
Abstract
In most cases, the amount of hematopoietic stem and progenitor cells (HSPCs) in a single cord blood (CB) unit is not sufficient for allogenic transplantation of adults. Therefore, two CB units are usually required. The ex vivo expansion of HSPCs from CB in coculture with mesenchymal stroma cells (MSCs) might be an alternative. It was investigated, whether bone marrow-derived MSCs, which have to be obtained in an invasive procedure, introduce a further donor and increases the risk of transmissible infectious diseases for the patient can be replaced by MSCs from amnion, chorion, Wharton's jelly, amniotic fluid, and CB, which can be isolated from placental tissue which is readily available when CB is sampled. In a two-step ex vivo coculture mononuclear cells from cryopreserved CB were cultured with different MSC-feederlayers in a medium supplemented with cytokines (stem cell factor, thrombopoietin [TPO], and granulocyte colony-stimulating factor). Expansion rates were analyzed as well, by long-term culture-initiating cell (LTC-IC) and colony-forming unit (CFU) assays, as by measuring CD34+- and CD45+-cells. Due to the comparably low number of 5×102 to 1×104 CD34+-cells per cm2 MSC-monolayer, we observed comparably high expansion rates from 80 to 391,000 for CFU, 70 to 313,000 for CD34+-, and 200 to 352,000 for CD45+-cells. Expansion of LTC-IC was partly observed. Compared to the literature, we found a better expansion rate of CD34+-cells with MSCs from all different sources. This is probably due to the comparably low number of 5×102 to 1×10 CD34+-cells per cm2 MSC-monolayer we used. Comparably, high expansion rates were observed from 80 to 391,000 for CFUs, 70 to 313,000 for CD34+-, and 200 to 352,000 for CD45+-cells. However, the expansion of CD34+-cells was significantly more effective with MSCs from bone marrow compared to MSCs from amnion, chorion, and Wharton's jelly. The comparison of MSCs from bone marrow with MSCs from CB and amniotic fluid showed no significant difference. We conclude that MSCs from placental tissues might be useful in the expansion of HSPCs, at least if low numbers of CD34+-cells per cm2 MSC-monolayer and a high TPO concentration are implemented in the expansion culture.
Introduction
We investigated, whether bone marrow-derived MCSs can be replaced by MSCs from amnion, chorion, Wharton's jelly, amniotic fluid, and CB, which are readily available and if these MSCs are suitable for ex vivo expansion of hematopoietic stem cells. It should be analyzed, if there are differences between fresh prepared and cryopreserved MSCs regarding the expansion data and which MSC-source yielded the optimal expansion.
Materials and Methods
Isolation of MSCs
MSCs from CB, amnion, chorion, Wharton's jelly, and amniotic fluid were isolated from healthy donors. MSCs from CB were isolated from mononuclear cells (MNCs). These cells were collected from the interface generated after Ficoll (Biotest AG) density separation and washed three times with phosphate-buffered saline (PBS) (Gibco Invitrogen Corp.). The MNCs from a CB unit were cultured in six-well-plates (Greiner Bio-One, GmbH), the cell number varies from 4.8×106–4.8×107 cells. Amnion and chorion were collected from placenta under sterile conditions, washed with PBS, supplemented with penicillin (104 U/mL)/streptomycin (10 mg/mL; StemCell Technologies), and minced in small pieces. These fragments were cultured in six-well-plates, 3 mL per well, without further treatment. Umbilical cord was also collected from placenta and the preparation was similar to amnion and chorion. About 2 cm fragments were cut alongside, vessels were removed carefully; Wharton's jelly was minced in small pieces of about 1 mm length and cultured in six-well-plates. Amniotic fluid was collected from pregnant women with a great volume of amniotic fluid (500 mL–2 L), which had to be reduced. After centrifugation, cells were cultured in 6-well-plates; the cell number varies from 2.9×106–6.2×106 cells. Bone marrow was isolated from patients undergoing hip arthroplasty between the ages of 60–80. For preparation, bone marrow was stored at room temperature for 20 min, the thereby developed fatlayer was discarded and so bone marrow was cultured in six-well-plates without further treatment. As soon as MSCs from amnion, chorion, and Wharton's jelly were visible, fragments were discarded. After 2–3 days, nonadherent cells were removed for cultivation of MSCs, medium was changed completely, and adherent cell culture continued in MSC culture medium until cells achieved nearly 80–90% confluence. Adherent cells were detached by trypsin-EDTA (0.25%; StemCell Technologies) and passaged into new six-well-plates.
Cryopreservation procedure
Adherent cells were detached by trypsin-EDTA, transferred to a 15 mL tube, centrifugated, and the supernatant was discarded. The pellet was resuspended in 1 mL 4°C cold cryopreservation medium and transferred to cryopreservation tubes. The cryopreservation medium consisted 70% fetal calf serum (Biochrom), 20% culture medium (StemCell Technologies), and 10% dimethylsulfoxide (DMSO). The tubes were stored for 1–2 h at −20°C, and then overnight at −80°C. For long-term storage, the tubes were taken to liquid nitrogen.
The cryopreserved cells were thawed (37°C), centrifugated, the supernatant (with DMSO) was discarded, and the pellet was resuspended in culture medium.
