
Abstract
The basement membrane (BM) of the corneal epithelium presents biophysical cues in the form of topography and compliance that can impact the phenotype and behaviors of cells and their nuclei through modulation of cytoskeletal dynamics. In addition, it is also well known that the intrinsic biochemical attributes of BMs can modulate cell behaviors. In this study, the influence of the combination of exogenous coating of extracellular matrix proteins (ECM) (fibronectin-collagen [FNC]) with substratum topography was investigated on cytoskeletal architecture as well as alignment and migration of immortalized corneal epithelial cells. In the absence of FNC coating, a significantly greater percentage of cells aligned parallel with the long axis of the underlying anisotropically ordered topographic features; however, their ability to migrate was impaired. Additionally, changes in the surface area, elongation, and orientation of cytoskeletal elements were differentially influenced by the presence or absence of FNC. These results suggest that the effects of topographic cues on cells are modulated by the presence of surface-associated ECM proteins. These findings have relevance to experiments using cell cultureware with biomimetic biophysical attributes as well as the integration of biophysical cues in tissue-engineering strategies and the development of improved prosthetics.
Introduction
In vivo, ECM proteins such as laminin, collagen, and fibronectin (FN) are among the protein constituents that form the rich 3D topographic landscape with which cells intimately associate. Among the most important and widely studied ECM proteins/peptides that influence the cell–substratum interaction are FN, collagen (C), and the Arg-Gly-Asp (RGD)-containing peptides. Previous reports21–23 have demonstrated that surfaces modified with varying densities of FN or RGD facilitate wound healing because of increased cell migration. Spatially integrating ECM proteins on surfaces facilitates cell adhesion, morphology, and spreading. In addition, ECM peptides have been observed to regulate numerous cellular functions, including proliferation,24–27 differentiation,17,24,26,27 and protein expression. These behaviors are mediated by activation of various signaling28,29 pathways, frequently involving integrins and/or syndecans.27,28
The simultaneously presented biophysical and soluble signaling cues intrinsic to the microenvironment of cells are challenging to replicate in vitro. Corneal cells are simultaneously presented with the biophysical attributes of matrix topography, stiffness, and intrinsic surface chemistry in addition to soluble cytoactive factors. The differentiation of cells in the corneal stroma from the fibroblast to the myofibroblast phenotype is altered by both substratum topography and the presence of soluble cytoactive factors such as the transforming growth factor beta. 30
To improve the likelihood that results obtained in vitro will be predictive of the in vivo condition, it is imperative that in vitro models integrate biophysical cues along with relevant cytoactive factors in the experimental design. Numerous studies from our laboratories have documented that corneal epithelial cells respond to nano- and submicron anisotropic substratum topographic cues by altering alignment8,10,15,31 either parallel or perpendicular to the long axis of the underlying topographic features. The goal of this study was to investigate the integrated response (alignment, migration, and morphology) of corneal epithelial cells to the simultaneous presentation of surface chemical cues (using ECM proteins) and anistropically ordered topographic features.
Materials and Methods
Fabrication of NOA81 topographically patterned substrates with alternating ridges and grooves
Patterned silicon chips containing alternating ridge and groove features were fabricated using X-ray lithography as previously described. 4 Briefly, the silicon masters were fabricated containing an array of 6 (2×2 mm) areas with pitches (pitch=ridge width+groove width) of 400, 800, 1200, 1600, 2000, and 4000 nm separated by planar control areas. These arrays were termed “6-packs.” Patterns were of equal ridge and groove width with a groove depth of 300 nm. A composite stamp of the silicon chip master was made of poly(dimethylsiloxane) to retain the topographic features. The pattern was then replicated into a thin layer of NOA81 (Norland Products) optical adhesive 32 deposited onto 35-mm tissue culture dishes using a spin coater and cured in an XL-1500 UV cross-linker under 365-nm light for 100 min. NOA81, a proprietary mercapto-ester compound of Norland Products, is supplied as a single component liquid adhesive that readily cures as a rigid polymer with exposure to UV light. We have previously demonstrated that NOA81 is a suitable material for cell culture.6,10
Preparation of substrates for cell culture
In preparation for cell culture, all NOA81 surfaces were sterilized by 280-nm UV light for 20 min in a laminar flow hood. To determine the role of the ECM proteins immediately before cell seeding, the surfaces were treated for 15 s with the fibronectin-collagen (FNC) coating mix (Athena Enzyme Systems). The FNC coating is a proprietary mixture of FN and collagen.
