
Abstract
So what is the “big deal” about engineering of collagen materials? It is certainly not enough to produce the familiar line about it being “more physiological.” This phrase explains very little of the substance and gets nowhere near to the base of the question. We need to be clear about why this is important enough to justify a major new research investment.
Direct and Indirect Tissue Fabrication
The strategy of cell-dependent growth in 3D is logically constrained to focus on producing ever better, more controllable cells, as these are the essential workhorses. However, by the same analysis, the approach of direct (cell-independent) fabrication is forced to tackle the problem of directly fabricating native tissue materials, that is, the extracellular matrix. The most logical and practical base component for such a matrix material is almost certainly the fibrillar type I collagen, not least as the most abundant, mechanically important support protein in mammals.
Examples of direct tissue-engineering technologies include (a) matrix-rich and, potentially, (b) cell-rich layer engineering, (c) 3D cell or bio-printing and some forms of (d) electrospinning. The first two, the matrix and cell layer engineering, involve fabrication of thin layers (tens of microns thick) of either a dense native collagen mesh seeded with cells 2 (a) or a cohesive, self supporting cell sheet (b). Cell layer fabrication involves extended culture of sheet-forming cells (such as epithelial or muscle cells) on special thermoresponsive plastic surfaces.3,4 Such cell sheets are then stacked to produce a tissue, with a minimal matrix. In matrix-rich layer engineering (a), each sheet is made as an over-hydrated collagen cell gel layer, and then rapidly compressed (∼100-fold) to the desired density and layered into heterogeneous stacks or spirals.1,5
Layer-engineering works by assembling very thin sheets with defined structures in their X-Y planes. However, as their thickness (Z-plane) is negligible, the effective Z- plane structure of the final material is produced by a separate, layer assembly stage (e.g., 20×50-μm collagen layers give a 1-mm tissue). This separate Z-plane assembly stage is a key difference with comparable bio-printing (c) or electrospinning (d) techniques, where the aim is to assemble cell and matrix elements, without damage, all in one X-Y-Z-plane assembly process. This difference is the key as the potential advantages of printing and spraying must be balanced by the technical difficulties (yet to be solved) of achieving such a high target in one stage, without cell protein damage. This means that, for now, layer engineering is the single practical approach routinely in use for direct collagen tissue engineering.
Our Engineering Approach Affects Our Choice of Support Material
An interesting consequence of indirect engineering, by growing cells, has been to make it reasonable to employ temporary, nonbiological, scaffold materials. After all, given that the cultivation process itself is likely to involve periods of weeks or longer, it is reasonable to have an extra stage where the support material dissolves away, over similar periods to leave a cultivated cell matrix. So, while synthetic polymer scaffolds are an extra stage, with their own problems, they are an acceptable extra. However, this rationale changes completely for direct fabrication strategies. The great advantage of direct tissue fabrication is its rapidity, taking minutes or hours rather than weeks for bioreactor culture.2,5 Consequently, users of direct, rapid fabrication face a stark problem with synthetic polymer (i.e., nonphysiological) support materials. As a result of their long cell-seeding stages and slow scaffold disappearance, most of the advantages of direct engineering are lost (i.e., process rapidity and/or matrix bio-mimesis). So, for users of direct tissue fabrication, non-natural support materials are an intermediate, which compromises their process.
This is the analysis, which most clearly shows why engineering of natural polymers, especially collagen, is becoming the new big deal.
What Sort of Collagen Is Needed?
This, in part, explains the developing view that when we genuinely aim to fabricate tissue equivalents, then we must also take over, from our cells, the task of making a native, if simple matrix material. Even though a native material can also be highly simplified, the idea of engineering a consistent, safe, mimetic biomaterial out of collagen polymer is a high hurdle. However, if as much research resource had been invested engineering collagen polymer materials as into synthetic biodegradable polymers over the last 20 yrs, we might now be close.