Characterization of MSCs
Flow cytometry
Flow cytometric analyses were performed to determine the phenotype of the different MSC-types (fresh and thawed). MSCs were tested for CD14, CD34, CD45, HLA-DR, CD29, CD90, CD73, CD44, HLA-I, and CD105 expression. With the exception of anti-CD105 FITC (Exbio), all antibodies were purchased from Becton Dickinson.
Differentiation
MSCs were incubated with special differentiation medium according to the manufacturer's instructions.
Ex vivo expansion of HSPCs with MSCs
MNCs from cryopreserved CB were plated over different MSC-feederlayers in a coculture medium: α-minimum essential medium (Hyclone Laboratories) supplemented with penicillin/(104 U/mL) streptomycin (10 mg/mL; StemCell Technologies), and 10% irradiated fetal bovine serum (Fluka, Sigma-Aldrich) and containing 100 ng/mL each of granulocyte colony-stimulating factor (G-CSF; StemCell Technologies), stem cell factor (SCF; StemCell Technologies) and thrombopoietin (TPO; StemCell Technologies). Coculture was first performed for 7 days. At day 7, the CD34+-data of the nonadherent cells were analyzed. Subsequently these cells were plated with the same volume of CD34+-data as at the beginning over fresh confluent MSC-feederlayer. Adding fresh medium, culture continued for additional 7 days (14 days total). At day 14, nonadherent cells from both cultures were harvested and CD34+-, colony-forming unit (CFU)- and long-term culture-initiating cell (LTC-IC)-data were analyzed. Further adherent cells from the MSC-coculture were released by trypsin and CD34+-cells were analyzed by flow cytometry.
Flow cytometry
For analyzing CD34+-cells were stained with 10 μL phycoerythrin-conjugated anti-CD34 (Becton Dickinson), 10 μL fluorescein isothiocyanate-conjugated anti-CD45 (Becton Dickinson), 5 μL 7-AAD (Becton Dickinson) and incubated for 15 min at room temperature. 7-AAD was used to determine dead cells. Afterwards, 1000 μL FACSFlow (Becton Dickinson) was added to each sample and measuring was according to the manufacturer's instruction.
LTC-IC assay
Cultures were plated over the stroma-cellline M2-10B4, which supports human myelopoiesis in long term assays. For each sample, three LTC-IC assays were performed and cultured for 5–6 weeks, to identify multipotent progenitor cells. The assays were refed weekly with LTC-IC medium: myelocult H5100 (StemCell Technologies) and hydrocortisone (StemCell Technologies). After that time cells were harvested and CFU assays were started. About 20 days later, colonies were scored “positive” or “negative” using a microscope.
CFU assay
For analyzing, cells were diluted with Iscoves MDM+2% FCS (StemCell Technologies) according to the manufacturer's instructions of StemCell Technologies. The diluted suspension was mixed 1:10 with methylcellulose medium (Methocult GF H4435; StemCell Technologies). For each sample, duplicate CFU assays were performed. The assays were cultured for 14–16 days in a humidified incubator (37°C, 5% CO2 in air atmosphere). Colonies were enumerated using a microscope.
Statistical methods
We analyzed data using IBM® SPSS® Statistics (release 19.0.0.1; IBM). Normal distribution of variables was tested using Lilliefors- and Shapiro-Wilks-test. Statistical analysis was done using nonparametric methods if results were not distributed normally. Comparisons between groups were performed by Mann–Whitney U-test. We considered differences to be significant when p-values were<0.05.
Results
Isolation of MSCs
MSCs from CB could be isolated in 14% and passaged for more than 10 times. Also MSCs from amniotic fluid, chorion, Wharton's jelly, and bone marrow were passaged over 10 times, but MSCs from Amnion stopped growing very soon after isolation.
Cryopreservation of MSCs
After thawing, MSCs were seeded in 6-well-plates supplemented with culture medium until they achieved nearly 80–90% confluence. For the experiments, it was not relevant to count the cells.
In previous experiments, the recovery rate of CD34+-cells was 83.3%±15.5%, the vitality of CD34+-cells was 91.4%±6.6%. 11
Characterization of MSCs
Flow cytometry
MSCs from all sources showed a unique pattern in multiparameter flow cytometry. They were negative for the markers CD14, CD34, CD45, and HLA-DR and positive for CD29, CD90, CD73, CD44, HLA-I, and CD105. These analyses were performed with fresh and cryopreserved MSCs and were conserved to repeated cell passages (data not shown).
Differentiation
MSCs differentiated in osteo-, adipo-, and chondrocytes.
Ex vivo expansion of HSPCs with MSCs
To find the best MSC-source for HSPC-expansion, the experiments were started in parallel, using MNCs from one CB with all different kinds of MSCs at one given time. The number of CD34+-cells varied between 5×102 to 1×104 per cm2 MSC-monolayer, depending on the quality of the CB samples. The control samples were started analog, without MSCs. After 14 days of ex vivo coculture, the cells were harvested and the cell number of CD34+- and CD45+-cells was determined. LTC-IC and CFU assays were started.