Cell culture
Immortalized human corneal epithelial cells (hTCEpi), 33 kindly provided by Dr. James V. Jester (UC Irvine), were routinely cultured in the EpiLife® medium supplemented with EpiLife defined growth supplement (EDGS; Life Technologies) and 1% (v/v) penicillin/streptomycin (Life Technologies) and were used between passages 40–60 for all experiments.
Atomic force microscopy
To determine the fidelity of topographic features following protein coating, polymeric topographically patterned substrates with and without FNC treatment were imaged in Dulbecco's phosphate-buffered saline (PBS), pH 7.4, in a contact mode using an atomic force microscope (MFP3D-Bio).
Immunocytochemistry
Cells were plated on 6-pack NOA81 substrates with or without FNC coating and incubated overnight. Cells were fixed in either 100% ice-cold methanol for 5 min or 2.5% formaldehyde in PBS (pH 7.4) for 10 min. Formaldehyde fixed cells were repermeabilized in 0.1% Triton X-100 in PBS for 5 min. Cells were immunostained with Alexa Fluor 568 Phalloidin (actin), or antibodies (Abcam) for beta-tubulin, vimentin, or FN for 20 min, and counterstained with DAPI for 5 min. Images were acquired with an Axiovert 200M microscope (Carl Zeiss).
Orientation analysis
Images of the actin cytoskeleton and the nucleus on each of the six packs were obtained using a 10× objective lens. A minimum of four separate images were taken from each patterned pitch and flat areas separating the topographies. Approximately 300–400 cells for a given topography were analyzed. Image analysis was performed using built-in functions in Igor Pro 6.1 (Wavemetrics, Inc.) along with user-defined functions to ensure that cellular orientation, elongation, and area data were determined and directly registered to the respective nucleus. Not all cells were included in our analyses. Inclusion criteria required that the cell body had to be fully contained within the borders of the image, not in physical contact with another cell and not undergoing mitosis (e.g., cells with more than one nucleus). The nuclei inclusion criteria also required that the numbers of nuclei and cells were identical and could be linked to each other.
Time-lapse microscopy and migration analysis
Epithelial cells were seeded at a density of 8000 cells/cm2 onto patterned NOA81 surfaces and allowed to adhere for 1h at 37°C, 5% CO2. Using Zeiss Axiovision 4.6 and an automated stage, sequential phase-contrast images were obtained every 10 min over 12 h on each topographically patterned pitch. Only individual cells that remained within the field of view, did not contact another cell or undergo mitosis during the course of the experiment were analyzed. The centroid of each cell was identified and using the Axiovision Image Tracker module, the x and y coordinates of the cell were obtained. Using these, the cell trajectories, and migration rate were determined. At least 50 cells per pitch were analyzed.
Statistical analysis
Statistical analysis was performed by one-way ANOVA followed by the Dunnett's multiple comparison test compared with respective controls (cells on planar surfaces or without FNC treatment). For determining the differential effect of FNC on the axis ratio and area, we first performed a box plot on the data. Results suggested that the median values for the distribution were similar, but multiple outliers were present. Thus, to mimimize misinterpretation of the outliers, measures of skewness and kurtosis for the data were determined.
Results
Molecular coating of FNC altered the contact guidance response of corneal epithelial cells
A molecular coating of FNC had a pronounced effect on how corneal epithelial cells responded to anisotropically ordered topographic features. On 400-nm pitch patterned substrates, a much smaller percentage of cells (less than half as many) aligned parallel to the long axis of the underlying topographic features in comparison with cells on substrates without FNC coating (Fig. 1). With increasing pitch, the alignment of cells parallel to the ridges and grooves increased on FNC-coated surfaces; however, an increase in alignment in the absence of FNC coating was not observed compared with the 400-nm surfaces. Also, a greater percentage of nuclei were aligned to the ridges and grooves on the 400- and 4000-nm surfaces in the absence of FNC coating. The orientation of cells and nuclei at each of the increasing intervals of 10° [parallel (0°–10°) to perpendicular (80°–90°)] is illustrated in Supplementary Figure S1 (Supplementary Data are available online at www.liebertpub.com/tea). By atomic force microscopy, it was verified that the molecular coating of FNC did not mask the fidelity of the underlying topographic patterns (Fig. 2A). Immunocytochemistry documented that the localization of tubulin in cells was more pronounced above the ridges of the FNC-coated topographies of greater than1200-nm pitch. This was not observed on non-FNC-coated surfaces of the same pitch (Fig. 2B). In contrast, vimentin was more randomly organized in cells cultured on FNC-coated surfaces, but became localized above the ridges of topographic patterns with pitches greater than 1200 nm that lacked FNC coating (Fig. 2C). On the smaller pitch topographic features, the localization of both tubulin and vimentin was random and similar to those of cells cultured on FNC-coated or -uncoated planar surfaces (data not shown).
