
Abstract
Neural progenitor cells (NPCs) derived from pluripotent stem cells (PSCs), including embryonic stem cells (ESCs) and induced PSCs, are promising cell source for the treatment of various neurological diseases. NPC derivation from PSCs is regulated by microenvironment factors that influence cell fate via paracrine and autocrine effects. In this study, ESC-derived NPC aggregates were replated in the secretomes of bone marrow-derived human mesenchymal stem cells (hMSCs) generated under hypoxia or normoxia to investigate the effects of hMSC secretome on NPC cellular behaviors. The results demonstrated that hMSC secretomes stimulated endogenous secretion of extracellular matrices from NPC aggregates and enhanced cell adhesion and proliferation. NPC functional differentiation measured by migration length, neurite extension, and the yield of neural and glial cells were also increased by threefold to fourfold. Inhibition of fibroblast growth factor-2, transforming growth factor-β1, and brain-derived neurotrophic factor signaling differentially reduced the adherent cell number, migration length, and neurite extension, suggesting the regulatory effects of a broad spectrum of hMSC-derived factors. In summary, ESC-derived NPC aggregates in hypoxic hMSC secretomes may represent a suitable combination to promote the engraftment and neurogenesis in vivo.
Introduction
N
Mesenchymal stem cells (MSCs) have been tested in preclinical and clinical studies for the treatment of neurodegenerative diseases.14,15 However, transplanted MSCs have short life span in vivo and their capacity to undergo neuronal differentiation for long-term engraftment is being intensely debated.16,17 Recent studies have shown that the primary mechanism of MSC's therapeutic effects is the secretion of bioactive molecules that induce anti-inflammatory, anti-apoptotic, neural trophic, and immune-modulating effects, which promote tissue healing by modulating the milieu of injury sites, suppressing immune responses,15,16 and/or stimulating endogenous neurogenesis.18–20 For examples, co-culturing MSCs with adult NPCs enhanced cell proliferation and promoted neurite extension by upregulating Notch signaling and the expressions of cell adhesion molecules and ECM proteins (i.e., N-cadherin, fibronectin [FN], and laminin [LN]).21,22 These enhancements are thought to be induced by MSC secretome because addition of conditioned medium (CM) has been found to have similar effects on NPC differentiation.14,20,23 While these studies demonstrated the benefits of MSC's trophic effects on NPC differentiation and CNS tissue repair, the mechanism underpinning the trophic effects and the impact of hypoxic secretome are not fully understood. 14
NPC derivation from PSCs has been carried out either in monolayer cultures or from so-called neurospheres through the formation of embryoid bodies (EBs).24,25 NPCs derived from monolayer cultures have a high degree of homogeneity but lack terminal neuronal maturation with limited neurite network. 26 In contrast, NPC aggregates with a heterogeneous neural cell population generated a dense neurite network and mature neuronal electrophysiology.24,26 Additionally, transplantation of NPCs in 3D microenvironment either as neural aggregates or with 3D ECM scaffolds promoted cell retention, adhesion, survival, and differentiated function.27,28 These studies suggest that the increased ECMs and the formation of proper milieu in the NPC aggregates play important roles in enhancing NPC proliferation, differentiation, and functional neural development.26,29
The goal of the present study is to investigate the influence of human MSC (hMSC) secretome generated under hypoxia or normoxia on cellular behaviors of ESC-derived NPC aggregates. It is hypothesized that hMSC secretome provides a milieu of macromolecules that promote NPC aggregate attachment, survival, proliferation, and migration and that hypoxic precultures could further enhance the trophic effects of hMSC secretomes. Understanding the role of hMSC secretome in regulating cellular events of PSC-derived NPCs will reveal the paracrine effect of hMSCs on endogenous neurogenesis and provide a novel approach to derive NPCs from PSCs for potential co-transplantation with hMSCs.
