
Abstract
Aim:
Despite our previous study that demonstrates that human embryonic stem cells (hESCs) can be used as seed cells for tendon tissue engineering after stepwise induction, suboptimal tendon regeneration implies that a new strategy needs to be developed for tendon repair. We investigated whether overexpression of the tendon-specific transcription factor scleraxis (SCX) in hESC-derived mesenchymal stem cells (hESC-MSCs) together with knitted silk-collagen sponge scaffold could promote tendon regeneration.
Methods and Results:
hESCs were initially differentiated into MSCs and then engineered with scleraxis (SCX+hESC-MSCs). Engineered tendons were constructed with SCX+hESC-MSCs and a knitted silk-collagen sponge scaffold and then mechanical stress was applied. SCX elevated tendon gene expression in hESC-MSCs and concomitantly attenuated their adipogenic and chondrogenic potential. Mechanical stress further augmented the expression of tendon-specific genes in SCX+hESC-MSC-engineered tendon. Moreover, in vivo mechanical stimulation promoted the alignment of cells and increased the diameter of collagen fibers after ectopic transplantation. In the in vivo tendon repair model, the SCX+hESC-MSC-engineered tendon enhanced the regeneration process as shown by histological scores and superior mechanical performance compared with control cells, especially at early stages.
Conclusion:
Our study offers new evidence concerning the roles of SCX in tendon differentiation and regeneration. We demonstrated a novel strategy of combining hESCs, genetic engineering, and tissue-engineering principles for tendon regeneration, which are important for the future application of hESCs and silk scaffolds for tendon repair.
Introduction
T
It is well-known that fetal tendons possess high regenerative potential. 22 Interestingly, this capacity is intrinsic to the fetal tendon itself. 22 Hence, the cells may in fact be crucial for the regenerative potential of tendons. Embryonic stem cells (ESCs), which are primary embryonic cells, offer great promise for tissue regeneration. As demonstrated in our previous work, ESCs can be used for tendon tissue engineering by stepwise induction. 20 However, complete maturation of tendon-like tissues and complete regeneration of injured tendons have not yet been achieved due to the lack of strategies to specifically drive the differentiation of human ESCs (hESCs) into a tenocyte fate.
Scleraxis (SCX), a basic helix-loop-helix transcription factor, is the earliest known marker of tendon and the somatic compartment of tendon progenitors (syndetome).23–25 It may be involved in the upstream regulation of other genes that are strongly expressed in tendon tissues, such as collagen I, collagen XIV, tenomodulin (Tnmd), and fibroblast growth factor 8.26,27 Moreover, SCX gene knockout has been shown to result in severe defects in force-transmitting tendons. 26 Further, overexpression of Scx induces teno-lineage differentiation of bone marrow MSCs 27 and improves rotator cuff tendon–bone junction healing.28,29 These results suggest that SCX plays a critical role in tendon differentiation and regeneration.
The scaffold is another key component in tendon tissue engineering by providing a three-dimensional (3D) environment for cell implantation and tissue regeneration. We have previously fabricated a composite scaffold of knitted silk and collagen matrix,30–32 which meets tendon scaffold standards, including biodegradability, biocompatibility, providing mechanical support, and promoting tissue formation. The knitted silk scaffold provides sufficient mechanical strength, while the collagen sponge encourages the ingrowth of neotissues and enhances neoligament regeneration by promoting extracellular matrix (ECM) gene expression and collagen fibril assembly. 30 In our previous study, we showed that the engineered tendon constructed from hESC-MSCs and the knitted silk-collagen scaffold improves Achilles tendon regeneration. However, the structure and function of the repaired tendon were still much inferior to those of normal tendon. In this study, we evaluated the hypothesis that tissue-engineered tendon constructed by combining hESC-MSCs genetically engineered with SCX (SCX +hESC-MSCs) and knitted-silk collagen sponge scaffold improves functional tendon regeneration. Our results demonstrated that SCX enhanced the teno-lineage differentiation of hESC-MSCs and promoted tendon regeneration in vivo.
Materials and Methods
Cell culture
hESC-MSCs were obtained as in our previous study and the protocol was approved by the Zhejiang University Ethics Committee (No. ZJU2010102003). 20 Briefly, to derive hESC-MSCs, hESCs were trypsinized, centrifuged, and resuspended in knockout Dulbecco's modified Eagle's medium (Gibco, www.invitrogen.com) supplemented with 10% (v/v) fetal bovine serum (FBS; Invitrogen) and 5 ng/mL fibroblast growth factor 2 (Gibco-BRL, Inc.). Upon reaching confluence, the cells were trypsinized and replated at a density of 102 to 103 cells/cm2 in the same culture medium. Cells between passages 6 and 9 were used.
SCX transfection
The human SCX gene (NM_001080514.1) was synthesized by Jin-Si-Te Co. SCX was subcloned into the pleti-Gateway vector for constructing lentivirus (Invitrogen, Inc.). The packaging cell line Hek293T was transfected with 5 μg of control lentiviral LacZ/GFP-gateway plasmid, SCX gateway, and 12 μg of package mix-plasmid, and 1.5 mL of DMEM without supplements. The DNA mixture with 45 μL Lipofectamine 2000 (Invitrogen Corp., www.invitrogen.com) was added to the Hek293FT cells and then incubated for 12 h. The cells were then cultured for 48 h and the harvested supernatant containing infectious particles was filtered through 0.45-μm filters (Millipore), prior to being added to semiconfluent cultures of hESC-MSCs (1×105 cells/6 wells in DMEM with 10% FBS) at passages 7–9 for 4 to 24 h. SCX hESC-MSC transfer was obtained by transfection with SCX-lentivirus followed by selection with blasticidin (2 μg/mL) for 2 weeks. The lacZ genes were used as controls. The cells were assayed for recombinant expression of genes by quantitative PCR. Cells that demonstrated a high level of transgene expression were propagated and frozen. Selected cells were expanded in the presence of blasticidin and the selection pressure was maintained during subsequent manipulations; multiclonal cells were used in this study. Transduced cells at passages 3 and 5 after selection were used for functional studies.
Immunohistochemical analysis
Cells were fixed in 4% formalin for 10 min, blocked in phosphate-buffered saline (PBS)/1% Tween-20/1% bovine serum, and incubated at room temperature for 1 h. After incubation with the primary antibody (rabbit anti-SCX polyclonal antibody, Ab58655; Abcam) the slides were rinsed in PBS, and a secondary Alexa Fluor 546 antibody (goat anti-rabbit antibody secondary, A21430; Lifetechnology), diluted 1:100 in PBS, was applied to the slides at room temperature for 30 min. After the slides had been washed with PBS (twice for 5 min), they were mounted with 50% glycogen for confocal microscopy imaging.