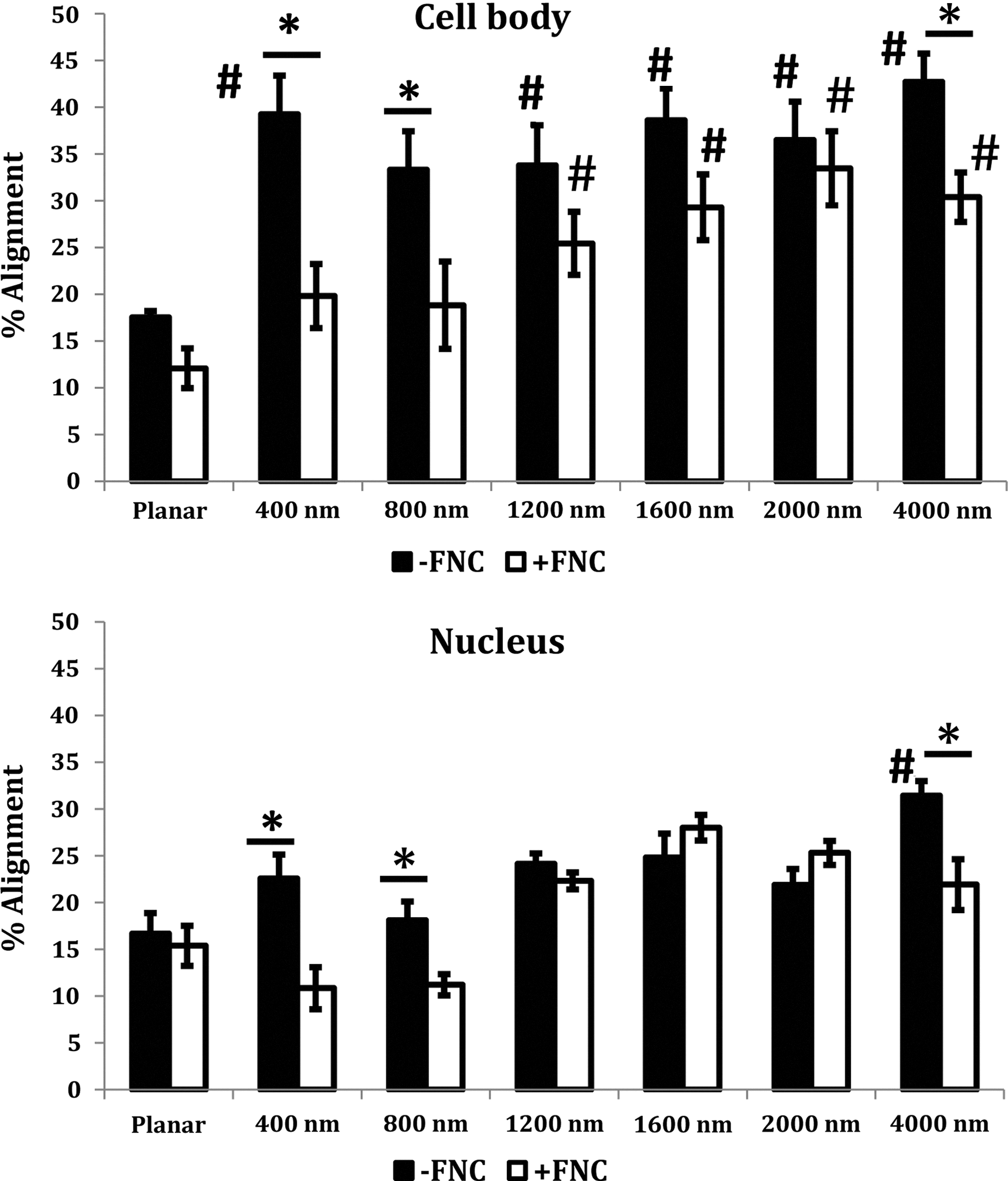
Corneal epithelial cell bodies and nuclei aligned significantly with underlying topographic features in the absence of extracellular matrix protein coating. However, in the presence of FNC coating, alignment of cell bodies increased with increasing topography. Results are mean±SEM. Statistically significant changes are indicated as follows: differences between topographic features and planar features for the same condition are marked with # (#p<0.05); differences between orientation as a consequence of the presence or absence of FNC for each pitch are marked with * (*p<0.05). FNC, fibronectin-collagen.
