noncoated), indicating an anti-inflammatory effect. Likewise, CCN1 reduced fibrotic degeneration, as noncoated grafts showed 1.4-fold increased scores of PTFE (p<0.05), but CCN1-coated grafts a 0.8-fold decrease (p<0.01 vs. noncoated). This was confirmed by specific staining of elastin fibers by EvG staining (Fig. 7A, B). Noncoated grafts showed highly fractured elastin fibers and fibrotic areas indicating disruption of the internal microstructure, which was not present in CCN1-coated grafts. As described above, endothelialization was enhanced by CCN1 (1.77-fold higher than noncoated, p<0.05), which was, however, not significantly different from the endothelialization of PTFE grafts in this assessment. As PTFE endothelialization occurs predominantly on the neointima, it most likely does not prevent graft occlusion (see Table 1). In line with this, atherosclerotic deposits were reduced in bioartificial noncoated grafts by 0.64-fold (p<0.01) and even more pronounced in CCN1-coated grafts (0.34-fold, p<0.05 vs. noncoated) indicating the beneficial effects of CCN1 on atherosclerosis. Calcification, which was specifically stained by vKo staining (Fig. 7C, D) was as low as in PTFE grafts in bioartificial grafts regardless of their coating and also the neovascularization did not show any significant difference. As