
Abstract
Cementum covering the tooth root provides attachment for the tooth proper to the surrounding alveolar bone via non-mineralized periodontal ligament (PDL). Cementum protein 1 (CEMP1) has been shown to induce a cementoblastic phenotype in cementoblast precursors cells of PDL. Oxygen availability is a critical signal for correct development of many tissues; however, its role in tooth root and periodontium development remains poorly understood. In this study, we demonstrated that reduced oxygen tension increased CEMP1 expression, mineral deposition, and alkaline phosphatase activity in human dental stem cells such as PDL stem cells and periapical follicular stem cells. Since an oxemic state is transduced by the transcription factor, hypoxia-inducible factor-1 (HIF-1), we performed experiments to determine whether this protein was responsible for the observed changes. We noted that when HIF-1 was activated by gene introduction or chemically, CEMP1 expression and mineralization increased. In contrast, when HIF-1α was silenced, CEMP1 expression and mineralization did not increase in vitro. Furthermore, we showed for the first time that mouse tooth root and periodontium development occurs partly under hypoxic conditions, particularly at the apical part and latently at the PDL space in vivo. Desferrioxamine, an HIF-1 stimulator, enhances CEMP1 expression in the mouse PDL space, suggesting that hypoxia affects cementogenesis of PDL cells lining the surface of the developing tooth root in an HIF-1-dependent manner. These results suggest that HIF-1 activators may have the ability to stimulate regeneration of the tooth root and cementum formation.
Introduction
T
Despite the important role that cementum plays in the reparative process after periodontal disease, very little is known about its progenitors, cementoblasts. Initially, cementum protein 1 (CEMP1) was isolated and characterized as a human cementoblastoma-derived protein 5 and in situ hybridization revealed that CEMP1 mRNA was expressed in the cementoblast cell layer lining the cementoid surface of cementum. 6 Transfection of the CEMP1 coding region (GenBank Accession No. NM_001048212) into human gingival fibroblasts showed that CEMP1 promotes cell attachment and osteoblastic and/or cementoblastic cell differentiation.7,8 Recently, it was found that purified human recombinant CEMP1 has strong affinity for hydroxyapatite and promotes octacalcium phosphate (OCP) crystal growth as a part of the biomineralization process. 9 OCP is presumed to be a necessary precursor of biological apatites in hard tissue calcifications. 9 Furthermore, immunohistochemical staining of human periodontal tissue confirmed that the CEMP1 protein localized to cementoblast cells as well as to restricted PDL cell subpopulations (cementoblast precursors) in the PDL space (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). These results strongly suggest that CEMP1 may regulate cementoblast differentiation during development and in the adult PDL space.
Oxygen is both a potent biochemical signaling molecule and a major gene regulator during an organism's development. 10 During development, oxygen participates in the intricate balance between cellular proliferation and commitment toward differentiation. Furthermore, oxygen concentration significantly influences the way in which cells remodel their environment, for example, matrix regulation.11,12 The response to hypoxia is cell-type dependent, but oxygen affects critical cellular processes such as adhesion, proliferation, metabolism, apoptosis, growth factor expression, extracellular matrix secretion, and differentiation patterns.10,13–16 Hence, oxygen affects cellular characteristics and tissue-remodeling processes, and may have the potential to direct cell fate.17–19 The hypoxia-inducible factor (HIF) pathway is the central pathway for sensing and responding to changes in local oxygen availability in a wide variety of organisms. 20 HIF-1α dimerizes with HIF-1β (ARNT) to form the HIF-1 transcription factor, and HIF-2 is similarly formed via the association of HIF-2α and HIF-1β: Both HIF-1 and HIF-2 activate transcription. HIF-1 is known to play an important role in modulating cell fate, including osteogenic differentiation.20,21 However, little is known about the role of hypoxia and HIFs in tooth development.
Human dental stem cells (DSCs) are recognized as a promising source for cell-based, autologous tissue regeneration.22,23 Thus, understanding the cellular and molecular biology of these cell populations is the current focus of translational research. Accumulating evidence suggests that local oxygen tension is a critical modulator of mesenchymal cell growth and differentiation. 17 The purpose of this study was to investigate the effects of hypoxia on cementoblastic differentiation of human DSCs, including periapical follicular stem cells (PAFSCs) and periodontal ligament stem cells (PDLSCs). We hypothesized that hypoxia affects the differentiation potential of DSCs and tooth development. Here, we show for the first time that mouse tooth root and periodontium development occurs partly in a hypoxic state, particularly at the apical part and latently at the PDL space. In addition, we demonstrate that hypoxia stimulates CEMP1 expression, at least partly, via a pathway involving HIF-1 in vitro and that mice treated with desferrioxamine (DFO), a chemical inducer of HIF-1 protein, show an increase in CEMP1 expression during tooth root development in vivo. Our data indicate that hypoxia stimulates CEMP1 expression and enhances cementum formation in a developing tooth root in an HIF-1-dependent manner.
Materials and Methods
Primary cell culture from human dental tissue
Human third molars were collected from healthy young men (18–25 years) under the protocol approved by the institutional review board of Seoul National University Dental Hospital, Seoul, Korea (IRB Number 05004). Periapical follicles were separated from the root of the tooth germs at the root-forming stage, and they were aseptically dissected to obtain human PAFSCs. PDL tissues were separated from the surface of the root to obtain human PDLSCs. Separated tissues were digested in a solution of 3 mg/mL collagenase type I (Worthington Biochem, Freehold, NJ) and 4 mg/mL dispase (Boehringer, Mannheim, Germany) for 1 h at 37°C. Single-cell suspensions were obtained by passing the cells through a 40 μm strainer (BD Labware, Frankin Lakes, NJ) and cultured in alpha modification of Eagle's medium (GIBCO BRL, Grand Island, NY) supplemented with 10% fetal bovine serum (GIBCO BRL), 100 μM ascorbic acid 2-phosphate (Sigma Aldrich, St. Louis, MO), 2 mM glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin (GIBCO BRL) (growth medium) and incubated at 37°C in 5% CO2 (normoxia). All primary cells used in this study were passage 2 or 3 cells. Hypoxia was induced by exposure to a gas mixture of 8% O2, 5% CO2, and 85% N2. Hypoxyprobe™-1 kit (Hypoxyprobe, Inc., Burlington, MA) was used to detect the in vitro cellular response to hypoxia. Cells were incubated under hypoxia for 48 h in the presence of 200 μM Hypoxyprobe™-1 followed by immunocytochemical staining. Cellular hypoxia was also confirmed by immunocytochemical staining of HIF-1α.
Flow cytometric analysis
To characterize the immunophenotype of human PDLSCs and PAFSCs, expression of mesenchymal stem cell (MSC)-associated surface markers at passage 3 was analyzed by flow cytometry as previously described. 24 Briefly, cells in their third passage (1.0×106 cells) were fixed with 4% PFA for 10 min and resuspended in 1% bovine serum albumin (BSA) phosphate-buffered saline (PBS) for blocking for 30 min. Cells were then incubated with specific antibodies for CD13 (0.1 μg/mL), CD34 (0.1 μg/mL), CD90 (0.05 μg/mL), or CD146 (0.1 μg/mL) at 4°C for 1 h, followed by the addition of fluorescently labeled secondary antibodies at room temperature for 1 h. All antibodies were purchased from BD Biosciences (San Jose, CA). The percentage of CD13-, CD90-, and CD146-positive and CD34-negative cells was measured with an FACS Calibur (Becton Dickinson, CA), and the results were analyzed using Cell Quest Pro software (Becton Dickinson).
Mineralization induction and Alizarin red S staining
For mineralization, cells were cultured in osteogenic medium (growth medium supplemented with 10 mM β-glycerophosphate, 50 μg/mL ascorbic acid, and 100 nM dexamethasone) for 3 weeks. For undifferentiated samples, cells were maintained in growth medium for the same duration. Mineral nodule formation was observed by staining the cultures with 40 mM Alizarin red S (pH 4.2), and the amount of Alizarin red S that bound to the minerals in each dish was quantified by destaining the samples in 10 mM sodium phosphate containing 10% cetylpyridinium chloride (pH 7.0) for 15 min at room temperature. The amount of Alizarin red S in the destaining solution was measured at OD 562 nm and compared with a standard solution of the dye. 25
Chondrogenic and adipogenic differentiation
To induce chondrogenic or adipogenic differentiation, cells were cultured in StemPro Chondrogenic or StemPro Adipogenic differentiation media (GIBCO BRL), respectively, with appropriate supplements. At week 2 of postchondrogenic or postadipogenic induction, cells were washed with PBS and fixed in 4% paraformaldehyde for 10 min. Cells were stained with 1% Alcian Blue (Sigma Aldrich) and 0.3% Oil Red O dye (Sigma Aldrich) to detect proteoglycans and fat vacuoles as an indication of chondrogenic and adipogenic differentiation, respectively. Cells were visualized under an inverted light microscope (Olympus U-SPT; Olympus, Tokyo, Japan).