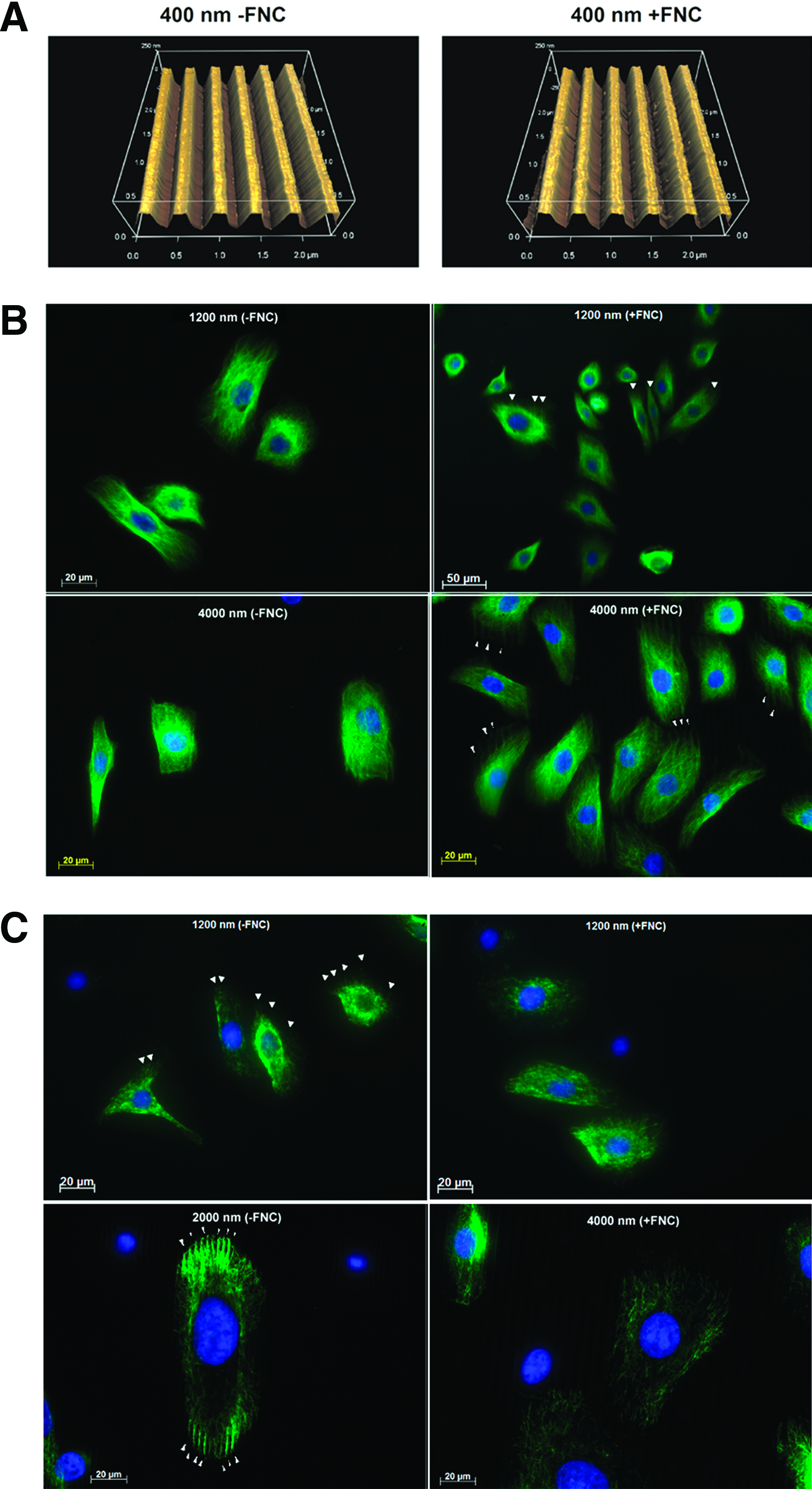
Cell area and elongation ratio are influenced by FNC coating
The median values for area and elongation ratios of cell bodies and nuclei on a given pitch for topographically patterned surfaces were similar when cells were cultured either in the presence or absence of FNC coating. However, a box plot of the raw data revealed that a greater number of cell bodies and nuclei had a larger area when cultured on FNC-coated surfaces (Fig. 3A). This was indicative of greater cell spreading. Also, from the box plots, a greater number of cells were observed with increased elongation/axis ratio of both cell bodies and nuclei when cultured in the absence of FNC coating (Fig. 3B) although the median values were similar. The skewness and kurtosis (Table 1) of the data for both the axis ratio and areas document significant differences in the two parameters with and without FNC coating on the topographies. A greater kurtosis number indicates that the distribution is skewed greater to the right. For example, the kurtosis in the axis ratio was significantly higher in cells on 400-nm pitch without FNC coating compared to those with FNC coating. Conversely, the kurtosis for the area was significantly lower for cells on 400-nm pitch without FNC coating compared to those with FNC coating. Taken together, this indicates that the cells on 400-nm surfaces