
Abstract
Embryonic stem cells (ESCs) emerge as a promising tool for tissue engineering and regenerative medicines due to their extensive self-renewal ability and the capacity to give rise to cells from all three-germ layers. ESCs also secrete a large amount of endogenous extracellular matrices (ECMs), which play an important role in regulating ESC self-renewal, lineage commitment, and tissue morphogenesis. ECMs derived from ESCs have a broader signaling capacity compared to somatic ECMs and are predicted to have a lower risk of tumor formation associated with ESCs. In this study, ECMs from undifferentiated ESC monolayers, undifferentiated aggregates, or differentiated embryoid bodies at different developmental stages and lineage specifications were decellularized and their capacities to direct ESC proliferation and differentiation were characterized. The results demonstrate that the ESC-derived ECMs were able to influence ESC proliferation and differentiation by direct interactions with the cells and by influencing the signaling functions of the regulatory macromolecules such as retinoic acid. Such matrices have the potential to present regulatory signals to direct lineage- and development-specific cellular responses for in vitro applications or cell delivery.
Introduction
P
Stem cell-derived ECMs have been used to support in vitro cell expansion and differentiation as well as in vivo tissue regeneration.12–14 Tissue-specific ECMs derived from mesenchymal stem cells (MSCs) directed MSC-lineage specification and augmented tissue regeneration by extending site-specific MSC retention.13–17 In addition, microcarriers made from MSC-derived ECMs promoted MSC adipogenesis and demonstrated in vivo compatibility, indicating the feasibility of their use in large-scale cell production and cell/matrix delivery.14,15,18,19 The ECMs decellularized from ESC cultures provided a permissive microenvironment for tissue remodeling and fibroblast repopulation.11,12 The decellularized ECMs of ESCs have also been shown to contain ESC-secreted factors such as Lefty, which inhibited the growth and migration of tumor cells.20,21 Compared to the ECMs derived from adult stem cells or somatic tissues, the decellularized matrices from PSCs may have a broader spectrum of signaling capacity owing to their embryonic origin, serving as novel biomaterials for in vivo cell and/or matrix delivery as well as in vitro cell expansion and differentiation.6,10 The PSC-derived ECMs may have the reduced risk of tumor formation compared to the live cells, significantly improving their prospects in clinical applications.
In vitro, PSCs have been grown as undifferentiated monolayers or 3D aggregates for expansion, or as embryoid bodies (EBs) for differentiation into specific lineages.22–25 These organizations have been shown to affect the secretion of ECM proteins and autocrine factors, producing an ECM microenvironment of distinct characteristics.20,26,27 For example, deposition of endogenous transforming growth factor-beta inhibitor, Lefty, into ECMs has been reported for undifferentiated ESCs but not EBs, whereas the EBs displayed a unique ECM-shell structure and different cytokine secretion profiles compared to undifferentiated ESCs.20,28–30 PSCs expanded as aggregates upregulated E-cadherin expression and downregulated Wnt signaling upon differentiation compared to PSCs cultured on 2D substrates, suggesting the impact of intercellular interactions on cell signaling. 26 Parallel to the organizational dependence, the characteristics of PSC-derived ECMs are also influenced by lineage specifications.6,23,29 For example, cerberus, a small antagonist of bone morphogenetic protein (BMP), was detected in the secretome and ECMs of ESCs undergoing cardiac differentiation, but not neural differentiation.6,28 Finally, the ECMs derived at the early or late stage of EBs exhibited different signaling capacities, suggesting the influence of the developmental stage on ECM characteristics.10,11,28
The goal of the present study is to evaluate the influence of ESC organizations (i.e., undifferentiated monolayers, undifferentiated 3D aggregates, and differentiated EBs) and lineage commitment on the characteristics of the decellularized ECMs. The ECMs at the early or late stage of EB differentiation were also derived from days 3 or 10 EBs, respectively.23,28 To create the ECMs with lineage-specific cues, EBs were treated with retinoic acid (RA) to induce germ-layer differentiation or BMP-4 for mesoderm differentiation.31–33 To probe the role of RA signaling in the characteristics of decellularized ECMs, the cellular expression of a RA receptor (RAR) and the effect of a pan-RAR antagonist were investigated. The results suggest that the decellularized matrices derived from ESCs are able to preserve the developmental cues and have a distinct capacity in directing the ESC fate.
Materials and Methods
Undifferentiated ESC culture
Monolayer culture
The murine ES-D3 line (American Type Culture Collection) was maintained on 0.1% gelatin-coated six-well culture plates (Millipore) in a standard 5% CO2 incubator. The growth medium consists of the Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% ESC-screened fetal bovine serum (FBS; Hyclone), 1 mM sodium pyruvate, 0.1 mM β-mercaptoethanol, penicillin (100 U/mL), streptomycin (100 μg/mL) (all from Invitrogen), and 1000 U/mL LIF (Millipore). The cells were seeded at 2−4×104 cells/cm2 and subcultured every 2–3 days.
Aggregate culture
The aggregates of ES-D3 cells were obtained by seeding the cells from ESC monolayer into Ultra-Low Attachment (ULA) six-well plates (Corning, Inc.) at a seeding density of 2−4×104 cells/cm2 in 3 mL of growth media. 34 The aggregates were subcultured every 2–4 days and serially passaged for 5–15 passages. For passaging, the aggregates were trypsinized with 0.05% trypsin-EDTA for 5 min at 37°C, neutralized with the media containing FBS, and resuspended in the fresh media.
Differentiated EB culture
For EB formation, 1×106 cells were seeded in the ULA six-well plates. Three types of EBs were cultivated in a differentiation medium for up to 10 days: (1) spontaneously differentiated EBs; (2) EBs treated with 1 μM all-trans RA (Sigma-Aldrich); (3) EBs treated with 5 ng/mL BMP-4 (R&D Systems). The differentiation medium consists of the DMEM supplemented with 10% FBS, 0.1 mM β-mercaptoethanol, penicillin (100 U/mL), and streptomycin (100 μg/mL).
Derivation of decellularized ECMs from monolayers, aggregates, and EBs
Four decellularization reagents, including (1) 1% Triton X-100, (2) 20 mM NH4OH, (3) 1% SDS, and (4) 1% Triton X-100+20 mM NH4OH (all from Sigma), were tested and compared (Table 1).11,17,35 All the decellularization reagents were sterilized by membrane filtration before the treatment. Briefly, about 600–1000 EBs (150–300 μm in diameter) were distributed into each of the 1.5-mL microcentrifuge tubes and treated with different decellularization solutions for 30 min. After the treatment, the samples were spun down at 18,000 g for 2 min, rinsed twice with phosphate-buffered saline (PBS), and incubated with 2000 unit/mL DNAse I for 15–30 min. The samples were centrifuged at 18,000 g for 2 min and rinsed twice with PBS before characterization or cell reseeding.