Materials and Methods
Undifferentiated ESC culture and generation of NPCs
Murine ES-D3 line (American Type Culture Collection, Manassas, VA) was maintained on 0.1% gelatin-coated six-well plates in a standard 5% CO2 incubator. The expansion medium is composed of Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% ESC-screened fetal bovine serum (FBS; Hyclone, Logan, UT), 1 mM sodium pyruvate, 0.1 mM β-mercaptoethanol, 100 U/mL penicillin, 100 μg/mL streptomycin (all from Invitrogen), and 1000 U/mL leukemia inhibitory factor (Millipore, Billerica, MA). The cells were seeded at 2–4×104 cells/cm2 and subcultured every 2–3 days.
To derive NPCs, ESCs were seeded at 1×106 cells into Ultra-Low Attachment six-well plates (Corning Incorporated, Corning, NY) in 3 mL of DMEM-F12 plus 2% B27 serum-free supplement (Invitrogen) using a 4−/4+ protocol. 30 At day 4, all-trans retinoic acid (Sigma-Aldrich, St. Louis, MI) was added at 1 μM in the differentiation medium. NPC aggregates were collected at day 8 to assess the effects of hMSC secretomes (Fig. 1).
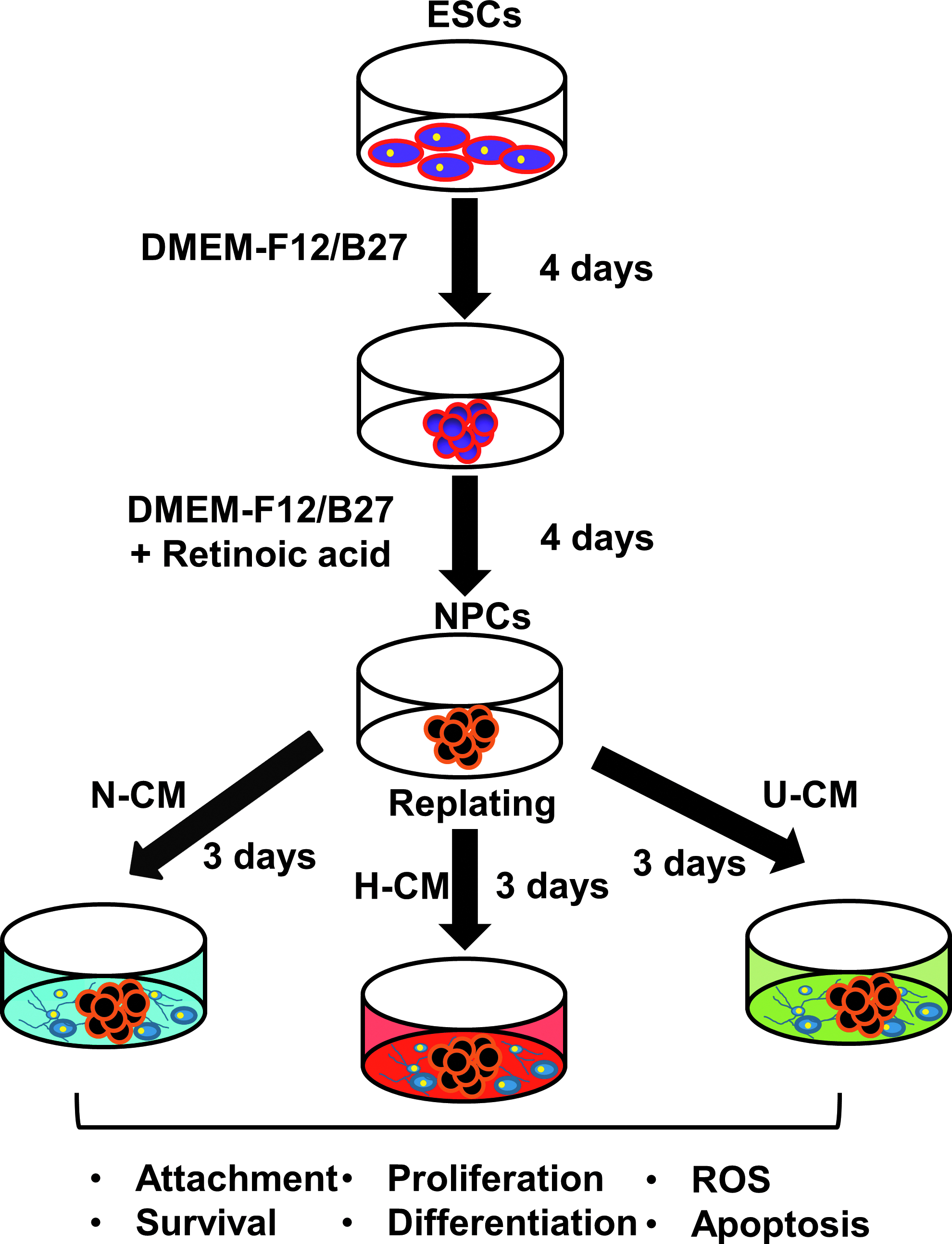
Schematic diagram of the experimental procedure. Embryonic stem cells (ESCs) were seeded in low attachment plates to form embryoid bodies in Dulbecco's modified Eagle's medium (DMEM)-F12 plus B27 for 4 days. Afterwards the medium was supplemented with retinoic acid for another 4 days to obtain neural progenitor cell (NPC) aggregates. NPCs were replated on gelatin-coated dishes in conditioned media under hypoxia (H-CM), conditioned media under normoxia (N-CM) or unconditioned media (U-CM) and cultured for 3 days before the characterizations. ROS, reactive oxygen species. Color images available online at www.liebertpub.com/tea
hMSC culture and collection of CM
Standardized frozen hMSCs from multiple donors were obtained from the Tulane Center for Gene Therapy and cultured as previously described.31,32 The MSCs were isolated from the bone marrow of healthy donors ranging in age from 19 to 49 years based on plastic adherence, negative for CD34, CD45, and CD117 (all <2%) and positive for CD29, CD44, CD49c, CD90, CD105, and