
Abstract
Substrate topography influences cell adhesion, proliferation, and differentiation. In this study, poly (ɛ-caprolactone) (PCL) films with a well-defined honeycomb structure of porosity 3–4, 5–6, 10–11, or 15–16 μm were contrasted with flat surfaces for their ability to support primary rat osteoblast adhesion and mineralized extracellular matrix deposition in vitro. Immunofluorescent visualization of vinculin and rhodamine phalloidin binding of actin were used to investigate cell adhesion and morphology. Localization of the alkaline phosphatase activity and Alizarin Red staining were performed to assess the osteoblast activity and deposition of a mineralized matrix. Scanning electron microscopy together with energy-dispersive X-ray spectroscopy was used to provide morphological analysis of cell–film interactions, the deposited matrix, and elemental analysis of the mineralized structures. After 24 h of culture, there were no differences in cell numbers on porous or flat PCL surfaces, but there were changes in cell morphology. Osteoblasts on honeycomb films had a smaller surface area and were less circular than cells on flat PCL. Analysis of cells cultured for 35 days under osteogenic conditions revealed that osteoblasts on all substrates acquired alkaline phosphatase activity, but levels of mineralized matrix were increased on films with 3–4-μm pore sizes. The bone-like matrix with a Ca:P ratio of 1.69±0.08 could be identified in larger areas often aligning with substrate topography. In addition, smaller spherical deposits (0.5–1 μm in diameter) with a Ca:P ratio of 1.3±0.08 were observed at the surface and particularly within the pores of the PCL film. Localization of vinculin showed significant decreases in the number of focal adhesion structures per unit cell area on 5–6, 10–11, and 15–16-μm surfaces compared to flat PCL, while focal complexes with a smaller area (0–2 μm2) were more abundant on 3–4 and 5–6-μm surfaces. Observation of cell interaction with these surfaces identified cytoplasmic protrusions that extended into and sealed the pores of these PCL films creating an extracellular space in which, the conditions could influence the deposition and formation of the mineralized matrix.
Introduction
Biomaterial scientists have long understood that the combination of surface chemistry and structure have the potential to be tailored to ensure that an appropriate tissue response is elicited by an implant. This is particularly evident in orthopedics where surface structuring has been used to improve osseointegration, the process of bone growth directly onto the biomaterial, ensuring long-term fixation and functional effectiveness.14–17 Deposition of bone is achieved by osteoblasts 18 and these cells deposit a proteinaceous matrix that is predominantly type I collagen, and then mediate mineralization by the release of matrix vesicles 19 that drive the formation of hydroxyapatite crystal formation within the protein fibrils.
Previous studies have demonstrated that osteoblast adhesion, proliferation, and differentiation are enhanced by textured surfaces.20,21 However, increased surface roughness may also negatively influence cell adhesion and bone formation.22,23 The gene expression profile, particularly those involved in mesenchymal cell differentiation, has been shown in bone marrow stromal cells to be regulated by surface topography. 24 In addition, microtopography also modulates how osteoblast cells respond to osteotropic regulatory factors such as 17β-estradiol, 25 1, 25(OH)2D3, 8 and BMP-2. 26 It is therefore clear that biomaterial surfaces have the potential to influence cell adhesion, profile of gene expression, and response to growth factors and hormones.
What remains unclear are the exact architectural parameters that influence bone cell function and the mechanisms that control the cell phenotype. Detailed studies on the influence of regular porous films will help clarify the mechanisms underlying the bone cell activity on biomaterials. We have previously found that regular structures are formed during the casting of biodegradable polymer solutions on solid surfaces; for example, self-organized honeycomb-patterned (honeycomb) films27–33 with highly regular porous structures can be prepared under humid casting conditions. In this study, planar substrates with highly controlled honeycomb surface features were created in poly (ɛ-caprolactone) (PCL) and assessed for their ability to support osteoblast adhesion, spreading, cytoskeleton and focal adhesion complex formation, and long-term growth under osteogenic culture conditions.
Materials and Methods
Materials
Films were constructed using PCL (MW=70,000–100,000) and a copolymer of didecylacrylamide and ω-carboxyhexylacrylamide (CAP; MW=22,000). The synthesis of CAP has previously been described 34 and was used as an emulsifying agent in our studies. Polymer surfaces for cell culture were prepared on cover glasses (diameter: 15 mm; Matsunami Glass Industrial Ltd.). Corning® Costar® (Corning, Inc.) cell culture plasticware was used throughout these studies.