For DNA removal, the scale +++ indicates that 85–100% of DNA is removed (the remaining DNA is 0–15%); ++ indicates 70–85% removal (the remaining DNA is 15–30%);+indicates 30–70% removal (the remaining DNA is 30–70%); − indicates less than 30% removal (the remaining DNA is more than 70%). Please refer to Figure 2 for the detailed DNA expression for each condition.
ECM, extracellular matrices; ND, not determined.
Based on the residual DNA and preservation of the ECM structure, 1% Triton X-100 and 30 min DNAse I treatment were selected and used to obtain different decellularized ECMs (DE) from monolayers, aggregates, and EBs (Table 2). The monolayers were decellularized similarly to aggregates, but centrifugation steps were omitted. For undifferentiated ESCs, DEs were obtained from day 3 cultures only because the prolonged culture induces differentiation. For EBs, DEs were obtained after culturing for 3 or 10 days.
Reseeding was not performed because the small difference in ECM expression compared to DE-SE. Instead, day 10 samples were reseeded for the comparison with DE-SE scaffolds.
ESC, embryonic stem cell; RA, retinoic acid; LIF, leukemia inhibitory factor; BMP-4, bone morphogenetic protein; DE, decellularized ECMs.
DNA assay
The residual DNA after decellularization was measured using a DNA assay. 36 A DNA standard was prepared by dissolving salmon testes DNA in TEX (10 mM Tris, 1 mM EDTA, 0.1% Triton X-100 at pH 8) and a standard curve was constructed for each assay. The decellularized samples were lysed with 0.1 mg/mL proteinase K (Fisher Scientific) at 50°C overnight. The lysate (100 μL) were placed in triplicate into a 96-well plate and 100 μL of Picogreen (Molecular Probes) were added to each well. The plate was incubated for 5 min in the dark, and then read on a fluorescent plate reader (FLX800; Bioinstrument, Inc.).
Scanning electron microscopy and atomic force microscopy
For scanning electron microscopy (SEM), the samples were washed with PBS, fixed in 2.5% glutaraldehyde for 30–60 min, and dehydrated in graded ethanol solutions. The samples were dried by hexamethyldisilazane evaporation, mounted, and sputter-coated with iridium. Observations were made using a Nova 400 Nano SEM (FEI) under low-vacuum conditions. For atomic force microscopy (AFM), the samples were fixed with 4% paraformaldehyde (PFA), dehydrated at 60°C, and then analyzed in contact mode using a Dimension 3000 scanning probe microscope (Digital Instruments). The roughness and topography were assessed from the acquired information.
Immunocytochemistry
For ECM expression, the monolayers, aggregates, or EBs before and after decellularization were fixed with 4% PFA. 34 The samples were permeabilized with 0.2–0.5% Triton X-100, blocked, and incubated with primary ECM antibodies, including rabbit polyclonal FN, laminin (LN), collagen IV (Col IV), and vitronectin (VN) (Abcam). For fluorescence staining, the samples were incubated with Alexa Fluor® 488 goat anti-Rabbit IgG (Molecular Probes), counterstained with 4′,6-diamidino-2-phenylindole (DAPI), and visualized using a fluorescence microscope (Olympus IX70) or a Zeiss 510 confocal microscope (Leica TCS SP2 AOBS). The negative control with a secondary antibody only revealed no signal. For the detection of glycosaminoglycans (GAGs), the samples were incubated in 1% Alcian blue 8GX (Sigma) that was diluted in 3% acetic acid (pH 2.5) for 30 min, washed with water, and analyzed under a light microscope.
To quantify the ECM contents, the samples were incubated with donkey anti-Rabbit IgG conjugated with horseradish peroxidase (HRP; Rockland Immunochemicals, Inc.) after the incubation with primary antibodies. After washing, 1 mL of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (Thermo Scientific) was added to the samples, incubated for 5–25 min, and stopped by a solution of 0.16 M sulfuric acid. The absorbance units (AU) were measured using a microplate reader (Biorad) at a wavelength of 405 nm with background subtraction at 655 nm. The AU values were also corrected by subtracting the absorbance of negative control stained with HRP-IgG only. The measurements were normalized to the cell number for comparison.
To detect differentiation markers, the cells were incubated with the mouse or rabbit primary antibody against Nestin (Millipore), β-Tubulin III (Sigma), α-Actinin (sarcomeric; Sigma), or FOXA2 (Millipore). After washing, the cells were incubated with the corresponding secondary antibody Alexa Fluor 488 goat anti-Mouse IgG1 (for Nestin and α-Actinin) or IgG2b (for β-Tubulin III), or Alexa Fluor 488 goat anti-Rabbit IgG (for FOXA2). The localization of RAR-α was also assessed using the goat anti-Rabbit RAR-α antibody (Abcam) detected by Alexa Fluor 594® donkey anti-Goat IgG.
Cell reseeding and cultivation on the decellularized ECMs
To assess cell proliferation, ESCs of 0.5×105 per mL from the stock culture were seeded on the decelluarized ECMs and cultivated for 4 days in expansion media without LIF (Fig. 1). About 30–50% of seeding efficiency was observed for 3D ECMs and 50% for 2D ECMs. During the culture period, the unattached cells died out or self-assembled into aggregates, which may become associated with ECM scaffolds during culture with an estimated additional 20–30% increase in seeding efficiency. Cell numbers were determined at days 0, 2, and 4 using a hemocytometer after trypsin/EDTA dissociation. The specific growth rates were calculated as the slope of the proliferation curve. Cell viability was determined by the incubation with 5 mg/mL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma) solution. Afterward, the formazan crystals were hydrolyzed with dimethyl sulfoxide (Sigma) and measured at 500 nm using a microplate reader. For the test of RA signaling, soluble RA (1 μM) or BMS 493 (1 μM; Santa Cruz), a pan-RAR antagonist, was supplemented in the media. To assess the differentiation ability after culturing on the decellularized ECMs, ESCs were replated on 0.1% gelatin-coated 12-well plates and cultured for 7 days in the differentiation medium without any inducing growth factors. The cells were evaluated for the expression of Nestin (ectoderm), β-Tubulin III (ectoderm), α-Actinin (mesoderm), and FOXA2 (endoderm).