CD147 markers (all >95%), and possess tri-lineage differentiation potential upon induction in vitro.33,34 Briefly, hMSCs were expanded at a density of 1.7×103 cells/cm2 using αMEM (Invitrogen) supplemented with 10% FBS and 1% penicillin/streptomycin in a standard 5% CO2 incubator. At ∼80% confluence, adherent cells were harvested with 0.25% trypsin–EDTA (Sigma-Aldrich) and further propagated. Passage 5 cells were used for collecting CM.
CM collection was performed as previously described. 35 Briefly, hMSCs were seeded at a density of 1500 cells/cm2 and incubated under hypoxia (2% O2) or normoxia (20% O2). After 7 days, the cultures were gently washed three times with phosphate-buffered saline (PBS), and fed with serum-free αMEM supplemented with 1% penicillin/streptomycin. After 24 h, conditioned media under normoxia (N-CM) or hypoxia (H-CM) were collected and filtered through a 0.2-μm filtration device. The αMEM without exposing to hMSCs was used as control (unconditioned media: U-CM). For neural differentiation, N-CM, H-CM, or U-CM were mixed at 50% v/v with DMEM-F12 plus B27 media.
NPC aggregate culture in the presence of hMSC secretomes
At day 8, NPC aggregates were replated on 0.1% gelatin-coated 24-well plates and cultured for 3 days in N-CM, H-CM, or U-CM. Adherent cell numbers were determined at 24, 48, and 72 h using a hemocytometer after trypsin/EDTA dissociation. Floating aggregates in the culture supernatants were also collected, dissociated by trypsin/EDTA, and counted for cell numbers. To assess the role of growth factor signaling in hMSC secretome during NPC differentiation, the following inhibitors were added in the media: PD173074 (20 nM; Sigma-Aldrich), a FGFR1/FGFR3 inhibitor, 36 to test the effect of fibroblast growth factor (FGF)-2; SB431542 (10 μM; Sigma-Aldrich), a SMAD2 inhibitor, 37 to test the effect of transforming growth factor (TGF)-β1; rabbit anti-BDNF (1:100; Millipore) to test the influence of BDNF. The cultures were treated with the inhibitors or anti-BDNF for 72 h and evaluated for cell growth, migration, and neurite length during this period.