Cell proliferation/cytotoxicity assay
Cell proliferation was measured using the colorimetric 3-(4,5-dimethylthazol-2-yl)-2,5-diphenyltetrazolium bromide assay kit (Promega, Madison, WI). Briefly, human PDLSCs and PAFSCs (3.0×103 cells/well) were seeded in 96-well plates and cultured for 24 or 48 h. Cells were then exposed to 8% oxygen or chemicals, including DFO (Sigma Aldrich) or YC-1 (Sigma Aldrich) at various concentrations for the indicated durations. Fifteen microliters of premixed optimized dye solution was added 4 h before the treatment was stopped. Each condition was prepared in triplicate, and reactions were assessed using an ELISA reader at OD 595 nm (reference, 655 nm).
Western blot analysis
Human PDLSCs and PAFSCs (1.0×106 cells/dish) were seeded in 100-mm culture dishes and cultured for various times. Protein concentrations of whole cell lysates were determined using a DC Protein Assay Kit (Bio-Rad Laboratories, Hercules, CA). Equal amounts of protein (30 μg/lane) were resolved by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis followed by transfer to a polyvinylidene difluoride membrane (GE Healthcare, Chalfont St. Giles, Buckinghamshire, United Kingdom). Specific antibodies against CEMP1 were produced as previously described. 26 Primary antibodies against hemagglutinin (HA; 0.01 μg/mL) (Invitrogen, Carlsbad, CA), HIF-1α (0.01 μg/mL) (Epitomics, Burlingame, CA), and β-actin (Santa Cruz Biotechnology, Santa Cruz, CA) were used for blots that were finally developed using horseradish peroxidase-conjugated secondary antibodies (Santa Cruz Biotechnology) and visualized by enhanced chemiluminescence (enhanced chemiluminescence kit; GE Healthcare, Pittsburgh, PA).
Immunocytochemical staining
Cells were fixed in 4% paraformaldehyde in PBS at room temperature for 10 min. After rinsing with PBS, cells were blocked with 5% BSA and 0.2% Tween-20 in PBS for 30 min at room temperature. Cells were then treated with primary antibodies for Hypoxyprobe-1 (pimonidazole) (0.05 μg/mL), HIF-1α (0.05 μg/mL), or CEMP1 (0.5 μg/mL) for 16 h at 4°C. Normal mouse, rabbit, or goat IgGs (Santa Cruz Biotechnology) for each primary antibody were used as controls. Alexa Fluor® dyes-conjugated second antibodies (Invitrogen) were used for double staining. DAPI was used for counterstaining. Cell staining was evaluated using a fluorescence microscope (Olympus BX51; Olympus).
DNA constructs and transfection
A full-length open reading frame of HIF-1α (Accession No. NM_010431) was cloned into the pENTR D-TOPO vector (Invitrogen). All expression constructs were modified to include the nine amino-acid HA epitope YPYDVPDYA fused directly to the C-terminus. For expression in cell culture, a derivative of the Piggybac transposon system that facilitates high efficiency expression was employed. 27 PBX2.2 empty vector and PBX2.1-EGFP for GFP expression were kindly provided by T. Sanders (University of California, San Francisco). Transfection experiments were performed with Lipofectamine LTX and PLUS reagent (Invitrogen). A 2:1 molar ratio of Piggybac transposase helper plasmid 27 was combined with the transposon expression construct to mediate integration and high levels of expression.
RNA interference
At 24 h before transfection, cells were seeded onto a 24-well plate and grown to 60–70% confluence. Cells were transiently transfected with 20 nM HIF-1α siRNA (Santa Cruz Biotechnology) and control siRNA (Santa Cruz Biotechnology) for 4 h using Lipofectamine™ RNAiMAX reagent (Invitrogen) according to the manufacturer's instructions. After 48 h, transfected cells were harvested for RNA and whole cell lysate preparation.
Alkaline phosphatase assay and staining
Quantitative alkaline phosphatase (ALP) activity was determined by an assay based on the hydrolysis of p-nitrophenylphosphate (p-NPP) to p-nitrophenol (p-NP). Human PAFSCs and PDLSCs cultured in induction media for 21 days were harvested in lysis buffer solution containing 0.1% Triton X-100. Cell lysates (60 μL) from DSC samples were added to 60 μL 0.2 M diethanolamine and 30 mM p-NPP. Samples were then incubated at 37°C for 30 min, and the reaction was stopped by the addition of 60 μL 0.3 N sodium hydroxide. ALP activity was determined from the absorbance of p-NP at a wavelength of 410 nm using a microplate reader. For ALP staining, cells were fixed in 10% formalin, incubated with 0.1% Triton X-100 for 5 min, and then stained with the Leukocyte ALP kit (Sigma Aldrich) according to the manufacturer's protocol.
RNA preparation and real-time quantitative PCR analysis
To evaluate gene expression levels in human PAFSCs and PDLSCs, 5.0×105 cells were seeded in a 60 mm culture dish and cultured for 3 weeks under mineralization differentiation induction conditions. Total RNA was prepared using an RNeasy Mini kit (QIAGEN, Valencia, CA) according to the manufacturer's instructions, and cDNA was synthesized from 2 μg of total RNA using Superscript II reverse transcriptase (Invitrogen). Real-time PCR was performed with SYBR Green PCR Master Mix (Applied Biosystems, Warrington, Cheshire, United Kingdom) by following the manufacturer's protocols. Reaction conditions comprised 40 cycles of 15 s of denaturation at 95°C and 1 min of amplification at 60°C. All reactions were run in triplicate, and expression was normalized to that of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (Gapdh). Relative levels of transcript expression were quantified using the ΔΔCt method. 28 The calculation was performed by using the Ct value of Gapdh to normalize the Ct value of the target gene in each sample to obtain the ΔCt value, which was then used to compare different samples. Relative mRNA expression was compared in a histogram. Specific primers sets used in the analysis are listed in Table 1.
Ca IX, carbonic anhydrase IX; Gapdh, glyceraldehyde-3-phosphate dehydrogenase; Glut1, Glucose transporter 1.
In vivo experiments
Animals
All animal-use procedures and experimental protocols were conducted according to the guidelines of the institutional animal care and use committee (No. SNU −120427-2, ilar.snu.ac.kr, IACUC) at Seoul National University. ICR mice were obtained from Orientbio, Inc. (Sungnam, Korea) and maintained on a daily feed of rodent food in housing quarters with regulated temperature, cycled light (12 h on/off), and sterile water.
Detection of tissue hypoxia
To detect mouse tissue hypoxia, we employed the Hypoxyprobe™-1 kit that contains pimonidazole hydrochloride and detection antibodies (Hypoxyprobe, Inc., Burlington, MA). Pimonidazole was injected intraperitoneally on postnatal days 7 and 14 in ICR mice (60 mg/kg) 90 min before sacrifice (n=3). Tissue fixation was performed by in vivo perfusion to eliminate artificial staining. 29 All animals were anesthetized with a mixture of ketamine HCL (Parke-Davis, Morris Plains, NJ) plus 20% xylazine I.P. (Mobay Corporation, Shawnee, KS) and perfused with 0.1 M PBS (pH 7.4) for 30 s and then 4% PFA solution through the left ventricle for 5 min (5 mL/min) using a three-way stopcock that enabled the operator to quickly switch from rinsing to fixation solution, thus avoiding a drop in perfusion pressure. After tissue preparation, immunohistochemistry (IHC) was performed to detect tissue hypoxia using antibodies included in the Hypoxyprobe™-1 kit.
In vivo DFO treatment
To study the effects of DFO on cementum formation during tooth developmental stages in vivo, we divided ICR mice into two groups: a DFO group (n=6); 25 mg/kg/day of DFO was injected intraperitoneally everyday from postnatal days 7 to 13, and a control group (n=6); the same volume of saline was injected at the same time points as used for the DFO group. Mice from each group were sacrificed at postnatal day 14 for histological analysis.
Tissue preparation and IHC
Whole murine heads were divided into two pieces by cutting the cranial sagittal suture, then fixed in 4% PFA solution for 48 h at 4°C, and decalcified with 12% EDTA for 21 days. After paraffin embedding, sections were cut serially in the sagittal plane from the most ligual side. During sectioning, the mid-sagittal plane of the tooth germ (P7) was defined as the plane where the highest point of the crown and the most middle portion of the pulp chamber appeared at the same time. Semi-serial 5 μm sections of first and second molars were prepared and stained with hematoxylin and eosin (H&E) and IHC technologies. Primary antibodies for Hypoxyprobe-1 (pimonidazole) (0.05 μg/mL), HIF-1α (0.05 μg/mL), bone sialoprotein (BSP; Santa Cruz Biotechnology) (0.05 μg/mL), or CEMP1 (0.5 μg/mL) were applied to paraffin-embedded tissue sections. Deparaffinized sections were immersed in 0.6% H2O2/methanol for 20 min to quench endogenous peroxidase activity. Sections were then incubated in 5% BSA in PBS for 30 min and incubated overnight at 4°C with each antibody (1:100). Sections were incubated for 1 h at room temperature with the secondary antibody and reacted with avidin-biotin-peroxidase complexes (Vector Laboratories, Burlingame, CA) in PBS for 30 min. After color development with 0.05% 3,3′-diaminobenzidine tetrahydrochloride (Vector Laboratories), sections were counterstained with hematoxylin. For Hypoxyprobe-1 staining, Alexa Fluor dye-conjugated secondary antibodies (Invitrogen) were used for double staining, and DAPI was used for counterstaining.