Schematic diagram of the experimental procedure. ESCs were seeded as monolayer, aggregates, or EBs for 3 to 10 days. The monolayers, aggregates, or EBs were decellularized, and ESCs were reseeded on the ECM scaffolds for 4 days without LIF as short-term expansion. The cells were replated and grown for 7 days in the absence of inducing growth factors for differentiation. Decellularization was also performed for EBs treated with BMP-4 or RA. ESCs, embryonic stem cells; ECM, extracellular matrices; ESCs, embryonic stem cells; EBs, embryoid bodies; RA, retinoic acid. BMP-4, bone morphogenetic protein. Color images available online at www.liebertpub.com/tea
Bromodeoxyuridine proliferation assay
The cells were incubated in the media containing 10 μM bromodeoxyuridine (BrdU; Sigma) for 30 min to allow BrdU, a synthetic thymidine analog, incorporating into the DNA during the S-phase of cell cycle. The cells were then fixed with 70% cold ethanol, followed by a denaturation step using 2 N HCl/0.5% Triton X-100 for 30 min in the dark. The samples were reduced with 1 mg/mL sodium borohydride for 5 min and incubated with mouse anti-BrdU (Invitrogen) in the blocking buffer (0.5% Tween 20/1% bovine serum albumin in PBS), followed by the incubation with Alexa Fluor 488 goat anti-Mouse IgG1. The cells were mounted with DAPI and visualized using a Zeiss 510 confocal microscope.
Flow cytometry
For flow cytometry, 1×106 cells per sample were fixed with 4% PFA and washed with a staining buffer (2% FBS in PBS). The cells were stained with DNA-intercalating dye ethidium monoazide bromide (Sigma) and permeabilized with 100% cold methanol. The samples were incubated with primary antibodies against Oct-4 (Millipore), Nestin, β-Tubulin III (Millipore), α-Actinin, or FOXA2 followed by the corresponding secondary antibody Alexa Fluor 488 goat anti-Mouse IgG1 (for Oct-4, Nestin, α-Actinin, and β-Tubulin) or Alexa Fluor 488 goat anti-Rabbit IgG (for FOXA2). The cells were acquired with the BD FACSCanto™ II flow cytometer (Becton Dickinson) and analyzed against isotype controls using FlowJo software.
Statistical analysis
Each experiment was carried out at least twice. The average values of two or three independent experiments were presented and the results are expressed as mean±mean absolute deviation. In each experiment, triplicate samples were used. To assess the statistical significance, ANOVA followed by Fisher's LSD post hoc tests or t-tests were performed. A p-value<0.05 was considered statistically significant.
Results
Derivation and characterization of decellularized ECM from ESCs
Among the various decellularization methods tested, SDS condition displayed the highest efficiency of DNA removal with less than 5% DNA remaining in the acellular structure (Fig. 2A). Treatment with Triton X-100 was also effective when DNAse incubation increased from 15 to 30 min (55% vs. 21%). NH4OH treatment alone did not reduce the DNA content and its combination with Triton X-100 did not add beneficial effects. For ECM proteins, SDS treatment maintained the expression of FN, LN, and Col IV, but reduced VN slightly (Fig. 2B). Treatment with Triton X-100 plus 30 min DNAse was comparable to SDS condition, whereas cotreatment with NH4OH reduced LN and Col IV. Because the SDS-treated ECMs displayed a viscous, semifluidic appearance that was difficult to handle, Triton X-100 plus 30 min DNAse treatment was chosen in the following studies (Table 1).
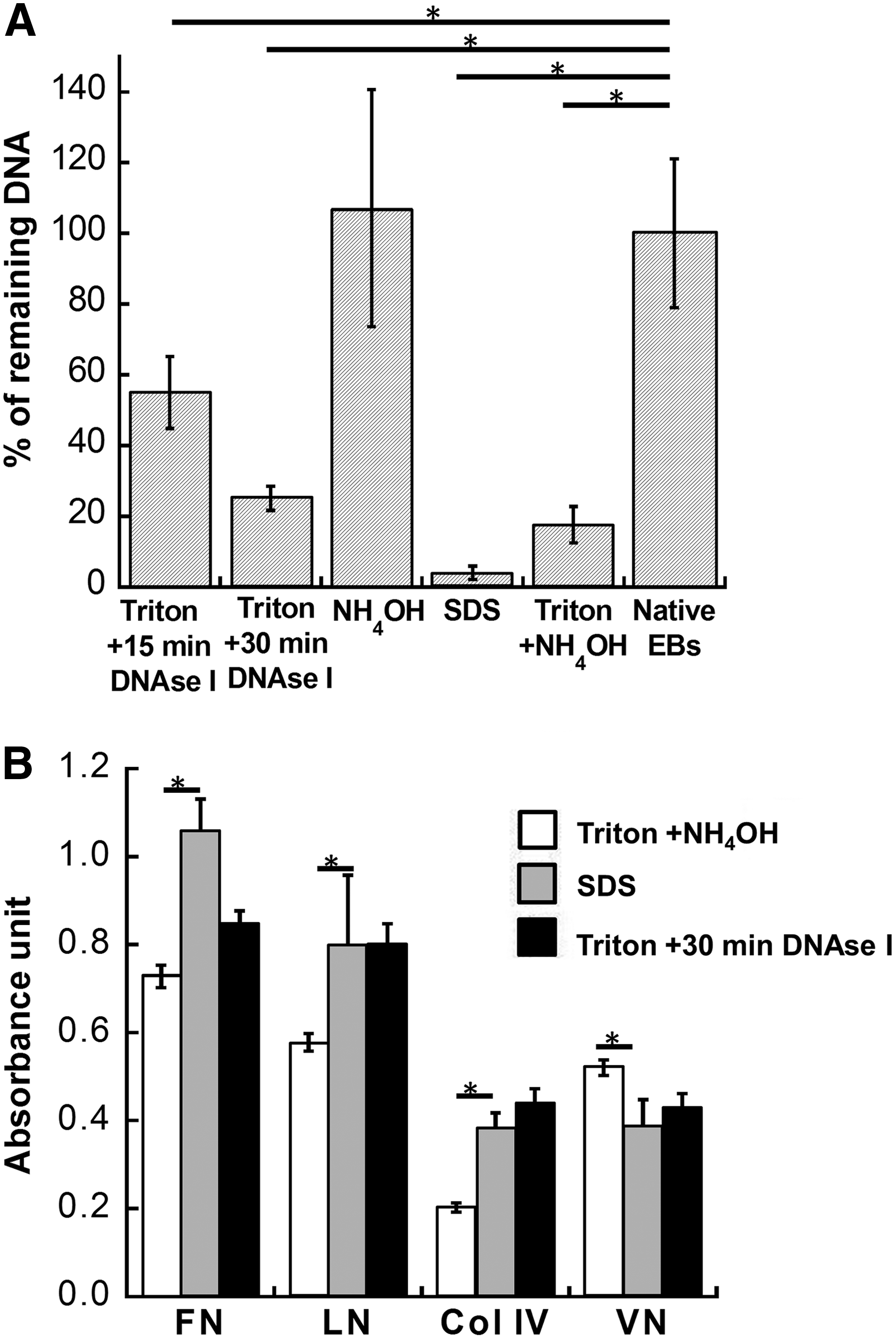
Comparison of decellularization methods for DNA removal and ECM preservation. EBs after 3-day culture were decellularized with different methods.