Fabrication of hESC-MSC-engineered tendon
To construct engineered tendon for in vivo repair, scaffolds (1×1 cm2; thickness, 0.2 cm) were seeded with SCX- or GFP-transfected hESC-MSCs (3×105 cells/scaffold). The cell-seeded scaffolds were cultured in a 5% CO2 incubator at 37°C for 24 h. Cellular proliferation was quantified on days 1, 4, 7, and 10. Scanning electron microscopy (SEM) imaging of cellular morphology was carried out on days 1, 4, and 14. For dynamic mechanical stimulation in vitro, the hESC-MSC-seeded scaffolds (4×0.5 cm2; thickness: 0.2 cm; see “Supplementary Materials and Methods” section [Supplementary Data are available online at www.liebertpub.com/tea]; 6×105 cells/scaffold) were placed in the chambers of a custom-made bioreactor, 31 with the two ends anchored and stretched at 10% elongation. The frequency of the bioreactor was set at 1 Hz. The control group was cultured in another chamber in the same bioreactor, but was not subjected to mechanical stimulation. The transcript levels of various gene markers (tendon genes: collagen Ia1, collagen Ia2, collagen XIV, Tnmd, and Eya2; bone gene: Ocn) in both groups were determined by RNA extraction and quantitative polymerase chain reaction (qPCR) at 4 and 24 h.
Ectopic implantation of tissue-engineered tendon
To determine the potential of ectopic tendon formation with the tissue-engineered construct under mechanical loading in vivo, a nude mouse model was used as previously reported. 31 The Zhejiang University Animal Care and Use Committee approved the study protocol (ZJU2010102003). After anesthesia, an incision was made on the dorsum and a 2-cm subcutaneous pocket was used to expose the fascia nuchae on the dorsonuchal area and the fascia of the musculi sacrospinalis in the sacral area. The entire length of the tissue-engineered tendon was 5 cm. To provide mechanical loading in vivo, part of the tissue-engineered tendon (3 cm) was sutured to the fascia nuchae cranially at the level of the tenth thoracic vertebra and to the fascia of the musculi sacrospinalis at the level of the second sacral vertebra. This enabled natural movement of the back to provide cyclic mechanical loading on the tissue-engineered tendon. For the control group, the engineered tendon (2 cm) was implanted within the subcutaneous pocket without suturing. Four weeks after implantation, the animals were subjected to euthanasia, and the implanted tissues were harvested for various analyses.
Rat Achilles tendon repair model
To assess the in vivo regeneration potential of the engineered tendon, an Achilles tendon gap wound model was used; full-thickness, 6-mm-long segment of the Achilles tendon in rat is cut to form a critical gap. Tendons were cut transversely 1.5–7.5 mm proximally to its calcaneal insertion (Supplementary Fig. S5). Scaffolds (1×1 cm2; thickness, 0.2 cm) seeded with SCX-transfected hESC-MSCs (3×105 cells/scaffold) were sutured to the remaining tendon using a nonresorbable suture (6-0 nylon). Treatment of animals was in accordance with the standard guidelines and approved by the Zhejiang University Ethics Committee (ZJU2010102003). After 2 and 4 weeks, the repaired tendon was evaluated. Sprague-Dawley rats weighing 200–220 g were given general anesthesia. A portion of the Achilles tendon on the left limb was removed to create a 6-mm-long defect and the control-engineered tendon. Tendons were cut transversely 1.5–7.5 mm proximally to its calcaneal insertion. hESC-MSCs seeded on knitted silk-collagen sponge was implanted into the defect (control group, n=14). On the right limb, the composite scaffold (SCX+hESC-MSCs seeded on knitted silk-collagen sponge) was implanted to repair the defect (SCX+group, n=14). After surgery, the animals were allowed free cage activity. Upon sacrifice, three limbs from each experimental group were used for histological evaluation and gene expression analysis at the 2- and 4-week time-points. Five limbs were used for mechanical testing, and three randomly selected ATs were imaged by transmission electron microscopy (TEM) 4 weeks after surgery.
Multipotent differentiation, cell labeling and detection, TEM, qPCR, histological examination, and Masson's trichrome staining are described in Supplementary Materials and Methods section 20 ).
Statistical analysis
All data are expressed as mean±SD, unless otherwise indicated. The t-test was performed between two groups. The interaction effect of mechanical stress and SCX on tendon differentiation in vitro was analyzed using factorial design analysis-of-variance. Values of p<0.05 were deemed to be statistically significant.
Results
SCX modulates mesodermal differentiation
MSCs were derived from hESCs as previously described, 20 and displayed low expression levels of SCX (Fig. 1A, B). Subsequently, hESC-MSCs were infected with lentiviral vectors encoding SCX or control genes. After blasticidin selection for 2 weeks, clonal hESC-MSCs stably expressing SCX were established (termed SCX+hESC-MSCs) (Figs. 1A and 4A). Immunofluorescent staining with anti-SCX antibody revealed that SCX was highly expressed in the transduced hESC-MSCs after 2 weeks (Fig. 1B). Western blotting confirmed the overexpression of SCX in hESC-MSCs after infection (data not shown). Next, we evaluated whether SCX overexpression autonomously initiates tenogenesis in hESC-MSCs. We selected at least three clonal lineages of SCX-hESC-MSCs and control cells, and found that recombinant SCX increased the expression of collagens I and XIV (Fig. 1C) but not other tendon-related genes (COL III, BGN, SIX1, and NFATC4; data not shown). Moreover, SCX enhanced cell-sheet formation, indicating more collagen deposition by the cells (Fig. 1D). During this procedure, cells did not express oct4 and telomerase, but did express rex1 and nanog (Supplementary Fig. S1).
