
Abstract
Although bone morphogenic protein (BMP)-2 is known to potently induce osteogenic differentiation of human mesenchymal stem cells, strong individual differences have been reported. In part, this is due to internal antagonists of BMP-2 for example, noggin and chordin, secreted by differentiating cells. This enabling study was performed to prove the hypothesis that osteogenic effects of BMP-2 can be improved by transient nonviral gene silencing of chordin. We investigated the effect of siRNA against chordin on osteogenic differentiation in human adipose tissue-derived stromal cells (hASC). Cells of two different donors were isolated after liposuction and proliferated for passage 4 or 5. On seeding, hASCs were transfected with siRNA using a commercial liposomal transfection reagent. Subsequently, cells were differentiated in the presence or absence of BMP-2 (100 ng/mL). Noncoding siRNA as well as siRNA against noggin served as a control. Osteogenic differentiation of hASC was determined by alkaline phosphase (ALP) activity and matrix mineralization. ALP activity of hASC treated with siRNA against chordin was increased for cells of both donors. In contrast, silencing of noggin had no effect in any of the donors. In combination with BMP-2, silencing of either chordin or noggin showed strongly improved ALP activity compared with the control group that was also supplemented with BMP-2. Mineralization was observed to start earlier in groups that received siRNA against chordin or noggin and showed increased amounts of incorporated calcium on day 15 compared with the control groups. Silencing chordin in hASCs was successful to increase BMP-2 effects on osteogenic differentiation in both donors, while effects of noggin silencing were reliably observed only in one of the two investigated donors. In contrast to noggin silencing, chordin silencing also increased osteogenic differentiation without supplemented BMP-2.
Introduction
B
The reduced efficiency of BMP-2 in human has been connected to several natural antagonists of BMP-2, such as noggin, chordin, gremlin, and twisted gastrulation, which play a pivotal role in regulating the balance between bone formation and resorption.14,15 Chordin, noggin, gremlin, and twisted gastrulation are secreted glycoproteins that act as extracellular antagonists through binding BMP-2 and preventing it from interacting with its membrane receptors.16–20 Moreover, it was demonstrated that supplemented twisted gastrulation and chordin inhibit differentiation and mineralization in MC3T3-E1 cells. 21 There is a feedback mechanism between BMP-2 and its antagonist in healthy tissue. 22 For example, the induction of noggin by bone morphogenetic proteins limited the growth factors' activity in cultured rat osteoblasts 23 and C2C12 cells. 12 Hence, in order to improve the effects of BMP-2 and possibly reduce the necessary therapeutic dosage for bone formation, a local gene silencing might be a promising strategy.12,24–27
For noggin as the most frequently investigated BMP-2 antagonist, various gene silencing strategies have been shown to be successful. Among them, retroviral transfection of MC3T3-E1 cells and mouse-derived primary calvaria osteoblasts, lentiviral shRNA and minicycle siRNA transfection of human adipose tissue-derived stromal cells (hASC) have been shown to be effective in stimulation of bone formation. 27 Moreover, also nonviral liposomal transfection of C2C12 cells 12 and human bone marrow stromal cells (hMSCs) 28 and electroporation for in vivo transfection were successful. 12 Finally, a biomaterial-based approach showed that a copolymer of lactic acid, dioxane, and PEG can serve to deliver siRNA. 24 To the best of our knowledge, only Kwong et al. 29 showed effects of silencing chordin in human mesenchymal stem cells. That study was performed without external addition of BMP-2.
In our opinion, a possible therapeutic silencing strategy needs to be transient in order to allow for normal bone remodeling after the initial period of accelerated bone formation. Finally, viral transfection strategies may not find acceptance for therapeutic approaches. Hence, we, among others, think that a transient nonviral transfection of bone forming cells is a desirable way to improve bone formation induced by BMP-2.30,31
Adipose tissue-derived stromal cells (ASC) are recognized as a valuable source for in vitro studies on differentiation to several cell types of mesodermal origin and for cell-based regenerative approaches.27,32–35 The major advantage of these cells is their easy availability by liposuction and purification. Hence, the aim of this study was to investigate the hypothesis that transient gene silencing of chordin by siRNA can improve the effects of BMP-2 on osteogenic differentiation in a relevant human stem cell system, hASCs. To this end, we first investigated the transfection of hASC with a standard liposomal transfection reagent. Effects of silencing chordin and noggin, the latter as a positive control, were compared with a noncoding control with and without supplementation of BMP-2. As a readout, we analyzed alkaline phosphatase (ALP) activity and matrix mineralization.
Materials and Methods
Materials
The plasticware for all cell culture applications was obtained from Corning; DMEM (high glucose), PBS, and penicillin/streptomycin (P/S) were received from PAA; FBS was from Biochrome; and collagenase I was from Roche Diagnostics. The supplier for dexamethasone, ascorbic-2-phosphate, and 4-nitrophenyl phosphate was Sigma Aldrich. The supplier for β-glycerolphosphate, Trition X-100, papain, isopropanol and ethanol was Applichem. The reagents and materials for qPCR analyses were obtained from Applied Biosystems. The transfection materials and siRNAs were received from Qiagen or Riboxx as well as from Invitrogen/Life Technologies. The Quant-iT™ PicoGreen® dsDNA reagent was from Invitrogen/Life Technologies, the Calcium CPC kit was from Rolf Greiner BioChemica, the DNase was from Fermentas, GelRed™ was from Biotium, TRIsure was from Bioline, and the Phase Lock Gel™ tubes for RNA isolation was from 5 Prime.