The ultrastructure analysis of the decellularized ECMs revealed contrasting morphologies before and after decellularization, with tightly packed cells becoming indistinguishable after the treatment (Fig. 3A). Compared to the gelatin-coated surface, the roughness (Ra) increased from 3 to 48 nm for the decellularized monolayers. The surface roughness was comparable for the decellularized aggregates and EBs (246 vs. 285 nm) and both were much higher compared with the decellularized monolayers (Fig. 3B), suggesting distinct ultrastructure characteristics of the decellularized ECMs from different ESC organizations.

Characterization of surface topography of decellularized ECMs. Monolayers, aggregates, and EBs were decellularized after 3 days of culture.
ECM protein expression before and after decellularization
ECM expression, including FN, LN, Col IV, VN, and GAGs, was preserved after decellularization. The ECMs from aggregates and EBs exhibited denser structures due to the compaction by centrifugation (Fig. 3C and Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
The specific ECM expression decreased by three- to fivefolds over 3 days of monolayer culture (Fig. 4A and Supplementary Fig. S2). Similarly, for the ESC aggregates, specific ECM expression also significantly decreased (two- to fourfolds) with culture time (Fig. 4B and Supplementary Fig. S2). After decellularization, the expression of ECM proteins in the monolayers (DE-M) was unchanged (Fig. 4C), indicating that decellularization did not modify the ECM composition. For the decellularized aggregates (DE-A), the expressions of FN, LN, and VN were maintained with a slight decrease in Col IV expression (Fig. 4D). Despite the higher specific ECM expression at day 1 compared to day 3, day 1 ECMs of monolayers contained lots of empty surface; thus, the restriction of the reseeded cells to the decellularized ECMs would be more variable compared to day 3 ECMs. So the decellularized ECMs from day 3 monolayers or aggregates were used in the subsequent studies.

Quantitative analysis of ECM composition pre- and post-decellularization of undifferentiated ESCs.
For the spontaneously differentiated EBs, extended culture from 3 to 10 days increased the specific ECM expression by two- to fourfolds (Fig. 5A). Following decellularization, the expressions of FN and LN were maintained, whereas the Col IV expression was reduced (Fig. 5B). The EBs were also treated with RA or BMP-4 to favor neural or mesodermal cells (Supplementary Fig. S3). For the growth factor-treated EBs, the differences in ECM expressions among the DE-SE, DE-RA, and DE-BMP were more pronounced for day 10 samples compared to day 3 samples, with the DE-RA10 and the DE-BMP-10 having a higher specific VN and Col IV expression compared to the DE-SE10 (0.8–1.1 vs. 0.2–0.3) (Fig. 5C and Supplementary Fig. S2). For FN and LN, the expression was differentially observed for the DE-SE10, DE-RA10, and DE-BMP-10 (0.4–0.6, 0.7–1.0, and 1.7–1.8, respectively). To ensure sufficient signals, day 10 samples of the DE-BMP and the DE-RA were used to further evaluate cellular responses. Taken together, these results indicated that the decellularized ECMs were dynamically remodeled during ESC expansion and differentiation.
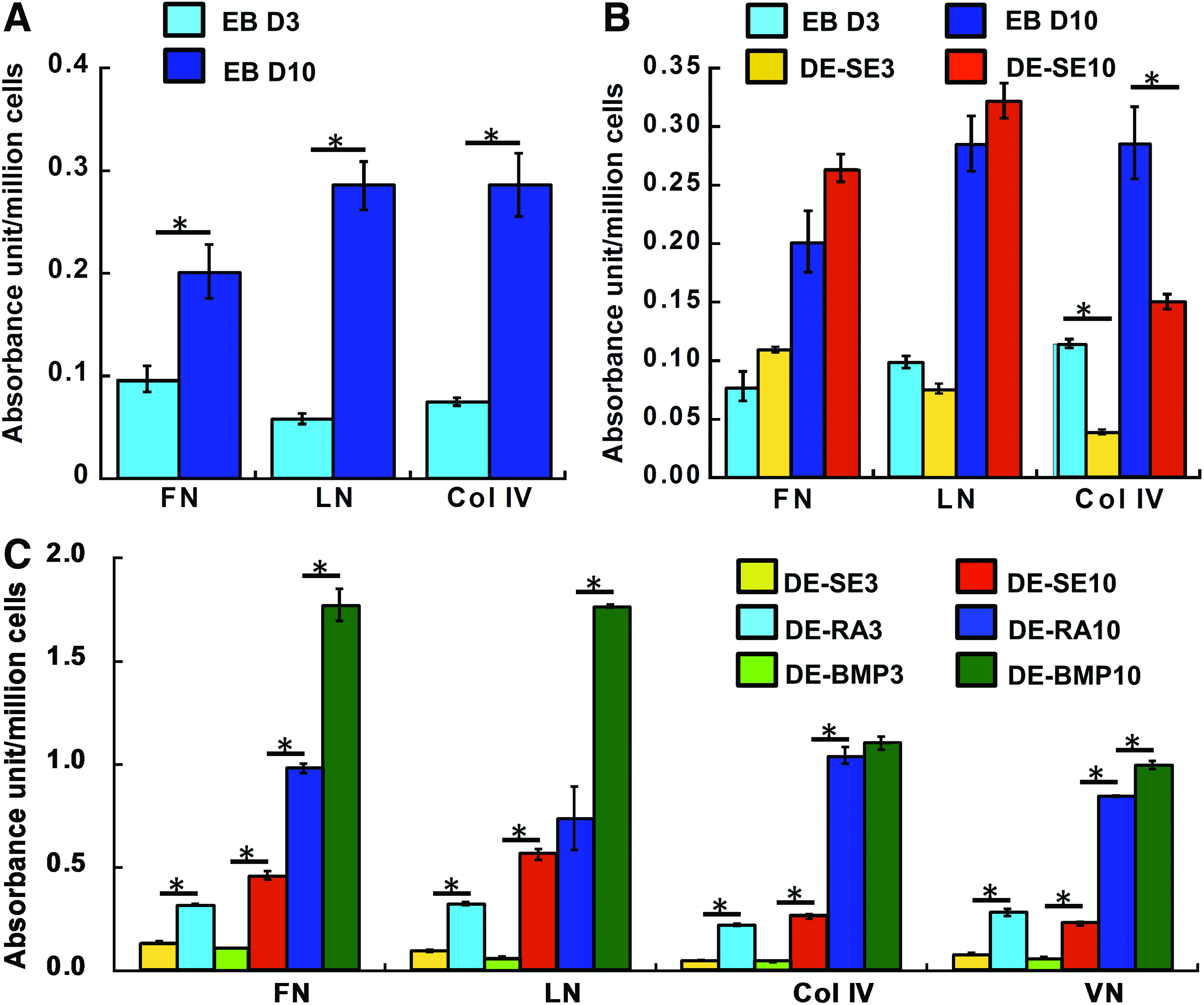
Quantitative analysis of ECM compositions pre- and post-decellularization of EBs.
