
Abstract
Introduction:
The aim of the study reported here was to investigate the molecular responses of human mesenchymal stem cells (MSC) to loading with a model that attempts to closely mimic the physiological mechanical loading of bone, using monetite calcium phosphate (CaP) scaffolds to mimic the biomechanical properties of bone and a bioreactor to induce appropriate load and strain.
Methods:
Human MSCs were seeded onto CaP scaffolds and subjected to a pulsating compressive force of 5.5±4.5 N at a frequency of 0.1 Hz. Early molecular responses to mechanical loading were assessed by microarray and quantitative reverse transcription-polymerase chain reaction and activation of signal transduction cascades was evaluated by western blotting analysis.
Results:
The maximum mechanical strain on cell/scaffolds was calculated at around 0.4%. After 2 h of loading, a total of 100 genes were differentially expressed. The largest cluster of genes activated with 2 h stimulation was the regulator of transcription, and it included FOSB. There were also changes in genes involved in cell cycle and regulation of protein kinase cascades. When cells were rested for 6 h after mechanical stimulation, gene expression returned to normal. Further resting for a total of 22 h induced upregulation of 63 totally distinct genes that were mainly involved in cell surface receptor signal transduction and regulation of metabolic and cell division processes. In addition, the osteogenic transcription factor RUNX-2 was upregulated. Twenty-four hours of persistent loading also markedly induced osterix expression. Mechanical loading resulted in upregulation of Erk1/2 phosphorylation and the gene expression study identified a number of possible genes (SPRY2, RIPK1, SPRED2, SERTAD1, TRIB1, and RAPGEF2) that may regulate this process.
Conclusion:
The results suggest that mechanical loading activates a small number of immediate-early response genes that are mainly associated with transcriptional regulation, which subsequently results in activation of a wider group of genes including those associated with osteoblast proliferation and differentiation. The results provide a valuable insight into molecular events and signal transduction pathways involved in the regulation of MSC osteogenic differentiation in response to a physiological level of mechanical stimulation.
Introduction
Various systems have been developed to mimic mechanical stimuli for investigating bone turnover in vivo and in vitro. Most of these have mainly focused on studying osteocytes as mechanosensors and osteoblasts as bone forming cells in their models. Commonly a gel based scaffold or tissue culture plastic has been used as the substrate on which cells were mechanically loaded, and the strain employed has been in the range of 2%–20%.6,12–20 Neither the scaffold nor the strain of this magnitude represents the true environment or magnitude of physiological or pathological loading of bone in humans in vivo. Further, a number of studies in vivo have been carried out in an attempt to overcome these obstacles, but these are limited to animal models and therefore could be associated with differences in responses to loading and mechanisms involved from one species to another.21,22 In addition, the presence of a mixed population of cells within these settings makes it difficult to study responses from specific cell types such as osteoblasts or mesenchymal stem cells (MSC).
Therefore, the aim of this study was to investigate the molecular response of MSCs to transient or continuous mechanical stimulation using a model that attempts to mimic the situation in vivo. A monetite calcium phosphate (CaP) scaffold was used to model the mechanical properties of bone and load and strain were induced using a bioreactor. A mathematical model was used to calculate a load required to mimic changes in strain equivalent to physiological-scale effects that have been reported previously. 23 A global gene expression microarray was carried out to identify candidate transcriptional regulators and signaling mechanisms induced by mechanical stimulation.
Materials and Methods
Cell culture
Commercially obtained primary human MSCs from three separate donors (Lonza), at passage 2 when purchased were expanded in culture and used at passage 3–5. Cells were maintained in normal growth medium consisting of α-Minimal Essential Medium (α-MEM), penicillin (50 U/mL), streptomycin (50 μg/mL) (Sigma-Aldrich), Glutamax (2 mM) (Invitrogen), and 10% fetal bovine serum (FBS) (Sigma-Aldrich) at 37°C in a humidified 5% CO2:95% air atmosphere.
MSCs were characterized for cell surface marker expression using four-color flow cytometry analysis as previously described. 24 To induce osteogenesis, MSCs were seeded at a density of 1.2×104 cell/cm2 in 12-well plates and after 24 h were treated with osteogenic medium consisting of growth medium supplemented with 0.1 μM dexamethasone, 0.05 mM ascorbic acid, and 10 mM β-glycerophosphate (Sigma-Aldrich). Mineralization was assessed by staining with alizarin red dye. Briefly, cells were fixed (15 min with 4% formaldehyde in phosphate-buffered saline [PBS]), stained for 10 min with alizarin red S (1:100 dilution in H2O), washed in 50% ethanol, and air-dried. To induce adipogenic differentiation, cells were seeded at a density of 4×104 cell/cm2 in 12-well plates (Nunc) and incubated for 24 h before switching to adipogenic medium (growth medium supplemented with 1 μM dexamethasone, 0.25 mM isobutylmethylxanthine, 50 μM indomethacin, and 10 μg/mL insulin; Sigma-Aldrich). Adipogenesis was visualized by light microscopy following staining with Oil red O dye. Briefly, cells were fixed (15 min with 4% formaldehyde in PBS), stained for 15 min, and washed with 60% isopropanol and with PBS.
Mechanical stimulation
Macroporous monetite cements were prepared with a solid phase made of equimolar quantities of β-tricalcium phosphate (β-TCP; assay >96%) (Fluka) and monocalcium phosphate monohydrate (MCPM; assay min. 98%) (Sharlau) with 60% w/w sodium chloride (NaCl) crystals in respect to the CaP powder were added as a water soluble porogen. The polygonal shaped salt crystals were sieved to achieve a diameter in the range of 150–600 μm. The liquid phase was a solution consisting of 0.5 M of sodium dihydrogen citrate. The powder to liquid ratio (R) was fixed at a value equal to 3 (g·mL−1). The mixture was manually stirred for ∼1.5 min. Following mixing, the paste was poured into two different Teflon moulds of cylindrical shape (one of length: l=12 mm, diameter: d=6 mm and the other of length: l=4.2 mm, diameter: d=8.4 mm) to make samples for the evaluation of the mechanical properties by uniaxial or diametral compression (“brazilian”) test in accordance to the ASTM F451 and C496 standard respectively. The samples were left to set for 24 h at ambient conditions (20°C–23°C and 50%–60% humidity) and then immersed in distilled water for 5 days to let the porogen dissolve. Identification of monetite was achieved by means of X-ray diffraction on powder samples using a Bruker D8 Advance diffractometer equipped with a Lynx Eye detector, in flat plate geometry using Ni-filtered Cu kα radiation. The phase composition was checked using the reference patterns for β-TCP (PDF Ref. 09-0169), monetite (PDF Ref. 09-0080) and brushite (PDF Ref. 09-0077). The distribution of macropores and the volume of the pores on the cement were observed by SEM (Hitachi S-3500 machine) and micro-CT (Locus SP, microCT scanner; GE Healthcare) analysis set at a resolution of 7 μm.