Statistical analysis
Statistical analysis was performed using Statistical Package for Social Science 13.0 software (SPSS, Inc., Chicago, IL). Normal data with equal variance were analyzed using one-way analysis of variance with a Tukey procedure. Significance was defined as p≤0.05. Values in each graph represent mean±standard errors. All assays were performed at least thrice, and representative data were presented.
Results
Characterization of human DSCs and multipotential differentiation
International Society for Cellular Therapy (ISCT) has proposed a set of criteria for defining human MSCs, 30 and these minimum standards have been widely accepted. At passage 3, both human PAFSCs (Fig. 1A) and PDLSCs (Fig. 1B) were characterized by flow cytometry. The percentage of positive cells was determined by the relative intensity of antibody-binding cells. CD13, CD90, and CD146 were highly expressed, whereas CD34 was expressed at a low level in both stem cell types. ISCT has proposed that MSCs populations should be positive for CD90. In addition, these cells should exhibit low expression of hematopoietic antigens such as CD34 and markers for monocytes, macrophages, and B cells. 30 CD146 is a representative putative MSC marker. Our results indicated that the isolated cells were mainly mesenchymal and minimally contaminated with epithelial cells. To investigate the multipotent differentiation potential of human PAFSCs and PDLSCs, cells were supplemented with osteogenic medium to induce mineralization in vitro. After 3 weeks of induction, cultured DSCs formed an extensive amount of Alizarin res S-positive mineral deposits throughout adherent layers (Fig. 1C, left). Human PAFSCs and PDLSCs were also capable of undergoing adipogenic differentiation in response to treatment with adipogenic-inductive supplements for 14 days (Fig. 1C, middle). Moreover, human PAFSCs and PDLSCs formed Alcian Blue-positive nodules after 3 weeks of chondrogenic induction (Fig. 1C, right).
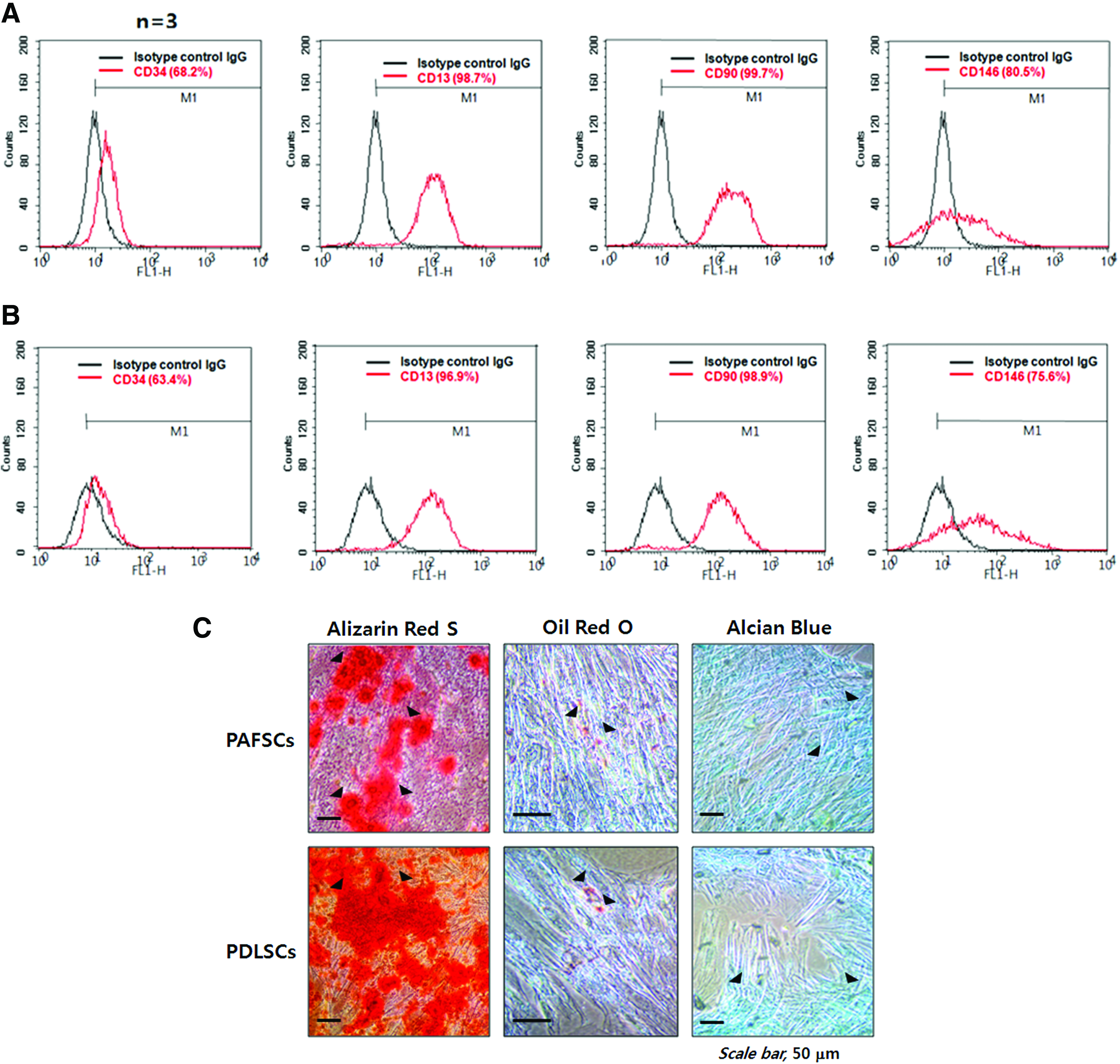
Characterization of human dental stem cells by flow cytometric analysis and multipotential differentiation. Flow cytometry assays were performed with human PAFSCs
Reduced oxygen tension stabilizes the HIF-1α protein and induces proliferation of human PDLSCs in vitro
Our preliminary data showed that anoxia and severe hypoxia (<2% oxygen) increased cell death substantially. Although 8% is higher than usual hypoxia when compared with other experiments, it is closer to the physiological oxygen tension experienced by embryonic stem cells, 10 somatic stem cells,10,17 and DSCs. 31 To confirm cellular hypoxia at 8% oxygen, we evaluated the accumulation of HIF-1α protein in vitro. Hypoxic response in these cells is characterized by an increase in pimonidazole adducts that form a stable covalent bond with the thiol groups of proteins 32 as shown by immunocytochemical analysis (Fig. 2A). HIF-1α protein expression in the nucleus of cells exposed to 8% oxygen for 24 h increased markedly. The hypoxic response in PDLSCs was similar to that in PAFSCs (data not shown). To examine the effect of the hypoxic state on cell viability, we performed cell proliferation assays. As shown in Figure 2B, PAFSCs and PDLSCs exposed to 8% oxygen proliferated to a greater extent (119.2%±19.6% and 132.8%±20.4%, respectively) than control cells.