Cell culture
hASCs were isolated from lipoaspirates via collagenase I digestion (Roche). Cells from a male (age of 40) and a female (age of 29) donor were obtained. Patients were informed by the physician and agreed, and none of the donors had any major medical comorbidities. Cells were cultivated until passage three in DMEM (PAA) containing 10% FBS (Biochrome) and 1% P/S (PAA); then, they were cryo-preserved in liquid nitrogen. For differentiation and transfection experiments, cells were thawed, proliferated again, and seeded in wells (Corning) in passage five at a density of 30,000 cells/12 well or 30,000 cells/48 well. The latter density was used in an experiment in which cells were seeded to confluence. One day after seeding, medium was changed to osteogenic medium (OM: 10 nM dexamethasone, 50 μM ascorbic-2-phosphate, Sigma Aldrich, and 10 mM β-glycerolphosphate, Applichem) or OM supplemented with 100 ng/mL BMP-2 (BOM). Medium in all cell culture experiments was changed every three days, and the cells were harvested at different times.
Recombinant production of BMP-2
Recombinant BMP-2 was produced at the Institute of Medical Physics and Biophysics, University of Leipzig, according to a previously described protocol. 36 In brief, Escherichia coli stain BL21 (DE3) containing a pUBS520 plasmid encoding BMP-2 was used to over-express the protein in inclusion bodies using a high-density cell fermentation in minimal medium. 37 Cells were lysed, and the inclusion bodies were separated by several centrifugation and washing steps and finally solubilized by 6 M guanidine hydrochloride in the presence of 200 mM DTT in Tris buffer at pH 8.5. DTT was removed by dialysis against 5 M guanidine hydrochloride buffer at pH 4.0 to induce protein refolding. Following the procedure described by 38 , the rapid dilution of the BMP-2 solution into degassed and prechilled arginine buffer (100 mM Tris; 1 M arginine; 5 mM EDTA, pH 8.3) also containing the redox shuffling system (100 μM GSH, 100 μM GSSG) induced refolding of BMP-2. The solution was stirred for approximately 10 days at 4°C. After concentrating the solution, the pH was adjusted to 5.5 and dialysed against urea buffer (100 mM Tris; 6 M urea; 5 mM EDTA; 75 mM acetic acid; and pH 6.0) at 4°C. Refolded dimeric BMP-2 was purified by heparin affinity chromatography according to the standard protocol. Finally, BMP-2 was dialysed against 10 mM ammonium acetate buffer (pH 4.8) at 4°C and lyophilized.
Testing of BMP-2 activity
Before BMP-2 was used for differentiation experiments with ASCs, its activity was tested using C2C12 cells.39,40 To this end, cells were seeded in 96 wells (30,000 cells/96 well) in DMEM containing 2% FBS and 1% P/S. The next day, the medium was changed and cells were induced with medium (2% FBS, 1% P/S) supplemented with different concentrations of BMP-2. A control group was cultivated in medium without BMP-2. Medium was changed every day. The cells were induced from day 1 (1 day after seeding) until day 4. On day 4, cells were harvested by aspirating the medium and by lysing them using a solution of Triton® X-100 (1% m/m) in PBS. This step was followed by measuring the ALP activity using 4-nitrophenyl phosphate (Sigma Aldrich) as a substrate and a plate reader (Infinite 200; Tecan) at 405 nm.
Transfection with siRNA
For a transient knockdown of chordin and noggin, hASCs were transfected with siRNAs (Riboxx) against these targets (Table 1) using Lipofectamine™ RNAiMAX (Invitrogen/Life Technologies) as a transfection reagent according to the manufacturer's protocol for forward transfection of cells. As a control, a nontargeting (noncoding) siRNA was used (Riboxx). On the day of transfection 30,000 cells were seeded in 12-well plates or 48-well plates. The cells were cultivated in 1 mL DMEM per well (high glucose, PAA) with 10% FBS (Biochrom) without P/S. Cells were transfected immediately after seeding. The final concentration of siRNA was 20 nM, which means that 1 μL siRNA was taken from a 20 μM stock solution for each well and combined with 2.5 μL Lipofectamine RNAiMAX and finally, diluted with medium to 1 mL per well. To silence chordin, three siRNAs were tested before, and the one leading to the most effective knockdown was chosen (data not shown). To compare the results with another target, noggin siRNA was used as described in literature. 41
Transfection efficiency and silencing efficiency
In order to determine the transfection and silencing efficiency of siRNA with Lipofectamine RNAiMax (Invitrogen) as transfection reagent, a fluorescently labeled, noncoding siRNA (AllStars Neg. siRNA AF 488; Qiagen) was used. To determine the effectiveness of transfection, cells were observed with a fluorescence microscope (Nikon, Eclipse TE 2000-S). Furthermore, cells were analyzed by flow cytometry (BD FACSCalibur™) and CLSM (Nikon, Eclipse TE 2000-U with D-Eclipse C1 488 and 543 nm) after transfection with fluorescently labeled siRNA. In addition, a Cell Death siRNA (AllStars Hs Cell Death siRNA; Qiagen), which leads to a knockdown of ubiquitously expressed genes resulting in cell detachment and death after 48–96 h that can be observed by light microscopy, was chosen.
Real-time PCR
For RNA isolation, cells were washed with PBS (PAA), homogenized in TRIsure (Bioline), and frozen at −80°C. After thawing, cells were transferred into Phase Lock Gel tubes (5 Prime), and isolation was performed using chloroform for phase separation, isopropanol for RNA precipitation, ethanol (75%) for washing (all solvents RNAse free; Applichem), and DEPC-water to desolve the RNA. RNA content was determined by absorption measurements at 260 and 280 nm. RNA integrity was verified by agarose gel electrophoresis and subsequent staining with GelRed (Biotium). Before reverse transcription, a DNase digestion was performed (Fermentas). Thereafter, cDNA was prepared using the High-Capacity cDNA Archive Kit (Applied Biosystems) and a thermal cycler (Eppendorf ).
For qPCR, TaqMan Gene Expression Assays for chordin (Hs00415315_m1) and noggin (Hs00271352_s1) as well as 18s RNA as endogenous control and the TaqMan® Universal PCR Master Mix, No AmpErase® UNG (Applied Biosystems) were used according to the manufacturer's instructions. The reaction was performed in 96-well optical reaction plates on a 7300 Real-Time PCR System (Applied Biosystems), and the relative quantification (RQ) values were calculated after the ΔΔct-method.