Effects of the decellularized ECMs on ESC proliferation
ESC proliferation on the decellularized matrices was assessed. The specific growth rate of ESCs on the DE-M3 or the DE-A3 was similar to gelatin control (4×10−2 h−1) (Fig. 6A). Cells on the DE-SE3 had a slightly lower growth rate without statistical significance (3.1×10−2 h−1). The ESCs seeded on the DE-RA10 had a significantly lower specific growth rate compared to the DE-BMP-10 and the DE-SE10 scaffolds (1.6 vs. 3.5−4×10−2 h−1). The cell viability determined by MTT assay was consistent with the specific growth rates (Fig. 6B). BrdU staining revealed the spatial distribution of the proliferating ESCs after reseeding. Whereas the BrdU+ cells evenly distributed in the DE-SE10 and the DE-BMP-10, less BrdU+ cells were observed in the central region of the DE-RA10 (Fig. 6C).
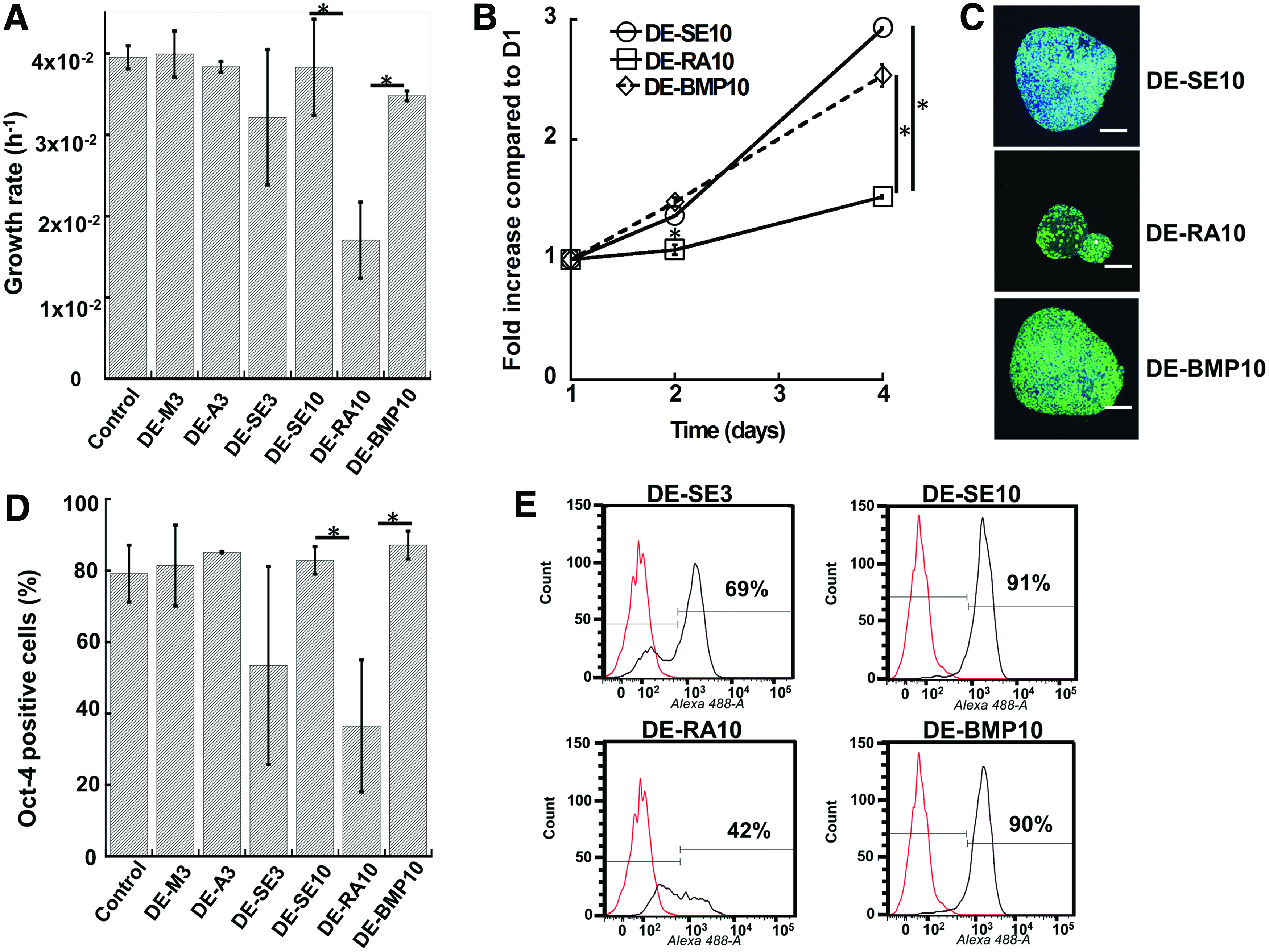
Proliferation and Oct-4 expression of the reseeded ESCs on the decellularized ECM scaffolds.