that lacked FNC coating were longer, but had smaller areas. This can further be correlated with the increased orientation on 400 pitch surfaces without FNC coating. There was no trend observed for cells on planar surfaces or other pitches.
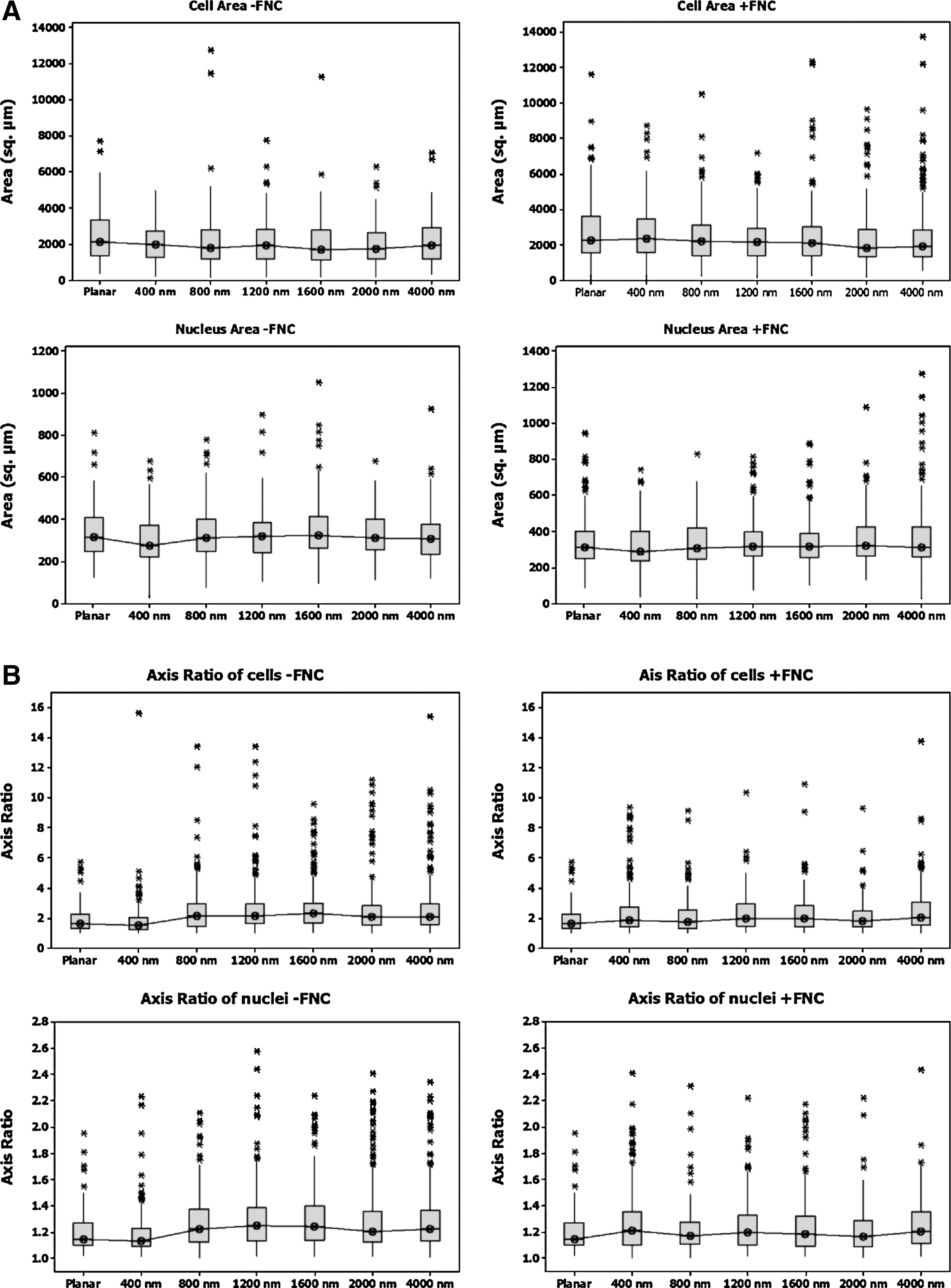
Box plots for the cell body and nuclei showed a wide distribution of
Significant differences in skewness and kurtosis were seen in the axis ratio and area measurements in cell bodies and nuclei of cells cultured on 400-nm topographic surfaces although the median values were very similar between both FNC treatment groups. An increase in the axis ratio accompanied by a reduced cell area on 400-nm surfaces suggests cell confinement and elongation along the ridges of the topography.
FNC, fibronectin-collagen.
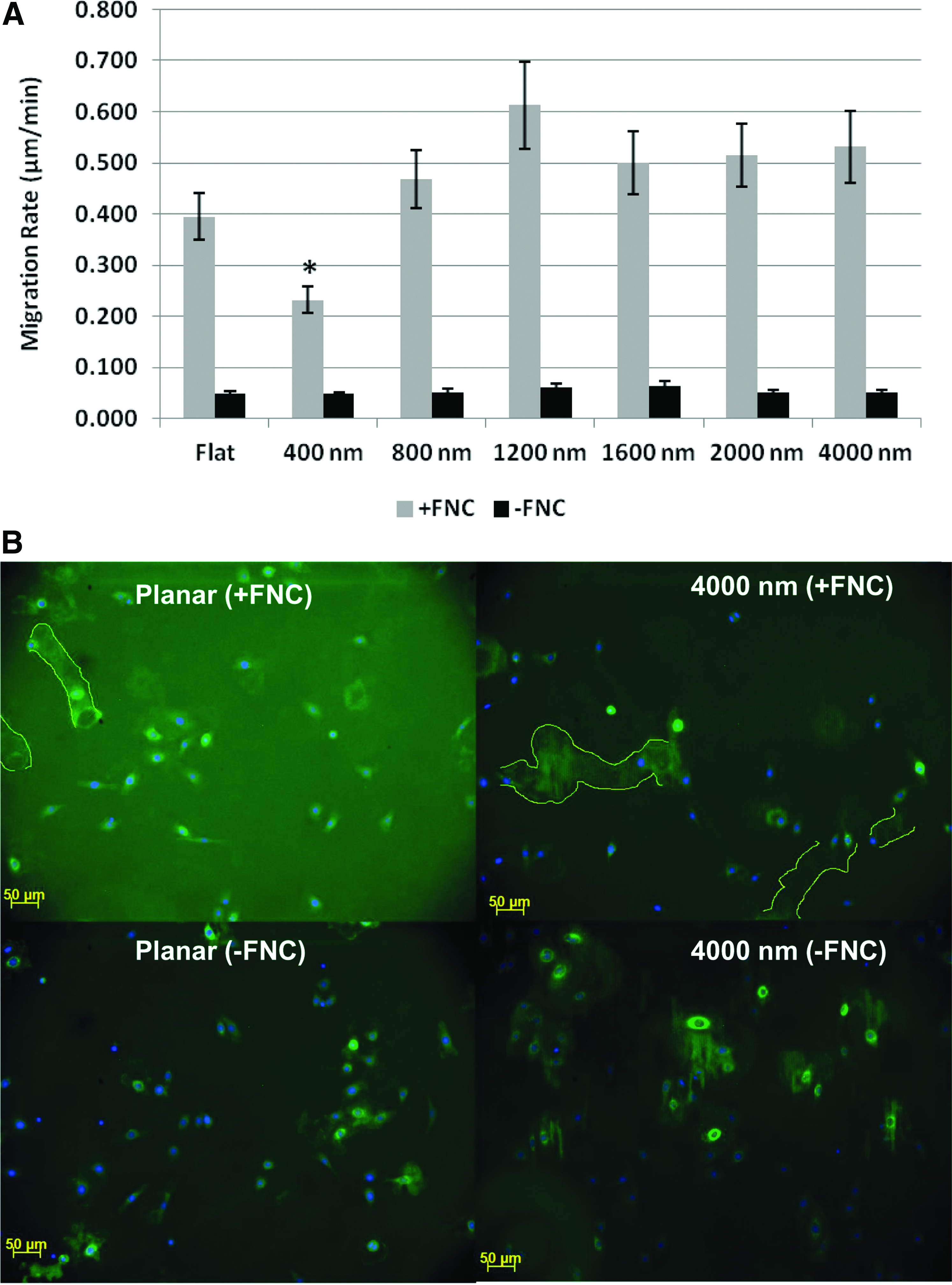
Corneal epithelial cell migration was inhibited by biologically relevant substratum topography and promoted by the presence of FNC coating
On substrates that lacked FNC coating, cell migration was not observed regardless of the presence or absence of topographic features. On FNC-coated surfaces cell migration was observed and was influenced by the scale of topographic features present (Fig. 4A). Migration was significantly slower on the 400-nm-size scale ridges and grooves compared to the planar surface, while it was highest on the 1200-nm pitch patterned