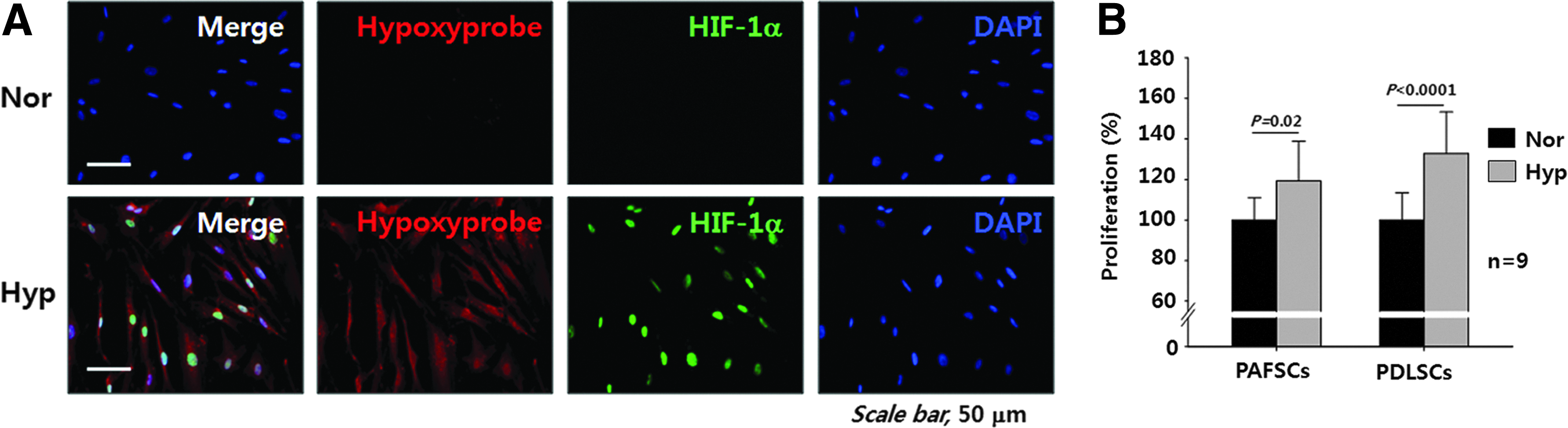
Reduced oxygen tension (8%) induced cellular hypoxic response and proliferation in vitro. Immunocytochemical staining was performed to test the cellular hypoxia. Hypoxyprobe (pimonodazole adduct)-positive cells were strongly positive for nuclear HIF-1α expression in PAFSCs exposed to 8% O2 (Hyp) for 48 h compared with normoxia (Nor) control
Hypoxia induces the expression of cementum-associated genes in vitro
To examine the hypothesis that cementoblast differentiation is regulated by oxygen tension, human PAFSCs and PDLSCs were cultured at 8% or 20% oxygen. To evaluate the activity of HIF-1α as a functional transcription factor, the expression of HIF-1α target genes was examined by real-time PCR in human PAFSCs (Fig. 3A) and PDLSCs (Fig. 3B) exposed to 8% oxygen. The expression of a HIF-1α target gene, carbonic anhydrase IX (Ca IX) 33 was up-regulated by 1.2-fold and 2.9-fold in human PAFSCs and PDLSCs treated with 8% oxygen for 21 days, respectively. To examine the effect of hypoxia on the expression of the cementum-associated genes, namely Cemp1 and Ocn, 34 we analyzed the expression level of those genes in human PAFSCs and PDLSCs after differentiation. Cemp1 expression was dramatically up-regulated in human PAFSCs and PDLSCs treated with 8% oxygen for 21 days by 11.6-fold and 4.8-fold, respectively. The expression of Ocn was also up-regulated in human PAFSCs and PDLSCs treated with 8% oxygen for 21 days by 5.6- and 5.2-fold, respectively. The expression of other cementum-associated genes, namely protein-tyrosine phosphatase-like member-a cementum attachment protein (Ptpla-cap) 35 and F-spondin, 36 was also examined in vitro; however, expression levels of these genes did not change significantly in response to oxygen treatment (data not shown).
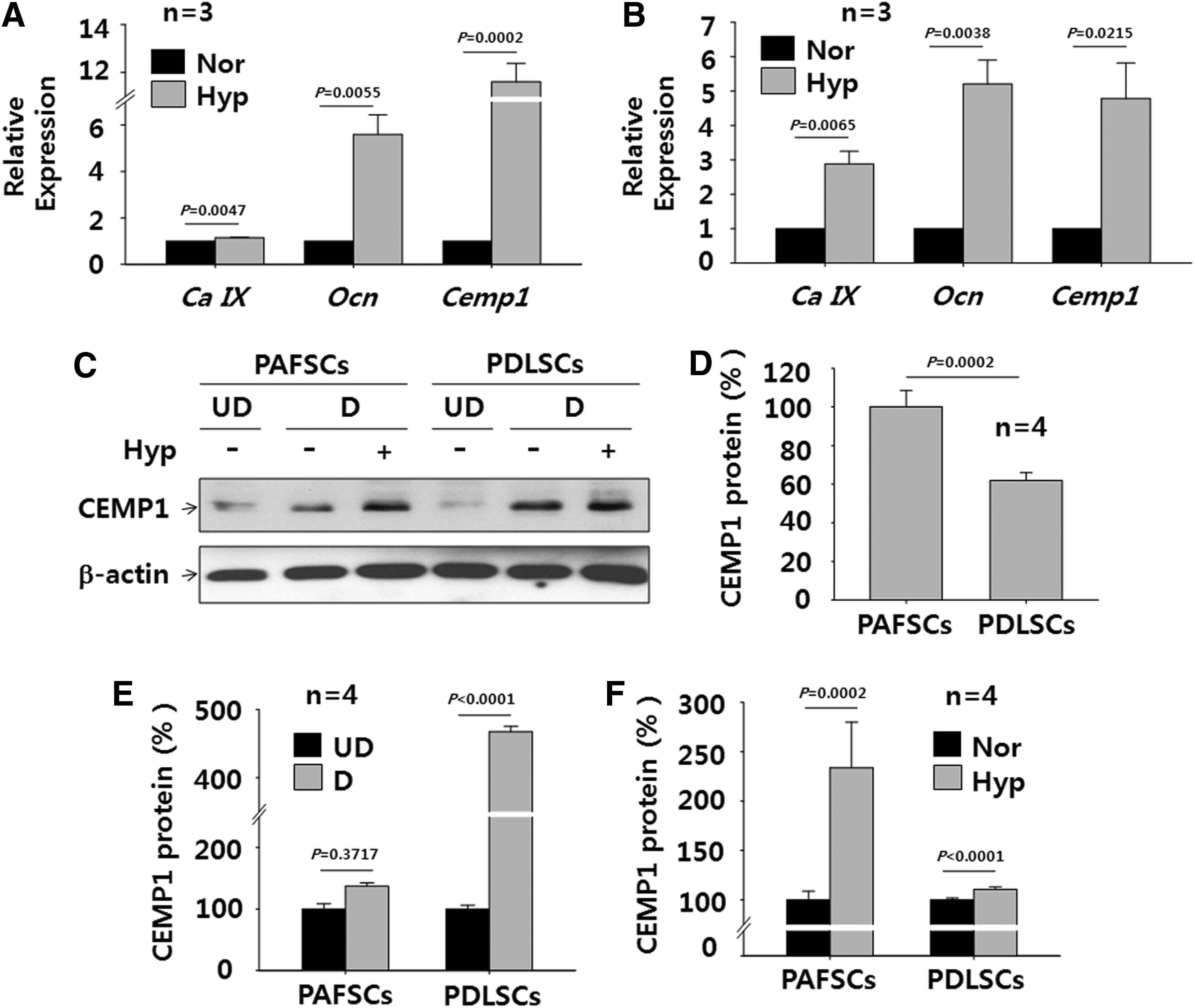
Hypoxia induces the expression of cementum-associated genes and CEMP1 protein in vitro. To examine the effect of hypoxia on differentiation into cementoblast-like cells, quantitative real-time PCR was performed after differentiation for 21 days in human PAFSCs
Hypoxia induces CEMP1 protein expression in vitro
Next, we evaluated the protein level of CEMP1 in human PAFSCs and PDLSCs differentiated in osteogenic medium for 21 days by western blot analysis (Fig. 3C). Intrinsic expression of CEMP1 protein was lower in undifferentiated PDLSCs (62.0%±3.9%) than in undifferentiated PAFSCs (Fig. 3C, D). However, CEMP1 protein expression in differentiated PDLSCs increased dramatically to 467.6%±8.0% relative to the undifferentiated control (Fig. 3C, E). CEMP1 protein expression in PAFSCs also increased in response to osteogenic differentiation to 137.2%±5.2%. When cells were treated with 8% oxygen, CEMP1 protein expression increased to 233.8%±46.2% in human PAFSCs and 110.5%±2.5% in PDLSCs (Fig. 3F). These data indicate that CEMP1 protein expression in both human PAFSCs and PDLSCs is significantly up-regulated when these cells are exposed to low oxygen tension during differentiation. Although CEMP1 protein induction in PAFSCs was more sensitive to hypoxia, CEMP1 induction was higher in PDLSCs than in PAFSCs, suggesting that the response of DSCs to hypoxia is cell-type specific.
Hypoxia enhances mineralization and ALP activity in vitro
During 21 days of osteogenic differentiation, both PAFSCs and PDLSCs exposed to 8% oxygen showed higher mineralization than control cells as shown by Alizarin red S staining (Fig. 4A). Mineralization in human PAFSCs and PDLSCs treated with 8% oxygen increased to 116.3%±11.0% and 164.2%±4.5%, respectively (Fig. 4B). Furthermore, ALP activity in human PAFSCs and PDLSCs treated with 8% oxygen increased to 117.4%±1.5% and 127.8%±4.3%, respectively (Fig. 4C). Both human PAFSCs and PDLSCs treated with 8% oxygen also showed increased ALP protein staining (Fig. 4D).