Bromodeoxyuridine proliferation assay
The proliferation of the cells surrounding NPC aggregates at day 2 after replating was assessed using bromodeoxyuridine (BrdU) staining. Briefly, the cells were incubated in a DMEM-F12 plus B27 medium containing 10 μM BrdU (Sigma-Aldrich) for 90 min. The cells were then fixed with 70% cold ethanol, followed by a denaturation step using 2 N HCl/0.5% Triton X-100 for 30 min in the dark. The samples were reduced with 1 mg/mL sodium borohydride for 5 min and incubated with mouse anti-BrdU (1:100; Invitrogen) in blocking buffer (0.5% Tween 20/1% bovine serum albumin in PBS), followed by the incubation with Alexa Fluor® 488 goat anti-Mouse IgG1 (1:100; Molecular Probes, Grand Island, NY). The cells were mounted with 4′,6-diamidino-2-phenylindole (DAPI) and visualized using a fluorescent microscope (Olympus IX70, Melville, NY). Both the samples without incubation with BrdU and the samples treated with BrdU but stained with secondary antibody only were used as negative controls.
Cell cycle analysis
Two days after replating, the distribution of attached NPCs in various phases of cell cycle was assessed after the trypsin/EDTA dissociation. 38 Cells were fixed and permeabilized with 70% ethanol at 4°C for 40 min. The samples were then centrifuged and resuspended in a solution containing 20 μg/mL propidium iodide (PI; Sigma-Aldrich) plus 0.5 mg/mL RNAse A (Sigma-Aldrich). The samples without PI staining were used as negative control. After the incubation and washing, the cells were acquired with BD FACSCanto™ II flow cytometer and the results were analyzed using FlowJo software.
Neurite length and migration analysis
The migration length was assessed using a light microscope (Olympus IX70). The distance between the edge of the attached aggregates and the furthest migrated cells was determined at five distinct locations per aggregates using ImageJ software. For the neurite length, the distance between the edge of attached aggregates and the extremities of at least eight neurites per aggregates were calculated using ImageJ software. At least three spheres per condition were analyzed. The cells from U-CM group were used as negative control.
Live/Dead, reactive oxygen species, and poly-caspase assays
Live/Dead assay
Cell viability was assessed using LIVE/DEAD® staining kit (Molecular Probes). The attached cells were incubated in DMEM-F12 containing 1 μM calcein AM (green color for live cells) and 2 μM ethidium homodimer I (red color for dead cells) for 30 min. The samples were then washed and imaged under a fluorescent microscope. The live cell controls were the normal ESC stock cultures. The dead cell controls were prepared by treating the cells with 70% methanol for 30 min.
Reactive oxygen species assay
Reactive oxygen species (ROS) was assessed using Image-iT™ Live Green ROS Detection kit (Molecular Probes). 39 Briefly, the aggregates were washed in Hank's Balanced Salt Solution and incubated in a solution of 25 μM carboxy-H2DCFDA for 30 min at 37°C. The samples were imaged under fluorescence microscope to visualize ROS location in the aggregates. For quantification, the fluorescence intensity was analyzed using ImageJ software. As the positive control, the cells were incubated in 100 μM tert-butyl hydroperoxide (TBHP) prior to staining with carboxy-H2DCFDA. The negative controls were the cultures without staining with carboxy-H2DCFDA.
Caspase assay
Caspases were detected using Image-iT Live Green Caspase Detection Kits (Molecular Probes). Briefly, the aggregates were incubated for 1 h with fluorescent inhibitor of caspases (FLICA) reagent and imaged under fluorescence microscope. The fluorescence intensity was analyzed by ImageJ software. The cultures without staining with FLICA were used as negative controls.
Immunocytochemistry and enzyme-linked immunosorbent assay
For ECM expression, adherent or suspended NPC aggregates were fixed with 4% paraformaldehyde (PFA) and permeabilized with 0.2–0.5% Triton X-100. The samples were then blocked and incubated with primary ECM antibodies at 1:100 dilutions, including rabbit polyclonal FN, LN, collagen type IV (COL IV), and vitronectin (VN) (Abcam, Cambridge, MA).35,39 After washing, the samples were incubated with Alexa Fluor 488 goat anti-Rabbit IgG (1:100; Molecular Probes), counterstained with DAPI, and visualized using a fluorescence microscope. To detect differentiation markers, the cells were incubated with mouse or rabbit primary antibody against the following: Nestin (1:200; Sigma-Aldrich) and Musashi-1 (1:200; Abcam) for progenitors, β-tubulin III (1:200; Millipore) for neurons, and glial fibrillary acidic protein (GFAP, 1:200; Millipore) for astrocytes. After washing, the cells were incubated with the corresponding secondary antibody: Alexa Fluor 488 goat anti-Mouse IgG1 (for GFAP and β-tubulin III) or Alexa Fluor 488 goat anti-Rabbit IgG (for Nestin and Musashi-1) at 1:100 dilutions, and the cells were observed under fluorescence microscope. The samples with secondary antibody only were used as negative controls.