The mechanical tests were carried out by using a universal testing machine (Instron 5699) with a 500 N load cell and a cross-head speed of 1 mm/min. Twelve samples were used for each test. The tensile strength of the material from the diametral compression tests had an average of 0.54 (±0.01) MPa, corresponding to a maximum axial load of 30 N. This preliminary evaluation allowed us to define the dynamic load that can be applied in the bioreactor tests without risk of failure for the scaffolds.
MSCs were cultured to 80%–90% confluence in normal growth medium. Cells were then seeded on to the top of one side of the scaffold at a density of 1.5×105 in 70 μL of media and allowed to attach for 30 min. The scaffold was then turned and the same number of cells were seeded on to the top of the other side and left undisturbed for a further 30 min for the cells to attach. Normal growth media was then carefully added to the culture plate and the cell/scaffold was incubated at 37°C in a humidified 5% CO2:95% air atmosphere for 48 h prior to stimulation. Mechanical loading was performed using a BOSE bioreactor (ElectroForce BioDynamic test instrument; Bose) equipped with 200N load cell (Fig. 1A). Seeded scaffolds were positioned between two loading plates inside the bioreactor chamber (Fig. 1A[iv]) and loaded in the diametral compression mode by a pulsating compressive force of 5.5±4.5 N at a frequency of 0.1 Hz. The corresponding head displacement was 0.5–50 μm. The test was performed in load control and the evolution of the head displacement versus the pulsating compressive force was recorded by the bioreactor software (WinTest® controls). The stress distribution in the disk was analytically evaluated by the Timoshenko model. 25 For controls, cells were incubated for the same period of time on samples positioned between the loading plates of the bioreactor with no load applied.

The diametral compression test configuration was chosen because it generates a complex state of stress where all of the in-plane stress components (normal and shear stresses) are present. In this way the influence of the mechanical loading on cell behavior had a higher probability of being observed. The distributions of radial (σrr), circumferential (σθθ), and shear (τθr) stress components shown in Figure 2A were calculated with reference to the load factor (Eq. [1], where P is the applied load, R the radius of the circular sample, and t its thickness):
In the bioreactor tests, most of the cells were attached to the outer surface of the scaffold, where no stress is directly applied. Therefore, the situation is different from that of scaffolds submerged in a liquid current, where an effective shear stress is present at the surface and an increase of cell activity has been observed by others.26–28
In the present case however, a physical stimulation of the cells as a consequence of mechanical loading can occur on account of the stretching they undergo while following the elastic deformations of the scaffold. The maximum (ɛI) and minimum (ɛII) strains are related to the principal stresses σ
I
and σ
II
by Equations (2) and (3), respectively;
Accordingly, we have the distributions of ɛI and ɛII in the disk (referred to σ o /E), which are represented in the plot of Figure 2B where, E is the Young's modulus and ν is the Poisson ratio of the scaffold respectively.

The stress analysis of disk subjected to two concentrated diametral forces.
The averages of the maximum
Microarray analysis
For total gene expression analysis, samples from three primary MSC lines were loaded or left unloaded for 2 h in the bioreactor chamber. Samples were then removed from the bioreactor and the cells were either lysed for RNA isolation or incubated for a further 6 or 22 h in normal culture medium at 37°C in a humidified 5% CO2:95% air atmosphere prior to RNA isolation. Total RNA was extracted using TRI reagent (Ambion) and Phase Lock Gel Heavy tubes (5 prime; VWR) according to the manufacturer's instructions. RNA purity and quantity was assessed by nanodrop (Fisher Scientific) and Qubit 2.0 (Invitrogen). RNA integrity was determined using an Agilent 2100 Bioanalyzer (Agilent). An equal quantity of RNA from three separate experiments was pooled together for each MSC line and processed for microarray analysis. The gene expression analysis was performed using the Illumina whole-genome expression array HumanHT-12 v4.0 Expression BeadChip according to the manufacturer's instructions. Briefly, 500 ng of total RNA was processed and cRNA was hybridized to an Illumina whole genome expression chip. The Beadchips were scanned on the Illumina iScan System with iScan software. The raw data were processed for the background signal, normalized, and differences in gene expression were compared by the Differential Expression Algorithm using the Illumina custom error model of GenomeStudio software (Illumina, Inc.). Genes with a Diffscore of±13, equivalent to p<0.05, were considered as being significantly down- or upregulated; a DiffScore of±20,±30 or±40 was equivalent to a p-value of <0.01, <0.001, or <0.0001, respectively. Gene Ontology (GO) analysis was subsequently carried out on genes significantly up- and downregulated using the Database for Annotation, Visualization, and Integrated Discovery (DAVID) 6.7 Web tool to study associations corresponding to biological processes (BP-FAT). The software categorizes related genes into functionally related clusters and rank them based on the statistical calculations according to their cluster enrichment by EASE score, which is calculated as higher probability to get a GO term under the null hyperthesis than would be randomly selected from the total number of gene on the microarray.