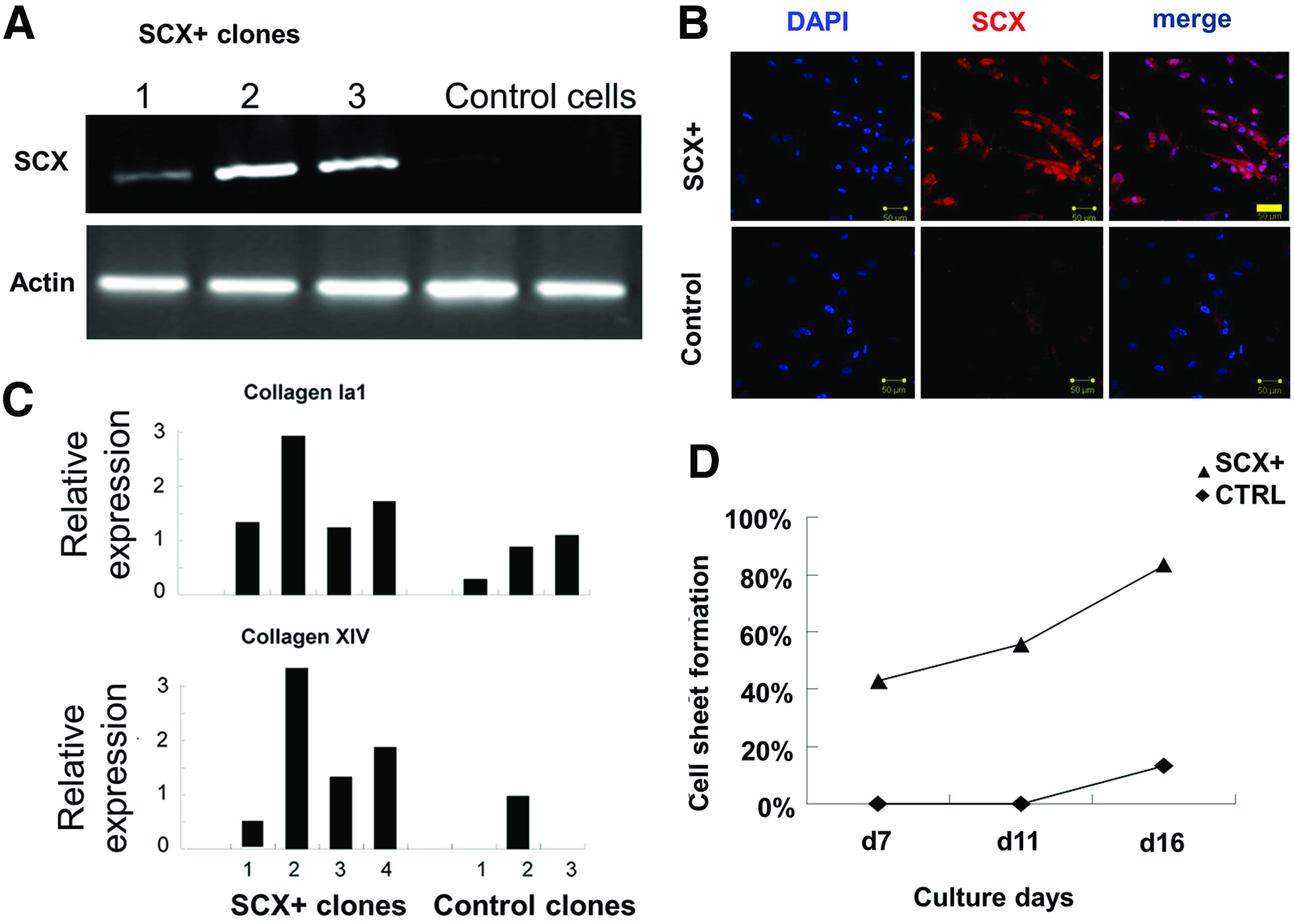
Recombined SCX overexpression in hESC-MSCs enhanced collagen expression.
To evaluate the function of SCX in differentiation into other mesodermal cell types, we examined the adipogenic and chondrogenic potential of SCX+hESC-MSCs. For adipogenesis, the SCX+hESC-MSC lines were induced under adipogenic conditions in vitro. Multicolony-derived SCX+hESC-MSCs demonstrated a decreased capacity to develop Oil red O-positive lipid-laden adipocytes in comparison with control hESC-MSCs (Fig. 2A). Quantitative analysis of the level of lipid production confirmed that the SCX+hESC-MSCs synthesized lower levels of triglycerides than the control cells (p<0.05, Fig. 2B). For chondrogenesis evaluation, hESC-MSCs were cotransfected with a reporter gene driven by the collagen II promoter, which is the main ECM marker of chondrogenesis. We found that SCX inhibited the collagen II promoter activation (Fig. 2C, D and Supplementary Fig. S2), suggesting that SCX plays an inhibitory role in chondrogenesis.
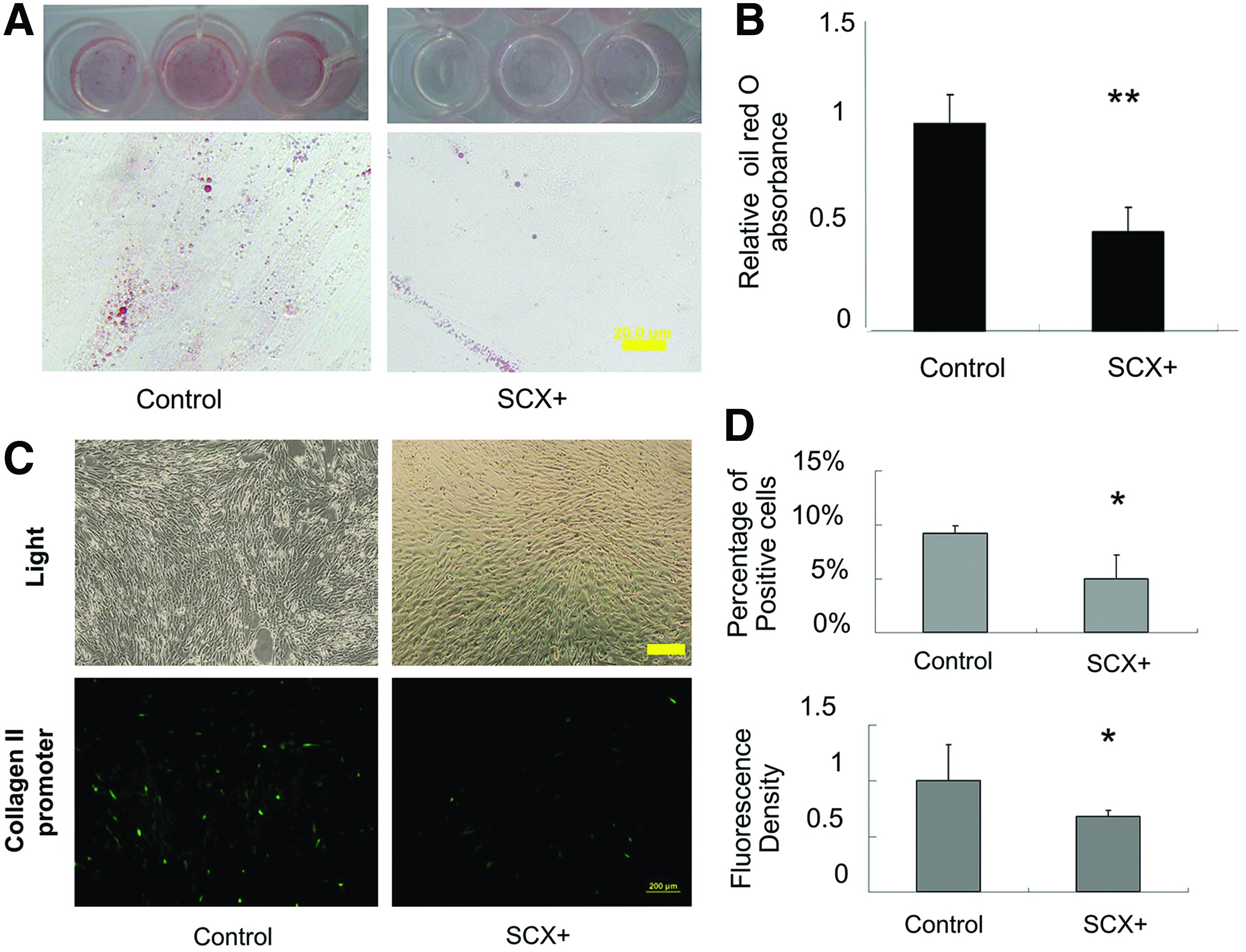
Effects of SCX on hESC-MSC differentiation.