surfaces. Immunostaining to FN on non-FNC-coated substrates revealed the presence of FN only in areas confined to the presence of corneal cells suggesting the deposition of FN. Immunostaining against FN on FNC-coated surfaces showed diffused labeling of FN on areas lacking cells; a dense region of FN indicative of the migratory path, which a cell had taken was also observed. It is not certain if this more concentrated area of FN represents the newly synthesized protein or a rearrangement of the FN, initially coated on the substrate that was reassembled by the cell (traced with a yellow line, Fig. 4B).
Discussion
The BM of the cornea possesses intrinsic surface chemistry and multiple biophysical cues (e.g., stiffness and topography) as well as serving as a reservoir for soluble cytoactive factors. The corneal BM is primarily comprised of collagen, laminin, FN, and fibrillin among other matrix proteins.34,35 In vivo, cells simultaneously integrate these disparate biophysical and biochemical cues to maintain homeostasis. The modulation of cellular behavior and differentiation by soluble factors,15,36–39 surface chemistry,40–44 and biophysical attributes of the extracellular microenvironment45–48 has been well established. While, the majority of studies have investigated only one type of biophysical or chemical cue in isolation, a limited number have explored the interaction between multiple biophysical cues,11,49 surface chemistry,19,50 or soluble cytoactive factors. 15
Here we have demonstrated that the simultaneous presentation of substratum topographic cues and surface-associated biochemical cues potently modulated cellular behavior. Specifically, we demonstrated that corneal epithelial cells responded differentially to substratum topographic cues in the presence or absence of an exogenously applied ECM protein coat. The response of cells to topographic cues in the absence of an ECM coat was consistent with previous reports from our laboratory.4,39 A greater number of cells aligned parallel to the long axis of ridges and grooves of the topography (on biomimetic size scale, 400 nm) in the absence of FNC coating; however, the migration of cells was significantly less on noncoated substrates. With the provision of a molecular coating of FNC, the migration of cells was promoted and varied with the size scale of the underlying topographic features. Migration of cells on 400-nm surfaces was significantly inhibited compared to cells on planar surfaces. Additionally, alignment along the ridges and grooves was less on the patterned surfaces possessing biomimetic size scale (400-nm pitch). Our results also demonstrated that on pitch sizes greater than 1200 nm, cell migration though not cell alignment was influenced by ECM coating of the substrate.