one hallmark of CCN1 is its enhancing effect on neovascularization, the tendency to reduce microvessel formation in CCN1-coated grafts prompted us to investigate this effect by immunohistochemistry and histology.

Evaluation of histological parameters of explanted alloplastic (PTFE) and bioartificial (noncoated and CCN1-coated dEAC; dEAC and dEAC+CCN1). Score 0=no observation; 1=minor; 2=moderate; 3=severe/strong. Shown are means±SEM of sections of 3 regions of 6 (PTFE) or 3 (dEAC and dEAC+CCN1) animals. *p<0.05, **p>0.01, and ***p<0.001 of the respective group versus the group left sided or as indicated, calculated by the Mann–Whitney U-Test for nonparametric values. n.s., not significant.

Elastica von Gieson
Effect of CCN1 on neovascularization
In near proximity to the grafts, numerous microvascular structures were found in noncoated dEAC, which seemed to be slightly reduced in CCN1-coated grafts. However, immunohistochemical staining showed that microvessels in noncoated dEAC appeared to be composed only of SMCs, whereas CCN1-coated dEAC showed capillaries containing both cell types (Fig. 8). This indicates a higher organized endothelialized capillary-like structure by CCN1. H&E staining confirmed this finding by indicating completely cellularized microvessels in CCN1-coated grafts.