ALP activity
Cells were kept on ice and lysed by scraping them with a cell scraper in TRIS buffer (pH 8) followed by ultrasonic treatment. The ALP activity was measured using 4-nitrophenyl phosphate (Sigma Aldrich) as a substrate. The yellow product, 4-nitrophenol, was quantified at 405 nm using a plate reader (Infinite 200; Tecan). The resulting ALP activity was normalized to the number of cells for each well.
Cell number
For cell counting, first cells were scraped with a cell scraper (Corning) in PBS (PAA) followed by papain digestion (Applichem) in PBE buffer (pH=6.5) at 60 °C for 16 h. Then, the DNA content was quantified using Quant-iT PicoGreen dsDNA reagent (Invitrogen/Life Technologies). The measurement was carried out in a plate reader (Infinite 200; Tecan) at wavelengths of 485 nm (excitation) and 530 nm (emission).
Calcium content
Cells were scraped with a cell scraper in hydrochloric acid (0.5 N) followed by ultrasonic treatment and shaking over 12 h. The measurement was carried out using a colorimetric method (Calcium CPC, Rolf Greiner; BioChemica), resulting in a coloured complex formed by calcium and Cresolphthalein, which shows an absorption at 570 nm. This was detected with a UV-Vis spectrometer (Genesys).
Statistical analyses
All experiments were done in triplicate, and data were expressed as mean for each group combined with standard deviations. Statistical analysis was performed using a one-way analysis of variance with Tukey's all-group comparison test. The p-values were included in the figures.
Results
Transfection and silencing efficiency of ASCs with Lipofectamine RNAiMax
The transfection of hASCs with fluorescently labeled siRNA resulted in 97% fluorescent cells being detected by flow cytometry (Fig. 1A, B). CLSM analysis revealed green fluorescence that was indicative of labeled siRNA in the cytosol around the nucleus (Fig. 1C, D). To evaluate the silencing efficiency, a cell death siRNA was used. Figure 1E and F show hASCs 72 h after transfection. In the group that was transfected with cell death siRNA (Fig. 1E), most of the cells were detached; whereas cells treated with the noncoding siRNA (NC) showed an almost confluent cell layer with the typical morphology for hASCs (Fig. 1F). Hence, there were no visible toxic effects caused by the transfection procedure.
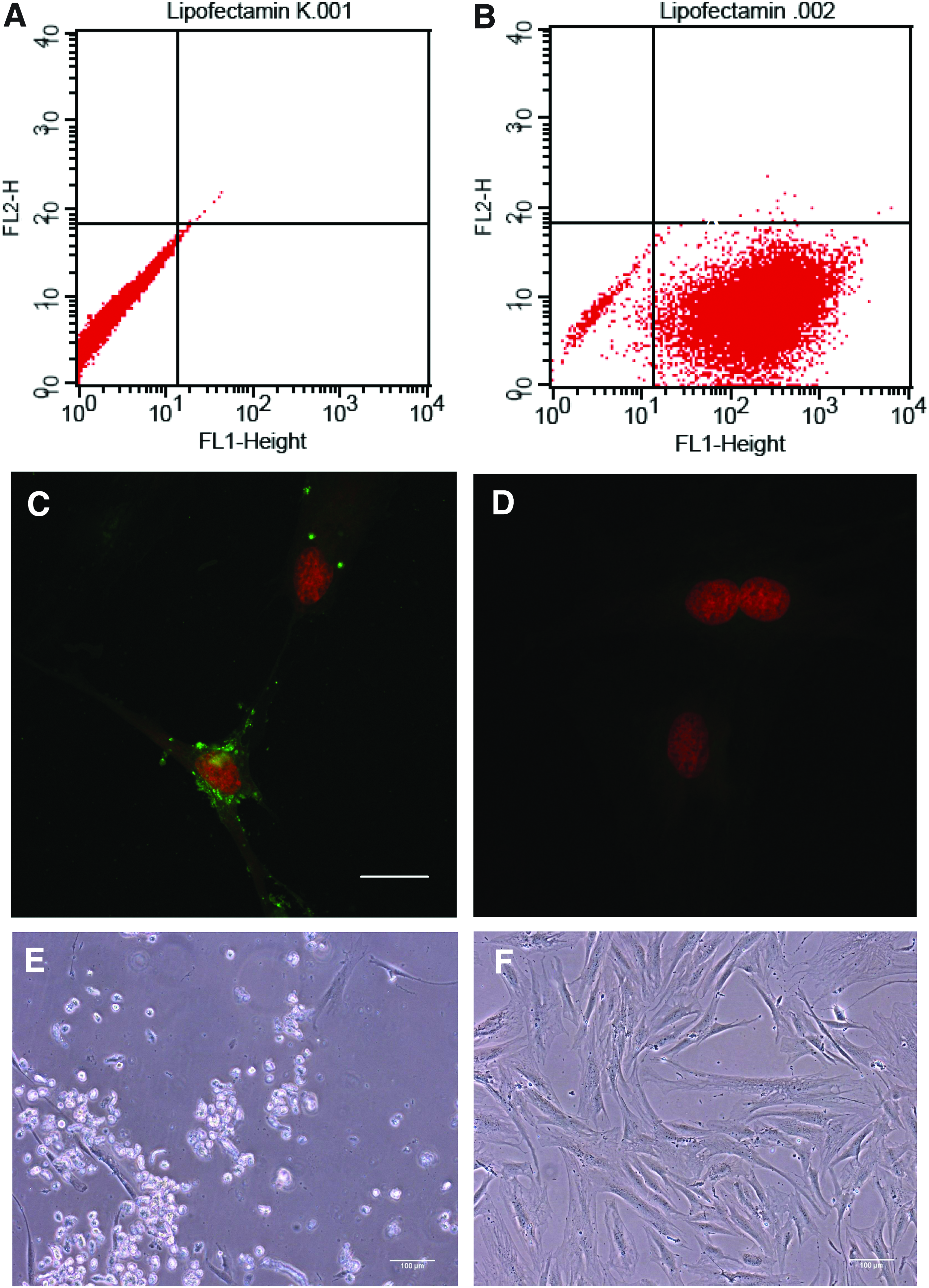
Transfection efficiency of hADSC with 20 nM green fluorescent siRNA (AF 488 nm) and Lipofectamine™ RNAiMAX;
Influence of BMP-2 on ALP activity and the expression of chordin and noggin during osteogenic differentiation of hASCs
Compared with OM without BMP-2, the addition of 100 ng/mL BMP-2 (BOM) increased the ALP activity significantly for both donors (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). The response to BMP-2 varied donor dependently and was higher for the male than for the female donor.