To quantify ECM contents, 40 the adherent or suspended NPC aggregates in different conditions were incubated with donkey anti-Rabbit IgG conjugated with horseradish peroxidase (HRP; Rockland Immunochemicals, Inc., Gilbertsville, PA) after the incubation with primary antibodies. After washing, 1 mL of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (Thermo scientific, Hudson, NH) was added to the samples, incubated for 5–25 min, and stopped by a solution of 0.1 M sulfuric acid. The absorbance units (AUs) were measured using a microplate reader (Bio-Rad, Richmond, CA) at a wavelength of 405 nm with background subtraction at 655 nm. The AU values were corrected by subtracting the values of negative control stained with HRP-IgG only. The measurements (i.e., indicating total ECM contents) were also normalized by the cell numbers to obtain specific ECM contents. To evaluate the effect of FGF signaling on extracellular signal-regulated kinase (ERK) pathway, cell lysates were collected and the levels of pERK were quantified by enzyme-linked immunosorbent assay (ELISA) following the manufacturer's instructions (Enzo Life Sciences, Farmingdale, NY). The specific pERK expression was calculated with AU values normalized to cell numbers. To quantify the growth factors in CM, the concentrations of FGF-2 and TGF-β1 in N-CM and H-CM were measured by ELISA according to the manufacturers' instructions (R&D Systems, Minneapolis, MN for FGF-2; Life Technologies for TGF-β1). The presence of BDNF was evidenced in blocking assay using anti-BDNF.
Flow cytometry
For flow cytometry, 1×106 cells/sample were fixed with 4% PFA and permeabilized with 100% cold methanol. After blocking, the samples were incubated with primary antibodies against Nestin (1:200), β-tubulin III (1:200), GFAP (1:200), or Musashi-1 (1:200) followed by the corresponding secondary antibody (1:100 dilutions): Alexa Fluor 488 goat anti-Mouse IgG1 (for GFAP and β-tubulin III) or Alexa Fluor 488 goat anti-Rabbit IgG (for Nestin and Musashi-1). The cells were acquired with BD FACSCanto II flow cytometer and analyzed against isotype controls using FlowJo software.
Statistical analysis
Each experiment was carried out at least three times. The average values of two or three independent experiments were presented and the results are expressed as (mean±mean absolute deviation). In each experiment, triplicate samples were used. To assess the statistical significance, ANOVA followed by Fisher's LSD post hoc tests. A p-value<0.05 was considered statistically significant.
Results
hMSC secretome promoted ECM secretion of ESC-derived NPC aggregates
After 8 days of differentiation, NPC aggregates were replated in the presence of N-CM, H-CM, or U-CM for an additional 3 days. The expression of endogenous ECMs in the attached cells was evaluated (Fig. 2A–H). FN was able to spread on the gelatin-coated surface to the area without cells, while LN, COL IV, and VN were mainly detected close to the cell membrane. Because the culture medium was serum-free and there was no exogenous FN, LN, COL IV, and VN used for coating, the expression indicated the endogenous ECM secretion from NPCs. The specific expressions of FN, LN, COL IV, and VN were similar for attached NPCs cultivated in N-CM, H-CM, and U-CM, while the expressions of total ECM proteins were enhanced by threefolds in N-CM/H-CM versus U-CM (Fig. 2I, J). The floating NPC aggregates were also assessed for endogenous ECM expression. N-CM and H-CM conditions displayed about twofold increase in specific expression of ECM proteins compared with U-CM (Fig. 2K). The expressions of total ECM proteins were also increased (1.3- to 2.0-folds) in N-CM/H-CM compared with U-CM (Fig. 2L). Hence, hMSC secretomes promoted endogenous ECM secretion of ESC-derived NPC aggregates.