Gene Expression
cDNA was prepared from 1 μg RNA using QuantiTect Reverse Transcription Kit (Qiagen) according to the manufacturer's instructions. Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) was performed on a rotor gene 6000 thermal cycler (Qiagen) using Brilliant III Ultra-Fast SYBR Green qPCR Master mix (Stratagene; Agilent Technologies) and primer pairs as listed in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/tea). PCR conditions consisted of 1 cycle of 95°C for 3 min and 40 cycles of 95°C for 10 s and 60°C for 10 s followed by melting analysis of 1 cycle with gradual increase from 65°C to 95°C. RPL13a was used as an invariant housekeeping gene. The relative quantitative expression of genes of interest, relative to housekeeping RPL13a gene was calculated and compared with untreated cells that were assigned a value of one. RPL13a was used as it has been shown by our group and others to be a stable housekeeping gene for qPCR analysis of MSCs during in vitro characterization and differentiation. 29
Western blotting analysis
MSC/scaffolds were incubated with serum-free α-MEM for 6 h prior to mechanical stimulation. Cells were then loaded for the times indicated then immediately lysed and subjected to western blot analysis as described previously 30 using antibodies against pAkt (Ser473) and p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (Cell Signalling; New England Biolabs). Proteins were visualized and photographed using ECL Plus detection reagent (GE Healthcare) and Molecular Imager Gel Doc XR+ documentation and analysis System with Image Lab Software (Bio-Rad). Equal sample loading was confirmed by probing with antibody to the housekeeping protein GAPDH.
Cell morphology and viability
Phalloidin-TRITC (Sigma) was used to stain for actin filaments. Briefly, MSC/scaffolds were fixed for 15 min with 4% formaldehyde, permeabilized for 10 min with 0.1% Triton X-100, and stained for 40 min with a 0.5-μg/mL phalloidin-TRITC conjugate solution in PBS. Samples were washed with PBS and visualized and imaged using a fluorescence microscope. Cell viability was determined using the Live/Dead, viability/cytotoxicity assay (Invitrogen). Cells were stained with 2 μM calcein AM and 4 μM ethidium homodimer-1 (EthD-1) in PBS for 20 min. Samples were visualized using a fluorescence microscope with live and dead cells staining green (calcein AM) or red (EthD-1) respectively.
Data analysis
For qRT-PCR analysis, statistical comparisons between means were made by Student's t-test (SPSS 16; SPSS). A p-value of less than 0.05 was considered statistically significant. The data were pooled from duplicate tests carried out in three separate experiments.
Results
MSC characterization
Prior to mechanical loading experiments, MSCs were assessed for their cell surface marker expression and differentiation potential. Using four-color flow cytometry analysis of panels of known negative and positive markers we showed that MSCs were 90%–100% positive for CD105, CD90, CD44, and CD146 but lacked the HSC marker (0%–5%) CD34 and CD45 expression (Fig. 3A). We also demonstrated the potential of MSCs to differentiate toward osteoblasts and adipocytes, by the increase in specific expression of markers of each lineage, that is, Runx-2, ALP, PPARγ, and FAB-4 and accumulation of calcium deposits and lipid droplets by the cells following culture for 14 days in differentiating media. (Fig. 3B, C).
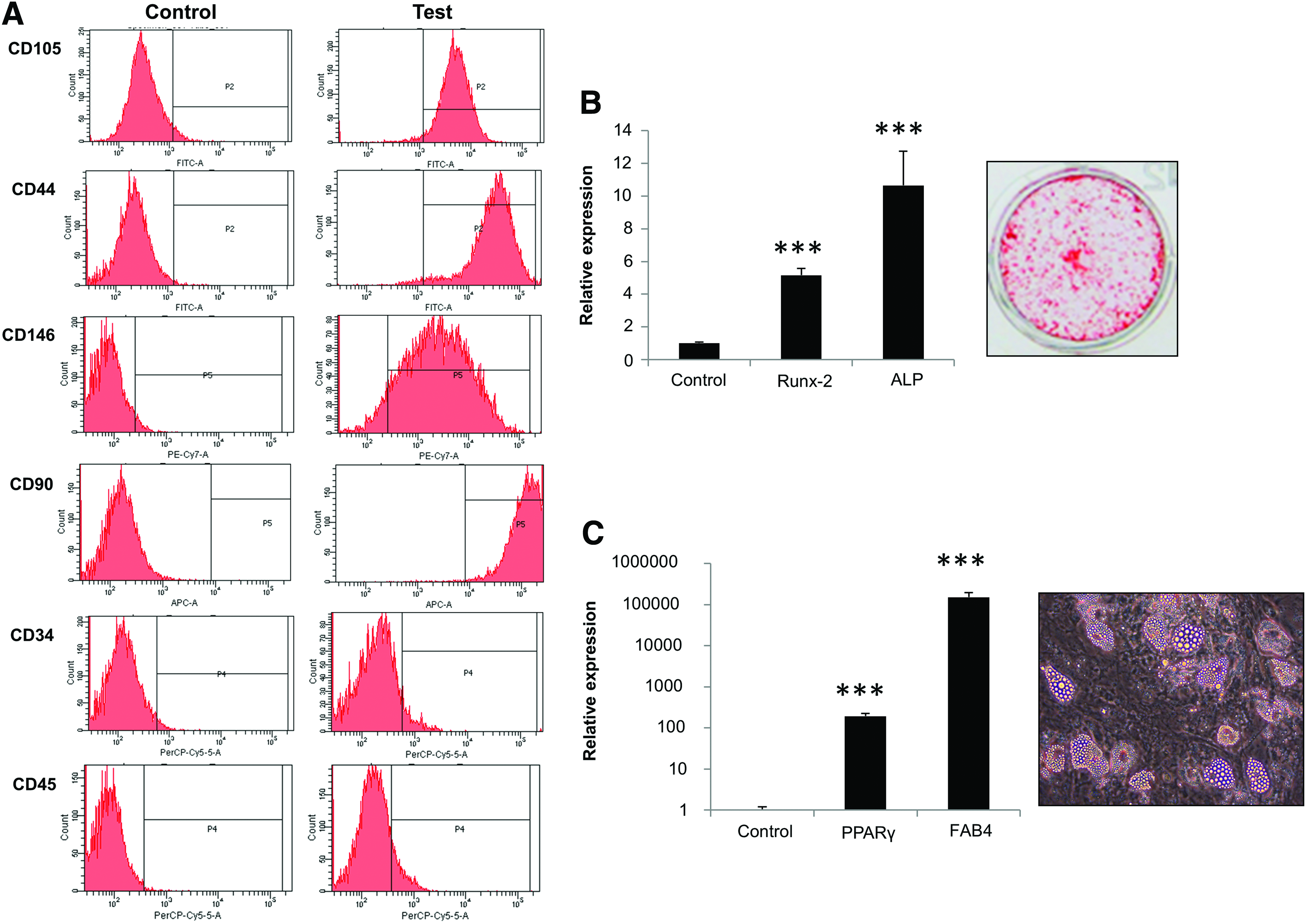
Characterization of mesenchymal stem cells (MSCs).