Construction of engineered tendon with SCX+hESC-MSCs and silk-collagen scaffold in vitro
SEM imaging of hESC-MSC morphology on the composite scaffolds showed cell attachment, spreading, and proliferation during a period of 14 days. After 3 days of culture, the engineered tendon showed considerable numbers of viable cells on the scaffolds (Fig. 3A). The cells displayed good proliferation and adhered to the scaffolds after 14 days of culture, as well as exhibiting some possible ECM formation (Fig. 3A). The MTS study showed that SCX overexpression did not impair the proliferation rate of the cells on the scaffold (Fig. 3B). Then, we investigated whether SCX reduced osteogenesis by the cells on the scaffold. After 14 days in osteogenic culture, Alizarin red S staining (Supplementary Figs. S2 and S3) and quantitative analysis showed that SCX+hESC-MSCs had less calcium nodule deposition (Fig. 3C), indicating that the SCX+hESC-MSC-engineered tendon had less bone differentiation potential.

Fabrication of engineered tendon that combination of SCX+hESC-MSCs and knitted silk-collagen sponge.
To assess the function of SCX in the engineered tendon under mechanical stress in vitro, we applied mechanical stress at 10% strain and 1 Hz. 31 The expression of tendon-related ECM genes, transcription factors, and surface markers was analyzed as tendon differentiation markers (collagen Ia1, collagen Ia2, collagen XIV, 33 Tnmd, 34 and Eya2). After 4 h, 24 h, or 7 days of mechanical stress on hESC-MSCs, the ECM genes collagen Ia1 and collagen Ia2 were upregulated, as well as the tendon-related transcription factor Eya2, indicating that mechanical stress initiates tendon differentiation. In the in-vitro-engineered tendon model, mechanical stimulation led to increased collagen XIV and Tnmd expression only in SCX+hESC-MSCs and not in control cells (Fig. 4A and Supplementary Fig. S4). Moreover, the ECM genes decorin and collagen III were downregulated (Fig. 4B and Supplementary Fig. S4). As these two genes were highly expressed in the repaired tendon compared with normal and impaired collagen fibrillogenesis, the downregulation of these genes in SCX-hESC-MSCs indicated a higher regeneration potential. And the osteogenic marker osteocalcin was also significantly inhibited by SCX (Fig. 4C and Supplementary Fig. S4). These data therefore indicated that SCX+hESC-MSCs have more potential for tendon differentiation and maturation in engineered tendon.
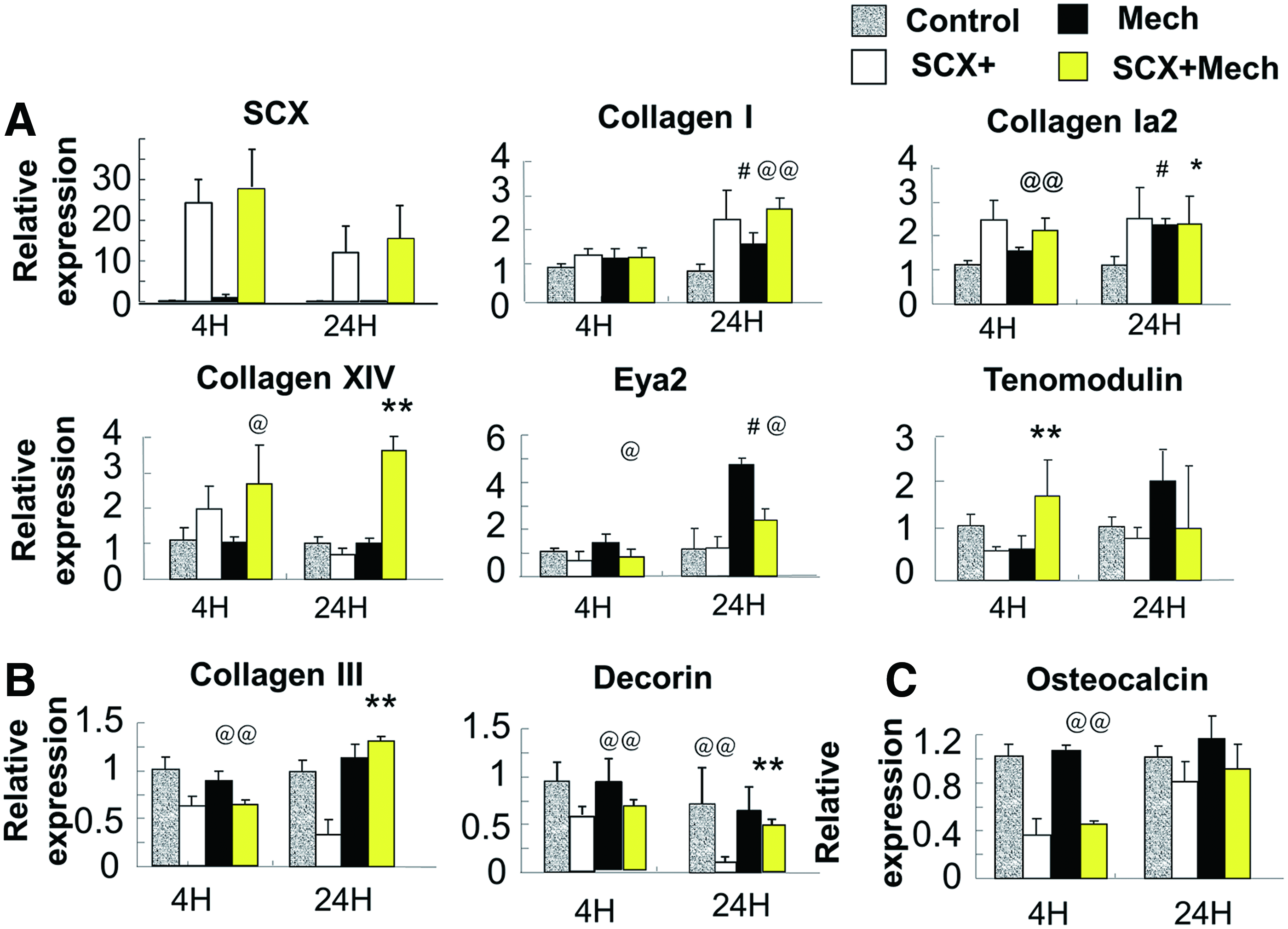
Expression of tendon-specific and related matrix genes in engineered tendon scaffolds after 4 and 24 h.