The perceived position can appear more complicated as there already appears to be a plethora of collagen support materials and implants. The many forms of collagen sponge and sheet as clinical hemostats or cell supports6–8 can suggest this is a nonproblem. Many are formed from slurries of shredded collagen or whole collagen tissue fragments, dried, and crosslinked into dense materials with or without cell seeding. A second strategy uses decellularized sheets of intact animal collagen 9 (e.g., from dermis or fascia), sometimes with chemical crosslinking. Are these not, then, “engineered collagen materials”?
Critical analysis of the technologies used suggests that such top-down processes (i.e., refining down complex tissues into less complex usable parts) make materials, which are better termed as “materials made of collagen.” In contrast, the engineer in the engineered collagen concept refers to the building up of complexity using the simplest possible building blocks. This is bottom-up fabrication. These are as different as bottom-up synthetic production of diesel fuels from plant sugars, versus top-down refining of Saudi crude to give petroleum.
Current collagen material processing mainly relies on simple treatments to refine out infective and immunogenic components. Restructuring of the refined fragments is normally to promote cell ingrowth and mechanical strength, often through focus on the pore size, interconnectivity, and wall properties. However, equivalent engineered collagen approaches would fabricate the material UP from its basic building blocks, such as amino acid sequences (as in recombinant collagens) 10 or soluble collagen monomers, purified from tissues,2,11 much as we might engineer sportswear from cotton and Lycra. In fact, it becomes clear that having collagen as the main component is just the first entry requirement for collagen materials.
Engineering Up Toward Complexity
The advantages of bottom-up engineering of collagen materials, as in our example of sports materials, are the ability to control the spatial and mechanical performance. The most immediate tissue engineering advantage of collagen materials is the entrapping of cells within their fibril structure, as it is built up (i.e., during fibril formation). 2 This not only abolishes traditionally slow, problematic cell-seeding stages, but positions those cells within, rather than on the material surface, their natural location.
Analysis of current tissue-engineering and regenerative medicine strategies highlights this simple requirement (interstitial cell distribution) as key to progress. 1 However, this high target brings constraints, including:
(i) The collagen monomer building-block stock must undergo rapid, effective fibrillogenesis, that is, it should rapidly form strong gels.
(ii) Incorporation of cells during gelling, at tzero as fibrils aggregate, means that all following processes must be cell friendly (physiological) and rapid.
(iii) The hierarchical nature of collagen aggregation and collagen-tissue structure requires that controls are focused at specific-length scales, ranging from the atomic to the gross.12–14
While (i) and (ii) are self-evident, the imperative for processes to operate at specified points in the hierarchy is less familiar, although the textile analogy is again useful. The Lycra polymer chemist may work at the atomic and monomer levels to produce polymer chains with distinct properties, but different specialists are needed to spin/blend and weave the fibers into fabrics and still others to sew fabric shapes into the gross garment. Similarly, we aim to aggregate collagen monomers (1.5 nm) into fibrils (50–250 nm) to trap and control individual cells (10's μm) and cell clusters/niches (100's μm), which will together mimic/replace the shape and mechanics of whole tissues (Fig. 1). While such hierarchies are functionally interconnected, their fabrication is only weakly interdependent (just as the polymer scientist cannot control the fit of a leotard).