Monetite scaffold characterization and strain values over the disk
X-ray diffraction confirmed that the final composition of the cement was monetite. 31 Figure 1B is an SEM image of the fracture surface of macroporous monetite cement tested by the uniaxial compressive test showing the pores produced by the dissolution in distilled water of the NaCl crystals. The micro-CT analysis reported an average porosity of 45% with pore diameters between 200 and 650 μm. 31 The pores were distributed throughout the cement while the structure maintained sound continuity along its multiple isosurfaces in all directions (X, Y, Z).
The mechanical properties of the monetite cements are reported in Figure 1C as average and standard deviation values of the maximum compressive strength (σc), Young's modulus (E), and maximum diametral strength (σt). The calculated distributions of radial (σrr), circumferential (σθθ), and shear (τθr) stress components are reported in Figure 2A. The average strain values in a loading cycle were calculated using Equations (4) and (5) above, where ν=0.3 is the Poisson ratio of the ε material. Their values in a loading cycle were as follows:
The strain amplitudes in a cycle (0.38; 0.18)% are comparable to those strains that have been previously reported to act on long bones. 23
Effect of mechanical loading on MSC gene expression
Global gene expression analysis of MSCs showed a total of 100 differentially expressed genes compared with unloaded MSCs following 2 h of loading. Among these, 75 genes were significantly upregulated and 25 downregulated in loaded MSCs compared to unloaded cells (Supplementary Tables S2 and S3). When MSCs were cultured for a further 22 h after 2 h of loading, 63 genes were also differently expressed, of which 40 were upregulated and 23 were downregulated when compared to unloaded cells (Supplementary Tables S4 and S5). When MSCs were rested in culture for 6 h following the loading, there were no significant differences in the levels of gene expression compared to unloaded controls (Supplementary Tables S6 and S7). To predict better the cascade of events that takes place during mechanical stimulation and the signaling mechanisms involved, we categorized the differentially expressed genes based on the biological processes that they are involved in. As shown in Tables 1–4 genes upregulated following 2 h of loading were mainly involved in regulation of transcription, protein kinase cascade, regulation of MAP kinase cascade, and activity and metabolic processes. However, when cells were rested for 22 h after 2 h loading, the upregulated genes were predominantly associated with cell surface receptor-linked signal transduction, regulation of metabolic processes, and cell division. In contrast, genes that were downregulated with 2 h of loading or after 22 h of rest following stimulation were mainly negative regulators of cell proliferation and cell cycle. Genes involved in programmed cell death and regulation of apoptosis were also downregulated with 2 h loading.
qRT-PCR validation of microarray data
To confirm the data obtained by microarray analysis, we carried out qRT-PCR analysis of RNA samples used for the microarray experiments. We selected five differentially expressed genes, namely FOSB, SPRY2, SERTAD1, CDKN1b, and CEBPδ based on their known roles in physiological processes involved in regulation of cell fate (differentiation or proliferation). The changes in expression of these genes obtained by microarray were 5.4, 1.94, 1.65, 0.64, and 0.55-fold, respectively compared to 10.3, 2.98, 5.3, 0.75, and 0.63-fold changes observed with qRT-PCR. As illustrated here and in Figures 4 and 5 qRT-PCR analysis confirmed the changes in these selected genes, showing significant changes in expression in accordance with the array data. The high consistency between assays gives us confidence of the accuracy of our microarray data. In addition, we tested the expression of the osteoblast-related transcription factor Runx-2. Although the microarray results did not show any significant differences in expression, when tested by qRT-PCR Runx-2 expression was significantly increased after 2 h of loading followed by a further 22 h resting (Fig. 6).