In vivo ectopic implantation of tissue-engineered tendon
In the in vivo ectopic implantation model, engineered tendons were implanted subcutaneously into SCID mice. The engineered tendons were given natural dynamic mechanical stimulation by the body movements of the mice. Cell tracking showed that implanted engineered tendons were present subcutaneously within mice for at least 4 weeks, as previously reported 31 (data not shown). Four weeks later, the HE and Masson trichrome staining results revealed a mass of dense tissue consisting of bands of collagen fibers formed in the SCX+hESC-MSC-engineered tendon under in vivo mechanical stress (Fig. 5A). In addition, in order to evaluate the maturation level of collagen, the ultrastructure of the collagen was examined. Although the collagen fibers were still relatively thin and uniformly small, TEM imaging showed that the SCX+hESC-MSC-engineered tendon had significantly more mature collagen and larger fiber diameters (Fig. 5B; 38.5±6.4 nm vs. 24.6±2.7 nm, p<0.05). These data thus indicated that SCX+hESC-MSCs successfully form engineered tendon with a silk-collagen scaffold in vivo.
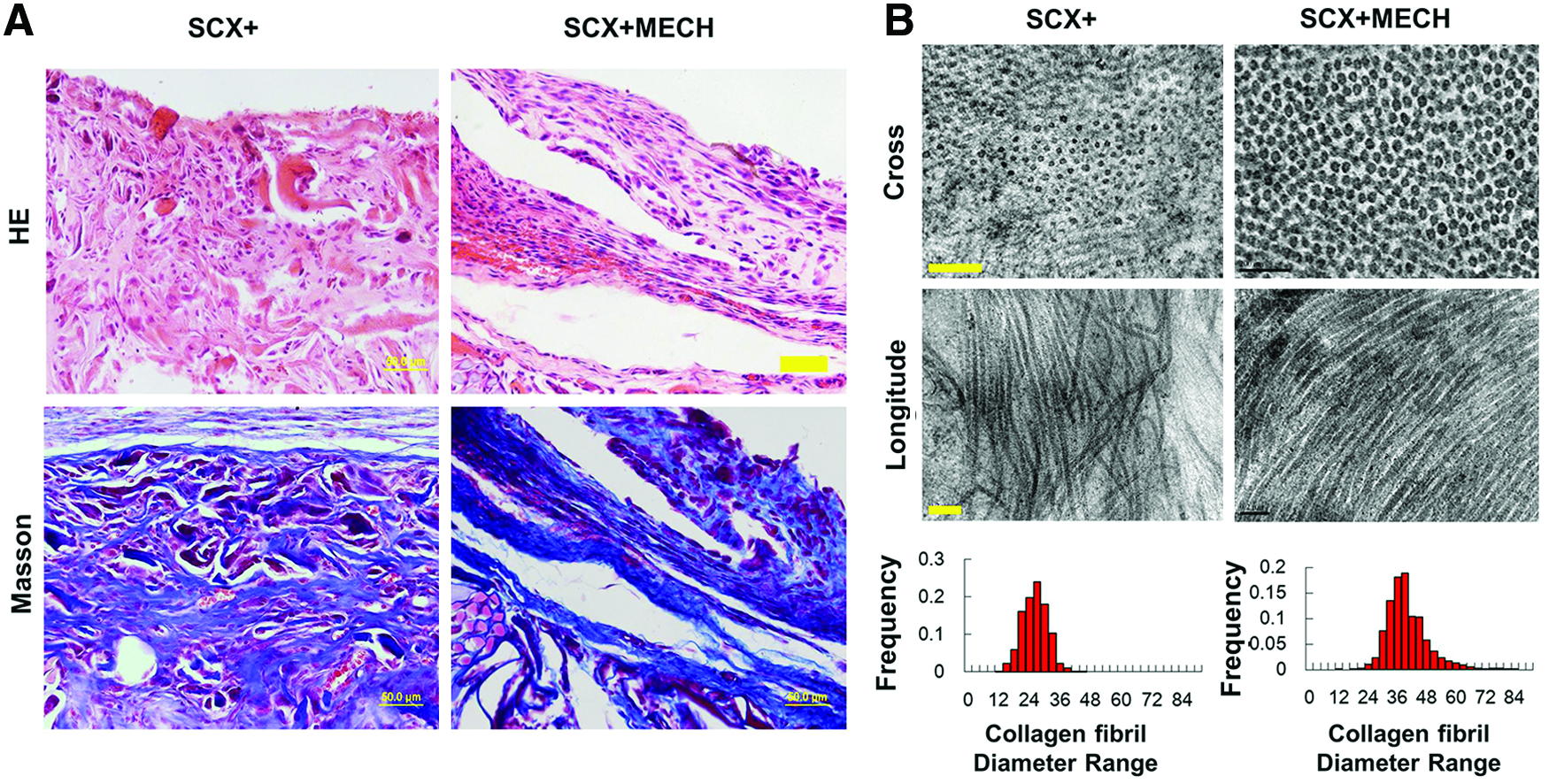
Histology and ultrastructure of tissue-engineered tendons 4 weeks after implantation in vivo.
SCX+hESC-MSCs increase the regeneration of rat Achilles tendon
To further evaluate the tendon regeneration potential of engineered tendons in vivo, an Achilles tendon defect model was used. hESC-MSCs transfected with GFP were used as control. Upon gross observation, all tendons had hypergrowth 2 and 4 weeks after surgery, and the injury was repaired with connective tissue, without inflammation or scaffold dislocation. The neo-tendons in both groups were easily distinguishable from native tendon as the repair site showed an amber color. The SCX+ group had more compact neo-tendon formation.