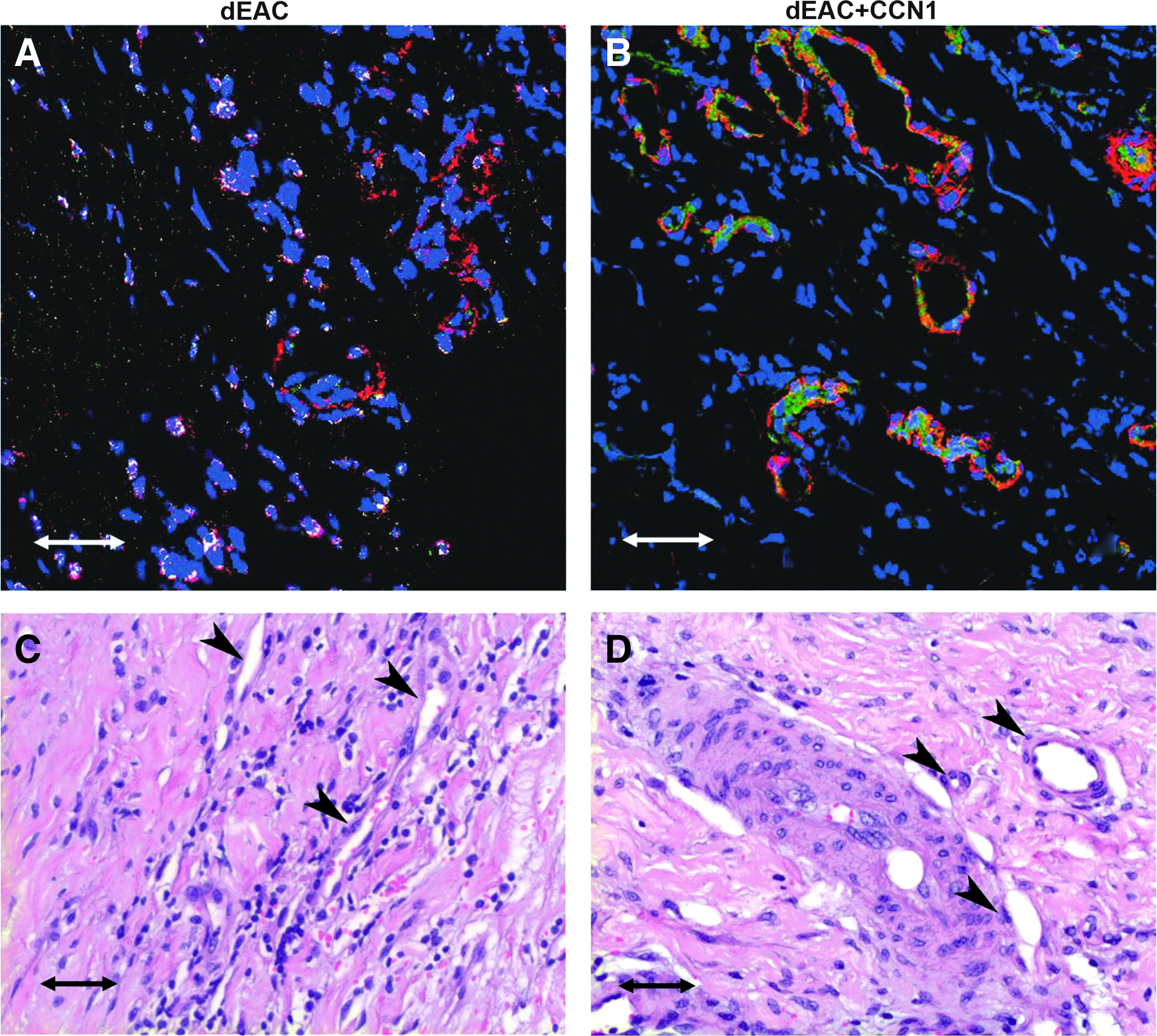
Effect of CCN1 coating on neovascularization of noncoated and CCN1-coated decellularized EAC (dEAC and dEAC+CCN1) after implantation in sheep. dEAC and dEAC+CCN1 were explanted after 14 weeks and evaluated for microvascularization in the tissue adjacent to the graft by hematoxylin-eosin staining
Taken together, these data show that CCN1 induced the formation of both, a structured neomedia and a highly organized vascularization and reduced local inflammatory reactions.
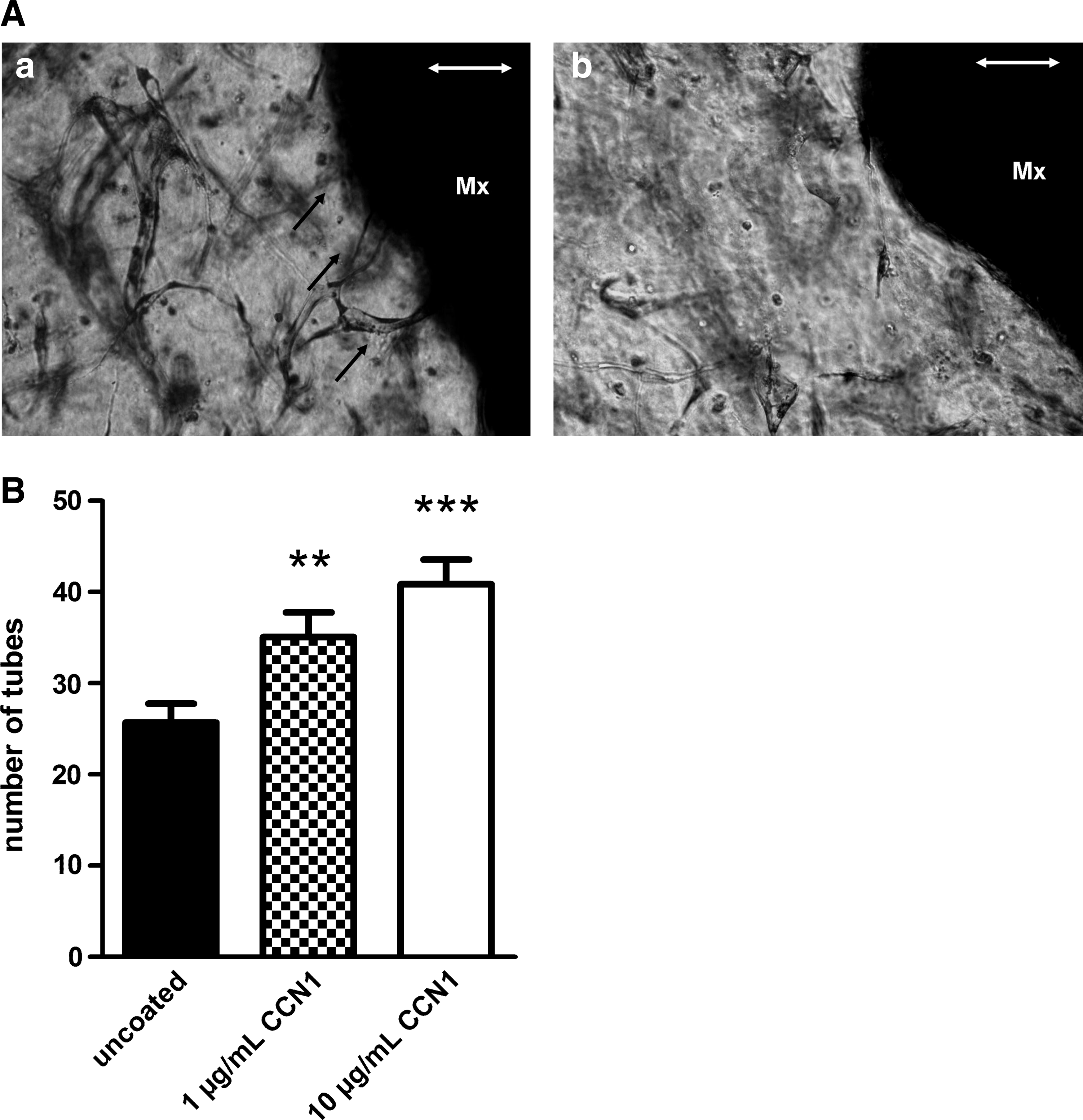
Three-dimensional tube formation assay
To confirm that CCN1 coating has a positive angiogenic effect, we performed a 3D tube formation assay, following a modified protocol by Davis.20,21 In addition to collagen and ECs, we included equally sized pieces of equine matrix coated with varying concentrations of CCN1. Tube formation was allowed to occur for 48 h (Fig. 9A). At this time point, tube formation was quantified by counting the tubes growing in direct apposition of and contact with the matrix. The number of tubes adjacent to the CCN1-coated matrices after 48 h was increased dose dependently for coating with 1 μg/mL CCN1 (1.3-fold) and 1.6-fold for the coating with 10 μg/mL CCN1 (p<0.01 and p<0.001; Fig. 9B) compared to the decellularized equine matrix without coating.