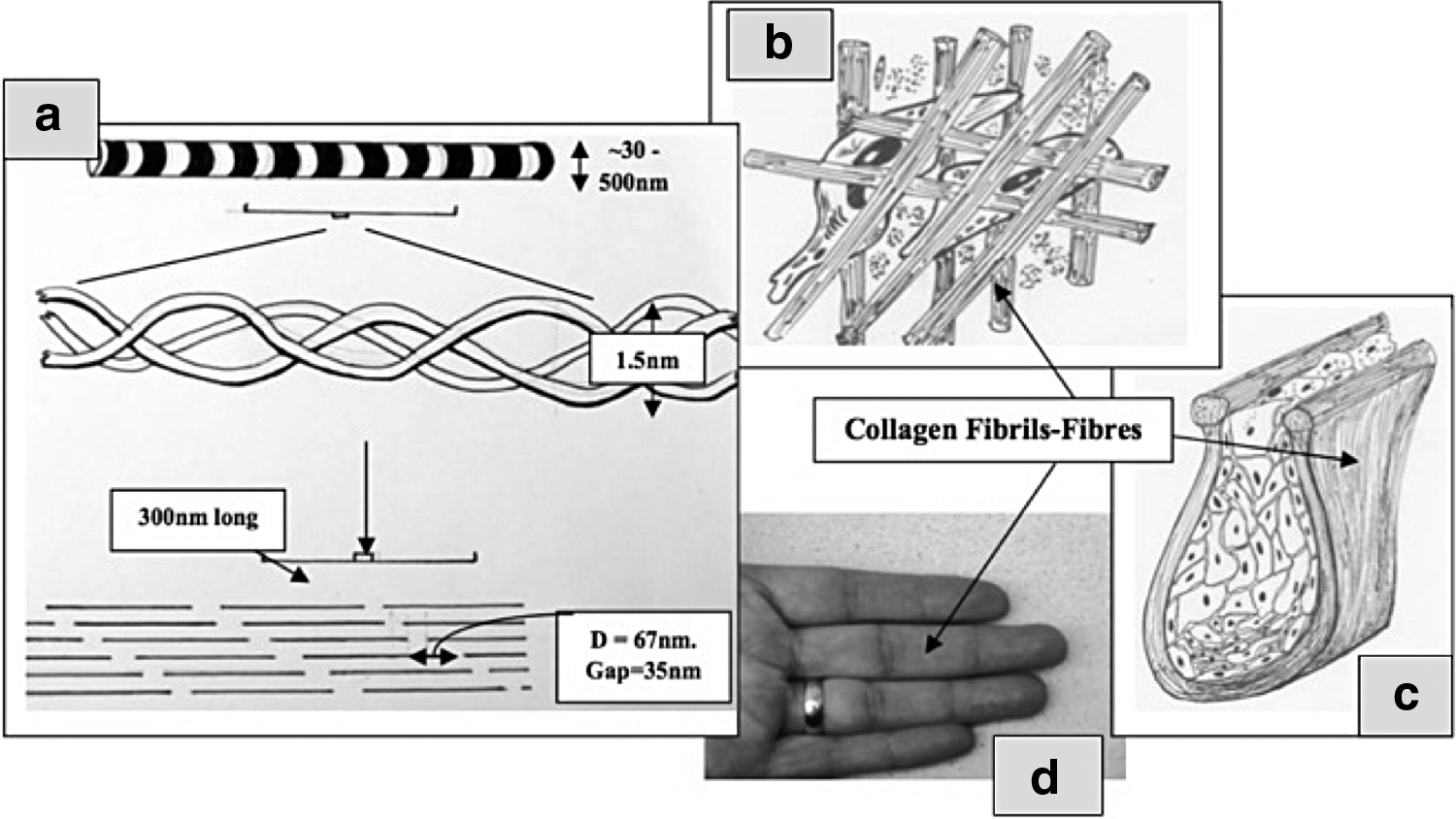
Illustration of four (nominal) hierarchies of collagen material organization. At first, molecular level
The size scales illustrated here (e.g., molecular, fibrillar, cellular, and tissue) are useful, if arbitrary, to illustrate the different expertise and problem sets involved. These are linked to the logic of bottom-up building of complexity. For example:
(a) Molecular-level collagen engineering (<2 nm) might aim to remove the variability of aggregation and minimize risks of infection, rejection, and inflammation. 15
(b) Fibril-level engineering aims to tailor the fibril network physical–mechanical properties by controlling fibril packing, alignment, and diameter/interconnection.12,14,16
(c) Cell-scale engineering aims to produce interconnections and 3D architectures to produce particular cues and microenvironments around cell(s).
(d) Tissue-scale engineering insert 3D structure aiming to control behavior/differentiation of cell clusters or multilayers to mimic native tissue structures, such as neurovascular ingrowth, stromal–epithelial layers, glands, ∼0.2–20 mm.