The Oct-4 expression of ESCs followed similar trends as observed in the specific growth rates. About 81–86% of the cells seeded on the DE-M3 and the DE-A3 were Oct-4+, similar to the gelatin-coated surface (Fig. 6D). The DE-SE3 supported a lower percentage of Oct-4+ cells, but the difference was not statistically significant compared to gelatin control. Whereas the DE-SE10 and the DE-BMP-10 maintained 84–91% Oct-4+ cells, Oct-4 expression of cells on the DE-RA10 was only 38% (Fig. 6D, E). A similar trend was observed for SSEA-1 (data not shown).
Effects of the decellularized ECMs on ESC differentiation
Cells seeded on various decellularized ECM scaffolds were assessed for the expression of differentiated markers, including Nestin, α-Actinin, β-Tubulin III, and FOXA2 (Fig. 7 and Supplementary Fig. S4). Due to the absence of inducing growth factors, the lineage-specific differentiation for cells on various ECM scaffolds was mainly attributed to the signals from the ECMs. The expression of α-Actinin in this study represented mainly skeletal muscle cells rather than cardiac muscle cells due to the low frequency of beating cells. For the cells grown on the day 10 DEs (DE-SE10, DE-RA10, and DE-BMP-10), higher expressions of FOXA2 (28–52% vs. 14–16%) and α-Actinin (15–32% vs. 4–5%) were observed compared to the cells grown on the day 3 DEs (DE-A3 and DE-SE3). Specifically, after differentiation, the ESCs seeded on the DE-RA10 had the highest expression of Nestin (46%), α-Actinin (32%), β-Tubulin III (36%), and FOXA2 (52%) compared to the cells seeded on the other ECM scaffolds (Fig. 7C, F, I, L). For the DE-A3 and the DE-SE3, which had low expression of the assessed markers, only about 5–10% of cells expressed Oct-4 (data not shown) indicating the loss of pluripotency. A more detailed analysis with markers to cover the development of lineage specification is warranted in future studies.
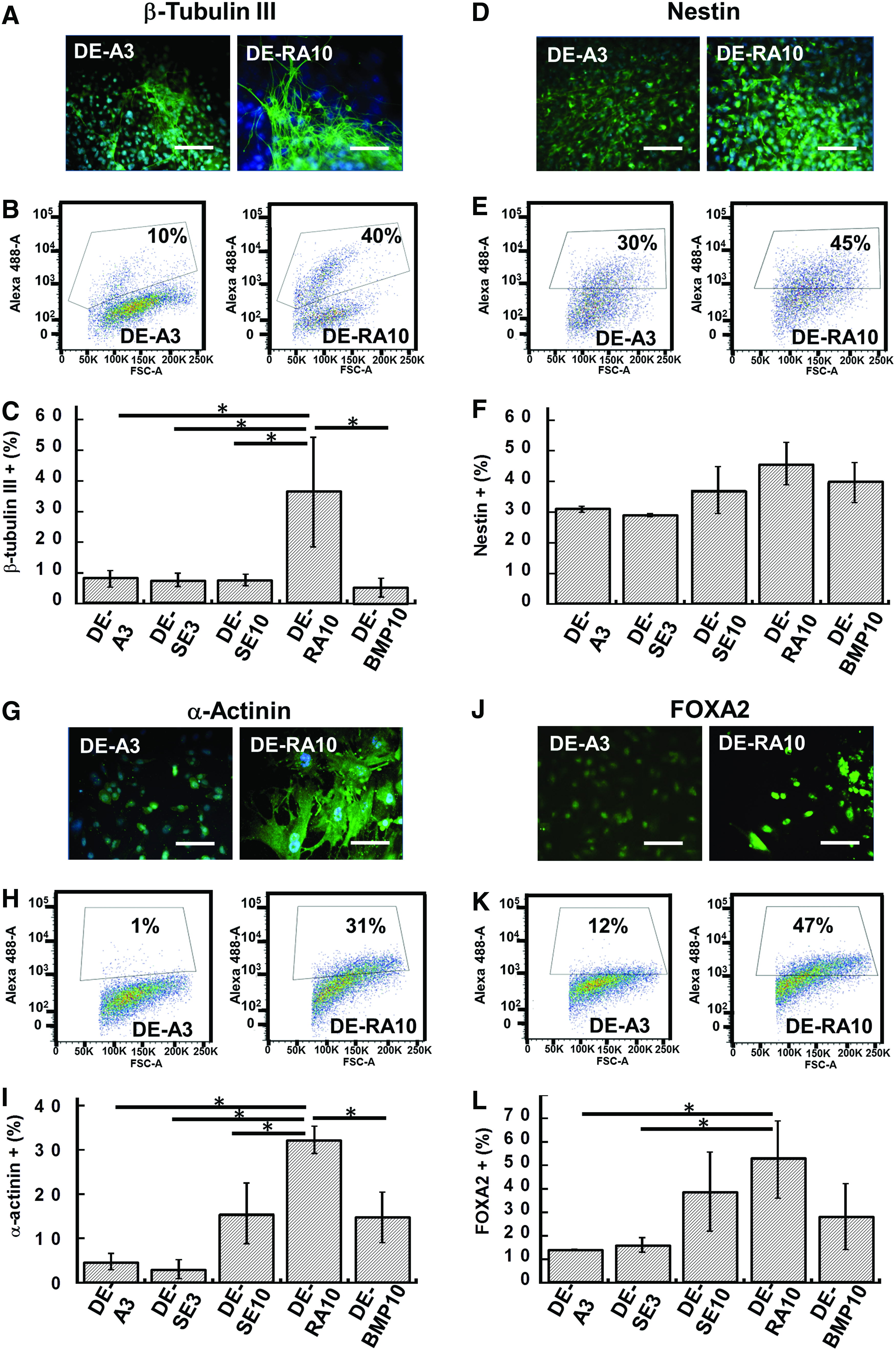
Differentiated marker expression of ESCs grown on decellularized ECM scaffolds. Representative fluorescence images and flow cytometry histograms of ectoderm markers β-Tubulin III
Role of RA–RAR interactions
To investigate the role of RA signaling in the ECM scaffolds, RAR-α expression by ESCs cultured on the DE-RA10 was evaluated. First, the translocation of RAR-α from the cytoplasm to the cell nucleus was confirmed for ESCs exposed to exogenous RA (Fig. 8A). Then, ESCs grown in DE-RA10 were assessed and the localizations of RAR-α both in the cytoplasm and cell nucleus were observed (Fig. 8B). Because no soluble RA was added in the DE-RA10 culture, the mixed pattern of RAR-α expression suggested active RA signaling presumably due to the presence of the decellularized ECMs.