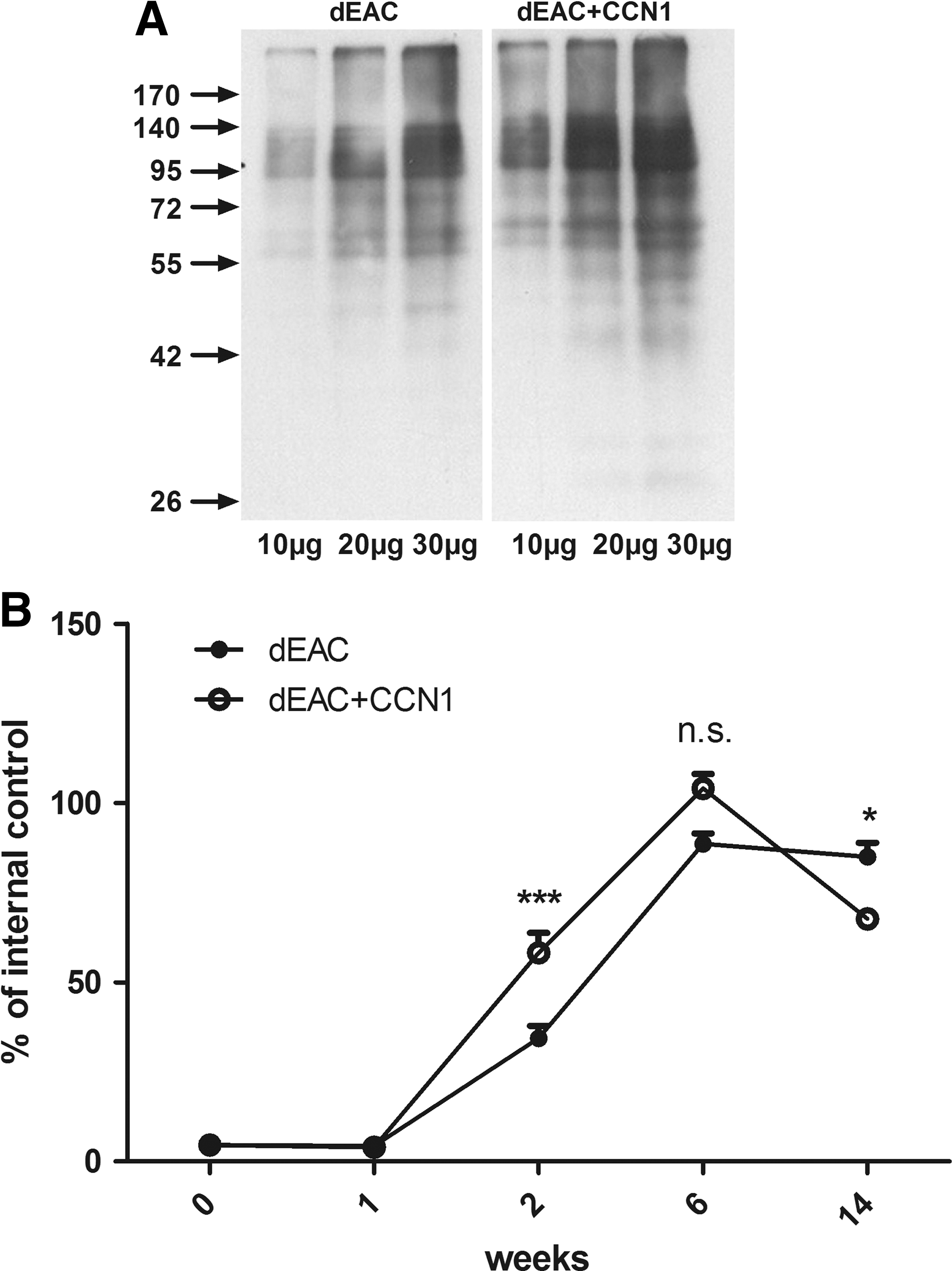
Graft-specific immune response in sheep implanted with noncoated or CCN1-coated dEAC
To elucidate the systemic immune response to implanted grafts, we investigated first the formation of graft-specific antibodies in the serum of sheep implanted with noncoated or CCN1-coated dEAC. Western blot analyses revealed strong staining of numerous bands in the range of 40 to 250 kDa (Fig. 10A). Staining was more intense in the CCN1-coated group, indicating a higher antibody titer. To quantify the antibody titer in serum taken at the respective time points, an ELISA was developed. ELISA plates were coated with extracts of dEAC as solid-phase antigen and probed with the sheep sera and anti-sheep peroxidase secondary antibodies. As the positive control and internal standard, serum obtained by immunization of mice with dEAC extracts was added. 23 The assay was validated using a sheep serum from week 14 after implantation of noncoated dEAC. Interassay variability evaluated in four experiments was 15.21% (coefficient of variation) and intra-assay variability of triplicates in four experiments was 1.49%±0.45%. These values indicated a robust and suitable assay. Quantification of dEAC-specific serum antibodies revealed a rapid increase starting after 2 weeks and a maximum after 6 weeks after implantation in either group (Fig. 10B). Antibody titers in the CCN1-group were 1.56-fold higher after 2 weeks (p<0.01) and 1.22-fold higher after 6 weeks (p<0.001). However, after 14 weeks, the antibody titer in the CCN1-treated group was decreased by 36.5% to a 0.8-fold lower level (p<0.01) than the titer of the noncoated group. Thus, CCN1 modulated the humoral immune response toward dEAC to higher levels at early time points and lower levels at delayed time points.