LTC-IC assay
LTC-IC assays were started with MNCs from CB at the beginning of the coculture experiments and after the harvest with increased stem- and progenitor cells after 14 days of MSC-coculture. MSCs from amniotic fluid and CB are not shown in Figure 1, because after the harvest no colonies were grown. The results (Fig. 1) showed that expansion of LTC-IC was partly observed.
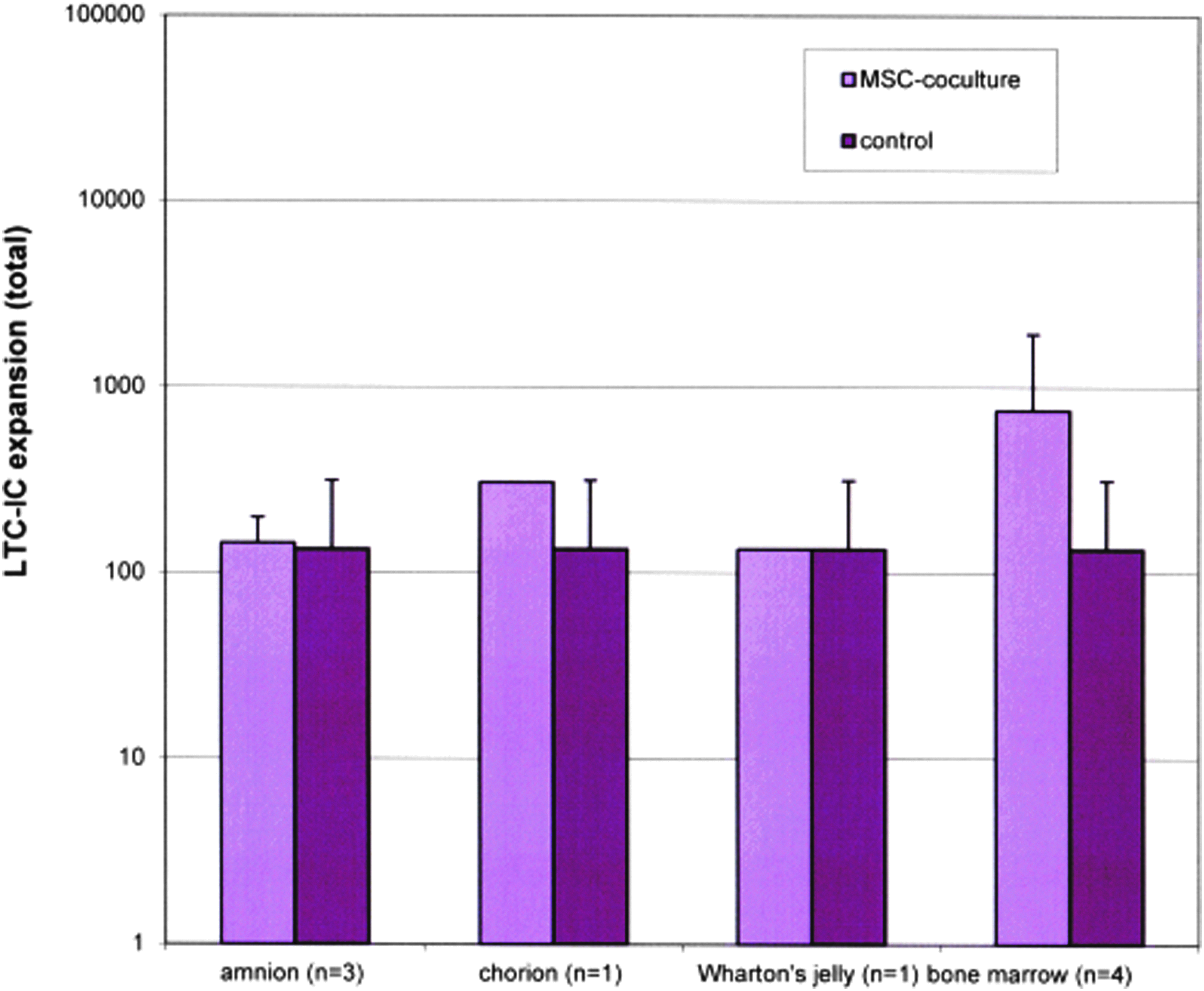
Long-term culture-initiating cell (LTC-IC) expansions; control samples without mesenchymal stroma cells (MSCs), n=4; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
CFU assay
CFU assays were started with nonadherent cells at the beginning (day 0) and at the end (day 14) of the coculture experiments. It was distinguished between fresh prepared and cryopreserved samples and different cell volumes at the beginning (a and b). The CFU-expansion data (mean±standard deviation) is shown in Figure 2 (fresh prepared) and Figure 3 (cryopreserved). The expansion data of the cryopreserved samples was highest with MSCs from bone marrow (a: 5.1-fold; b: 13.7-fold). The expansion data of the remaining MSC sources were 2.3-fold to 3.8-fold (a) and 1.6-fold to 4.8-fold (b).