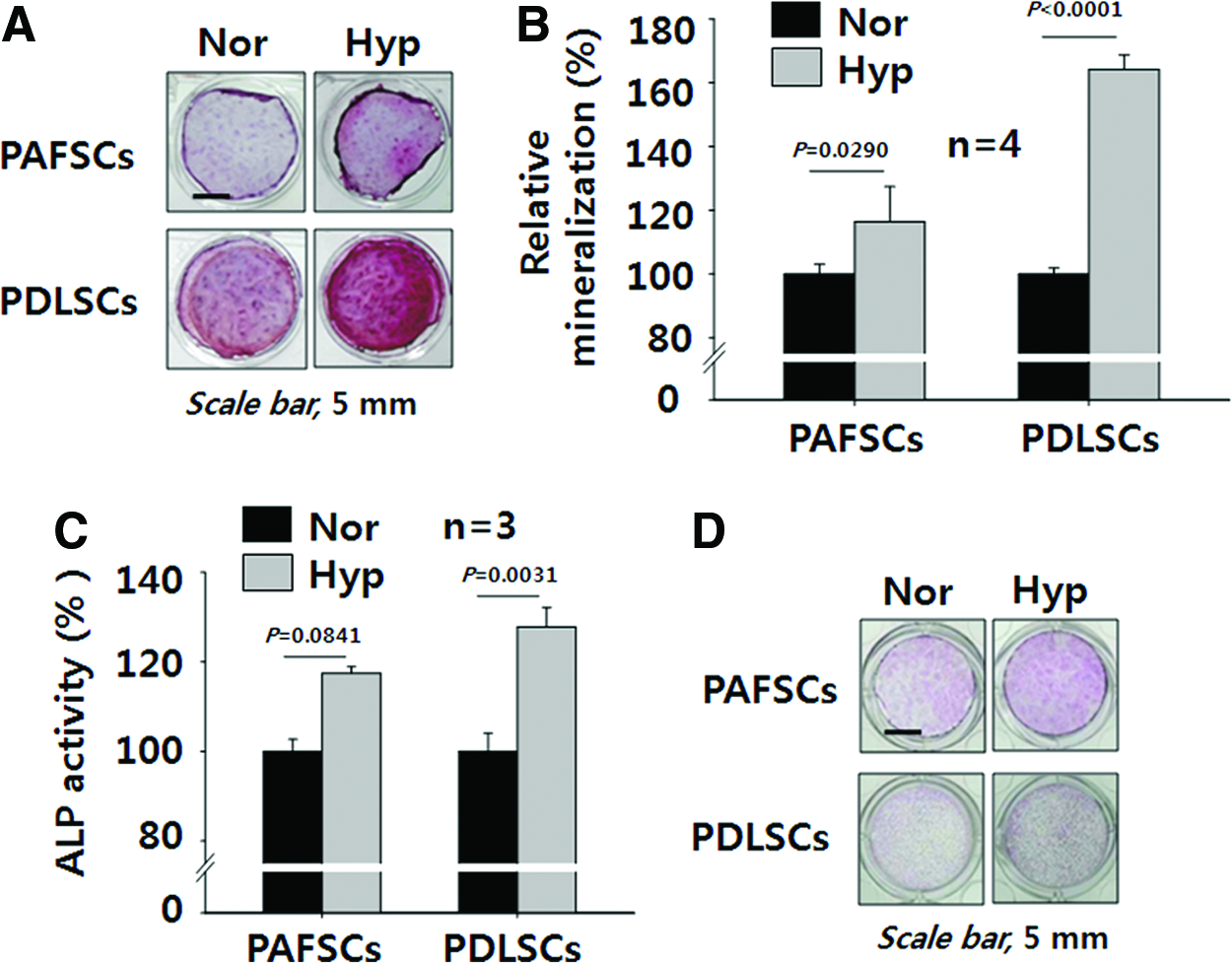
Hypoxia enhances mineralization and ALP activity in vitro. Mineralization levels of human PAFSCs and PDLSCs differentiated under hypoxia for 21 days were evaluated by Alizarin red S staining and compared with their normoxia control
Pharmacological activation of the HIF-1 pathway induces CEMP1 expression in vitro
A family of oxygen-sensitive prolyl hydroxylases (PHDs) hydroxylate HIFs under normoxia, which promotes their subsequent E-3 ligation and proteosomal destruction. 37 To differentiate between an increase in HIF-1α and other effects of a low oxygen environment, we blocked HIF-1α degradation with DFO, a known PHDs inhibitor, to activate the HIF-1 pathway at 20% oxygen. The optimum level of DFO was determined based on cytotoxicity analysis in both cell types (Supplementary Fig. S2). Minimum cytotoxicity in PDLSCs was observed at 1 μM DFO (94.6%±12.4% proliferation), but maximum mineralization was observed. DFO treatment of both PAFSCs and PDLSCs for 24 h at 20% oxygen increased HIF-1α and CEMP1 protein expression to levels similar to those observed for hypoxia (Fig. 5A). In addition, Alizarin red S staining showed that mineralization was also induced in PDLSCs treated with DFO at 20% oxygen (Fig. 5B) as well as in PAFSCs (data not shown).
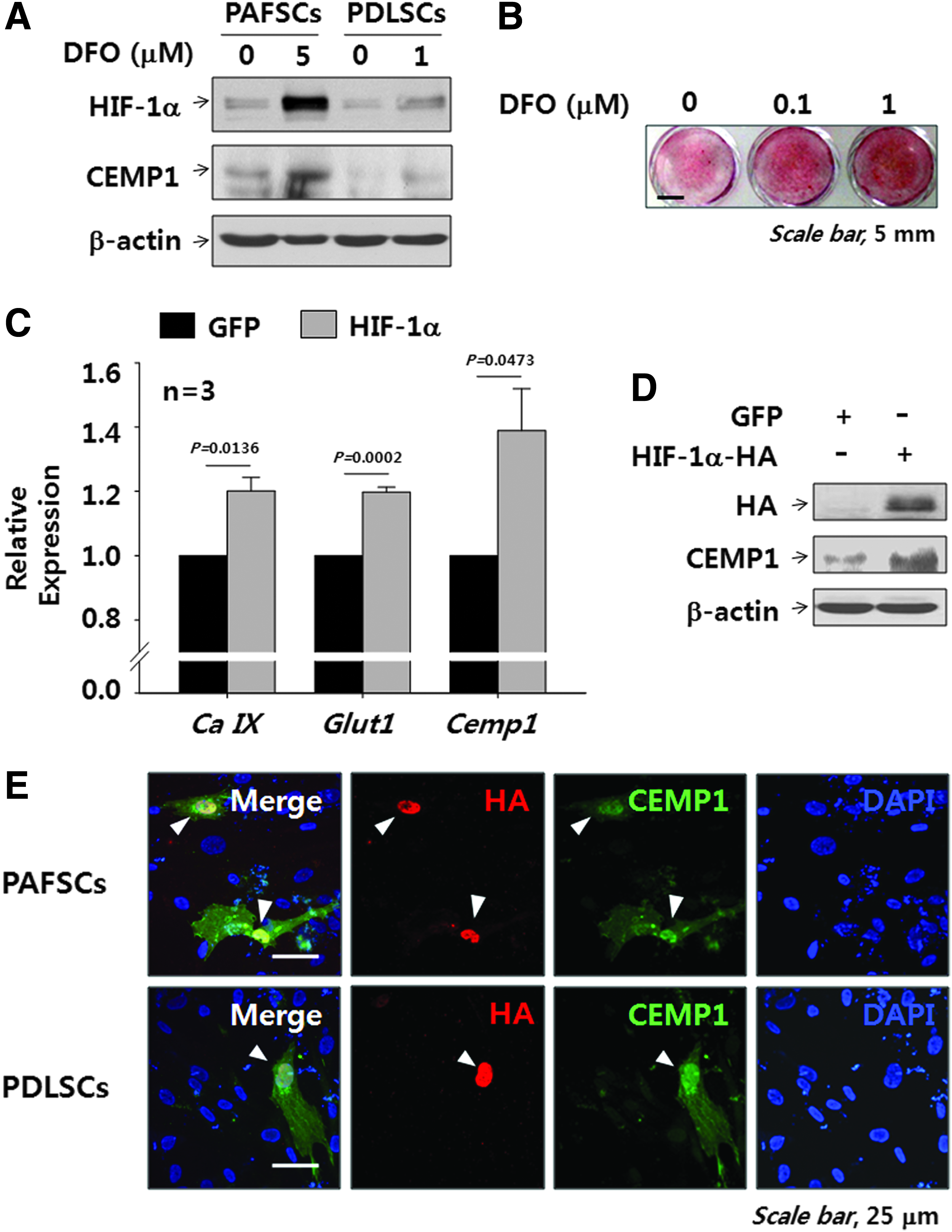
Phamacological and genetic activation of the HIF-1α increases CEMP1 expression and final mineralization in vitro. Human PAFSCs and PDLSCs were treated with DFO for 24 h, and Western blot was performed to analyze the level of HIF-1α and CEMP1 protein by using cell lysate
Enforced expression of HIF-1α enhances CEMP1 expression in vitro
Next, we introduced HIF-1α into the cells to investigate the effect of HIF-1 on CEMP1 protein expression and then performed real-time PCR and western blot analysis. As expected, HIF-1α up-regulated the expression of Cemp1 (Fig. 5C) as well as Ca IX and Glucose transporter 1 (Glut1), which are known HIFs-target genes. In addition, CEMP1 protein expression in PDLSCs transfected with a plasmid harboring HIF-1α fused with an HA tag (Fig. 5D) was confirmed by western blot analysis. Moreover, immunocytochemical images clearly showed correlated expression of HIF-1α and CEMP1 in PAFSCs and PDLSCs transfected with Hif-1α (Fig. 5E).