Preparation of flat and porous films
The porous honeycomb films of PCL were fabricated as previously described27–32 using water droplets as templates for regular pores. In brief, PCL and CAP (10:1 wt%) were dissolved in chloroform at a concentration of 5 g/L and the polymer mix layered over the cover glasses in a humid environment. The pore sizes of the films were controlled using the following casting conditions: a temperature of 22°C, 85% humidity, a gas flow rate of 1.0 L/min, and a casting volume of 1–15 mL. Films typically exhibited 50% porosity and the pore size was principally controlled by the casting volume with 1 mL giving a film with 3–4-μm pores, 5 mL a film with 5–6-μm pores, 10 mL a film with 10–11-μm pores, and 15 mL a film with 15–16-μm pores. Film thicknesses were equivalent to the scale of the porosity, for example, the film with 3–4-μm pores was ∼3-μm thick, while the 10–11-μm film had a thickness of 10 μm. Flat polymer layers of PCL/CAP were created by casting the solution over the cover glasses and then washing in 1-propanol for 12 min to remove CAP.
Cell culture
Primary calvarial rat osteoblasts isolated as previously described 35 were cultured in the Dulbecco's modified Eagle's medium (Sigma) supplemented with 10% (v/v) fetal calf serum (Sigma), and 2 mMol glutamine (Sigma). Cells from 2nd or 3rd passage were plated onto plane glass, flat PCL, or the porous PCL films at 10,000 cells/well (5000 cells/cm2) in short-term experiments (up to 72 h) to investigate cell morphology and at 20,000 cells/well (10,000 cells/cm2) in studies to osteoblast matrix deposition and mineralization. In long-term experiments, the culture media were exchanged after 48 h to include 5 mM β-glycerophosphate (Sigma), 100 μg/mL L-ascorbic acid phosphate (Wako Ltd.), and 10 nM dexamethasone (Sigma) to induce osteoblastic differentiation over periods up to 35 days. Throughout the course of the experiments, the cultures were fed every 3 days with osteogenic media. All cell cultures were performed at 37°C in 5% CO2, humidified atmosphere.
Immuno/histochemical analysis
To visualize vinculin, cells were fixed in 4% (w/v) parformaldehyde, washed, and incubated with a primary antibody (V9131; Sigma) at 1:200 for 2 h at room temperature. After a wash step, a FITC-goat anti-mouse antibody (Zymed Laboratories) in PBS containing 50 μg/mL TRITC-phalloidin (Sigma) was added to the samples for 1 h in a dark chamber. Following a further wash step, cells were overlayered with VectaShield containing DAPI (Vector Labs) and mounted for microscopy. The alkaline phosphatase activity was localized with Naphthol AS MX-PO4 and Fast Blue RR salt and the mineralized matrix with Alizarin Red staining. Digital images were captured on a Leica DMLB microscope and analyzed with ImageJ. To study cell morphology, individual cells were manually outlined using a digital graphics tablet and the area calculated together with circularity, which was defined as 4π (area/perimeter2). To assess the mineralized matrix, red channel images were split from the randomly acquired RGB images and thresholded. The area of the mineralized matrix was then determined for each binary image. The assessment of vinculin-positive structures was performed by thresholding the grayscale image of stained cells, manual removal of perinuclear structures, then creation of a ROI equivalent to an individual cell as outlined above. Vinculin-positive staining within the ROI was then quantified using the menu command Analyze—> Analyze particles. In addition, confocal laser scanning microscopy (CSLM; Olympus FV-300) was also used to observe the specimens.
Scanning electron microscopy
Samples that had been in culture under osteogenic conditions for 35 days were fixed in 1.25% gluteraldehyde (Polysciences Europe GmbH) in PBS for 30 min. Specimens were then washed in PBS and water before dehydration in graded alcohol through to 100% ethanol and then t-butylalcohol. Critical point drying was then carried out and the samples coated with carbon. All analyses were performed on a scanning electron microscope (Hitachi S-4800) with Energy-dispersive X-ray spectroscopy (EDX) capability. EDX analysis was performed on eight independent sites of matrix mineralization at the surface of the films and eight points inside the pores of the films. Ca/P ratios were calculated and a mean±standard deviation is presented.
Statistical analysis
Similar findings were observed in at least three independent experiments for each experimental approach with data from a single experiment being presented as representative of these studies. Statistical significance between data sets was calculated using either an ANOVA with a Tukey post-test or Kruskal–Wallis one-way analysis of variance with a Bonferroni post-test. Differences were considered significant when p<0.05.
Results
Porous honeycomb films control osteoblast morphology
Primary rat osteoblasts were seeded onto the glass and PCL substrates and cultured for between 16–18 h. Samples were then investigated using immunofluorescence microscopy to localize vinculin, cytoskeletal actin, and nuclear DNA. Cells were observed to adhere, form focal adhesion complexes, and robust actin cytoskeletal structures on all of the substrates investigated in this study (Fig. 1). Analysis of rat osteoblasts cultured on glass, flat and porous films showed that a similar number of cells could be observed after 16–18 h (Fig. 2a). There were, however, significant differences in cell morphology between the surfaces (Fig. 2b, c). Cells cultured on flat PCL were spread more extensively and rounder than cells plated on honeycomb films. There were no significant differences in terms of the cell area or roundness between the different porous surfaces.