The expression of chordin and noggin mRNA was quantified using qPCR on day 4 of osteogenic differentiation (Fig. 2). The level of mRNA for noggin and chordin was strongly increased in groups that were supplemented with BMP-2 (BOM) during osteogenic differentiation compared with pure OM. Similar results were reported by Gazzerro et al. for cultured rat osteoblasts. 23 A profile of noggin and chordin expression over 10 days of culture, normalized to day 2 for another male donor, is shown in the supplemented data (Supplementary Fig. S2A). Here, we also see increased gene expression for chordin and noggin for the BMP-2 treated groups. For this donor, however, gene expression of chordin was stronger up-regulated by BMP-2 on days 2 and 4 than noggin. On day 7, noggin expression was stronger increased by BMP-2. On day 10, noggin as well as chordin mRNA expression was decreased compared with day 2.
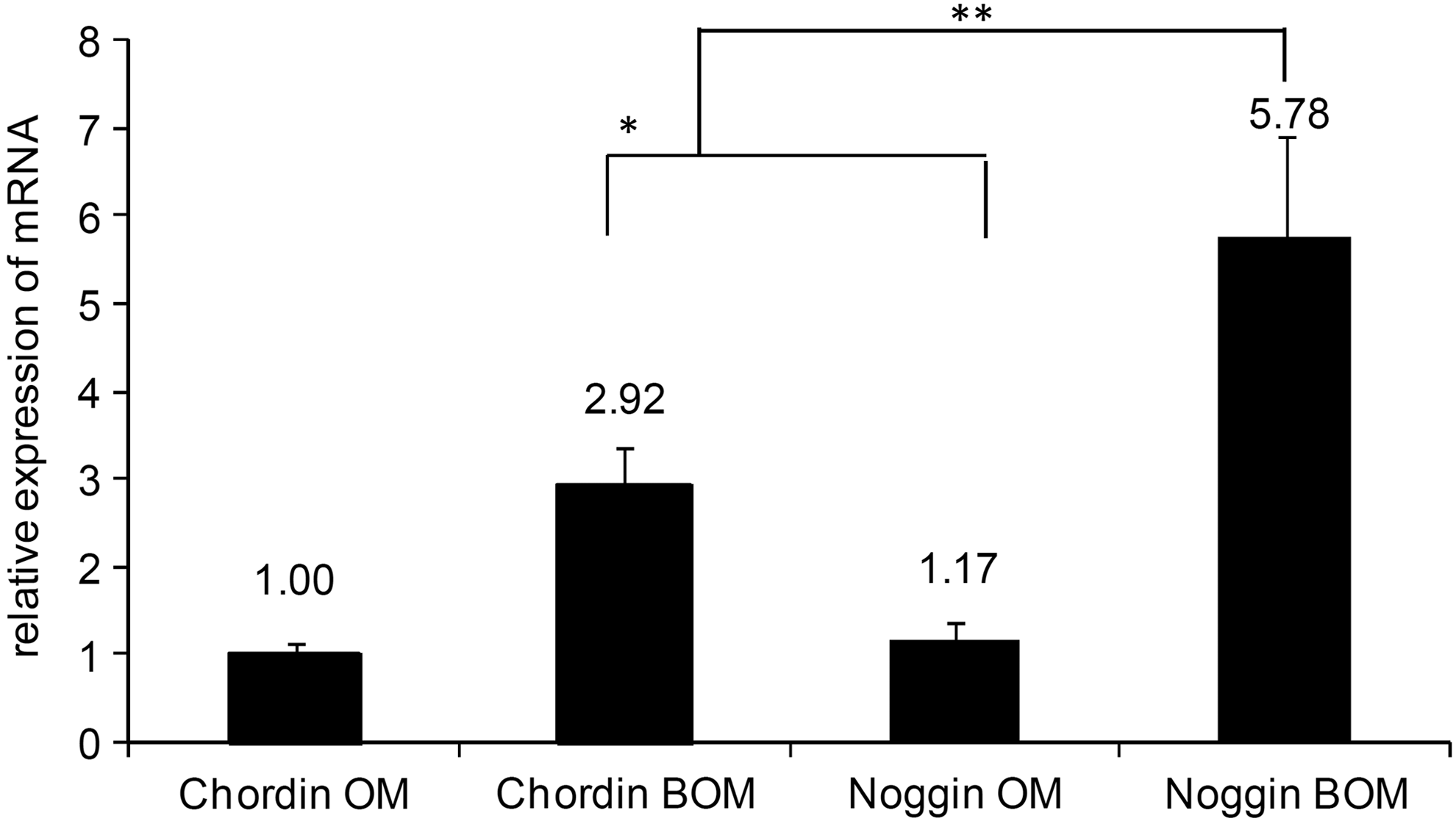
Expression of chordin and noggin mRNA in ASCs on day 4 during osteogenic differentiation; osteogenic medium (OM) compared with OM with additional BMP-2 supplementation (BOM, 100 ng/mL), *p<0.05, **p<0.01.