The survival of implanted cells in vivo was monitored using a noninvasive CCD tracking system and fluorescence. Image tracking showed positive DiI labeling at the repair site 2 weeks after surgery and the presence of DiI-labeled cells was confirmed by fluorescence microscopy (Supplementary Fig. S6). 31 Imaging with fluorescence microscopy confirmed that the DiI-labeled cells contributed to tendon regeneration at the injury site.
Histology of repaired tendon
After 2 weeks, the histology showed that more collagen fibers had been deposited in the SCX+ group than in the control group. A more mature morphology (based on color and continuity) was observed in the SCX+ group. The SCX+ group showed that denser connective tissue filled and wrapped the scaffold (Fig. 6A). Masson trichrome staining showed that more dense collagen fibers and continuous tissue at the junction site were formed in the SCX group than in the control group (Fig. 6A). The histology score also confirmed the histology results (8.32±0.83 vs. 11±0.44, p<0.05; Fig. 7A). This indicated that SCX+ cells induced more fibroblast-like cells and fewer inflammatory cells in the repair site than in the control group (fibroblast ratio: 77.8%±4.2% vs. 71.3%±3.7%, p<0.05; Fig. 7B), which enhanced tissue regeneration at an early stage. The sections were also observed under polarized light microscopy to evaluate collagen fiber maturation. The SCX+ group showed more continuous collagen fibers in the repair site (Fig. 6A).
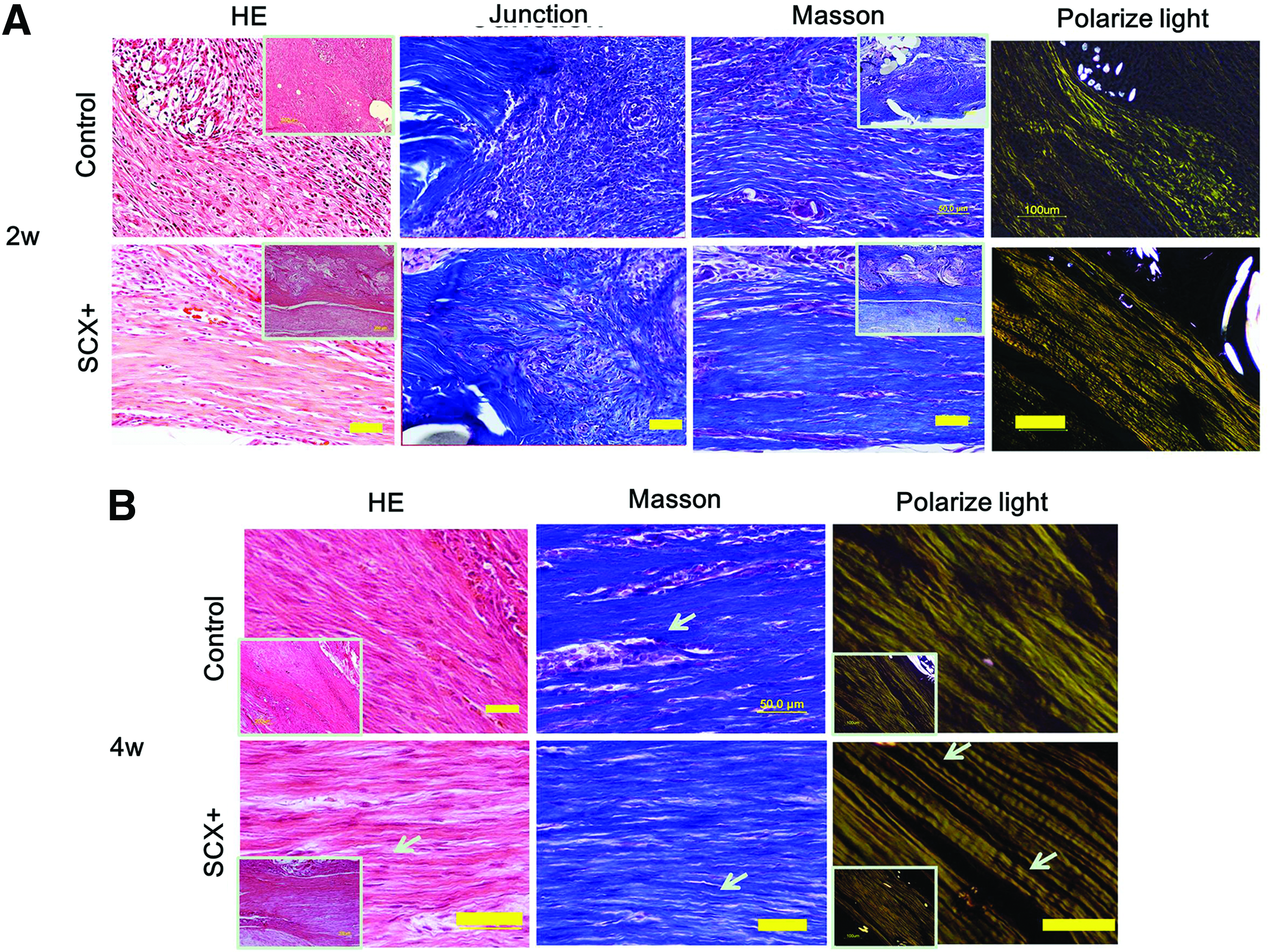
HE, Masson staining, and polarized light microscopy images from regenerated tendon 2 and 4 weeks after surgery.