In a second approach, the cellular immune response toward noncoated or CCN1-coated dEAC was investigated. Peripheral mononuclear blood cells (PBMCs) were isolated from blood at the time of explantation and restimulated with extracts of dEAC prepared as described above. Restimulation resulted in an increase in proliferation assessed by 3 H thymidine incorporation indicating the reactivation of T-lymphocytes by the specific antigens (Fig. 11). In line with the reduced antibody titers after 14 weeks, the proliferation of PBMCs from sheep implanted with CCN1-coated dEAC was decreased by 26% compared to those from sheep implanted with noncoated matrices, which was highly significant, indicating that CCN1 coating attenuates both, the specific humoral and cellular immune response after 14 weeks.
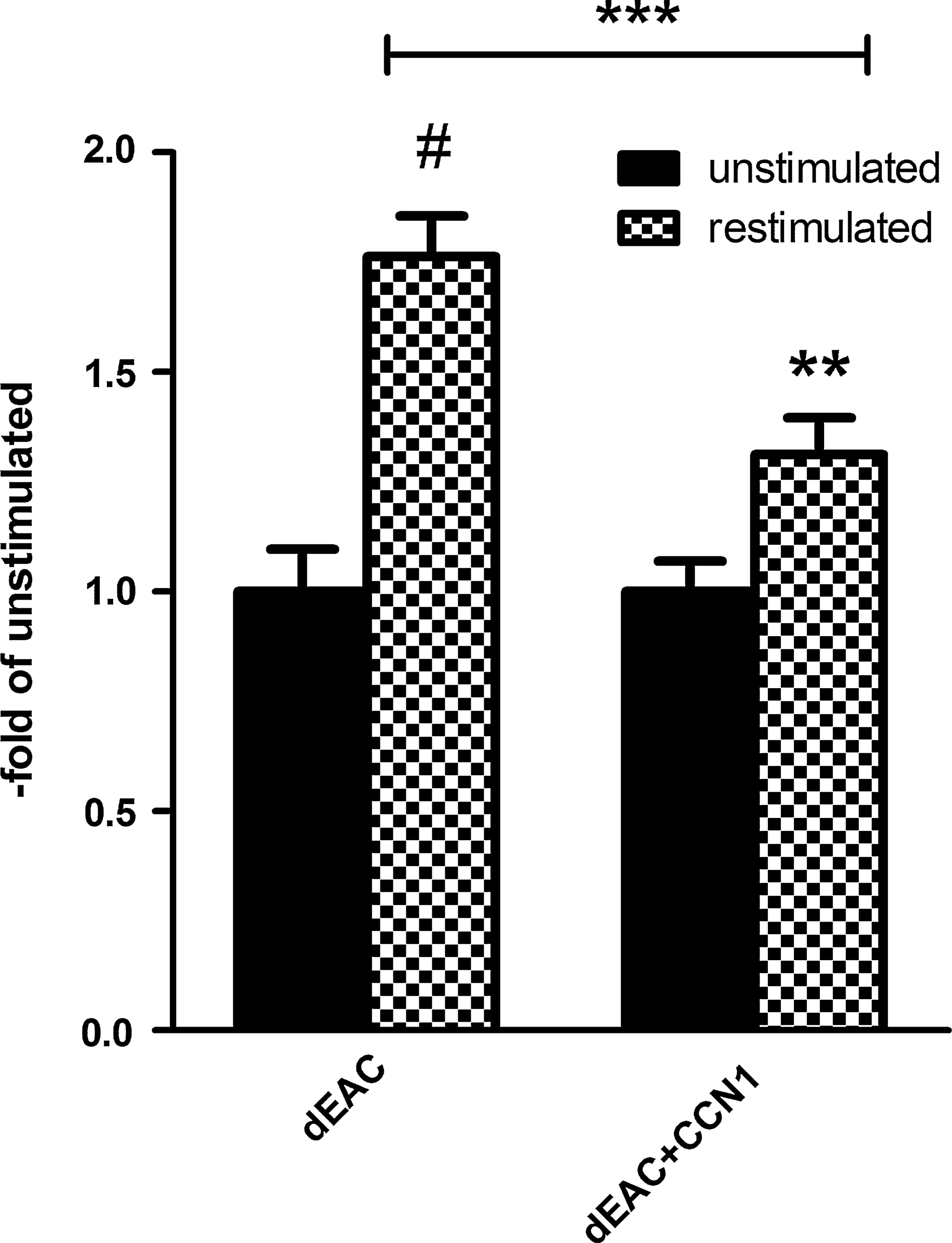
Cellular immune response to noncoated and CCN1-coated EAC. Peripheral blood mononuclear cells were isolated from sheep implanted with noncoated or CCN1-coated EAC after 14 weeks and restimulated with extracts of dEAC for 7 days. Proliferation was analyzed by 3H-thymidin incorporation for 24 h. Shown are means±SEM expressed as the fold increase of unstimulated PBMCs isolated from three animals and analyzed in 30 wells for each stimulus. **p<0.01; ***p<0.001; #p<0.0001 by the Mann–Whitney U-test for nonparametric values.
Discussion
Remodeling and integration of decellularized scaffolds is a fundamental process in regenerative medicine. Reconstruction/regeneration starts with the infiltration of autologous cells, includes the proteolytical breakdown of the matrix and the production of the autologous extracellular matrix, and finally, results in an autologization of the graft over time. 5
Spontaneous reseeding of different acellular scaffolds, however, seems to be dependent on the origin of the tissue (allogeneous versus xenogenous), of its structure, and its anatomical location within the body. Whereas human decellularized heart valves showed repopulation and adaptive growth in humans, 26 xenogenous heart valves in few cases did not. They were rejected and lacked endothelialization.27,28 In contrast, thin sheet-like structures like decellularized porcine small intestinal submucosa 29 or dermal scaffolds 30 were completely infiltrated by fibroblasts and remodeled after several weeks.