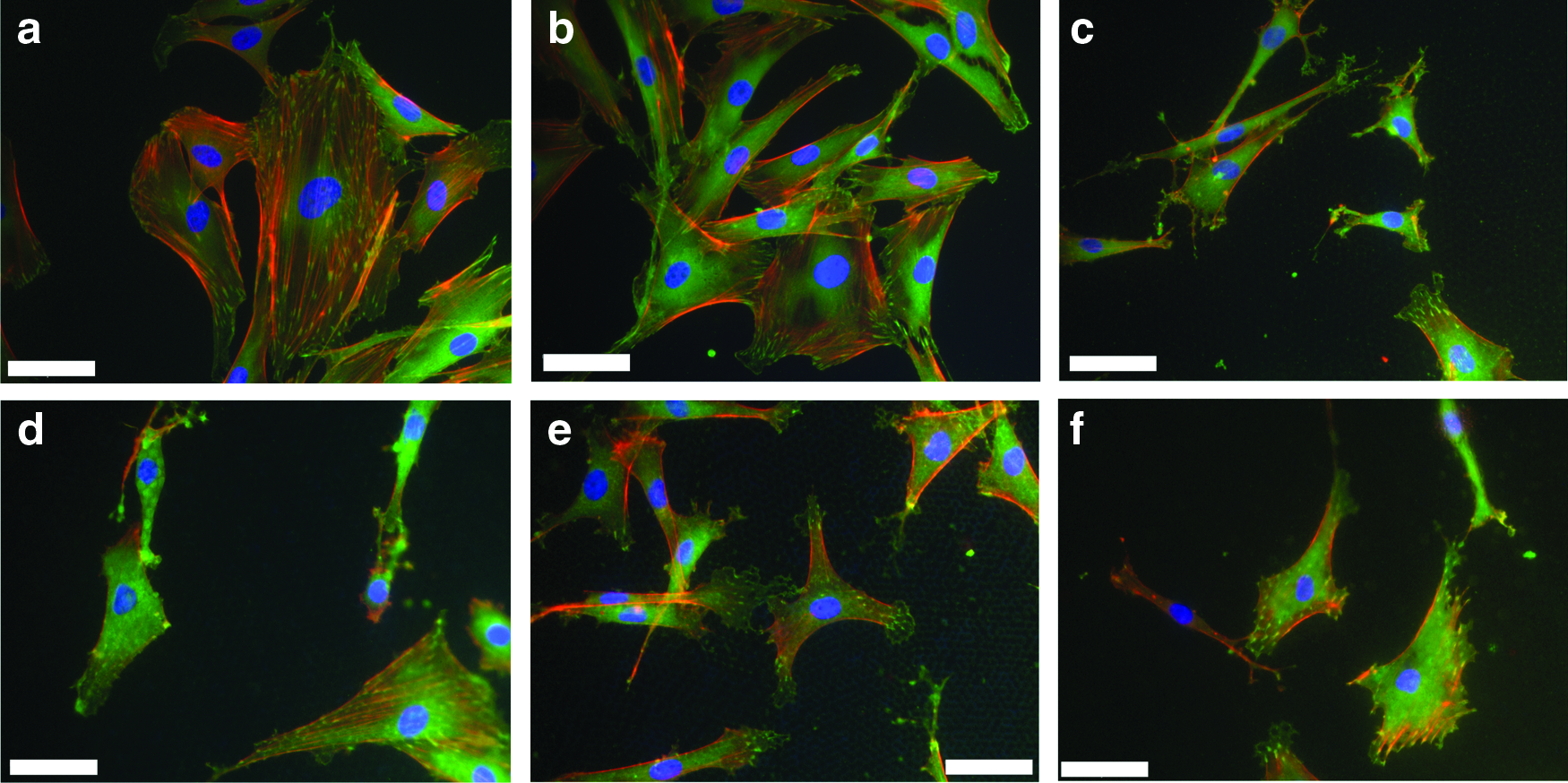
Primary rat osteoblasts adhere and spread on porous PCL films. Cells (5000 cells/cm2) were seeded on
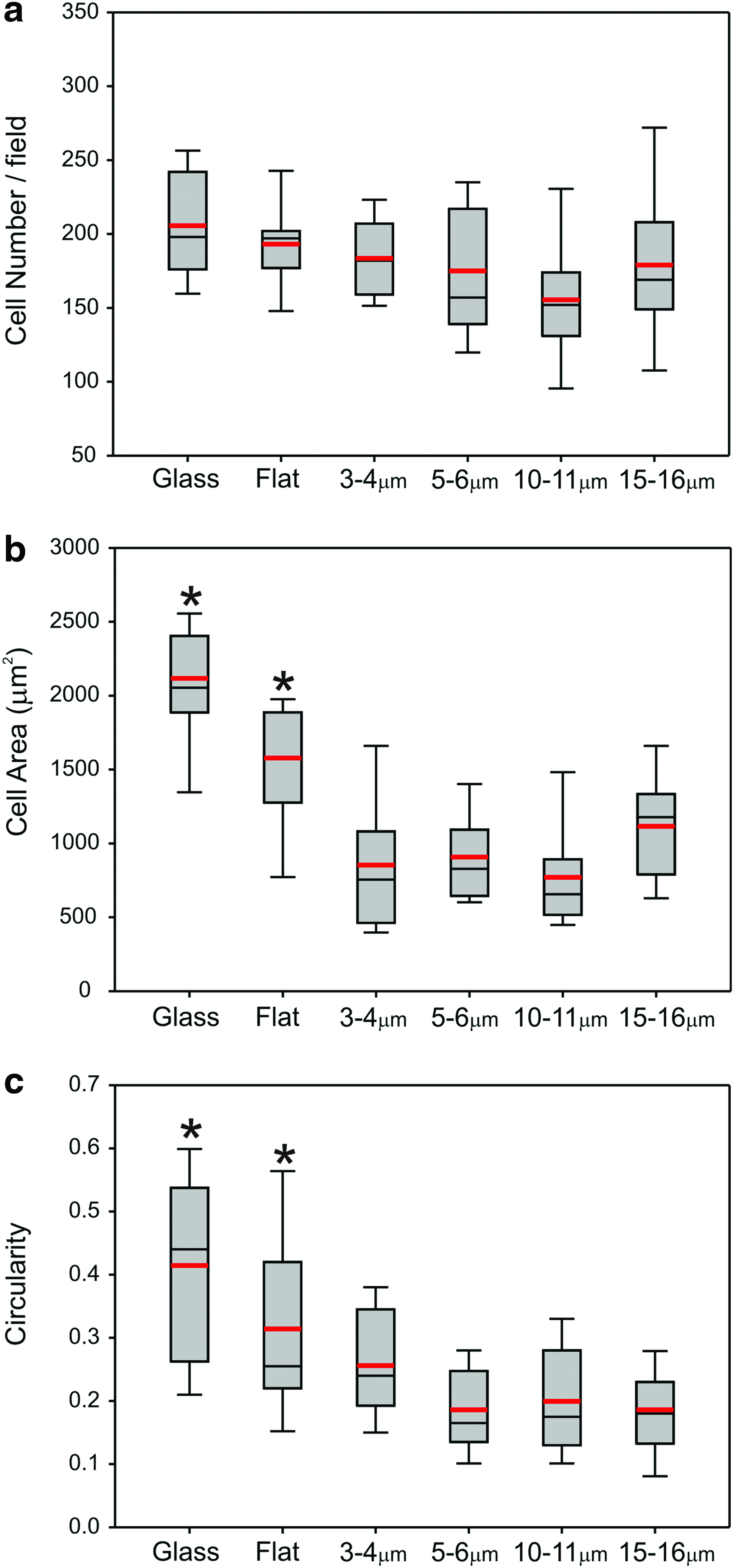
Analysis of immunofluorescence images of cells at 24 h on the PCL surfaces assessed for
Porous PCL films support enhanced mineralized matrix deposition by osteoblasts
Rat osteoblasts were cultured on the substrates for 35 days under osteogenic conditions. Samples were then processed to localize the alkaline phosphatase activity and the formation of the mineralized matrix (Figs. 3a–e and 4a). On all of the different substrates investigated, there was evidence of an extensive alkaline phosphatase activity with cell aggregates displaying intense alkaline phosphatase activity typical of cells undergoing osteogenesis. Alizarin Red S staining revealed the deposition of the mineralized matrix and identified increased abundance of bone-like nodules on the porous surfaces compared to flat PCL. Image analysis of Alizarin Red S staining identified the significant levels of calcium deposition on the 3–4-μm porous PCL film compared to flat PCL with a progressive decrease in the levels of the mineralized matrix as the pore size increased (Fig. 3f).