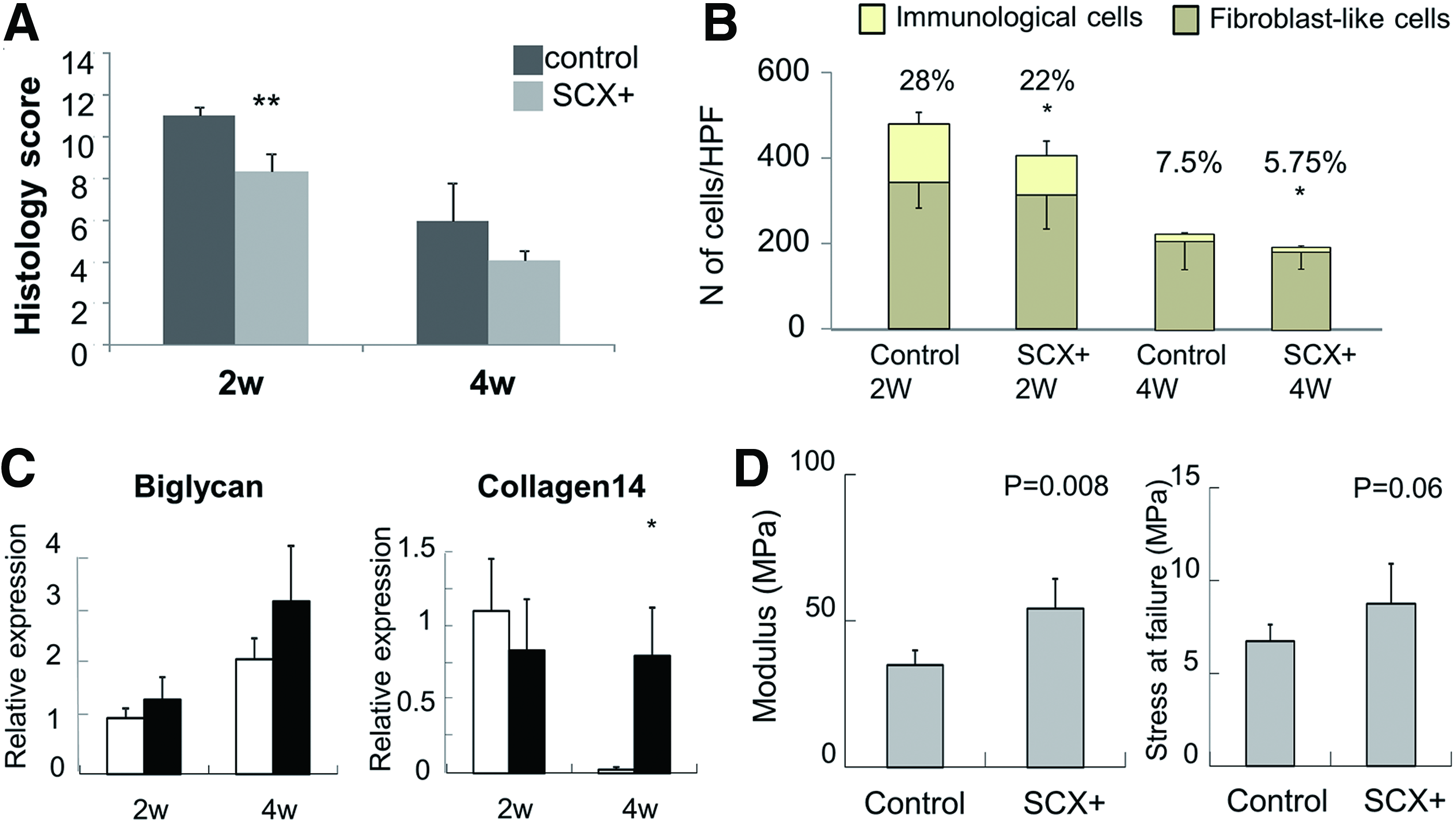
Gene expression and mechanical properties of the repaired tendon.
After 4 weeks, the histological appearance of the neo-tendons was more like native tendon with a reduced cellular component and more bands of dense collagen fibers filling in the repaired tissue of both groups. More crimped collagen fibers were found in the SCX+ group, and more cells exhibited spindle-shaped morphology along the axis of tensile load (Fig. 6B). Moreover, there were fewer immunological cells (immunological cell ratio: 5.8%±1.1% vs. 7.5%±0.7%, p<0.05; Fig. 7B), a lower total cell number (cells per high-power field: 172±38 vs. 195±62), and more fibroblast-like cells in the SCX+ group than in the control group. We also found host immune cell infiltration and a few vascular components in all groups at 2 weeks postsurgery, while these were reduced at 4 weeks. Consistent with the histology and cell counts, the histology score showed that the SCX+ group was more similar to the native tendon (4.07±0.48 vs. 5.94±1.8231), but the difference was not significant. This may due to the relatively small sample size (n=3).
Gene expression of repaired tendon
Tendon-related ECM gene expression in repaired Achilles tendons was analyzed by qPCR (n=3). The expression levels of collagen XIV were similar in the two groups after 2 weeks, but were much higher in the SCX+ group at 4 weeks. Biglycan expression in the SCX+ group was higher than in the control group. The higher expression of the collagen XIV and biglycan genes in the SCX+ group indicated that SCX-hESC-MSCs influence endogenous collagen maturation (Fig. 7C).
Mechanical properties of repaired tendons
To further correlate structural features of the tissues with their mechanical properties, harvested tendons (n=5 for each group) were subjected to mechanical testing. As shown in a previous study, 31 repaired tendons also exhibited a toe region for up to ∼5% of elongation in the typical load–elongation curves of healed tendons. Both groups had better mechanical properties than noncell controls. At 4 weeks, the modulus (60.63±17.6 MPa vs. 48.4±13.6 MPa, 31 p<0.05) and stress at failure (8.73±2.15 MPa vs. 6.72±0.9 MPa) in the SCX group showed better results than the control group (Fig. 7D). At 8 weeks, the modulus (71.3±12.4 MPa vs. 53.1±16.9 MPa, p<0.05) and stress at failure (10.1±2.2 MPa vs. 7.9±1.8 MPa) in the SCX group also showed better results than the control group (Supplementary Fig. S7A, B). However, the mechanical characteristics of both groups were still much lower than normal tissues. The gene expression, histology, and mechanical properties indicated that SCX enhances the structural quality of neo-tendon formation, especially in the early stage.
Discussion
In this study, we demonstrated a novel strategy for functional tendon tissue engineering by incorporating SCX+hESC-MSCs within a knitted silk-collagen sponge scaffold under an inductive environment (i.e., dynamic mechanical stimulation). In particular, our results suggested that (1) transcription factor SCX expression is essential for initiating tendon differentiation and inhibiting other lineage differentiation. (2) SCX+hESC-MSCs can form engineered tendon by combining with knitted silk-collagen sponge scaffolds. Mechanical stress enhanced tendon differentiation of the engineered tendon, both in vitro and in vivo. (3) The engineered tendon enhances Achilles tendon regeneration at an early stage. The results thus indicated that a combination of the four elements, transcription factor SCX, hESC-MSCs, silk-collagen scaffold, and dynamic mechanical stimulation, can effectively differentiate hESC-MSCs into the teno-lineage and result in an improved tissue-engineered construction for enhanced tendon healing.
SCX is essential for initiating tendon differentiation
During embryonic development, SCX expression is highly specific, being the earliest known marker of tendon progenitor cells and tenocytes. 24 SCX plays a crucial role in tendon ECM gene expression and tendon-like tissue formation.23,24,35,36 In this study, we used two-stage-engineered tendon construction from hESCs: the initial generation of SCX+ progenitor cells followed by engineered tendon formation. We first overexpressed SCX into hESC-MSCs to generate tendon progenitor cells. SCX expression in turn promoted the synthesis of tendon-specific ECM and enhanced collagen deposition.