Extracellular matrix (ECM) expression of ESC-derived NPC aggregates in N-CM, H-CM, and U-CM.
hMSC secretome promoted adhesion and proliferation of ESC-derived NPC aggregates
The impact of hMSC secretomes on adhesion and proliferation of ESC-derived NPC aggregates were evaluated. N-CM and H-CM significantly increased cell attachment and outgrowth compared with U-CM based on morphology observation (Fig. 3A). In all conditions, the cells around the aggregates displayed a heterogeneous morphology composed of flat cells and bipolar cells with neurite projections, suggesting the presence of a mixed population of mature neurons and neural progenitors. 41 The adherent cell numbers (including the migrating single cells and the attached aggregates) were threefold to fivefold higher for N-CM and H-CM than U-CM, while no significant difference was observed between N-CM and H-CM (Fig. 3B). Consistently, the percentage of floating cells in U-CM was also higher (90% vs. 40–60%) compared with N-CM and H-CM (Fig. 3C), suggesting that N-CM and H-CM favored NPC aggregate adhesion.

Adhesion and proliferation of ESC-derived NPC aggregates in N-CM, H-CM, and U-CM.
To further analyze the status of adherent cells, BrdU incorporation was evaluated to detect the cells in S phase. More BrdU+ cells (about 2.8-folds) were observed for cells around NPC aggregates in N-CM and H-CM conditions compared with U-CM (Fig. 3D, E). To confirm this observation, cell cycle analysis was performed 24 h postreplating. About 28–29% NPCs in N-CM and H-CM were in S-phase, while only 17% for U-CM (Fig. 3F). Conversely, 74% of cells in G0/G1 were observed for U-CM compared with 60–63% in N-CM/H-CM conditions. Hence, N-CM and H-CM promoted NPC proliferation, which also contributed to the higher adherent cell numbers.