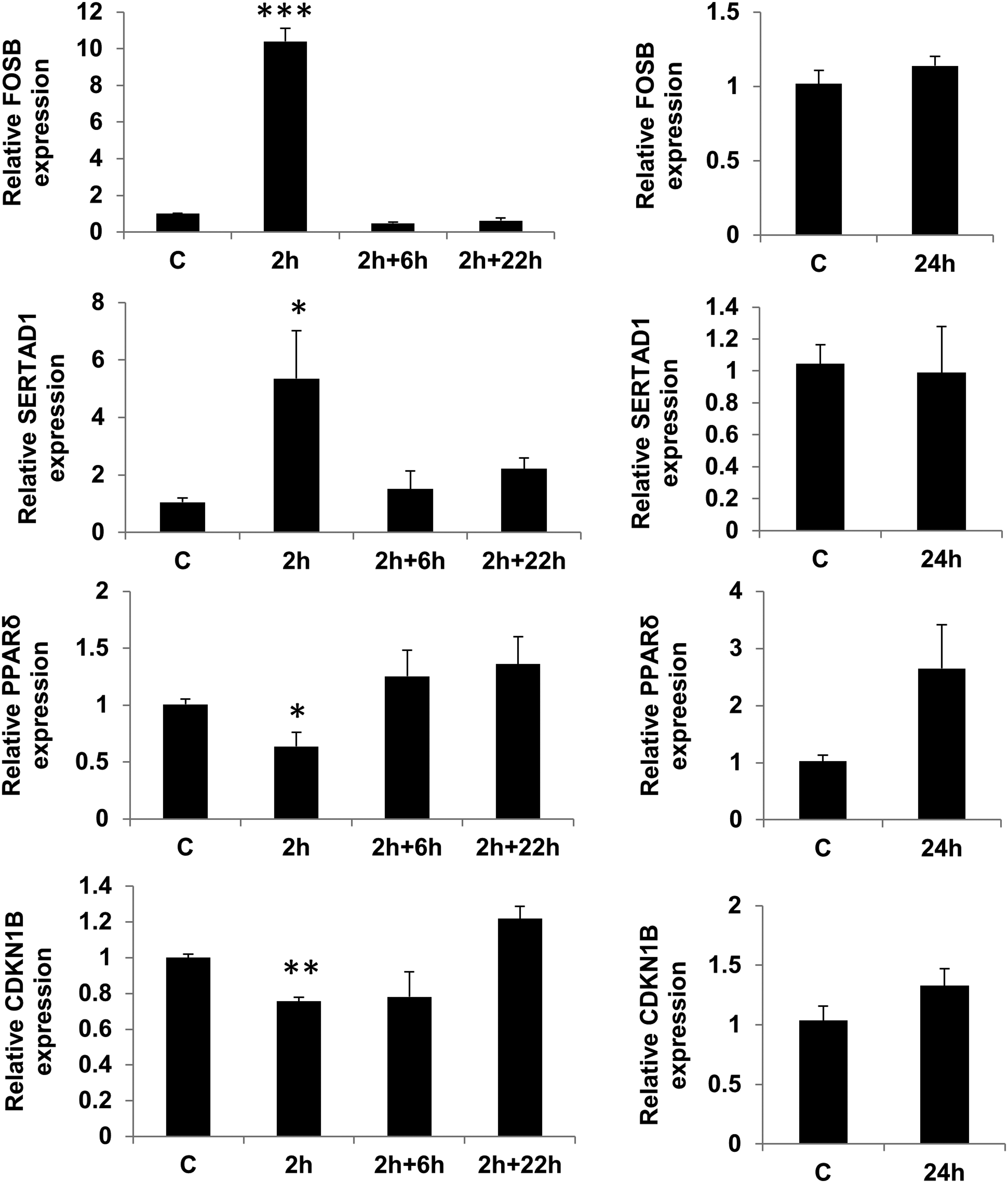
Evaluation of microarray data. qRT-PCR analysis was carried out on selected differently expressed genes (FOSB, SERTAD1, CDKN1B, and CEBPδ) on RNA samples used for the microarray experiments. A comparison was made between 2 h of loading, 6 h post 2 h loading (2+6), 22 h post 2 h loading (2+22), and following 24 h of constant loading. Each time point was normalized to housekeeping gene and relative expression was calculated in relation to appropriate unloaded samples and this ratio was assigned a value of one. (mean±SEM of three experiments of duplicates). *p<0.05, **p<0.01, and ***p<0.001 when compared with unloaded cells for the same time point.
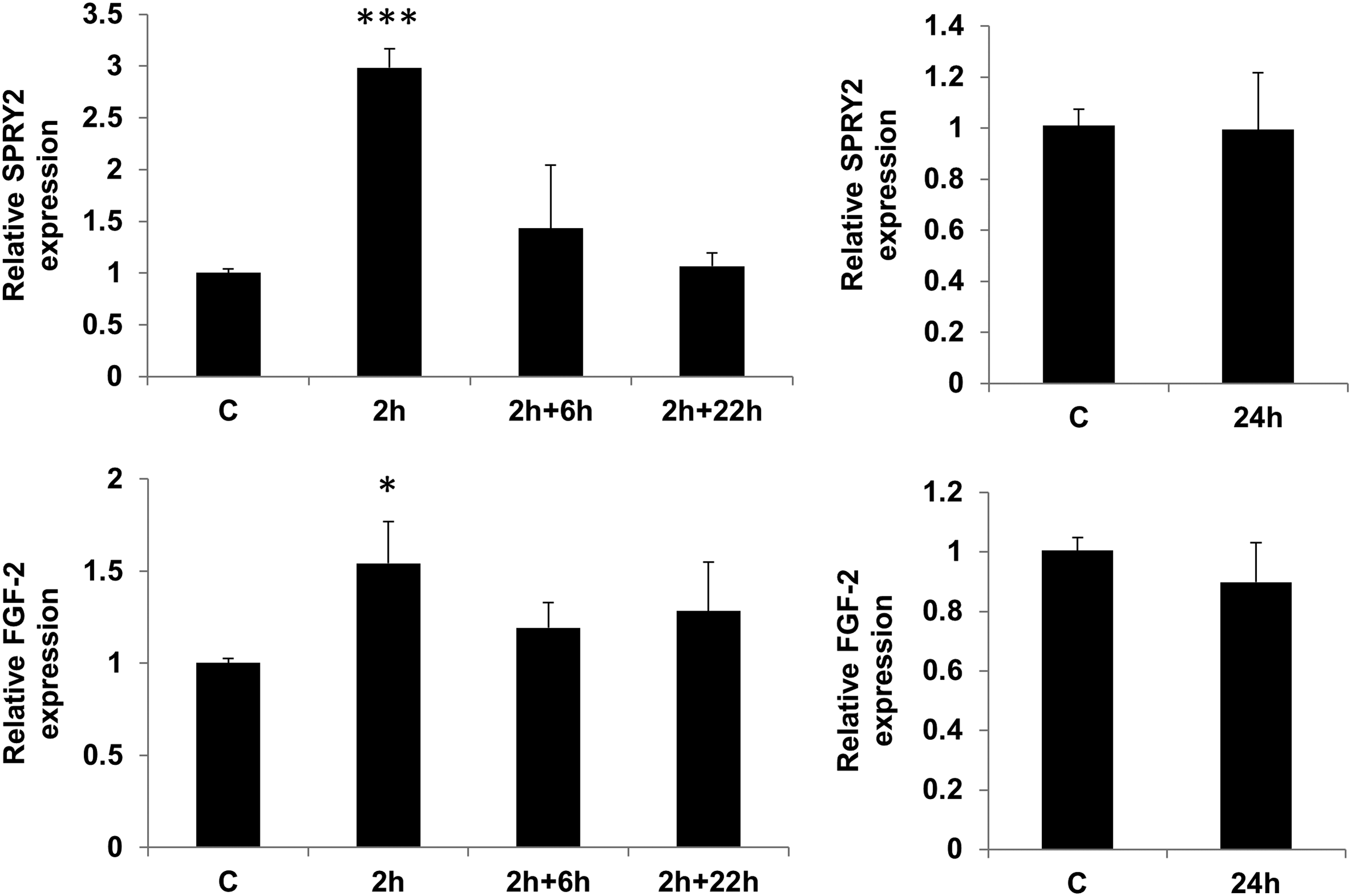
Effect of mechanical stimulation on FGF-2 and SPRY-2. Cells were loaded with 0.4% strain and analyzed after 2 h of loading, following 6 or 22 h rest post 2 h loading (2+6 and 2+22) and following 24 h of constant loading. mRNA expression for FGF-2 and SPRY-2 was determined by qRT-PCR and compared with unloaded cells after normalization to RPL13a. (mean±SEM of three experiments of duplicates; **p<0.01 and ***p<0.001 vs. untreated).