Inhibition of HIF-1 activity reduces CEMP1 protein expression and further mineralization in vitro
To determine whether HIF-1 is responsible for induction of the CEMP1 protein and mineralization, we suppressed HIF-1α expression by siRNA technology. The marked decrease in HIF-1α protein expression at 8% O2 indicates effective suppression of HIF-1 activity. CEMP1 protein expression was significantly lower at 8% oxygen in cells transfected with HIF-1α siRNA than in cells transfected with control siRNA (Fig. 6A). In addition, we treated cells with YC-1, a potent HIF-1 inhibitor, to investigate whether HIF-1 inhibition reduced mineralization. The optimum level of YC-1 was determined by cytotoxicity analysis in both cell types (Supplementary Fig. S3). One micromolar YC-1 was not cytotoxic to either PAFSCs or PDLSCs; the proliferation rates of these two cell types were 131.8%±16.4% and 110.9%±11.0%, respectively. As expected, mineralization of PDLSCs treated with YC-1 at 8% oxygen for 21 days decreased significantly (Fig. 6B); the same effect was observed in PAFSCs (data not shown).
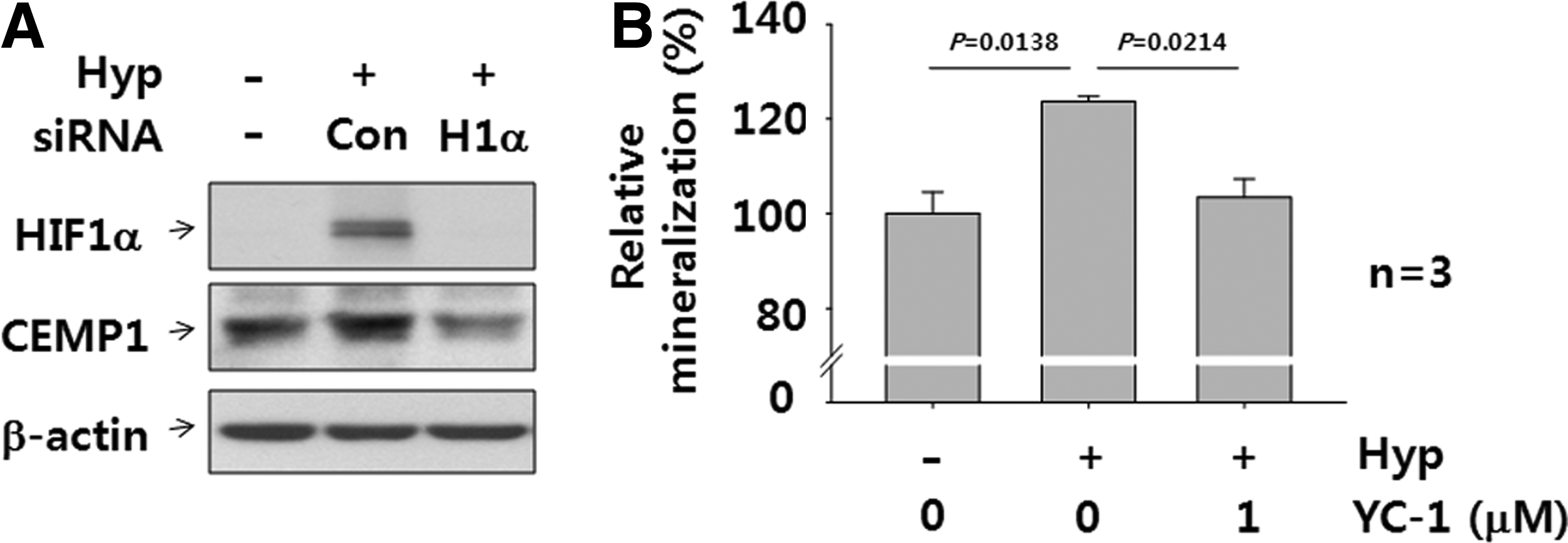
RNA interference and phamacological inhibition of the HIF-1α decreases CEMP1 expression and mineralization in vitro. Human PDLSCs were transfected with HIF-1α (H1α) and control (Con) siRNA for 24 h while exposed to hypoxia. Western blot was performed to analyze HIF-1α and CEMP1 protein by using cell lysate and compared with control siRNA group (Con)
Mouse tooth roots at postnatal day 7 develop in a hypoxic environment in the presence of CEMP1
To assess the role of tissue hypoxia in mouse tooth development, we employed the hypoxia marker Hypoxyprobe-1, (pimonidazole hydrochloride). 38 Hypoxyprobe-1 is converted by hypoxia-activated nitroreductases into a reactive intermediate that forms covalent pimonidazole adducts with cellular components in hypoxic regions. 29 To detect tissue hypoxia in developing mouse tooth germ, mid-sagittal sections of the mandibular second molars were analyzed by IHC. As seen in Figure 7C, Hypoxyprobe-1 was highly concentrated at the proliferative root apex of the tooth germ at postnatal day 7, suggesting that highly proliferative cells may be localized to this region and contribute to local low oxygen tension. Pimonidazole adducts were highly concentrated at the apical area of the root edge and the central apex of the outer face of molar roots (Fig. 7F) when compared with the vehicle-injected control (Fig. 7B, E). Of note, the cytoplasm of odontoblasts, which are typically arranged similar to a palisade along the pulp, was highly positive for Hypoxyprobe-1 at the apical area. However, the dental follicle (DF) surrounding the tooth germ appeared to remain non-hypoxic at this stage, while predentin exhibited weak nonspecific staining (asterisks in Fig. 7B, C). CEMP1 expression was highly correlated with tissue hypoxia as assessed by Hypoxyprobe-1 staining (Fig. 7I) and HIF-1α measurement (Fig. 7H), suggesting that hypoxia may influence overall root development of mouse molars as well as CEMP1 expression to regulate cementum formation in vivo. The CEMP1 protein of mouse tooth germ at postnatal day 7 was immunolocalized in the DF and dental papilla at the apical area of root edge and the central apex in the outer face of molar roots and some of the pulp cells (Fig. 7F), while that of human adult tissue localized primarily to a restricted area of the PDL space (Supplementary Fig. S1).
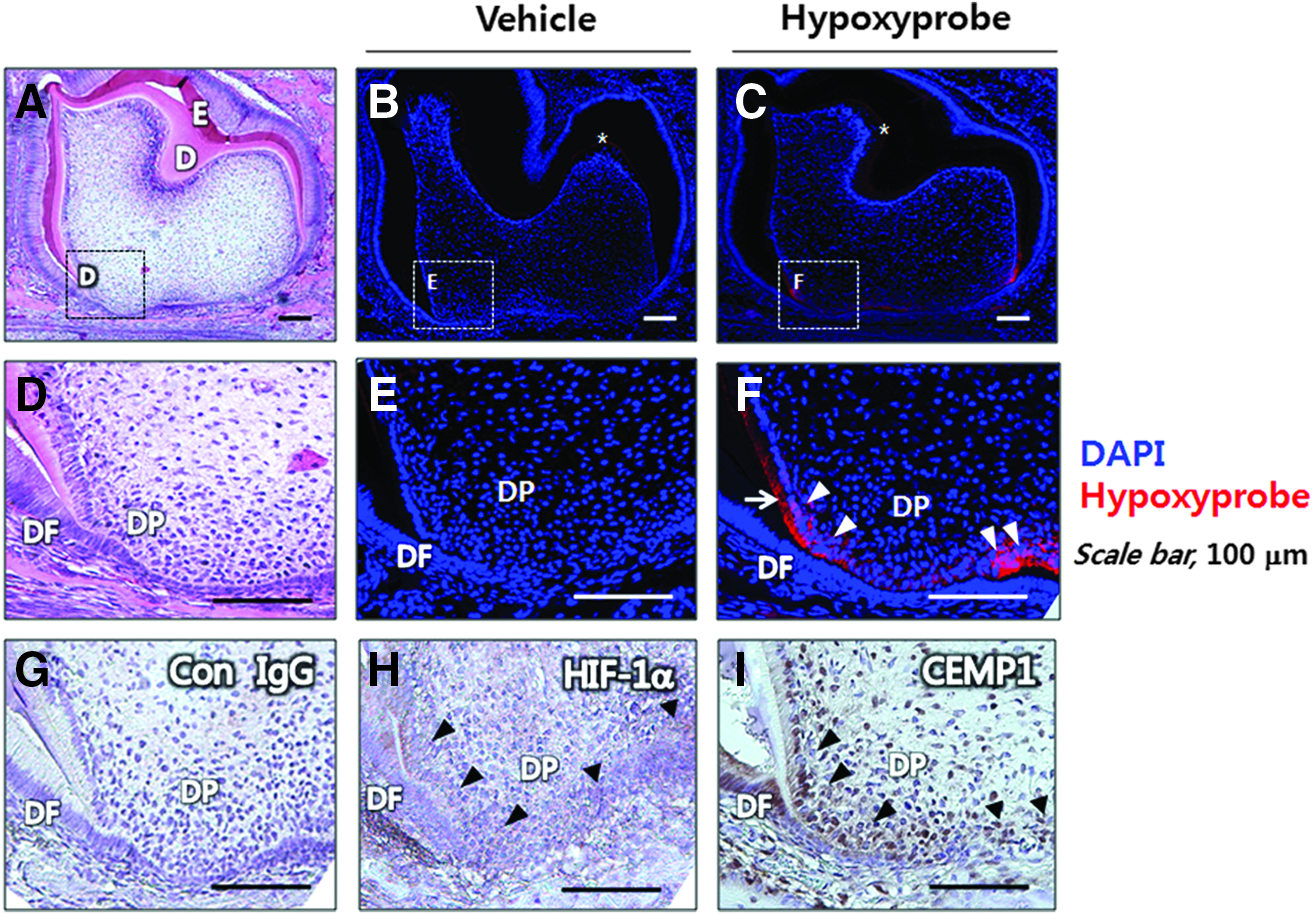
Mouse tooth roots develop in hypoxic environment with CEMP1 expression at postnatal day 7 in vivo. An H&E staining section shows a whole tooth germ of the second lower molar at postnatal day 7
Mouse PDL space is hypoxic after tooth root formation in vivo
To investigate tissue hypoxia in late tooth root development, we injected Hypoxyprobe-1 into mice at postnatal day 14 and fixed the tissue using an in vivo perfusion method to eliminate artificial staining. Days were selected to capture developmental time points of interest during tooth-bone interface development, including cementogenesis. 21 At postnatal day 14, root morphogenesis was almost finished and the root surface was covered with thin acellular cementum to facilitate attachment of the PDL. 39 At this stage, tissue hypoxia was localized mainly to the apical part of the PDL space and the furcation of molar roots (Figs. 8B, E, H) when compared with the vehicle-injected control (Figs. 8A, D, G). IHC revealed that most CEMP1-positive cells were localized to the PDL space at the furcation (Fig. 8F) and the apex (Fig. 8I) of molar roots.