Migration of cells into a wound is critical to wound repair. FN has previously been demonstrated to appear prominently on the corneal surface, within 1 h of wounding in rabbits, and remained until the epithelium had covered the wound. 51 In this study, we demonstrated that corneal epithelial cells deposited FN both in the presence or absence of an exogenously applied FNC coating. However, precoating surfaces with FNC promoted cell migration and this correlated with well-established reports that FN promotes epithelial cell migration in the cornea.52,53 We note that the migratory pattern in the immortalized hTCEpi cells used in this study differed from that observed in SV40 transformed corneal epithelial cells previously published by our group. 54 SV40 transformed corneal cells migrated parallel to the long axis of the underlying ridges and grooves and their migration rate on these topographically patterned substrates was significantly higher than that on planar surfaces. 54 However, in the current study, hTCEpi cells migrated perpendicular to the long axis of their cell body irrespective of their orientation with the ridges and grooves (Supplementary Movie S1). We verified this migratory pattern with primary corneal epithelial cells (data not shown) that also migrated perpendicular to the long axis of their cell body irrespective of the topography in the presence of FNC coating.
Differences in cell types (hTERT & primary vs. SV-40) may also be partially responsible for the differences in these studies. While we do not at present completely understand the significance of such a different migration pattern, it must be noted that the differential influence of soluble factors in a growth medium (the serum-free defined EpiLife medium versus serum-containing EP growth medium) or the dimensions of the topographies (equal size for ridges and grooves in this study versus unequal ridge/groove width) cannot be discounted. In previous studies from our laboratory,15,54 the dimensions of the ridges and grooves were asymmetric (i.e., unequal ridge (70 nm) and groove (330 nm) width for the same pitch (400 nm)) and cells were cultured in media containing serum and other soluble factors. In the present study, the dimensions of the ridges and groove width were equal (i.e., ridge width=groove width; e.g., for 400-nm pitch, ridge width=groove width=200 nm). Also, in this study, hTCepi cells were cultured in a proprietary defined medium (EpiLife) supplemented with selective growth factors without serum. It is thus possible that the differences observed between this study and previous may be dependent on dimensions and/or soluble factors.
Cytoskeletal remodeling, influenced by topographical cues, is mediated by complex signaling mechanisms initiated at the integrin-ECM interface and involves the Rho family proteins, Cdc42, PI3Ks, PTEN, Rac proteins, and many others. 55 An earlier study from our group using human umbilical vein endothelial cells demonstrated that over 3000 genes, many associated with the ECM and cell adhesion, cytoskeletal reorganization, DNA replication and repair, and spindle organization, were significantly altered by biomimetic size scale (400-nm pitch) topographic features. 56 It is known that while nanoscale topographic cues can modulate the expression of integrins in cells, 57 there is bidirectional signaling between the cytoskeleton and integrins. 58 Additionally, the mechanical link between integrins and various cytoskeletal filaments aids in stabilizing the nuclear structure and shape and this can in turn regulate gene expression.59,60 In this study, we report that both the cell body and nuclear shape were altered by both topography and exogenous FNC coating.
Microtubules (tubulin) were aligned parallel to the long axis of the topographic features on FNC-coated surfaces, but did not have a distinctive spatial orientation in the absence of FNC coating. On the other hand, intermediate filaments (vimentin) were observed to be aligned with the ridges in the absence of FNC coating, but lacked this oriention on noncoated surfaces. Our results suggest that a molecular coating of FNC alters the cytoarchitecture in corneal cells with the microtubules more responsive on surfaces that had been coated with FNC particularly on the larger pitches. In the absence of FNC coating, microtubules in corneal cells may not have been critically aligned to facilitate migration of the cell. Thus, the provision of an exogenous ECM coating may not only facilitate cell migration on topographically patterned surfaces, but may also alter how mechanotransduction and subsequent cell-signaling occurs. These biological consequences are relevant to the evolving strategies for design and fabrication of tissue scaffolds or synthetic materials for prosthetic devices.
Conclusions
Our results demonstrated that cells perceived and responded differently to the simultaneous presentation of biophysical cues and surface chemistry than to either cue in isolation. If the goal of in vitro studies is to best predict in vivo outcomes and if the goal of tissue engineering and prosthetic design is to best promote bio-integration, then results from this study and others highlight the need for integration of surface chemistry and biophysical cues in simulating the in vivo environment.
Footnotes
Acknowledgments
This project was supported by the National Institute of Health through grants from the National Eye Institute R01EY01613404 and P30EY12576 and unrestricted funds from Research to Prevent Blindness.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.