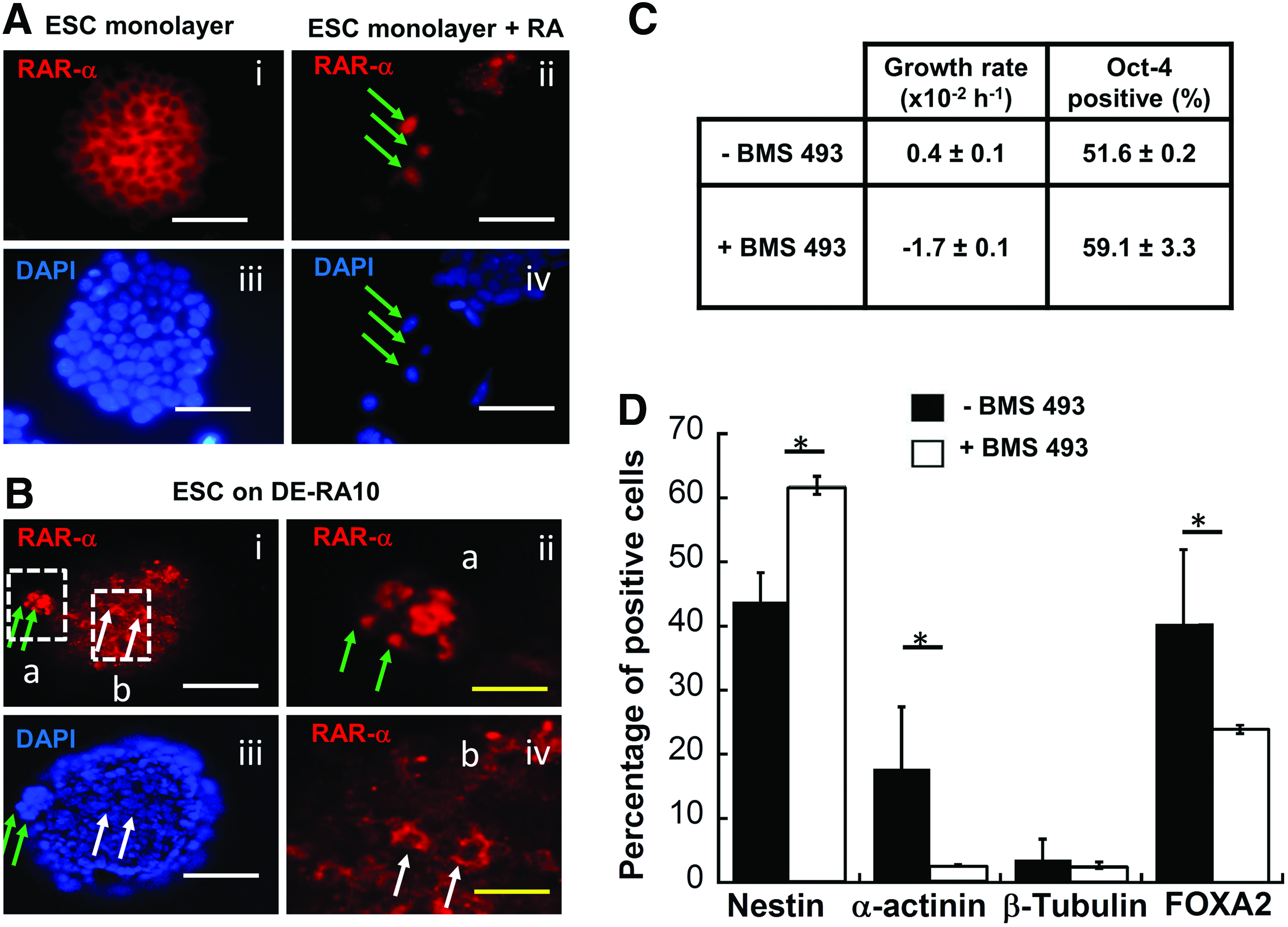
Role of RA–RAR interactions in ESCs cultured on the decellularized ECM scaffolds.
Addition of a pan-RAR antagonist, BMS 493, in the DE-RA10 culture reduced ESC proliferation but not Oct-4 expression (Fig. 8C). Upon differentiation, blocking RA–RAR signaling significantly reduced the expression of α-Actinin (3% vs. 18%) and FOXA2 (24% vs. 40%) compared to the untreated group (Fig. 8D). Conversely, Nestin expression was increased in the treated group with similar β-tubulin III expression. These results indicated that the effect of the DE-RA10 on the endodermal and mesodermal differentiation of the reseeded ESCs may be a consequence of the active RA signaling.
Discussion
Different from ECMs derived from somatic cells, the decellularized ECMs from PSCs recapitulate the developmentally relevant extracellular microenvironments and have a unique signaling capacity for directing cell fate.6,10 The results of the present study demonstrated the organization- and lineage-specific characteristics of ESC-derived ECMs and their differential signaling capacity in regulating ESC fate.
Characterization of ESC-derived ECM scaffolds
Formation of EBs is an important step in ESC differentiation in vitro and the derivation of decellularized matrices from spontaneous EBs has been demonstrated in several studies.11,12 However, ESCs are also able to form 3D aggregates while maintaining the undifferentiated phenotype, which facilitates their expansion in suspension. 25 The differences of ESC fate in the EB versus the 3D aggregate may be reflected in their ECM microenvironment, but the derivation and properties of the ECMs from the 3D aggregates have not been reported. In the present study, Triton X-100 followed by DNAse was effective in maintaining the 3D structures of ESC aggregates similar to those derived from EBs. Despite a slight decrease in Col IV, the main components of ECM proteins in the 3D ESC aggregates were preserved.