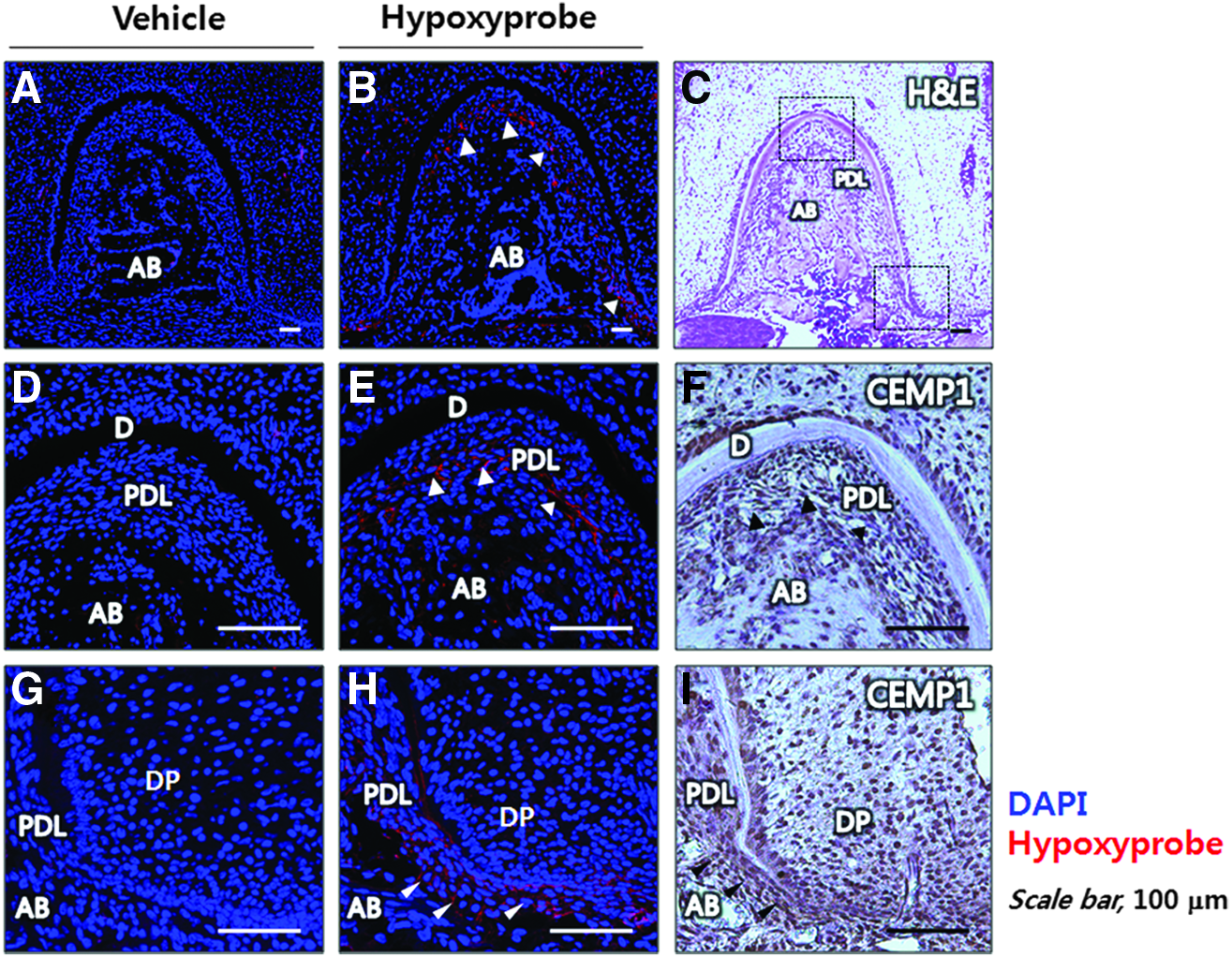
Mouse PDL spaces are hypoxic at the formation of the tooth-bone interface in vivo. To assess mouse tissue hypoxia of developing mouse tooth root, Hypoxyprobe-1was injected intraperitoneally into ICR mouse at postnatal day 14 (n=3) and analyzed by immunohistochemical staining
Stimulation of the HIF pathway enhances acellular cementogenesis in vivo
To examine whether a similar pathway is involved in acellular cementum formation, we analyzed the cervical area of molar roots at postnatal 14 days. Using a refined IHC technique, which allows more sensitive identification of differential pimonidazole adducts, we discovered that tissue hypoxia localized differentially to the line covering the cervical cementum area (Figs. 9A–C, note the arrowhead in 9B) when compared with the vehicle-injected control (data not shown). Interestingly, the pimonidazole adduct-positive line colocalized with BSP (Fig. 9K), which is a mineral-binding member of the SIBLING family with a role in the mineralization process 40 ; there is a close relationship between AEFC formation and the act of mineralization. To activate the HIF-1 pathway during tooth root development, we injected DFO daily into mice from postnatal day 7 to postnatal day 13. No significant differences in the general state of animals, including body weight or body temperature, were detected among groups during DFO injection. After 7 days of treating mice with DFO, there was a marked increase in pimonidazole adduct accumulation (Fig. 9E–G, note the arrowhead in 9F) and HIF-1α protein expression (Fig. 9M) in PDL cells lining the cervical acellular cementum of mouse molar roots. CEMP1 was widely expressed during molar root formation and was expressed more strongly in the PDL space in response to DFO treatment than in the control (Fig. 9L). Despite increased expression of CEMP1 and HIF-1α in the PDL space, overall cementum thickness did not change significantly as shown by immunolocalization of BSP (Fig. 9N). These results indicate that activation of the HIF-1 pathway induces CEMP1 protein expression and may affect cementogenesis during tooth root development in vivo.