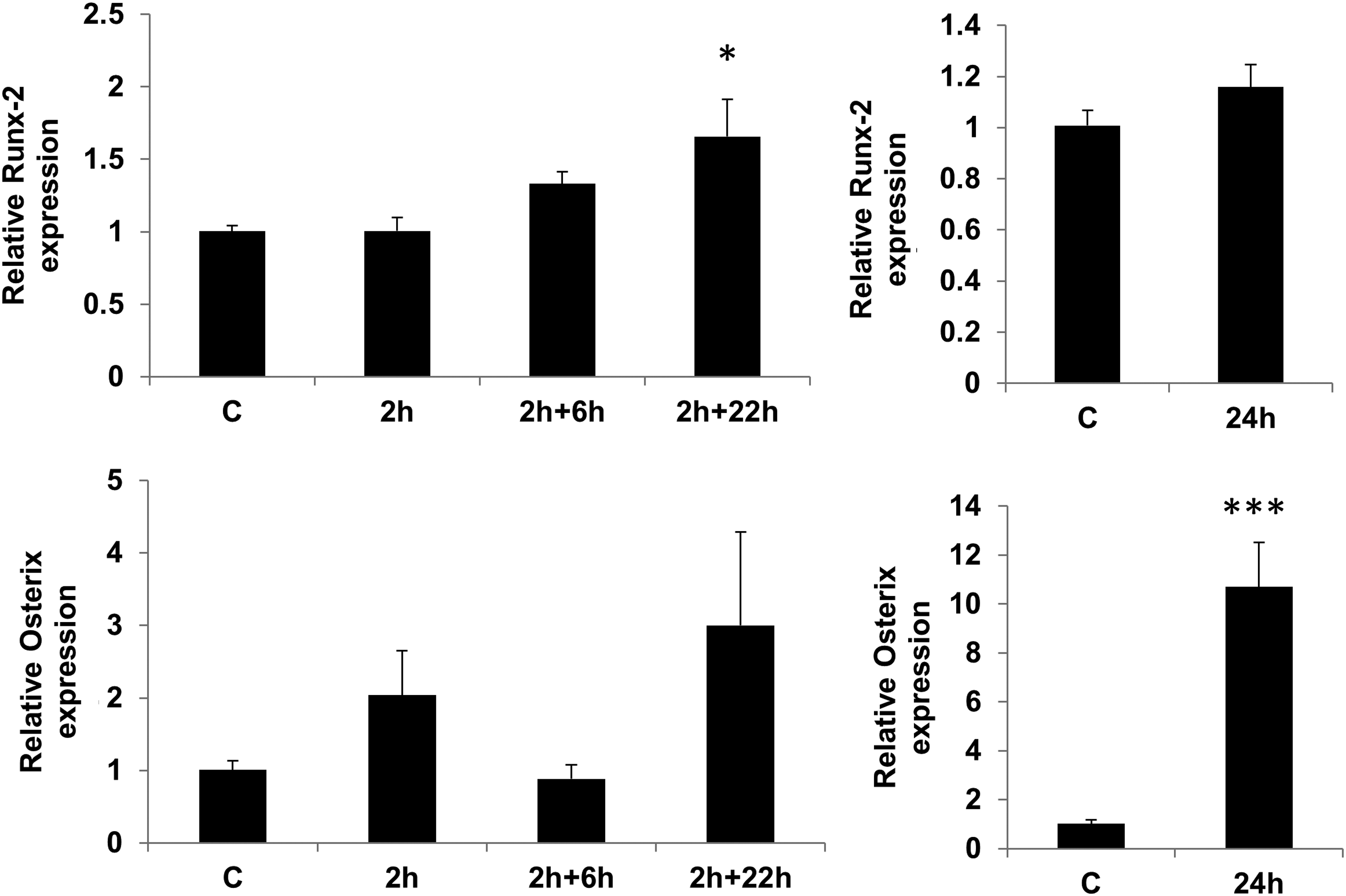
Effect of mechanical stimulation on transcriptional regulators of osteogenesis. Cells were loaded with 0.4% strain and analyzed after 2 h of loading, following 6 or 22 h rest post 2 h loading (2+6 and 2+22) and following 24 h of constant loading for expression of osteogenic (Runx-2 and osterix) transcription factors. Data (mean±SEM) from three experiments in duplicates. *p<0.05 and **p<0.01 when compared with unloaded at the same time point after normalization to RPL13a.
To determine the outcome of a longer period of mechanical stimulation, we loaded the cells continuously with the same strain for 24 h and compared the changes in expression of selected genes from microarray studies. None of the selected genes that were influenced by 2 h of loading were affected in the same manner with either 24 h continuous loading or following 22 h of resting after the load (Figs. 4 and 5). However, continuous loading for 24 h resulted in a dramatic increase in expression of osterix (Fig. 6).