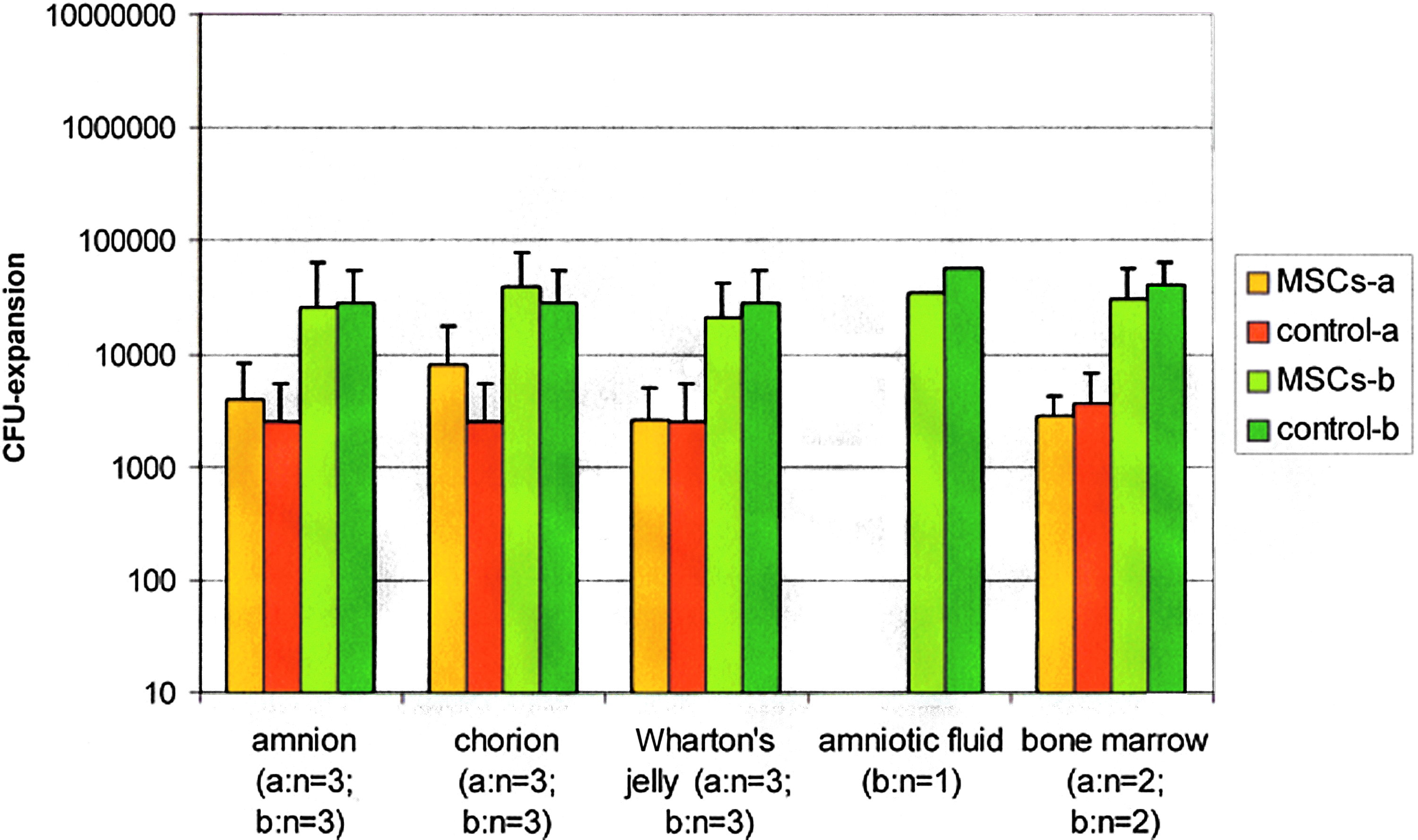
Colony-forming unit (CFU)-expansions of all coculture samples (fresh prepared) with MSCs of different sources (see figure) and mononuclear cells (MNCs) after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
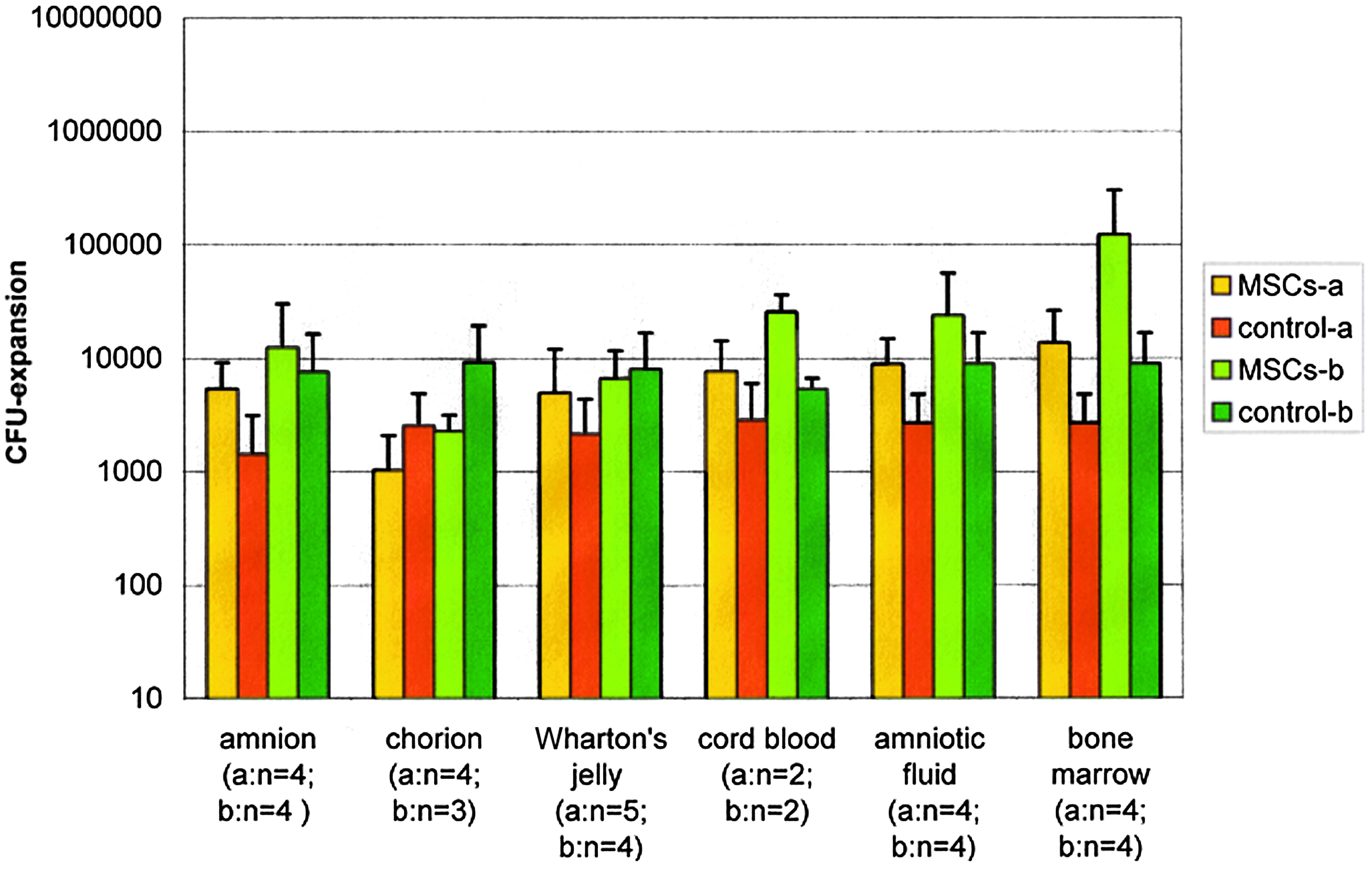
CFU-expansions of all coculture samples (cryopreserved) with MSCs of different sources (see figure) and MNCs after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
The comparison of CFU-expansions with fresh and cryopreserved MSC-samples and with MSCs from different sources using the Mann–Whitney U-test showed no significant differences (data not shown). The comparison of the CFU-expansion with different initial numbers of