SCX is also known to be an upstream regulator of bone- and cartilage-specific genes, such as collagen I, collagen II, aggrecan, and BMP4, and is therefore implicated in bone and cartilage differentiation and development.37–40 Consistent with previous studies, our results showed that SCX was not only involved in tendon differentiation, but also modulated bone and cartilage differentiation. However, the function of SCX varies in different cells and tissues. For example, SCX inhibits chondrogenesis in heart valve and increases collagen I expression in fibroblasts.41,42 Previous study also showed that the bMSC line is negative for SCX, and SCX overexpression inhibits osteogenesis and adipogenesis in bMSCs. 27 Here, we also found that hESC-MSCs did not express SCX, and SCX inhibited adipogenesis as well as downregulating osteocalcin and collagen II. This indicated that SCX may initiate tendon differentiation by modifying the function of other transcription factors. However, unlike the previous study, Tnmd could not be induced in hESC-MSCs.
Moreover, we found that the tendon expression pattern was different in two-dimensional (2D) culture than in 3D-engineered tendon. This is consistent with previously reported data that demonstrate that 3D culture is essential for maintaining the functional phenotype of tenocytes,43,44 may alter the cell physiology, and is essential for maintaining tenocyte gene expression patterns similar to normal tendon.33,43 We found that two ECM genes, decorin and collagen III, which are highly expressed in the repaired tendon and impair collagen maturation, were inhibited by SCX. Moreover, SCX reduced the osteogenesis of hESC-MSCs in the engineered tendon. The underlying mechanisms of the different functions of SCX within 2D and 3D cultures need to be further clarified.
SCX+hESC-MSCs and silk scaffold efficiently improve tendon tissue engineering
Our previous work showed that hESC-MSCs can be used to construct engineered tendon with a knitted silk-collagen sponge composite scaffold and promote tendon repair. The scaffold possesses the optimal microenvironment for cells to attach and grow with excellent mechanical strength and biocompatibility.30,31 However, the repaired tendon is still much inferior to native tendon, and this may be because the seed cells do not exclusively differentiate into the teno-lineage. In this study, we successfully fabricated engineered tendons with SCX+hESC-MSCs and a silk-collagen scaffold. As SCX expression alone is inadequate for inducing the terminal differentiation and maturation of tenocytes, we used mechanical stress as an additional induction signal. Mechanical stress is well-known to be an inducing factor for tendon differentiation.16,45 In the engineered tendon with mechanical stress, collagens Ia1 and collagen Ia2 were upregulated, as well as the tendon-related transcription factor EYA2, which indicated that mechanical stress initiates tendon differentiation. However, the mechanisms of tendon maturation induced by mechanical stress are poorly understood, and alone it is not enough to induce tendon maturation. In this study, we also found that SCX alone was inadequate for inducing terminal tendon differentiation and maturation, such as EYA2 and TNMD gene expression at 4 h. SCX is also known to be an upstream regulator of bone- and cartilage-specific genes, such as collagen I, collagen II, aggrecan, and BMP4, and is therefore implicated in bone and cartilage differentiation and development.37–40,46 However, the function of SCX varies in different cells and tissues. We used mechanical stress as a synergistic tendon-induction signal. Mechanical stimulation led to an increase of collagen XIV and Tnmd expression only in SCX+hESC-MSCs but not in control cells (Fig. 4A). This indicated that SCX with mechanical stress can induce hESC-MSCs to differentiate into tenocytes.
To evaluate whether the engineered tendon can also develop successfully in the in vivo environment, which is unpredictable and complicated, we implanted the construct ectopically in nude mice, as shown in our previous work.20,31 Under natural mechanical stress, cells showed a more physiological tenocyte-like morphology and the collagen fibers were more mature. In this study, we also found that the SCX group had more collagen deposition than the control cells from our previous work. 31 As collagen is the most abundant ECM protein and plays an integral role in maintaining the physiological structure and function of normal tendons, 20 these results indicated that SCX promotes engineered tendon function in the in vivo microenvironment.
SCX+hESC-MSC-engineered tendon with silk scaffold promotes in vivo Achilles tendon regeneration
In the model of Achilles tendon defect, recombinant SCX expression in engineered tendon induced denser connective tissues with more mature collagen fibers at an early stage and more crimped patterns were formed at later stages. More fibroblast-like cells were found in the repaired tendon, which correlates with tendon regeneration. 32 Consistent with the histological data, SCX+hESC-MSCs modestly, but significantly, improved the modulus and stress of the repaired tendons. As shown in our previous study, both collagen sponge 30 and seed cells 20 have the potential to improve collagen maturation within the repaired tendon. The results indicated that seed cells together with ECM have synergistic effects on collagen maturation and restoration of the mechanical properties of the repaired tendon.
As seed cells, SCX+hESC-MSCs may be more appropriate for repairing injured tendon. SCX expression may change the ECM composition and cytokine secretory profile of the cells in the in vivo environment and promote active endogenous tendon regeneration, such as biglycan and collagen XIV expression. However, the key ECM molecules and growth factors secreted by seed cells that promote collagen maturation need to be further identified.
Although SCX+hESC-MSCs with knitted silk-collagen improve the maturation of tendon-like tissues and enhance tendon regeneration in an animal model, there are still many challenges to be overcome before clinical application. As mature tendon formation and full tendon regeneration have not been achieved, further understanding of the mechanisms of tendon differentiation, tendon biology, and regeneration is necessary. Recent studies also showed that BMP2, Smad8, and tendon ECM are important for tendon stem-cell differentiation and lineage commitment.18,47 It will be interesting to determine the optimal inductive conditions for the tenogenic differentiation of hESCs by examining a combination of cytokines, transcription factors, and ECM.
Conclusion
Our results collectively demonstrate that hESCs can aid regeneration of functional tendons through the synergistic effect of recombinant SCX expression and a knitted silk-collagen sponge scaffold both in vitro and in vivo. The efficacy of this engineered tendon in promoting tendon repair was positively demonstrated in a rat model of Achilles tendon injury. As a result, engineered tendons derived from SCX+hESC-MSCs can serve as novel seed cells for the regeneration of injured tendons. These findings may have considerable importance in understanding the roles of SCX expression in tendon differentiation, as well as in developing novel therapeutics for tendon regeneration by using hESCs.
Footnotes
Acknowledgments
This work was supported by NSFC (81330041, 81125014, 31271041, 81201396, J1103603), Fundamental Research Funds for the Central Universities, International Science & Technology Cooperation Program of China (2011DFA32190). The National Key Scientific Program (2012CB966604), 863 Program (No. 2012AA020503). Sponsored by Regenerative Medicine in Innovative Medical Subjects of Zhejiang Province. Medical and health science and technology plan of Department of Health of Zhejiang Province (2013RCA010).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.