hMSC secretome regulated the survival of ESC-derived NPC aggregates
To interrogate whether the higher cell numbers in N-CM/H-CM versus U-CM is related to cell survival, cell viability, ROS, and caspase expression were assessed. NPCs cultivated in N-CM/H-CM displayed higher cell viability compared with U-CM (75% vs. 45%) (Fig. 4A, B). Similarly, caspase expression for NPCs in N-CM/H-CM was about 1.7-fold lower compared with U-CM (Fig. 4C). Because ROS may induce caspase expression, the ROS levels were also measured (Fig. 4D–F). ROS expression was about 1.8-fold lower in H-CM compared with N-CM and U-CM in the absence of TBHP (Fig. 4E). Upon ROS induction using TBHP, about threefold less ROS expression was observed in H-CM (Fig. 4F), indicating that H-CM may provide better antioxidant effect compared with N-CM and U-CM.
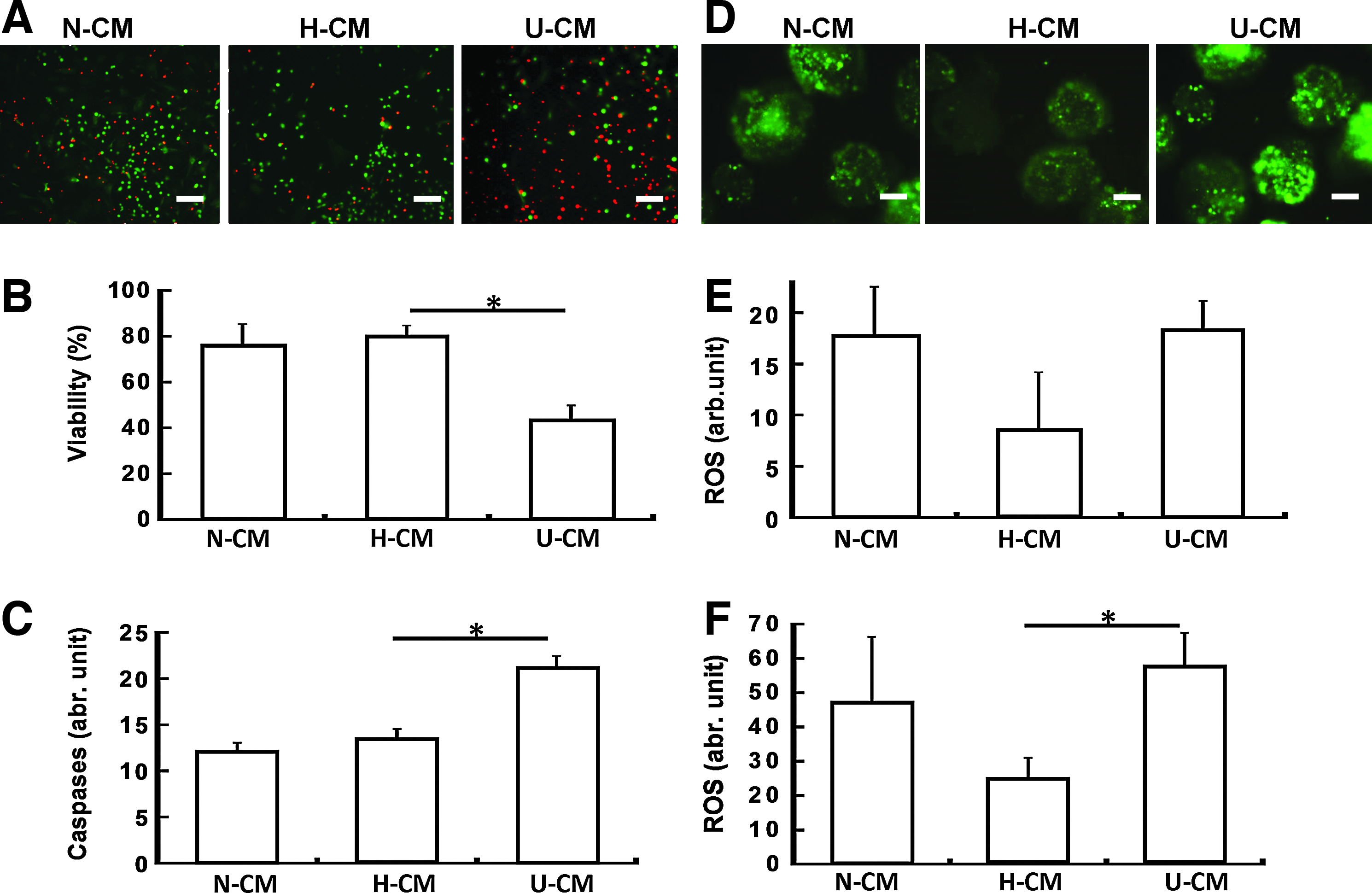
Cell survival, caspase, and ROS expression of ESC-derived NPC aggregates in N-CM, H-CM, and U-CM.
Effects of hMSC secretome on neural differentiation of ESC-derived NPC aggregates
After growing for 3 days, NPCs were assessed for the expression of neural markers Nestin (progenitor), Musashi-1 (progenitor), GFAP (astrocyte), and β-tubulin III (neuron) (Fig. 5). Similar percentages of marker expression were observed for all three conditions: 44–48% β-tubulin III, 20–24% GFAP, 58–68% Nestin, and 82–85% Musashi-1 (Fig. 5A, B). Before the treatment, day 8 NPCs expressed about 60–70% Nestin, 80–90% Musashi-1, and low levels of GFAP and β-tubulin III (<10%), and thus NPC aggregate replating in hMSC secretomes resulted in neural differentiation. About 2.5- to 3.0-fold increase in the positive cell numbers for each marker was observed for N-CM/H-CM conditions compared with U-CM condition (Fig. 5C), which may be directly linked to the increased total cell numbers in N-CM/H-CM.