Effect of loading on ERK and AKT pathways
To investigate the signal transduction pathways involved, we assessed the alteration in phosphorylation of Akt and P44/P42 MAPK in response to stimulation with mechanical loading. Serum starved MSCs were loaded for 15 min, 2 and 24 h and protein phosphorylation was determined using phosphor-specific antibodies against Akt (Ser473) and p44/42 MAPK (Erk1/2) (Thr202/Tyr204). The Erk1/2 were phosphorylated within 15 min and the level remained consistently high for the duration of 2 h and even further induced by 24 h of stimulation. In contrast there was no change in Akt phosphorylation (Fig. 7A).
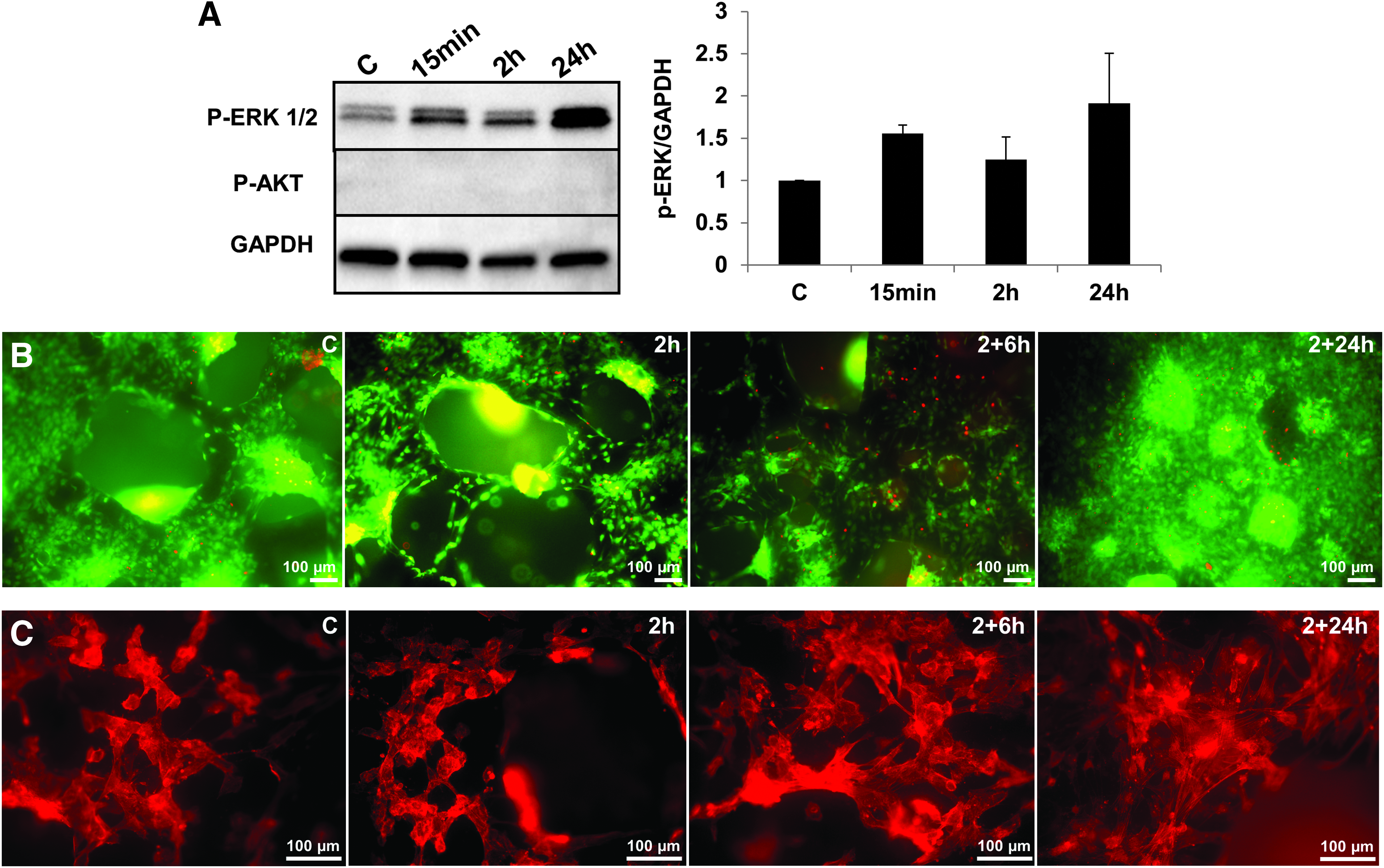
Signal pathways regulated by mechanical loading in MSCs.
Effect of loading on MSC morphology and viability
Microscopic analysis (Fig. 7B) showed a homogeneous monolayer of MSCs covering the scaffold surface. Almost all the cells were alive and only a few dead cells were detected in the unloaded control and following 2 h of loading. However, the number of dead cells slightly increased when cell were rested for 6 and 22 h following the 2-h of loading. As Figure 7C shows the cytoskeletal organization of the cells displayed alterations, with MSCs taking a more rounded shape when loaded for 2 h, while cells appeared more flattened with a more prominent filamentous actin network when rested for 22 h.
Discussion
The role of mechanical loading in skeletal tissue homeostasis has received considerable attention in the recent past. Osteoblasts and chondrocytes have been the main focus of many of these studies, but it is also evident that MSCs are responsive to mechanical loading. A number of studies have been conducted to study the mechanisms involved in mechanically induced MSC differentiation to osteoblasts in vitro. However, many of these studies have used loads that substantially exceed the physiological or even pathological equivalent loads in vivo and have not been able to closely mimic the bone environment.13,17,18,32 It is well known that surface stiffness and composition is a key regulator of MSC behavior, with stiff substrates with bone-like properties shown to support osteogenesis.33,34 Strain measurements in human bones physiologically indicate a strain magnitude of between 0.2% and 0.4%. 23 Biological responses to this loading have been reported following activities of more than 30 min duration and particularly following high intensity exercise of longer periods (>2 h) such as marathon running. 23 Consequently, in the present experiments we have used a CaP scaffold to mimic the biomechanical properties of bone and applied a carefully controlled load to study the mechanisms of mechanical loading in inducing bone formation using a strain and duration that may closely mimic physiological conditions and has been shown to initiate early biological responses in vivo.