Knockdown of noggin and chordin
The knockdown of noggin and chordin was confirmed for both siRNAs in both osteogenic media (BOM, OM) on day 4 by qPCR (Fig. 3 and Supplementary Figs. S2B and S3A, B). Data were normalized and compared with noncoding control siRNA (NC). After transfection with coding siRNA, a significant decrease of mRNA for chordin and noggin was determined in all groups. Similar data of silencing were found by Chen et al. for noggin silencing using the same transfection kit. 28 On day 4, BMP-2-supplemented cells (BOM) treated with siRNA showed higher amounts of remaining mRNA for noggin and chordin than cells that had received OM. A profile of chordin mRNA expression over 10 days normalized to the untreated control on day 2 showed efficient silencing over 10 day in the presence and absence of BMP-2 (Supplementary Fig. S2B).

Real-time PCR after silencing of chordin and noggin mRNA on day 4 for the male donor in BOM and OM normalized to the respective noncoding control (NC), experiment performed in 12 wells, *p<0.05, **p<0.01.
Effects of noggin and chordin knockdown on osteogenic differentiation and cell proliferation
ALP as a typical early marker and calcium content as a late marker for osteogenic differentiation were determined. Silencing of chordin in cells cultured in OM medium led to increased ALP activities for both donors, while silencing of noggin had no significant effect compared with the noncoding control (Fig. 4A and Supplementary Fig. S4A). This was confirmed by qPCR, where we found an increased ALP mRNA level in the chordin silenced group compared with the corresponding control on day 4 (Supplementary Fig. S5). The combination of BMP-2 supplementation (BOM) and silencing of chordin resulted in a synergistic effect on ALP activity for both donors. Silencing of noggin, however, showed significant effects on ALP activity only for the male donor.
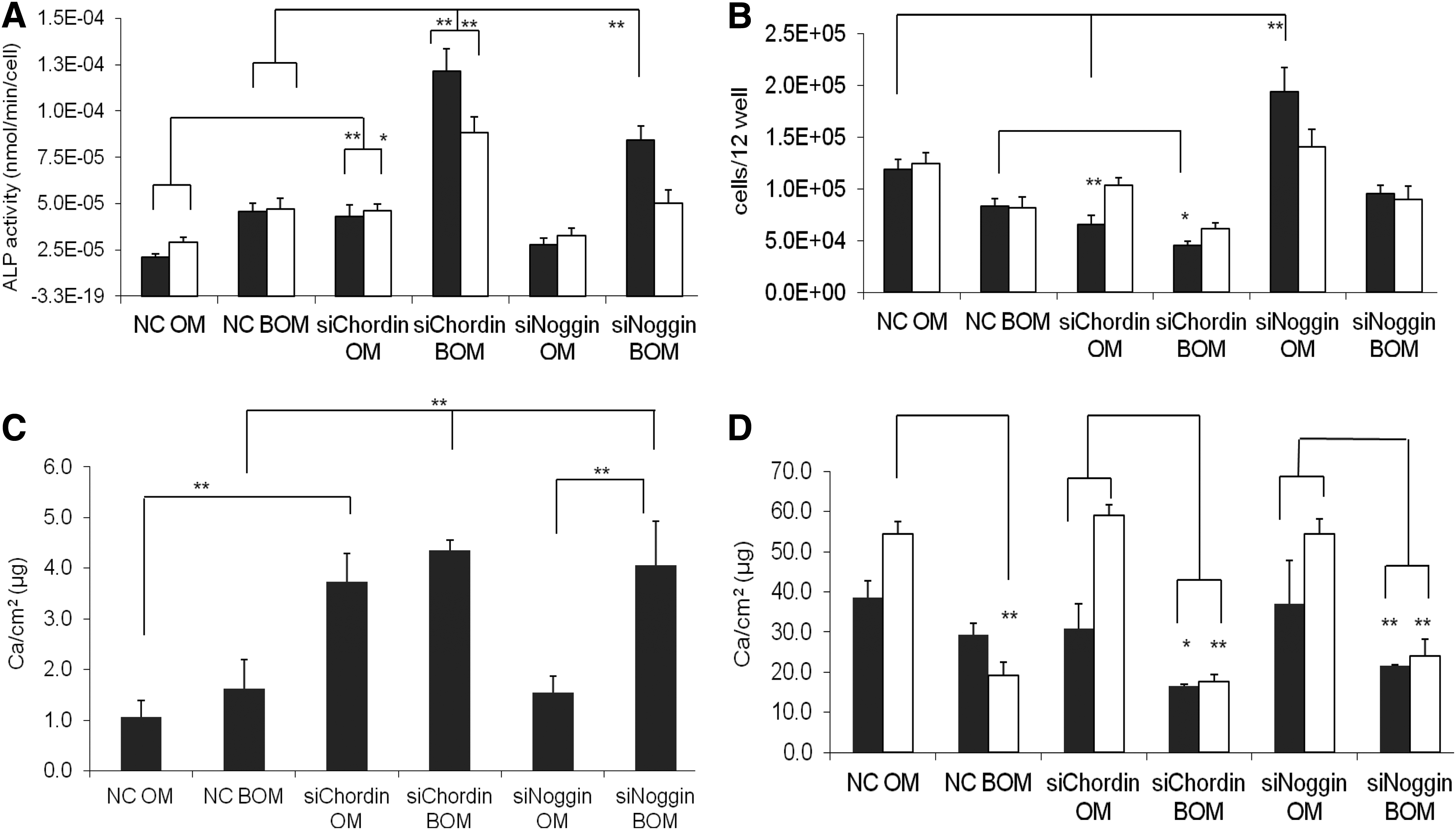
Effects of silencing chordin (siChordin) and noggin (siNoggin) on osteogenic differentiation in the presence (BOM) and absence of BMP-2 (OM) at a seeding density of 7900 cells/cm2 for a male (black columns) and a female (white columns) donor compared with the respective NC, *p<0.05, **p<0.01.
The supplementation of BMP-2 resulted in decreased cell numbers for both donors and in all groups (Fig. 4B). This effect was even stronger after silencing chordin. Silencing of chordin alone also reduced cell numbers significantly compared with the control. In contrast, positive effects on cell numbers were shown in one donor after silencing of noggin, while no significant effects on cell number were found in combination with BMP-2 compared with the corresponding control.