PCL films support differential mineralized matrix deposition by osteoblasts in vitro. Primary rat ostoblasts (20,000 cells per well/10,000 cells per cm2) were seeded onto
Deposition of a mineralized matrix can be influenced by film topography. After 35 days in osteogenic culture on a honeycomb film with porosity 4–5 μm,
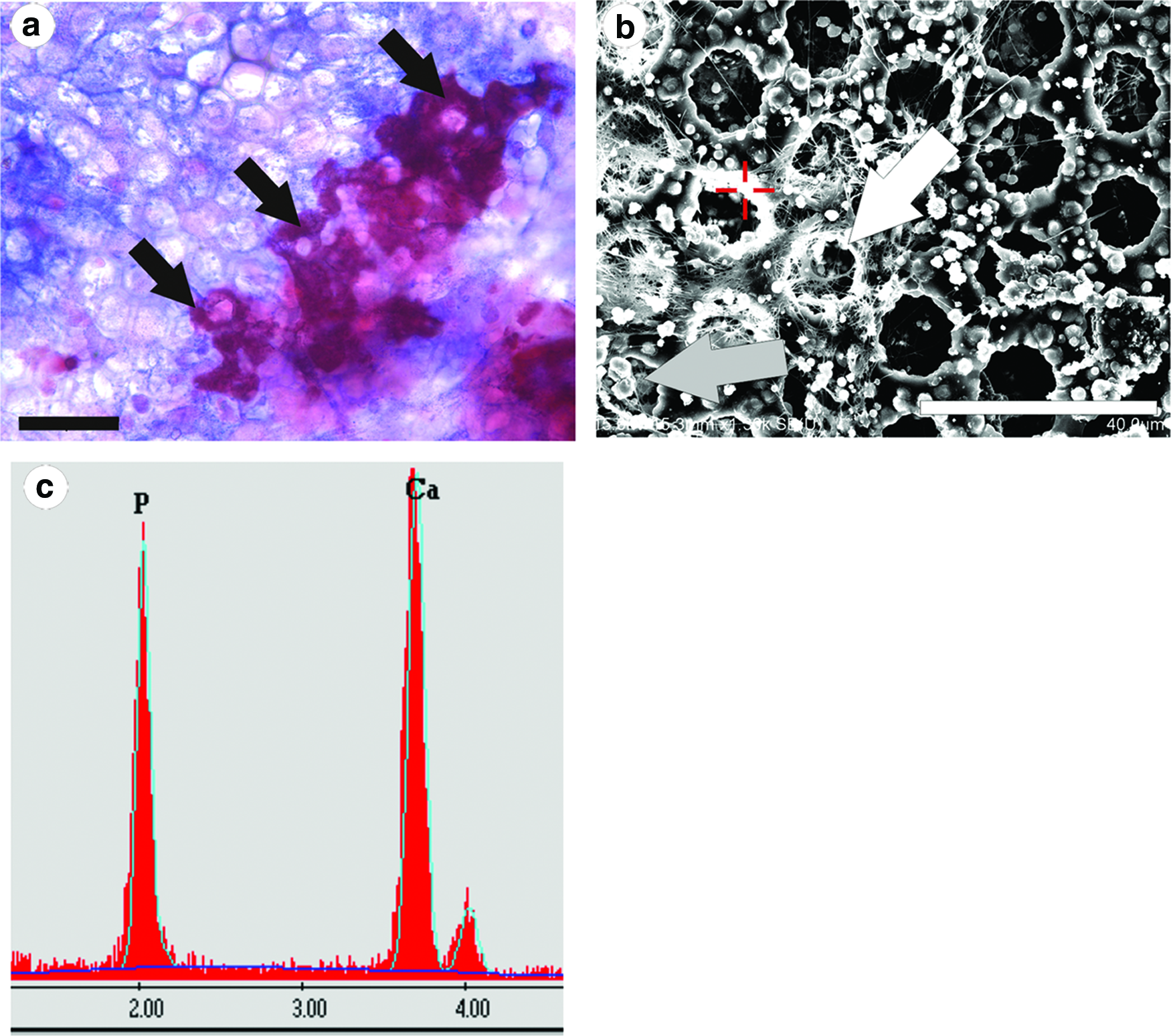
On flat PCL surfaces, Alizarin Red S staining was restricted to globular deposits with little obvious higher order structure other than the aggregation of surrounding cells. On the porous PCL films, it was observed that surface topography could guide matrix deposition with the mineralized matrix mapping to the rims of pores in some areas (Fig. 4a, b). Furthermore, EDX analysis (Fig. 4c) of these deposits typically gave a Ca/P ratio of 1.69±0.08, n=8 (independent samples).
Also evident on the surface of the PCL films and particularly within the pores were spherical matrix deposits, typically 0.5–1 μm in diameter (Figs. 4b and 5a, b). EDX analysis again identified that these structures consisted predominantly of calcium and phosphate (Fig. 5c, d) with a Ca/P ratio of 1.3±0.08, n=8 (independent samples).

Abundant accumulation of calcospherites within the pores of films with small dimensions. SEM analysis of 4–5-μm films
Cell attachment mechanisms on PCL films
It has previously been reported that cell adhesion, as reflected by the size and abundance of vinculin-containing focal adhesion complexes, influences cell differentiation and bone formation.9,24,36 Experiments were therefore performed that looked at vinculin-positive focal adhesion complexes in cells cultured on flat PCL films and also the porous substrates (Fig. 6a, b). Image analysis was performed assessing the abundance of focal structures (1–5 μm2 in size) in cells on each of the surfaces (Fig. 6c). These data show that when normalized for cell area, the number of focal adhesions was significantly reduced in cells grown on the surfaces with 5–6, 10–11, and 15–16-μm pores compared to the flat and 3–4-μm substrates. Further analysis identified the size distribution of focal complexes in cells on each of the surfaces (Fig. 7). On 3–4 and 5–6-μm pore size PCL, there was a clear shift to increased abundance of focal structures in the order of 0–2 μm2, whereas cells on 10–11 and 15–16-μm substrates had elevated levels in the size range 2–4 μm2. The vinculin-positive structures that were assessed in terms of focal complexes were at the surface of the PCL films, but our studies with rat osteoblasts also identified that immunoreactivity could also be found localized to the structure of the pore (Fig. 8a) extending the cell membrane inside the porous film (Fig. 8b).