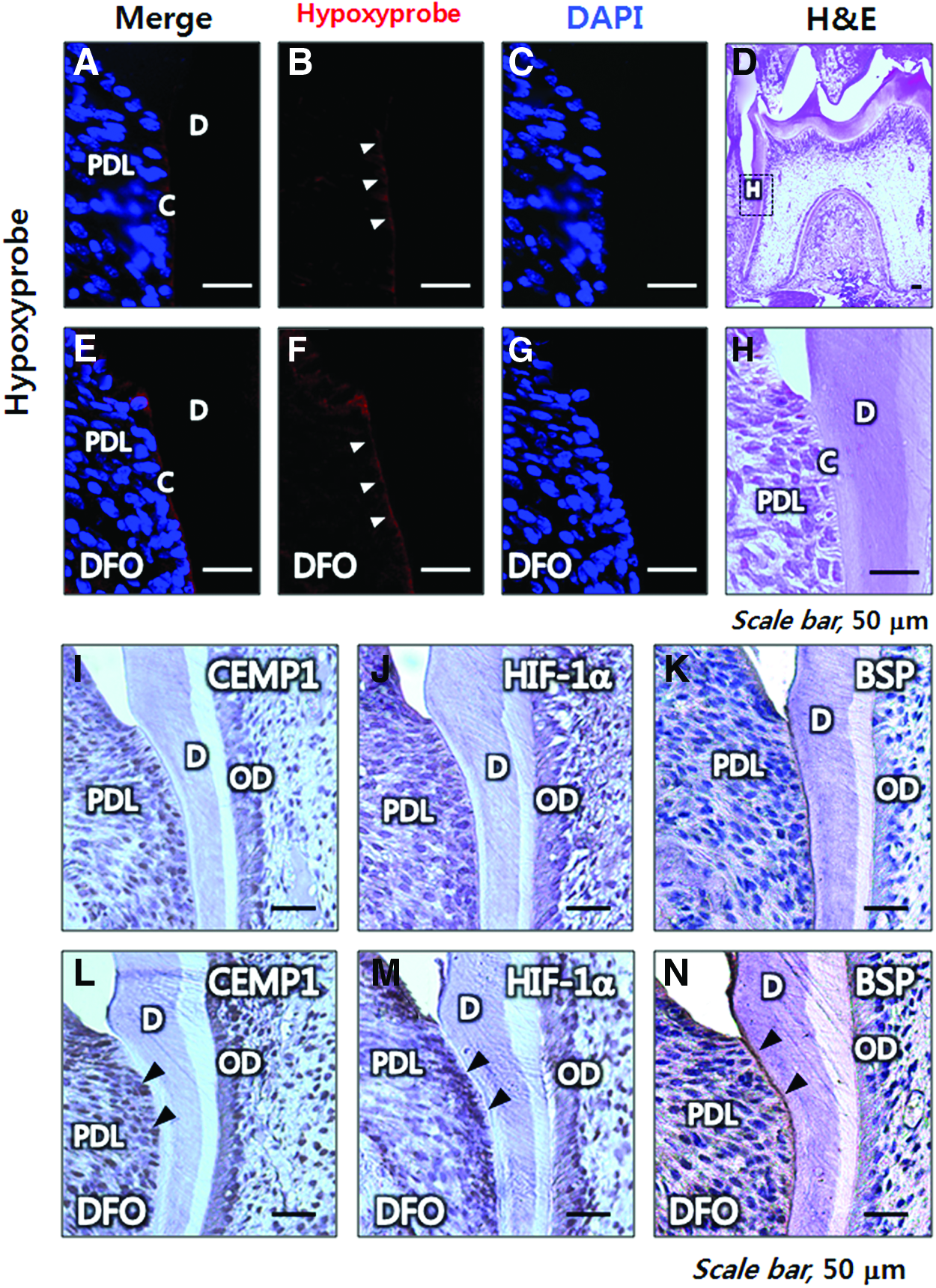
Acellular cementogenesis occurs in a hypoxic environment, and stimulation of HIF pathway enhances acellular cementogenesis in vivo. Hypoxyprobe-1was detected by immunohistochemical staining and analyzed to assess tissue hypoxia in cervical acellular cementum of mouse molar roots at the postnatal day 14. Hypoxyprobe-1 was also detected after DFO treatment to differentiate HIF-1 pathway in acellular cementum formation in vivo
Discussion
CEMP1 was recently identified and characterized as a protein participating in biomineralization of progenitor cells in the PDL. 9 The precise mechanisms which couple CEMP1 expression to cementum formation are still unknown, but it appears that CEMP1-positive progenitor cells localized in the PDL participate in mineralization and differentiation of progenitor cells for cementum formation.8,41 Indeed, previous studies showed that CEMP1 protein expression localized to the cementoblast cell layer lining the cementoid surface of cementum in both acellular and cellular cementum of human adult periodontal tissues.6,42 Our immunohistochemical staining for CEMP1 in human tissues confirm these previous findings (Supplementary Fig. S1).
There are abundant reports that hypoxia induces proliferation of human MSCs.15,19,43 It has also been reported that low oxygen tension during tissue culture has positive effects on mineralization and vascularization of bone.15–17,19,44 However, the effects of hypoxia on osteogenesis and mineralization are still controversial.15,45–48 Previous in vitro hypoxia experiments have been performed at lower oxygen tension (<2%) than the average physiological oxygen tension (4–8%). 49 Under low oxygen tension, bone marrow-derived MSCs show a decrease in osteogenic differentiation or spontaneous calcification.46–48 Our preliminary oxygen gradient data indicate that neither PAFSCs nor PDLSCs were able to proliferate or undergo osteogenesis under 1% oxygen tension in vitro (data not shown). To mimic physiological oxygen conditions more closely, we subjected cells to 8% oxygen for long-term differentiation. Interestingly, hypoxia induced significant cementogenesis of human PAFSCs and PDLSCs at this oxygen tension. However, the responses of PAFSCs and PDLSCs to hypoxia in terms of CEMP1 expression levels were different (Fig. 3F). Staining of human tissues for CEMP1 (Supplementary Fig. S1) indicated that PDLSCs could be a good source of progenitors for cementogenesis or regulation of cementogenesis, because PDLSCs expressed higher levels of CEMP1 protein during osteogenic differentiation than PAFSCs. Cells usually respond to external changes through crosstalk between cell surface proteins and cytoskeletal proteins. Therefore, cell surface molecules are likely to be involved in the differences in CEMP1 expression in the two cell types in response to hypoxia. The mechanisms underlying the difference in the expression of CEMP1 between PAFSCs and PDLSCs should be explored in future studies.
Our in vivo results indicate that oxygen tension was reduced by local proliferation of cells at the apical part of the tooth root during development (Fig. 7). At postnatal day 7, CEMP1 protein expression in the mouse tooth germ was highly correlated with tissue hypoxia. Interestingly, CEMP1 showed strong immunolocalization to the DF and dental papilla at the apical area of the root edge, the central apex in the outer face of molar roots, and some of the pulp cells (Fig. 7I), while human CEMP1 protein was localized to cementoblast cells and restricted PDL cell subpopulations (cementoblast precursors) in the PDL space (Supplementary Fig. S1). In addition, we have found that PDLSCs and other DSCs, including PAFSCs and dental pulp stem cells, express CEMP1 protein without osteogenic differentiation (unpublished data). These data support the possibility that CEMP1 has other roles in other types of cells during tooth development.
HIFs activate genes that regulate cellular events, and hypoxia and HIFs are responsible for several aspects of developmental morphogenesis. To obtain mechanical insights into cementogenesis in response to hypoxia, we demonstrated that HIF-1 activation by gene introduction or pharmacological activation induced CEMP1 expression in human PAFSCs and PDLSCs. Furthermore, we found that HIF-2α induced CEMP1 expression significantly. However, it is unclear whether the two HIF-α proteins have redundant roles in cementogenesis. In sum, our in vitro experiments employing DSCs provided a mechanistic platform for probing the proposed roles of hypoxia in cementoblast mineralization and gene expression according to HIF-1 expression.
A few studies have investigated the role of hypoxia in tooth development, including root formation and cementogenesis. Based on our in vitro findings, HIF-1 stimulates cementum-associated gene expression and mineralization. We examined whether hypoxia affects tooth root and periodontum development and whether an HIF-1-modulating drug enhanced CEMP1 expression and cementoblastic differentiation in vivo. At postnatal day 7, mouse molar tooth germs develop (late bell stage). Tissue staining indicated that there was proliferation of stem cells at the apical region, resulting in local hypoxic environment. At postnatal day 14, the number of hypoxic regions in the PDL space increased, suggesting that a rigid tooth-anchoring structure formed in the periodontal tissue between the tooth root and alveolar bone during this stage of mouse tooth development. Given that CEMP1-expressing cells could be a source of progenitors for cementoblasts or regulators of cementogenesis, the fact that CEMP1 expression and pimonidazole adducts had a proximal distribution indicates that the hypoxic PDL space supports cementogenesis of progenitor cells by increased CEMP1 expression at the apical part and furcation of molar roots, corroborating our in vitro results. DFO is a metal-chelating drug that is often used in aluminum-overloaded patients who require dialysis. DFO can promote bone mineralization and regeneration without decreasing aluminum levels in the bone and have indirect effects on bone cells via HIF-1α stabilization.50,51 In this study, DFO induced a marked increase in CEMP1 and HIF-1α protein expression in PDL cells in vivo. These findings strongly indicate that DFO may likewise regulate cementoblast differentiation of progenitor cells through the same pathway as osteoblast differentiation.
In summary, we demonstrated that hypoxia is accompanied by an increase in HIF-1α protein expression, which, in turn, increases CEMP1 protein expression and mineralization of human DSCs such as PAFSCs and PDLSCs, promoting cementogenesis in vitro. We also provided evidence that HIF-1 activity is required to stimulate CEMP1 production and to allow mineralization and an ALP enzymatic response in vitro. Furthermore, we found that mouse tooth root development occurs in a hypoxic state and that stimulation of the HIF-1 pathway enhances CEMP1 protein expression in mouse PDL spaces in vivo. Based on these findings, we suggest that the combination of hypoxia and human DSCs may be useful in regenerative therapies to repair periodontal defects.
Footnotes
Acknowledgment
This work was supported by a grant from the National Research Foundation of Korea (NRF) funded by the Korea government (MSIP) (No. 2011-0028922).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.