The more compact wall structure of vascular grafts and the shear stress of the blood stream, however, may account for the poor repopulation of bioartificial arterial vascular grafts as observed in the studies mentioned above.6–8 Our results with dEAC in a sheep model are in line with these findings. Although being patent throughout the observation period, and thus demonstrating a low thrombogenicity of the graft wall, we observed several shortcomings of this approach. The reseeding pattern for both, ECs and SMCs was not homogeneous after 14 weeks leaving parts of the graft cell free and thus, in ongoing contact to the blood stream. Moreover, a sustained leukocyte infiltration in the perivascular region was observed in combination with a foreign body reaction and fibrosis. Also, signs of atherosclerosis and a slight calcification were found completing the scenario of a mild, but sustained inflammatory reaction. Moreover, antibody formation was observed, which increased from week 2 to 6 and was sustained to the end of the observation period. However, the moderate local biocompatibility and immunogenicity of dEAC obtained by the protocol mentioned above was not completely unexpected. We could show previously, 23 that the process of decellularization does not eliminate extensively cellular proteins, but only depletes them to varying extents. Likewise, extractable proteins from decellularized matrices evoked a robust antibody formation in mice when used for immunization for 17 days. Thus, the local and systemic immune response observed after implantation in sheep could be a consequence of foreign body reactions, which will not cease as long as nonautologized parts of the graft surface are exposed to the surrounding tissue (a comprehensive review of foreign body reactions to biomaterials is given in 31 ). These observations prompted us to improve matrices by a factor, which has been shown previously to support in vitro endothelialization of a decellularized matrix. 15 Here we could show for the first time that CCN1 was capable for this purpose in vivo. Beyond that, we demonstrate that the beneficial effects of CCN1 are far more widespread than just serving as an anchor for the attachment of ECs.