Mineralization was investigated on day 14 for the male donor (Fig. 4C) and on day 26 for both donors (Fig. 4D). On day 14, supplementation of BMP-2 led to a slight increase in mineralization for the male donor compared with the OM controls. Silencing of chordin in the presence and absence of BMP-2 as well as silencing of noggin in the presence of the growth factor, however, resulted in a strong early increase in mineralization relative to the other groups. Incorporated calcium amounts increased up to day 26 in all groups. In contrast to day 14, the groups cultivated in OM presented significantly higher amounts of calcium than the corresponding groups supplemented with BOM (Fig. 4D and Supplementary Fig. S4B). Moreover, no positive effect of chordin or noggin silencing was found on the late mineralization at day 26, neither with OM nor BOM. Furthermore, the chordin silenced group supplemented with BMP-2 showed the lowest calcium amounts for both donors.
Effects of seeding density on the osteogenic differentiation
In order to evaluate whether the observed lack in effectiveness of gene silencing noggin or chordin on mineralization was due to the influence of a long proliferation period and remarkable differences in proliferation between the experimental groups during the osteogenic induction, cells of both donors were seeded in a nearly confluent density with the intention to reduce proliferation and induce faster differentiation of the cells. To this end, we seeded the same cell number that had been used for the 12-well format in a smaller well format (48 well) whereas all other conditions were kept as in the 12 well-format. This resulted in a seeding density of 31,600 cells/cm2 (compare 12 wells: 7900 cells/cm2). Due to the higher cell density, we expected the cells to show only minor proliferation and faster differentiation. Hence, ALP activity was demonstrated for day 8 (Fig. 5A). ALP activity showed the same pattern for the silencing groups as was found for the lower cell density (Figs. 4A and 5A), but with increased activities compared with the first experiment, indicating improved osteogenic differentiation. No negative effect of BMP-2 on cell numbers was determined in cultures with increased seeding density (Fig. 5B) compared with the controls. In groups receiving siRNA against noggin, however, cell numbers were significantly increased compared with the corresponding controls. For the female donor, this effect was found after silencing chordin in OM as well. Similar to the experiment at low seeding density, an early onset of mineralization was found, but this time, mineralization for the male donor was stronger than in the first experiment (Fig. 5C). On day 15, calcium content of the matrix was significantly increased in both chordin silenced groups as well as in the group that received the combination of BMP-2 and siRNA against noggin. The highest amount of calcium was reached for both silenced groups in combination with BMP-2, which indicates a stronger and earlier differentiation and mineralization. Synergistic effects of gene silencing and BMP-2 supplementation were determined when chordin or noggin was silenced. A similar pattern was found for the female donor. However, in contrast to the male donor, the mineralization was still weak on day 15, resulting in low calcium amounts and high standard deviations for some groups. No significant effect was found for the combination of BMP-2 and noggin silencing for the female donor at this time. However, the synergistic effect for silencing of chordin in combination with the growth factor compared with the corresponding control (NC BOM) was traceable (Fig. 5D).
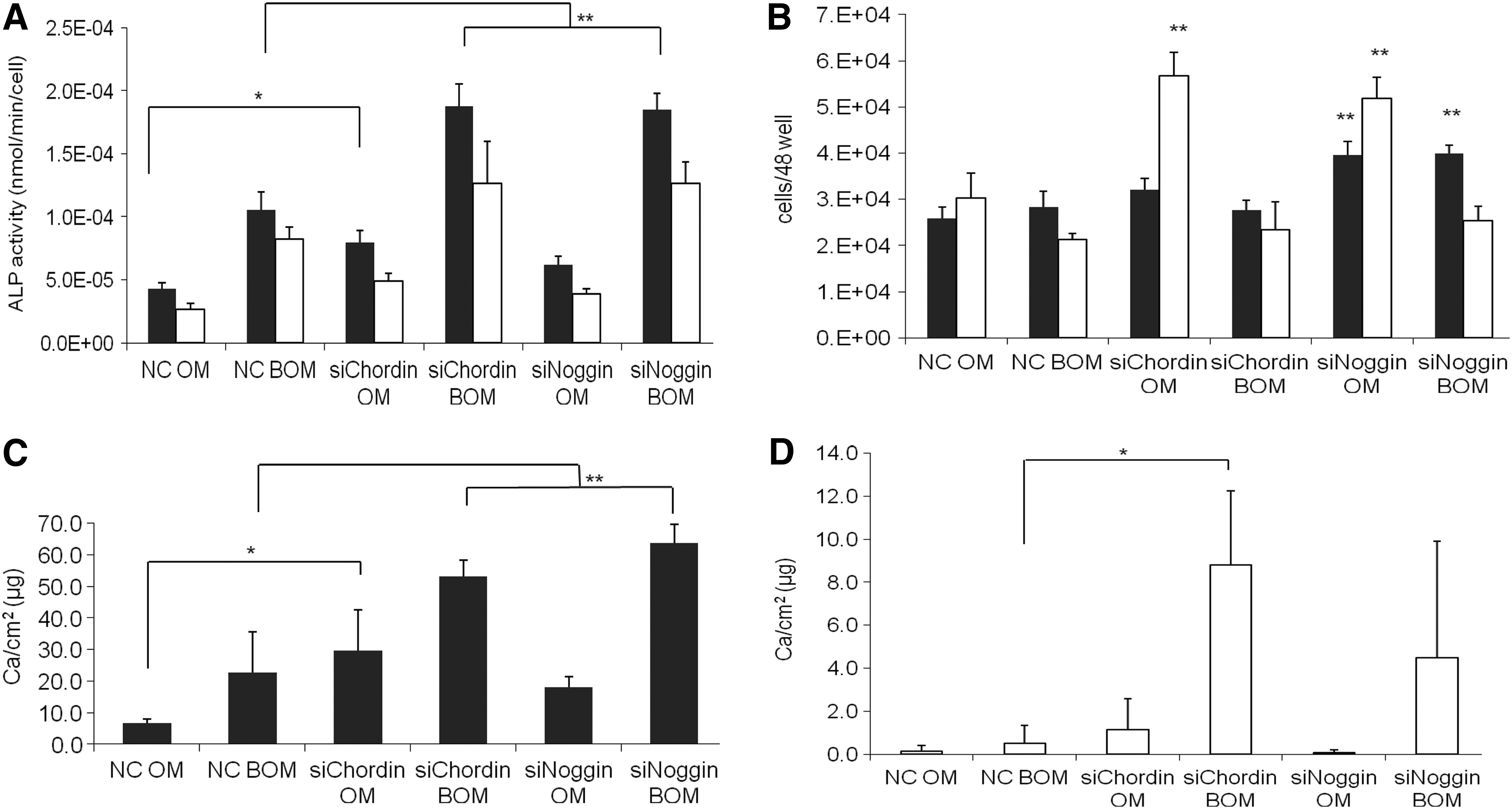
Effects of silencing chordin (siChordin) and noggin (siNoggin) on osteogenic differentiation in the presence (BOM) and absence of BMP-2 (OM) at a seeding density of 31,600 cells/cm2 for a male (black columns) and a female (white columns) donor compared with the respective NC, *p<0.05, **p<0.01.