CD34+-cells/cm2 MSC-monolayer represented by a and b (see Table 1) showed a significant better expansion in assays with lower initial number of CD34+-cells/cm2 MSC monolayer.
Mean±standard deviation.
Amniotic fluid: only one fresh sample.
ND, not done; CFU, colony-forming unit.
CD34+-expansions
CD34+-expansion data (mean±standard deviation) is shown in Figure 4 (fresh prepared) and Figure 5 (cryopreserved). The comparison of CD34+-expansions with fresh and cryopreserved MSC-samples using the Mann–Whitney U-test showed no significant differences (data not shown). The comparison of all samples, not distinguished between fresh and cryopreserved MSCs showed significant differences in the expansion of CD34+-cells between assays using MSCs from bone marrow and Wharton's jelly (p<0.002), bone marrow, and chorion (p<0.0004) and also among bone marrow and amnion (p<0.0004).
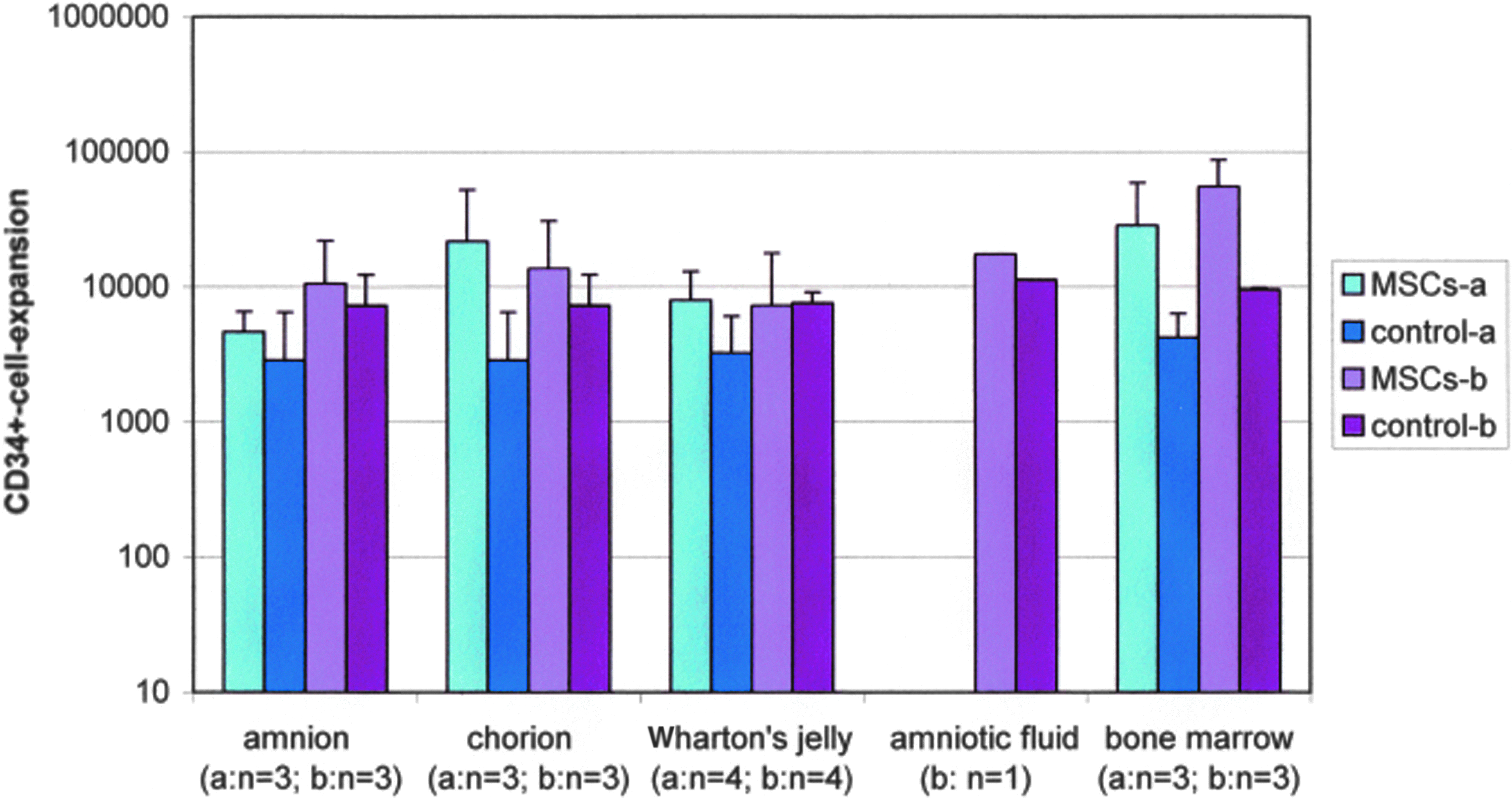
CD34+-expansions of all coculture samples (fresh prepared) with MSCs of different sources (see figure) and MNCs after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
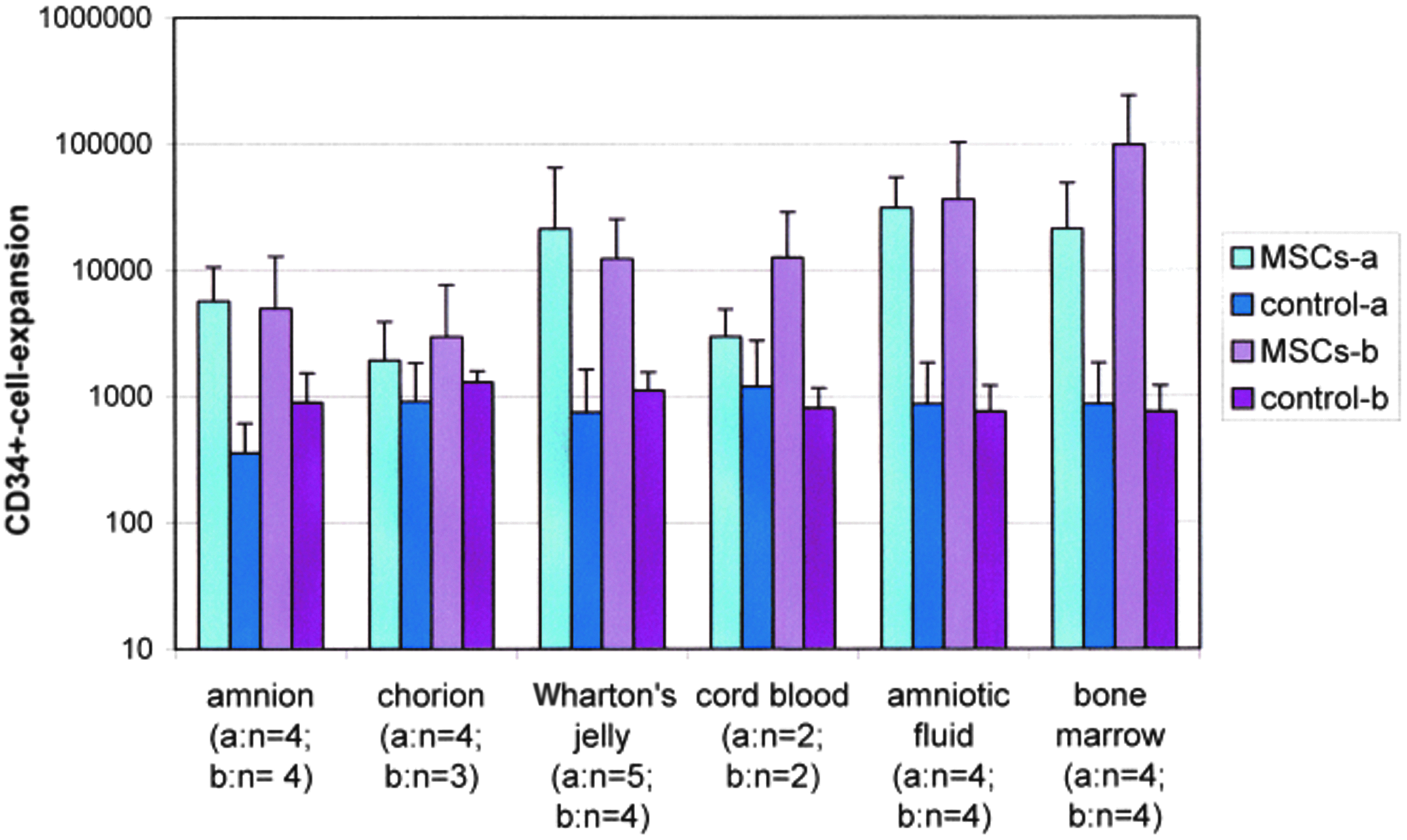
CD34+-expansions of all coculture samples (cryopreserved) with MSCs of different sources (see figure) and MNCs after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
The comparison of different initial numbers of CD34+-cells represented by a and b (see Table 2) showed no significant difference in CD34+-expansion (data not shown).