CCN1-coated grafts showed not only a completely closed endothelial lining, but also were infiltrated by SMCs, showed less severe local inflammatory responses, less fibrosis, less atherosclerosis, and a higher organized neovascularization.
The functions and signaling pathways of the CCN1 protein are numerous, including direct interaction by binding to integrins as well as indirect transcriptional activities when localized in the nucleus.14,32 Here, some of these actions, which may be responsible for the observed effects will be discussed, although the true scenario might be far more complex.
One of the first effects described for CCN1 is its angiogenic action. It has been shown that CCN1 induces the formation of new blood vessel structures in vitro and in vivo in the cornea. 33 The mechanism involved is the interaction of CCN1 with integrins αVβ3 and α6β1 on ECs leading to chemotaxis and proliferation, whereas binding to integrin α6β1 in interplay with heparan sulfate proteoglycans induces adhesion and chemotaxis of vascular SMCs (reviewed in 16 ). Moreover, Grote et al. completed the picture by stressing the role of circulating CD34+ EPCs in angiogenesis and endothelial regeneration. 34 Soluble CCN1, which has been shown to be present in human plasma in that study, stimulates EPCs via integrin αVβ3 and αMβ2 signaling to secret an array of growth factors, chemokines, and matrix metalloproteinases, which enhance endothelial proliferation in a paracrine manner. CCN1 also enables transmigration of EPCs through endothelial layers contributing to angiogenesis. Coating dEAC with CCN1 most likely enables these mechanisms in terms of endothelialization and neovascularization. CCN1 leads to attachment of EPCs from the circulating blood to the graft wall and induces the secretion of multiple factors leading to proliferation of likewise attached ECs and SMCs. Moreover, EPCs may transmigrate into deeper regions of the graft where secreted factors again will induce proliferation and angiogenesis. As the coating procedure is not limited to the graft lumen, but also soaks the decellularized tissue, the effect of CCN1 thus, may not be restricted to the blood site. As neovascularization was observed in the perivascular region, the contribution to EPC resident in the vascular wall of neighbored blood vessels 35 could contribute to effects in this region. To prove that effects seen in vivo are linked to the proangiogenic effect of CCN1 and are not only due to an inflammatory host response, we used an in vitro approach to assess this question. Coated decellularized matrices with varying amounts of CCN1 were used in an in vitro 3D tube formation assay. Assessment of tubes adjacent to the matrix could significantly show the proangiogenic potency of CCN1, leading to the conclusion that effects seen in vivo represent results due to the coating with the compound itself.
In addition to the effects on endothelialization and neovascularization, CCN1-coated grafts showed reduced inflammation and fibrosis. There have been few reports linking the action of CCN1 to inflammatory processes, which at the first glance show contradictory results. On one hand, CCN1 was shown to induce a proinflammatory genetic program in murine macrophages in vitro, 36 on the other hand, it was able to suppress inflammatory reactions in a mouse model for experimental autoimmune myocarditis. 37 As the first effect involves the recruitment and activation of macrophages via αMβ2, it can be assumed that the CCN1 coating also adheres to macrophages and induces the secretion of proinflammatory cytokines. As this seems to be an acute effect (stimulation in 36 up to 24 h), the inflammatory reaction most likely is enhanced in the early phase after implantation. Roh et al. demonstrated that an immediate adherence of macrophages from the blood is the prerequisite for successful remodeling of a tissue-engineered vascular graft. 38 Thus, the proinflammatory early effect of CCN1 may initiate the reconstruction of the graft. Moreover, Rother et al. 37 investigated an anti-inflammatory long-term effect of CCN1 as it was administered one week before inducing the autoimmunogenic myocarditis. CCN1-treated mice showed a significantly reduced histological severity score, which was attributed to a reduced migratory potential of splenic immune cells from these mice. Thus, also in the sheep model, the long-term anti-inflammatory effects of CCN1 leading to reduced immune cell infiltration seem to support the switch from an acute inflammatory reaction to sustained tolerance toward the graft. In line with this, as a long-term effect at the end of the observation period, we also observed an attenuated immune response in sheep implanted with CCN1-coated grafts that covers both, the specific cellular and humoral immune response. T-Lymphocyte proliferation after a challenge with extracts of decellularized porcine tissue has been shown previously; 39 however, as the human PBMCs