Discussion
In this study, human ASC (hASCs) of two different donors, a man and a woman, were investigated to evaluate the effects of BMP-2 in combination with dexamethasone-containing OM. In agreement with other reports,6,7,42,43 we found increased expression of BMP-2 antagonists noggin and chordin in response to BMP-2 supplementation (Fig. 2 and Supplementary Fig. S2A). ALP activity as an early osteogenic marker also increased in response to BMP-2 compared with cells treated with OM alone. Differentiation of hASC seeded at a density of 7900 cells/cm2 resulted in decreased mineralization of BMP-2-supplemented cultures compared with wells treated with normal OM. After seeding the cells at confluent density, cell numbers at day 8 in the BMP-2-treated groups were shown to be as high as in the controls. Under these conditions, positive effects on mineralization were observed after supplementation with BMP-2 and OM. Our results on BMP-2 effects are in agreement with findings of other groups who found reduced mineralisation when nonconfluent cultures were treated with strongly differentiation inducing media.44,45 In addition, Hyzy et al. even reported an apoptotic effect of BMP-2 in human osteoblasts, while only minor effects were observed in noncommitted MSCs. 46
The aim of this study was to investigate the hypothesis that the effects of BMP-2 on osteogenic differentiation can be increased in human cells by concomitant silencing of the BMP-2 antagonist chordin and supplementation of BMP-2. The effects were compared with cells in which noggin, another BMP-antagonist, was silenced. To this end, cells were transfected with siRNA on seeding using a commercial liposomal transfection kit. One day after transfection, cells received OM with or without BMP-2. ALP activity and mineralization were compared with cells transfected with a noncoding commercial siRNA as well as with nontreated cells (Fig. 4A and Supplementary Fig. S4A). In absence of BMP-2, we found improved ALP activity by gene silencing of chordin for both donors, while none of these donors reacted to the silencing of noggin. In combination with 100 ng/mL BMP-2, we observed a highly significant increase in ALP activity relative to the BMP-2-treated control after silencing of chordin for both donors and in the male donor after silencing noggin.
In the absence of BMP-2, similar positive effects of chordin gene silencing on osteogenic differentiation of human MSCs have been reported before by Kwong et al. 29 This study showed improved ALP activity as well as mineralization for human bone marrow-derived mesenchymal stem cells. In contrast to chordin silencing, where we found only one published study, a lot of knowledge exists with regard to noggin silencing with and without BMP-2 supplementation in vitro and in vivo.
For example, Wan et al. described the positive effects of noggin siRNA on cbfa1, osteopontin, and osteocalcin after adenoviral transfection of MC3T3-E1 cells and primary osteoblasts in the absence of BMP-2. 26 Levi et al. also observed increased ALP by quantitative staining in the absence of BMP-2 on day 3 after lentiviral transfection of hASC with shRNA against noggin. 27 Chen et al., however, did not observe positive effects of siRNA against noggin in hMSCs in the presence of BMP-2. 28
Consistent with the increased ALP activity, we determined an early onset in matrix mineralization in cells transfected with siRNA against chordin with or without BMP-2 and in cells of the male donor treated with noggin siRNA and BMP-2. At later time points, however, when considerable mineral had been accumulated, silenced groups did not show improved matrix mineralization. In case of the male donor, cells treated with BMP-2 and siRNA against chordin or noggin showed even significantly reduced mineral content compared with the control.
To investigate whether these negative results on matrix mineralization were caused by the reduced cell number and nonconfluent culture in the chordin silenced and in the groups additionally treated with BMP-2, cells were seeded in a nearly confluent density for the following experiment. On day 8, no significant differences in cell numbers were found with the exception that some groups treated with chordin or noggin siRNA showed increased cell numbers relative to the control. ALP was determined already on day 8, because osteogenic differentiation is known to be faster under confluent conditions. 47 ALP normalized to the cell number in the chordin silenced groups was then increased compared with the chordin silenced groups in the first experiment. Again, the effects of the combination of BMP-2 and siRNA seemed to be synergistic. Considerable matrix mineralization was already found on day 15 for the male donor. The determined amounts of calcium on day 15 were about the same as in the best groups in the experiment with low seeding density on day 26 when normalized to the well surface area.
The increased cell seeding density probably had two positive effects: First, it prevented dilution of siRNA by initial proliferation of the transfected cells and second, by increasing the cell number to confluent numbers, any negative effects of the strongly differentiation inducing treatments on cell proliferation and confluent conditions as prerequisites for successful mineralization were neutralized.
In contrast to our data for the low seeding density, Kwong et al. showed increased mineral content after transfection of hMSCs with siRNA against chordin. 29 In agreement with us, this study showed a reduced proliferation of hMSCs after transfection with siRNA against chordin. Kwong et al., however, apparently used a 15-fold higher concentration of siRNA for transfection. Thus, dilution by proliferation is probably less severe than under the conditions applied in the present study. Other differences might be due to different cell sources.