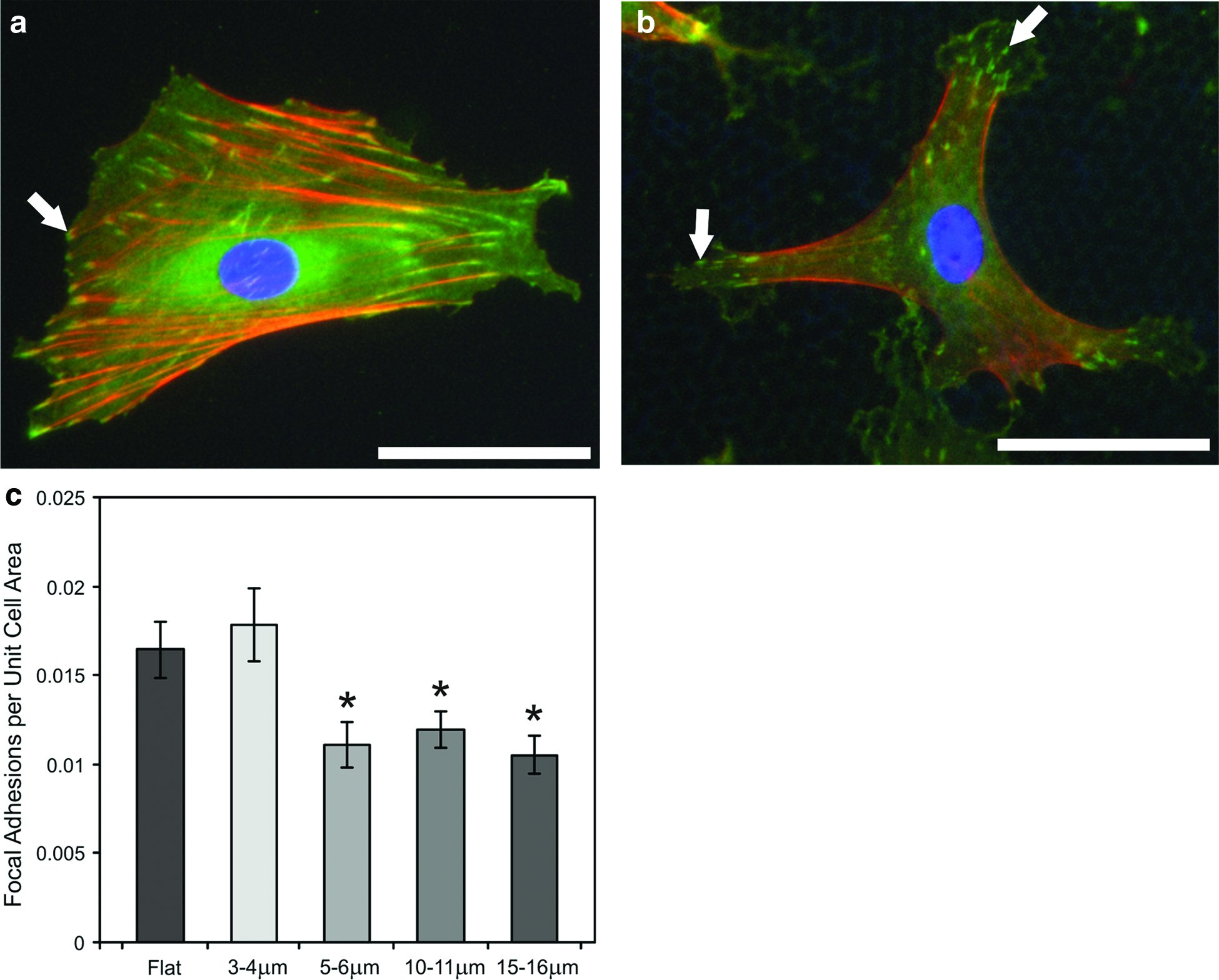
Focal complexes exhibit distinct patterns of distribution on porous PCL. Primary rat osteoblasts adhere and spread on porous PCL films. Cells (11,000 cells/cm2) were seeded on
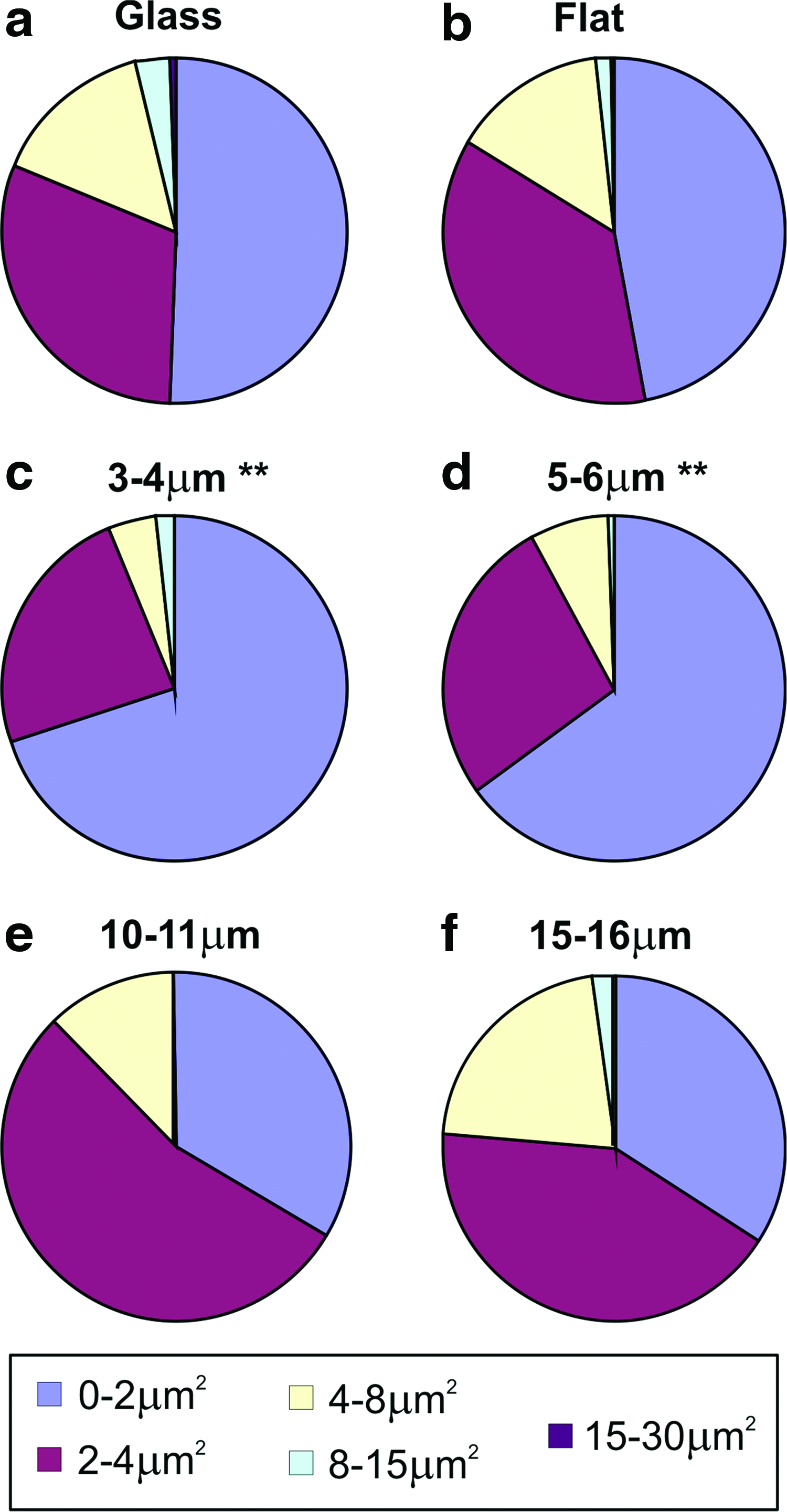
Focal complex size distributions are influenced by the PCL film structure. Image analysis identified the areas of focal complexes formed by 20 individual cells cultured on flat PCL and each of the porous PCL substrates. The numbers of vinculin-positive structures falling within the size boundaries 0–2, 2–4, 4–8, 8–15, and 15–30 μm2 were determined and plotted for

Cell membrane protrusions were identified extending into the porous PCL structure. Staining of cells with anti-vinculin (green) localized to cell–matrix contacts by immunofluorescence and actin cytoskeleton (red) was visualized with TRITC-phalloidin. Nuclei (blue) were identified by DAPI staining.
Discussion
Previous studies have demonstrated how topography at the nano- and microscale has the potential to influence osteoblast differentiation and bone formation.5,9,17,24,37 What is less well understood are the molecular mechanisms that underlie this observation. Much of the focus has been on the role that cell attachment mechanisms may play in the subsequent control of cell activity, correlating topographically influenced formation of cell adhesion structures such as focal adhesion complexes with changes in the cell structure and function.12,13 Evidence suggests that abundant large vinculin-positive adhesion structures and robust cytoskeleton can be linked with cell differentiation 36 and this may well be mechanistically analogous to the cellular response to substrate stiffness. 38 These observations have led to the suggestion that cytoskeletal tensioning in continuum with nuclear scaffolding proteins may directly influence the transcriptional activity and thus alter the cell activity.39,40
The studies presented here used topographically distinct surfaces with defined and highly reproducible structures that ranged in size from subcellular (3–4 μm) to cellular (15–16 μm) to investigate their ability to influence osteoblast interactions with the surface and the formation of the mineralized matrix. There was little difference in the number of cells that adhered to these surfaces at 24 h of culture (Fig. 2a), but parameters of cell morphology (area and circularity) illustrated that cells on the porous films were significantly smaller and less circular than those on the flat film (Fig. 2b, c). Analysis of individual cells confirmed that the surface topographical features of the films used in this study were influencing cytoplasmic projections and spreading of the cells (Fig. 1).