Dramatic alterations in cell structure and shape with excess strain may bring about a large body of nonspecific changes in pathways other than those involved in differentiation or proliferation such as cell stress and inflammatory responses. In fact, previous gene expression studies of cells subjected to large mechanical loads have reported regulation of stress and inflammatory genes.15,35,36 In comparison, there were no signs of upregulation of inflammatory or stress-related genes in the present study, supporting the suggestion of an appropriate level of strain being utilized. Our study also showed a markedly lower total number of changes in gene expression in response to a mechanical loading. A total of 100 genes were differentially expressed after 2 h stimulation and 63 genes when cells were subsequently rested for 22 h. In contrast, others have reported between 1540, 20 5449, 37 400–6000 36 861 38 differentially expressed genes in response to mechanical stimulation depending on the duration of induction in osteoblasts or MSCs. This large number of changes might cast doubt on the specificity of these effects and makes it difficult to draw conclusions on the mechanisms involved.
The limited number of changes in this study allowed us to divide the differentially expressed genes into only a few clusters associated with their biological processes. The largest cluster of genes activated with 2 h stimulation mainly belonged to regulators of transcription. Among these, FOSB has been shown to enhance bone formation in vivo and in vitro, and its expression is enhanced in a differentiation-related manner and in response to mechanical stimuli in mouse osteoblasts.39,40 These findings imply that FOSB might be a mechanoresponsive gene that activates the osteogenic program in osteoblasts in response to mechanical stress. Recent in vivo expression analysis of mouse osteoblasts also indicates an increase in members of the FOS family early in loading. 22 Our data concur with these findings and show that responses are not limited to mouse osteoblasts but also occur in human MSC. In addition to FOSB, which has also been associated with osteoblast proliferation, a group of genes was identified that function in the regulation of the cell cycle and proliferation. These included SERTAD-1, CDKN1b, GMNN, WDR37, WDR43, TP53RK, and SNIP-1. The changes in expression of these genes are likely to result in regulation of early cell cycle processes. Members of the MAPK family such as Erk1/2 are involved in cell cycle progression and also implicated as an important mechanically induced signal pathway involved in osteogenesis.7,41 Our data also showed upregulation of Erk1/2 phosphorylation with loading and we identified a number of possible genes (SPRY2, RIPK1, SPRED2, SERTAD1, TRIB1, and RAPGEF2) that may regulate this process. In particular, upregulation of Sprouty (SPRY)2 is interesting as this gene and other members of the Sprouty family have been extensively studied as negative regulators of FGF-2/MAPK signaling.42,43
Surprisingly, when cells were rested for 6 h after mechanical stimulation, gene expression returned to normal, although after further resting for a total of 22 h upregulation of a totally new group of genes was seen, which were mainly involved in cell surface receptor signal transduction and regulation of metabolic and cell division processes. The absence of any significant changes at 6 h may reflect the fact that gene expression profiles are dynamic, and the data overall suggest that 2 h mechanical loading induces an initial wave of expression of immediate-early response genes, which cascade to induce a second wave of later response genes many hours after the initial responses seen. These data are consistent with the biphasic effects of prostaglandin inhibition on bone following mechanical loading previously reported by Chow and Chambers. 44 However, additional studies would be useful to clarify this and to study further the possibility of gene responses at other time points within or after the times described here.
To study whether changes in expression of transcription factors and protein kinase cascades regulators could be translated to physiological changes, we also analyzed the expression of some key genes involved in osteogenic differentiation. As might be predicted, 2 h was not long enough to alter the expression of Runx-2 or osterix transcription factors. However, when cells were left for a further 22 h a significant increase in expression of Runx-2 was seen by qRT-PCR. Although the change in Runx-2 expression in the microarray was not statistically significant, it followed a similar trend in 22 h rested cells in comparison to unloaded samples. In general, there was a good correlation in trend of expression between microarray and qRT-PCR data; however, there were differences in level of expression and statistical significance that could be attributed to variation in detection method, data normalization, and analysis.
The transforming growth factor β (TGF-β) signaling pathway has also been shown to regulate bone formation in response to mechanical loading. Members of the TGF-β family, particularly TGF-β1 and -β3, have been shown to be differentially expressed and are involved in various stages of bone formation in response to loading.22,45,46 In the current studies, TGF-β receptor 3 upregulation was seen as an early response to loading while TGF-β3 itself was downregulated following a 22-h resting period. Members of other growth factor families (PDGFC and FGF7), chemotactic factors (CCL2 and CCL20) and members of the collagen family were among other genes previously implicated in mechanical loading gene expression studies, which were also identified as being differentially regulated in our study.22,38,46
In summary, we have used an in vitro model based on a CaP scaffold and induced mechanical loads relevant to physiological conditions to mimic in vivo mechanical stimulation responses in MSCs. We have shown that the early stage of mechanical stimulation is associated with upregulation of a group of transcription factors including FOSB, a regulator of bone cell proliferation and differentiation. Other genes involved in regulation of MAPK signal transduction cascade and cell cycle progression were also induced. Later responses to loading were very distinct, revealing new changes in expression patterns of genes involved in cell surface receptor signaling and metabolic processes and osteogenic transcription factors. These data may provide a valuable insight into the early molecular mechanisms and intracellular signal transduction pathways involved in the regulation of MSC osteogenic differentiation in response to a physiological level of mechanical stimulation. In addition, our data show for the first time the use of mechanical load in a bioreactor for induction of MSC proliferation and differentiation on porous BCP scaffolds. This could be used as a bone-like model for future study of mechanical loading or as a novel method for preparation of ex vivo implants to use in tissue engineering approaches for treatment of bone defects.
Footnotes
Acknowledgments
This work was supported by research funding from the Guy's and St Thomas Charity. The authors acknowledge financial support from the Department of Health via the National Institute for Health Research (NIHR) comprehensive Biomedical Research Centre award to Guy's and St Thomas' NHS Foundation Trust in partnership with King's College London and King's College Hospital NHS Foundation Trust. We also acknowledge support from the Wellcome/EPSRC Centre of Excellence in Medical Engineering.
Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.