Mean±standard deviation.
Amniotic fluid: only one fresh sample.
CD45+-expansions
The comparison of the expansion of CD45+-cells showed also no significant differences between assays using fresh and cryopreserved MSCs (Figs. 6 and 7). Looking at all samples together, we observed a significant difference in the expansion of CD45+-cells between assays using MSCs from bone marrow and Wharton's jelly (p<0.012), bone marrow, and chorion (p<0.007) and among amniotic fluid and chorion (p<0.016). The comparison of the expansion of CD45+-cells in assays with different initial numbers of CD34+-cells represented by a and b (Table 3) showed a significance of p<0.0004 with better expansion results in assays with a lower initial number of CD34+-cells.
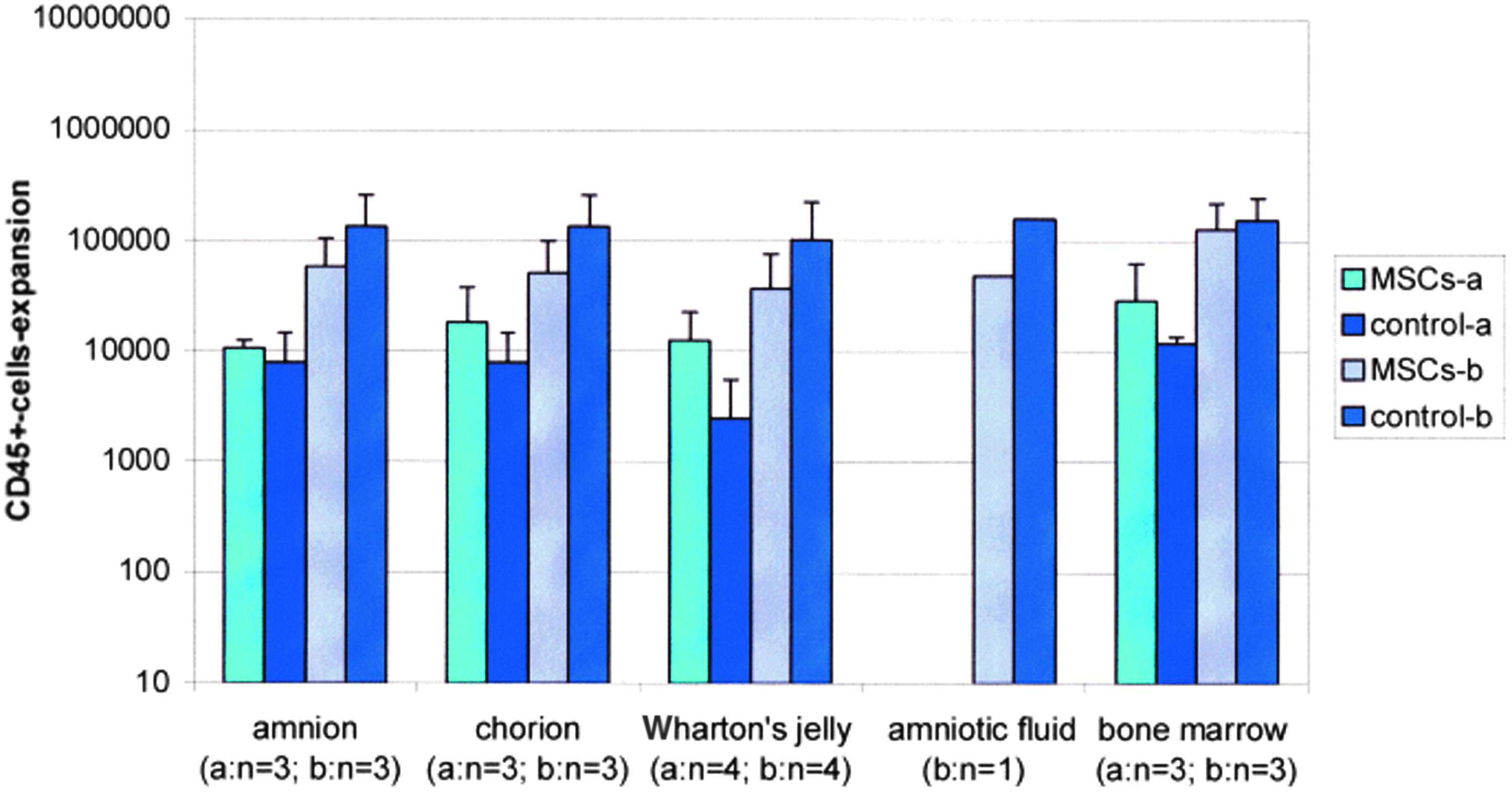
CD45+-expansions of all coculture samples (fresh prepared) with MSCs of different sources (see figure) and MNCs after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea

CD45+-expansions of all coculture samples (cryopreserved) with MSCs of different sources (see figure) and MNCs after 14 days (n=number of samples); control samples: without MSCs; (mean±standard deviation). Color images available online at www.liebertpub.com/tea
Mean±standard deviation.