Osteoblasts were cultured on the flat and porous films for periods of up to 35 days under osteogenic culture conditions to induce osteoblast differentiation. On all surfaces investigated, there were abundant numbers of cells that were positive for alkaline phosphatase and evidence of mineralized matrix deposits. In particular, an assessment of the area of calcified bone-like nodules revealed that they were increased on the 3–4 and 5–6-μm porous films compared to the other surfaces investigated in this study (Fig. 3). Observations of the mineralized matrix structures revealed that morphologically they could be divided into two classes: one, where the gross structure had, in part, been dictated by the surface architecture of the substrate material (Fig. 4) or a second spherical morphology that appeared not to be physically related to surface topography (Fig. 5).
The larger deposits of the mineralized matrix exhibited a Ca:P ratio of 1.69, which is close to the nominal value of pure hydroxapatite (1.67) and typical of the values observed for hydroxyapatite formed in vitro by osteoblasts. It was also observed that the macrostructural organization of these bone-like nodules often conformed to the topography of the substrates and, in particular, the lip or rim of the pores. A previous study with these types of films 41 highlighted that proteinaceous deposits selectively accumulate in these regions, and therefore in our cultures, it is likely that concentrations of collagen and other ECM molecules at the pore rim would form foci for mineralization. This organization of proteins and glycosaminoglycans on the membrane surface is perhaps dictated by a combination of altered surface charge and/or nanostructure, which influences the way biomolecules and water interface with the surface.42–44 Further organization of the extracellular matrix is then cell-dependent based upon the structural contact guidance cues.
The spherical deposits of the mineralized matrix were much smaller (0.5–1 μm in diameter) and could be found at the surface, but also particularly inside the pores of the PCL film (Fig. 5). These type of structures have been described previously in vivo and are believed to be a functional unit of mineralization that combines protein, glycosaminoglycans, lipoproteins, and calcium and phosphate ions.45–50 Often referred to as “calcospherites” or “calcospherulites,” these mineralized deposits are typically found in regions of rapid matrix deposition and maturation and are particularly localized to the mineralization front, where they are believed to be a part of the mechanism that leads to a uniform distribution of the mineral phase. EDX analysis revealed a Ca:P ratio of 1.3 in these deposits, which perhaps represents the relative immaturity of these mineralized structures.49,50
To try and gain mechanistic insight into the differential levels of mineralized matrix deposition observed on the different surfaces investigated in this study, we investigated the size, abundance, and organization of vinculin-positive focal adhesion complexes formed in the cells. Our data show that a number of focal adhesion complexes (1–5 μm2 in size) per unit cell area were significantly reduced on surfaces with 5–6, 10–11, and 15–16-μm pores (Fig. 6) and that the relative abundance of smaller focal adhesion complexes is increased in cells on 3–4 and 5–6 μm porosity PCL (Fig. 7). However, these observations do not absolutely correlate with the patterns of the mineralized matrix observed in this study suggesting that substrate-induced changes in cell shape, adhesion structure, and regulation of cell phenotype may not exclusively contribute to the patterns of matrix deposition.
Detailed CSLM revealed that the osteoblasts cultured on these surfaces exhibited globular structures that extended into the porosity of the PCL film (Fig. 8). This was particularly observed with the films that had 3–4 and 5–6-μm pores correlating with smaller vinculin-positive structures. It may be that on these surfaces, weaker cytoskeletal tensioning as a result of smaller vinculin containing anchoring points is more permissive for membrane extension and formation of membrane protrusions. Extensions of the plasma membrane have been previously described as podosomes or invadapodia with functions that range from bone resorption in osteoclasts to tissue invasion in tumor cells.51,52 The common function of these structures is their use to create an extracellular space in which the molecular environment (including pH, balance of proteases/inhibitors) is carefully controlled. Our studies lead us to hypothesize that in an analogous way the PCL films with particular pore dimensions allow osteoblasts to create an extracellular space in which, the conditions particularly favor the deposition and formation of mineralized matrix structures.
Conclusions
These data demonstrate that regular porous films with pore dimensions in the order 3–4 and 5–6 μm support differential osteoblast adhesion. In particular, these surfaces control cell morphology and cell:ECM interactions, which in turn result in enhanced osteoblast deposition of the extracellular matrix and its mineralization. These surfaces provide a reproducible tool to dissect the mechanistic basis for substrate control of cell function and may also provide the basis for new therapeutic implants.
Footnotes
Acknowledgments
This work is supported by Grants-in-Aid and Special Coordination Funds for Promoting Science and Technology of Ministry of Education, Culture, Sports, Science and Technology, Japan and the Nuffield Foundation in the UK.
Disclosure Statement
The authors affirm that they have no conflict of interests in relation to the work presented in this manuscript.