In a study on combined effects of BMP-2 and noggin siRNA on hASC, Ramasubramanian et al. employed even higher cell densities (∼38,000 cells/cm2) than we did in the high cell density experiment. 25 In agreement with our results for the high seeding density, this study reported increased matrix mineralization by noggin silencing, but after 37 days of osteogenic differentiation. In contrast to our results, however, Ramasubramanian et al. did not observe positive effects on mineral content by a combination of siRNA and BMP-2. Contrasting results of noggin silencing were found by Chen et al. 28 This study showed decreased ALP activity on day 14 after noggin silencing in the presence of BMP-2 as well as decreased mineralization on day 28 using hMSCs from five donors. In our experiments, no decrease in ALP activity was observed in hASC after silencing noggin. With regard to the female donor, however, no positive effect of noggin silencing on ALP was determined in the presence of BMP-2 in the low seeding density experiments. In agreement with our data, Chen et al. found decreased mineralization after noggin silencing when cells were seeded in low densities and additionally showed reduced proliferation compared with the control on day 3 and 6. Possibly due to similar reasons as discussed by us, this might have caused a decreased amount of mineral.
Besides the mentioned effects after silencing chordin or noggin, the presented data revealed strong differences between the investigated donors with regard to ALP activity as well as mineralization (Fig. 4). They suit well into the current controversial discussion on BMP-2. Recent studies on human cells revealed heterogeneous results with regard to dosage and donor dependencies in response to BMP-2.6,8,9,11,48,49 Zuk et al. even doubted that the use of BMP-2 in combination with hASCs is justified at this point of time. 11
While the supplementation of BMP-2 in experiments of our study showed elevated ALP activity in hASC, the mineralization for the low seeding density was decreased compared with the cultivation in OM. Similar effects of BMP-2 were also observed by other authors. 48 Krattinger et al. compared the effects of BMP-2 on human primary fetal bone cells (hFBCs) and hMSCs. 45 Interestingly, after BMP-2 treatment in OM, they found an enhanced proliferation for hFBCs but not for hMSCs. Vice versa, hMSCs showed increased levels of ALP and other osteogenic markers. Nevertheless, the BMP-2 supplementation failed to increase matrix mineralization in both cell types on day 21 compared with OM without BMP-2.
Our observation that cell numbers were also slightly reduced after supplementation of BMP-2 is further in line with results of other studies.25,50 In addition, there are clear differences in the response to BMP-2 depending on the donor. 9 Besides heterogeneities in the differentiation of cells depending on individual variations, for example, co-morbidities, hormone states, depot site, the age or changes in cells, and tissue microenvironments,7,32,51–56 sex-specific heterogeneities during the osteogenic differentiation with stronger osteogenic potentials for men were reported in several studies.7,52,57–59 Levi et al. discussed differences in blood supply, cytokine signaling, or gene expression of subcutaneous locations and visceral fat depots, resulting in gender heterogeneities. 60 In contrast, Siddappa et al. found donor dependencies in response to osteogenic signals, but no correlation to age, gender, or source of isolation. 55 Ray et al. intensely discussed the impact of sex-specific hormones on several tissues/diseases 61 and gender differences. Polymorphisms in estrogen receptors and changes in their ratios or interactions might influence interindividual variability and gender differences.62–64 Moreover, for dexamethasone, also a steroid hormone, differences in receptor distribution were found to be influenced by gender and depending on tissue. 53
On the background of donor and possible gender-related effects, we have to point out that most of the data presented in this study are based on two donors. More work needs to be done to make clear statements on donor variability as well as gender and age effects on the effects of chordin and noggin silencing. In addition, cells were treated with a single dose of siRNA on seeding. For long-lasting effects of silencing, controlled release of siRNA might be a prerequisite for reliable effects of this strategy.
Overall, we focused on the interactions between BMP-2 and two antagonists, chordin and noggin, showing new insights in their effects on the osteogenic differentiation of human ASCs. To the best of our knowledge, this is the first study describing effects of concomitant BMP-2 supplementation and chordin silencing in human ASCs. Moreover, none of the studies directly compared the effects of noggin and chordin silencing in combination with BMP-2 in hASC.
Conclusion
Silencing chordin had a positive effect on osteogenic differentiation of hASCs. By co-treatment with BMP-2, ALP activity as an early marker of osteogenic differentiation was synergistically increased. With regard to mineralization as a late marker, we found an earlier onset for the groups treated with siRNA against chordin with and without supplementation of BMP-2 than for the controls treated with noncoding siRNA. The combined treatment of hASCs with BMP-2 and siRNA against noggin resulted in a similar effect as observed for the chordin group for one of the donors. Positive effects on mineralization, however, were only found when confluent cell density was chosen. The presented results rely on a single siRNA treatment of the cells during the seeding procedure. In an up-coming study, we plan to investigate the effects of a prolonged treatment with siRNA. In addition, alternative carriers for the transfection of cells, which are suitable for in vivo applications, are under investigation.
Footnotes
Acknowledgments
The authors thank Harald Kaisers (M.D.) and Karl Heinz Horak (M.D.), Praxisklinik am Markt (Leipzig, Germany) for providing lipoaspirates. Many thanks are also due to Daniel Huster (Institute of Medical Physics and Biophysics, University of Leipzig) for providing the authors with BMP-2. In addition, the authors thank Tina Biltz, Juliane Schieferbein, and Katharina Schwabe for excellent lab work supporting the generation of the presented data. Furthermore, they thank Anja Steude and Andrea Robitzki (Institute of Biochemistry, University of Leipzig) for their friendly help involving CLSM analyses. Finally, we would like to thank Achim Aigner (Rudolf-Boehm-Institute for Pharmacology and Toxicology) for valuable discussions.
This work was supported by grants from the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation, TRR 67, A1), the Saxonian State Ministry for Science and the Arts (SMWK), and the DAAD.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.