Amniotic fluid: only one fresh sample.
In Tables 1–3, total numbers of CFUs (Table 1), CD34+-cells (Table 2) and CD45+-cells (Table 3) before and after 14 days of coculture with MSCs from different sources are shown; (mean±standard deviation). In Figures 1–7, the calculated expansions are shown.
Discussion
In preliminary studies, HSPCs from CB were successfully used for ex vivo expansion in coculture with MSCs from bone marrow. At the moment, bone marrow is the main source for MSCs, but it is not ideal for transplantation. An invasive procedure is necessary for isolation. The introduction of a further donor increases the risk of transmissible infectious diseases for the patient. At birth placental tissue, from which MSCs can be easily obtained, is basically available with each CB unit. It was investigated, whether these MSCs are suitable as feederlayers in ex vivo expansion of hematopoietic stem and progenitor cells. Data presented here demonstrate that MSCs from amnion, chorion, Wharton's jelly, amniotic fluid, and CB are basically useful as feederlayers for ex vivo expansion. Compared to the literature,1,12,13 we found a better expansion rate of CD34+-cells with MSCs from all different sources. Robinson et al. found an expansion of>40-fold for total nuclear cells (TNC), 20-fold for CFU using purified CD133+-cells from CB with MSCs from bone marrow. 1 The same group found an expansion of 8-fold for CD34+-cells, >200-fold for CFU and 10-fold for TNC using MNC from CB with MSCs from bone marrow where they cultured 8×104 CD34+-cells per cm2 MSC-monolayer. 1 This group used the same setting of recombinant growth factors, which was used in the expansion cultures for this article: TPO, SCF, and G-CSF, each 100 ng/mL. 1 McNiece et al. performed ex vivo expansion of CB MNCs on MSCs from bone marrow with a density of 2.8 to 4.5×103 CD34+-cells per cm2 MSC-monolayer using SCF, G-CSF and megakaryocyte growth and differentiation factor, each 100 ng/mL. 12 This resulted in a 10- to 20-fold expansion of TNC, a 7- to 18-fold expansion of CFU and a 16- to 37-fold expansion of CD34+-cells. 12 Li et al. expanded purified CD34+-cells from peripheral blood stem cells using MSCs from bone marrow, the density of CD34+-cells on the MSC-monolayer is not given in their article. 13 They used TPO 10 ng/mL, Flt-3 ligand 50 ng/mL, SCF 50 ng/mL and IL-6 10 ng/mL in their expansion cultures. They found expansion rates of 242-fold for TNC, 126-fold for CFU and 92-fold for CD34+-cells. 13 In the present work, comparably high expansion rates were observed from 80 to 391,000 for CFUs, 70 to 313,000 for CD34+- and 200 to 352,000 for CD45+-cells. This is probably due to a combination of two culture conditions which were one or both different compared to the studies described above1,12,13: a comparably low number of 5×102 to 1×104 CD34+-cells per cm2 MSC-monolayer and a comparably high concentration of TPO, which is known to have a major antiapoptotic effect on HPSCs. 14 However, the expansion of CD34+-cells was significantly more effective with MSCs from bone marrow compared to MSCs from amnion, chorion, and Wharton's jelly. The comparison of MSCs from bone marrow with MSCs from CB and amniotic fluid showed no significant difference. We conclude that MSCs from placental tissues might be useful in the expansion of HSPCs, at least if low numbers of CD34+-cells per cm2 MSC-monolayer and a high TPO concentration are implemented in the expansion culture.
The expansion of LTC-IC was successful in a small proportion of the experiments, contrary to several studies, which report no expansion of hematopoietic progenitor cells after 6 weeks LTC-IC assay.1,15
Only coculture expansion experiments with fresh MSC were reported in literature,1,13,16 but these data show cultures using cryopreserved MSCs, which yielded higher expansion rates and more often positive expansions compared to fresh MSC samples. These results could be positive for clinical use, because cryopreserved MSCs are always available; however, fresh prepared MSCs are not.
Furthermore, the clinical use of fetal MSCs compared to adult MSCs is more advantageous, because of a better HLA-compatibility 17 and a higher potential of expansion in vitro. 18
Footnotes
Disclosure Statement
No competing financial interests exist.