Whereas the decellularized matrices derived from EBs and ESC aggregates have similar macroscopic structures, they represent divergent developmental stages and thus may have different ECM compositions and signaling capacities. Indeed, quantitative assessment of the decellularized matrices revealed time- and organization-dependent ECM compositions. The specific expression of ECM proteins increased with time during EB development, in contrast to the decreasing trend for the undifferentiated ESC aggregates and the monolayer cultures. The reduced specific ECM expression in the monolayers and undifferentiated aggregates is thought to be a combined result of accelerated cell proliferation accompanied by limited endogenous ECM secretion by undifferentiated ESCs. Although the endogenous ECM proteins such as FN and LN are required for ESC self-renewal, such ECM proteins are limited in quantity in the undifferentiated ESCs to prevent the loss of self-renewal ability. 7 In contrast, it is well established that EB formation is closely coupled with ECM secretion, notably the formation of a basement membrane rich in LN and Col IV.10,23,37,38 The ECMs in the EBs not only play important roles in directing ESC differentiation, but also form supporting structures that sustain EB development. 23 To further probe whether the decelluarized ECMs maintain their lineage-specific properties, the EBs were treated with BMP-4 or RA to induce mesodermal or ectodermal differentiation.31,32 The significantly increased ECM expression and the differential expression patterns for both groups suggest the close association of ECM properties with lineage specification of ESCs. For example, RA-induced differentiation of pluripotent cells has been shown to alter the cellular proteoglycan composition. 39 Together, these results suggest that the decellularized ECMs recapitulate the cellular microenvironments associated with ESC development in vitro and that the characteristics of ECMs derived from ESC cultures depend on both spatial organization and lineage specification.
ESC proliferation on the decellularized ECM scaffolds
The decellularized ECMs, except the DE-RA10, supported short-term ESC proliferation and Oct-4 expression at a level comparable to that of the gelatin-coated surface. It is interesting to note that no significant difference in cell proliferation was observed between the DE-A and the DE-SE groups, two types of matrices with divergent ECM compositions. This may be explained by the fact that multiple components in the decellularized ECMs influence ESC proliferation. Compared to the purified ECM proteins, the decellularized ECMs contain a mixture of ECM proteins and provide a multitude of binding sites for both cell adhesions and growth factor sequestrations.5,6,40 Studies have shown that the combinatorial ECMs supported human ESC expansion more robustly compared to individual ECM proteins, probably due to the increased cell binding and integrin signaling.41,42 A limitation of the present study, however, is the short culture duration after reseeding, which may not be indicative of the long-term effects of cellular microenvironments on ESC fate.
The significant reduction in ESC proliferation and Oct-4 expression in the DE-RA10 scaffold indicates its unique ECM microenvironment. Obtained over the same culture period, the DE-SE10, the DE-BMP-10, and the DE-RA10 groups represent the microenvironments of three distinct developmental paths and have different patterns of ECM expressions. However, the differences in ESC proliferation and Oct-4 expression in the DE-RA10 cannot be explained by the differences in ECM composition alone because cell proliferation was not correlated with ECM expressions among these three matrices. Instead, regulatory molecules such as RA retained in the DE-RA10 are thought to play a role. Although unstable and susceptible to oxidative damage in aqueous solutions, RA is intrinsically hydrophobic and its binding to proteins such as serum albumin could significantly prolong its half-life and biological function.43,44 Although no soluble RA existed in the DE-RA10 culture, RA may form RA-albumin complexes and are subsequently retained in the ECMs, altering the endogenous retinoid signaling in ESCs grown in DE-RA10.44–46 Retinoid signaling is known to reduce ESC growth by increasing p27 accumulation and lowering the Oct-4 level by augmenting the Oct-4 chromatin/transcription factor complex. 47 Whereas further experiments are required to elucidate RA's bioactivity in the ECMs, the results support the notion that ECMs have the capacity to regulate stem cell fate through their interactions with the regulatory macromolecules.4,5
ESC differentiation on the decellularized ECM scaffolds
Corresponding to the reduction in cell proliferation, the DE-RA10 exhibited the most dramatic effects on ESC differentiation among all scaffolds with increased expression of four differentiated markers. RA is an important signaling molecule in embryonic development, and has been commonly used as an inducer to initiate ESC differentiation into ectoderm, mesoderm, or endoderm through the temporal- and concentration-dependent retinoid signaling.32,45,48–50 Besides neural lineage, the increased expression of α-Actinin and FOXA2 was also observed, which was consistent with the effect of RA on the upregulation of markers in three-germ layers. 45 Although the exact mechanism of RA signaling in ESCs grown in DE-RA10 requires further study, the coexpression of RAR-α in the nucleus and cytoplasm of cells grown in the DE-RA10 suggested the presence of active RA signaling in the absence of soluble RA. 51 The reduced α-Actinin and FOXA2 expression by blocking RA–RAR interactions using BMS 493 in the DE-RA10 provided further evidence that retinoid signaling played an active role in directing ESC differentiation. 48 The increased Nestin expression may be due to the RAR independent differentiation or the expression from nonectodermal cells. 52 Together, these results reveal the reciprocal interactions between the ECMs and the stem cells, and the capacity of such a microenvironment to regulate cell fate in the absence of the nonphysiological level of soluble RA. 50
The effects of decellularized ECMs derived at different developmental stages were also observed. Compared to the day 3 DEs, including both DE-A3 and DE-SE3, cells grown in the day 10 DEs expressed a higher level of α-Actinin and FOXA2. Due to the progression in differentiation, the day 10 DEs may contain cell-secreted signals (e.g., Cerberus) that are more inductive for differentiation compared to the day 3 DEs.10,28 In addition, the higher content of ECM proteins in day 10 DEs might also contribute to the increased α-Actinin and FOXA2 expression, because the incorporation of biomaterials in ESC aggregates has been shown to promote mesodermal and endodermal differentiation by affecting cell–cell interactions. 53
Conclusion
The ECMs derived from ESC cultures maintained their signaling capacities that are specific to cellular organizations, lineage specifications, and developmental stages. Such ECMs regulate ESC fate by providing cell adhesion sites as well as by the interactions with regulatory macromolecules. The ESC-derived ECMs have the potential to direct lineage-specific ESC responses and influence germ-layer differentiation for in vivo cell and/or matrix delivery as well as in vitro cell expansion and differentiation.
Footnotes
Acknowledgments
The authors would like to thank Dr. Eric Lochner of FSU Condensed Matter and Materials Physics (CMMP) facilities and Dr. Brett Kirkland from the FSU Chemical and Biomedical Engineering department for their help in AFM analysis. The authors also thank Ms. Ruth Didier of the FSU Department of Biomedical Sciences for her help in the confocal microscopy and flow cytometry analysis. This work is supported by the FSU start up fund. Funding support, in part, from the American Heart Association Grant-in-Aid (10GRNT3860040) to T.M. is also acknowledged.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.