Collagen Engineering and Hierarchy Positioning
Where we start, or what part of the material hierarchy we focus on arguably should be a pragmatic decision based on the greatest, most pressing need at any one time to achieve the goal. Do we need a safer starting material (i) or stronger, more mimetic materials (ii), do stem cell therapies need better 3D cell niches (iii) or improved host tissue ingrowth (iv)?
Best practice requires a critical analysis before we go too far with our strategies; for example, which level of collagen engineering gives the greatest repair/regeneration benefit? This should first establish the pragmatic bottom building block for our bottom-up processing. For example, to engineer collagen-based cell niches (>100 μm scale), it would be illogical to start by developing new collagen-like sequences from single amino acids. The problem here is to assemble collagen fiber bundles into strong 3D shapes, which are not solvable by altering the% hydroxylation of lysine or proline residues. However, if the logjam is in fibril formation or the need for collagen isotypes, then progress at the level of recombinant sequencing is the key.
This special issue focuses on the smallest scale of collagen material engineering—that of synthesizing human collagen amino acid sequences as fibril-forming monomers. A number of routes are available, but the promise here is in the modification of gene expression in plants, such that functional collagen monomer accumulates. Collagen expression in plants has a number of advantages, not least eliminating potential animal-derived infections. But, it also brings its own obstacles, involving both the collagen monomer itself, and the host plant.
The collagen itself is complex to generate synthetically, involving an unusually long sequence of post-translational (enzyme-dependent) modifications, site-specific hydroxylations, directional folding, glycosylation, precursor cleavage, and crosslink formation.14,15 Some of these are (i) basic to monomer stability, or (ii) give the collagen its tissue-specific fine detail, and (iii) influence downstream aggregation properties. In addition, plant enzyme systems may insert plant-specific glycosylation patterns with inflammatory of immunogenic tendencies in mammals.
Research summarized here, has focused on the additional insertion of mammalian enzyme systems to produce appropriate post-translational modifications, in sequence, at a reasonable cost and consistency. Furthermore, expressing synthetic collagen in plants has required stages, which change the normal pattern of protein glycosylation, as applied to collagen. The effectiveness of some of these stages is easy to verify because they affect the integrity of the basic collagen molecule (i). These could also influence the rate and consistency of collagen gelling under cell-friendly process conditions and influence cell responses at the next hierarchy scale (ii), for example, through the fibril diameter or helix-trimer composition. However, their functional influence at larger scales (iii & iv) will be modest, and much more difficult to predict.
Problems with plant-based, inflammation can be, and have been tested in this issue, by implantation into animals. It is also likely that the nature of the synthetic collagen monomer (e.g., hydroxylation, crosslink content) will to some currently unknown extent affect material mechanical properties at the cell and gross levels. Critically, however, until we understand how to aggregate, weave, bundle, layer, and zone the collagen at these different levels, we cannot in practice predict which basic monomer properties are needed. Small dispersed researcher communities have been trying to understand how to control/engineer the fibril density, 3D alignment, fibril/fiber diameter, and bundle structure, correlated with material mechanics. However, it is now clear that control of collagen aggregation assembly at larger scales is fast becoming a limiting step, at least as important as the monomer. First, we cannot really test recombinant collagens properly until we can form realistic materials from them. Second, we must eventually understand the control of assembly at this level anyway, using animal-extracted collagen monomer as the bottom, building block. Importantly, material remains the second key word in our starting phase collagen material engineering.
Summary
Current tissue-engineering approaches to directly fabricate tissues, with minimal cell reliance, favor the use of natural protein support materials over temporary synthetic scaffolds. Among the options for natural protein materials, collagen is a strong candidate as, in effect, it represents the finished biological support material for mammalians. Although a spectrum of collagen-derived materials has been in common clinical use for many years, it is argued that a new generation of engineered collagen is now needed as the feedstock for bottom-up tissue fabrication. The key to this research challenge is the need to focus on producing material properties over one range of the huge length scale (from nm to cm), which makes up the collagen aggregation hierarchy. The present special issue deals with cutting edge approaches to control production of material building blocks at the smallest scale, recombinant human, and fibril-forming collagen monomers—in this case, engineered in plants.
Footnotes
Disclosure Statement
No competing financial interests exist.