stimulated in that study had not been in contact with the matrix before, it was necessary to costimulate with an anti CD3 antibody to receive a response. In our study, PBMCs isolated from implanted sheep started to proliferate in the absence of a cofactor after being restimulated with the initial stimulus. Most likely, the cellular immune response to the matrices during the implantation induced the formation of specific T-cell clones, which start to proliferate when they are challenged with the antigens a second time. Interestingly, this response was attenuated in the CCN1 group, indicating an anti-immunogenic effect of CCN1 that explains partially the tolerance toward the graft. However, the antibody formation in the earlier time course seems to contradict these observations at the first glance as antibody titers in the CCN1 group were significantly higher up to 6 weeks and than dropped to levels below those of noncoated grafts. Similar effects in antibody formation to xenogeneic surfaces have been shown recently. 40 It was demonstrated that, in patients with porcine ligament transplantation, IgG antibody titer increased up to 6 months after transplantation and declined to pretransplantation levels within 24 months. This change has been explained by gradual remodeling of the xenograft leading to replacement by human tissue and thus, elimination of the xenoantigens. Due to the comparably short observation time, we were not able to see such an effect with the noncoated grafts. However, in the CCN1-coated group, the dropping antibody levels after 14 weeks may indicate an accelerated remodeling process, which is in line with the improved local biocompatibility and the attenuated cellular immune response at this time. It is tempting to speculate that the antibody titer also displays the switch from acute proinflammatory to delayed anti-inflammatory and remodeling effects of CCN1. The upregulated cytokines in macrophages include also IL-1ß stimulating antigen presentation by macrophages in an autocrine manner and IL-6 stimulating Th2 lymphocyte differentiation, which support B-cell activation. 36 Thus, antibody formation could be induced by CCN1 in the early phase. As the decline in antibody titer seems to depend on matrix remodeling, finally the effect of CCN1 on fibrosis and wound healing is highlighted.
The antifibrotic effect observed in the explanted grafts most likely is in coincidence with the observed matrix remodeling. In wound-healing processes, it was shown, that CCN1 by an integrin-mediated mechanism induced an antifibrotic senescence switch in myofibroblasts. This means the change from extracellular matrix protein production to the secretion of matrix metalloproteases and the downregulation of collagen I and TGFβ leading to matrix degradation. 41
Taken together, these results demonstrate that CCN1 coating can promote cellular repopulation and remodeling and may help to overcome the insufficient biocompatibility of dEAC. As the matrix itself is not changed, it is most reasonable to assume that these effects on the one hand are due to the described signaling effects of CCN1; on the other hand, the result of a rapid in vivo autologization protecting the body from direct contact with xenogeneic proteins. As far as one can judge, the effects of CCN1 coating are comparable with autologization of a graft or, in terms of neovascularization, even superior to it. However, a group by group comparison of modified grafts is desired to clarify this interesting question. Taking into account the tremendously higher effort of in vitro autologization, a comparable outcome of such a study would justify undoubtedly the use of CCN1-coated grafts for further development. Further studies will be initiated in the near future investigating the long-term effects of CCN1 coating and, thereby, evaluating the potential and suitability for clinical use of these grafts.
Conclusions
We could show that CCN1 not only facilitated attachment of ECs to the matrix, but induced beneficial effects to the graft such as formation of a neomedia containing SMC and EC lining, highly organized neovascularization, reduced local inflammatory reactions, and induction of immunologic tolerance. Thus, CCN1 coating improved remodeling and biocompatibility of decellularized xenogenous matrices significantly. As the coating procedure itself is extremely easy to handle, this matrix modification may represent a reasonable and effective tool for the generation of bioartificial matrices.
Footnotes
Acknowledgments
The work was funded by the “Innovation Fund of Lower Saxony, Germany: GMP model laboratory tissue engineering” and by the “Else Kroener-Fresenius-Stiftung,” Germany.
We thank Wiebke Baars and Reinhard Schwinzer for the quantification of the 3 H thymidin incorporation.
The excellent veterinary care carried out by Tanja Meyer, Carolina Theodoridis, Katharina Bobe and Christin Gebhard is gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.