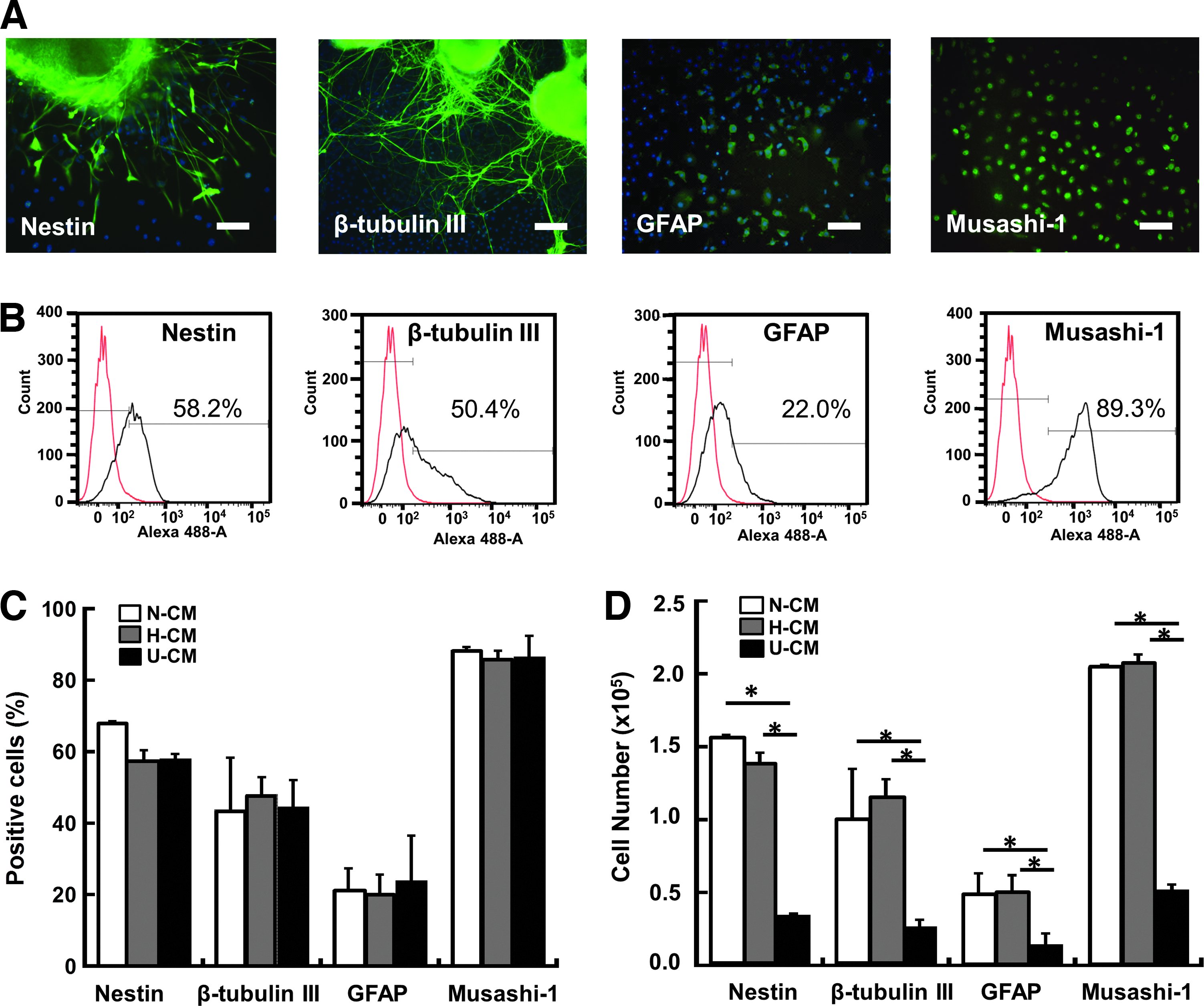
Neural differentiation of ESC-derived NPC aggregates in N-CM, H-CM, and U-CM.
Although the percentages of neural markers were similar, quantitative analysis indicated about threefold to fourfold longer neurite length in N-CM/H-CM compared with U-CM (i.e., 600 μm vs. 180 μm at 72 h) (Fig. 6A, C). In addition, NPC migration length from the aggregates was increased by twofold to threefold in N-CM/H-CM compared with U-CM condition (i.e., 340–450 μm vs. 170 μm at 72 h) (Fig. 6B, D). No significant difference in neurite length was observed between N-CM and H-CM treated cells, while migration length was slightly longer for H-CM than N-CM treated cells.
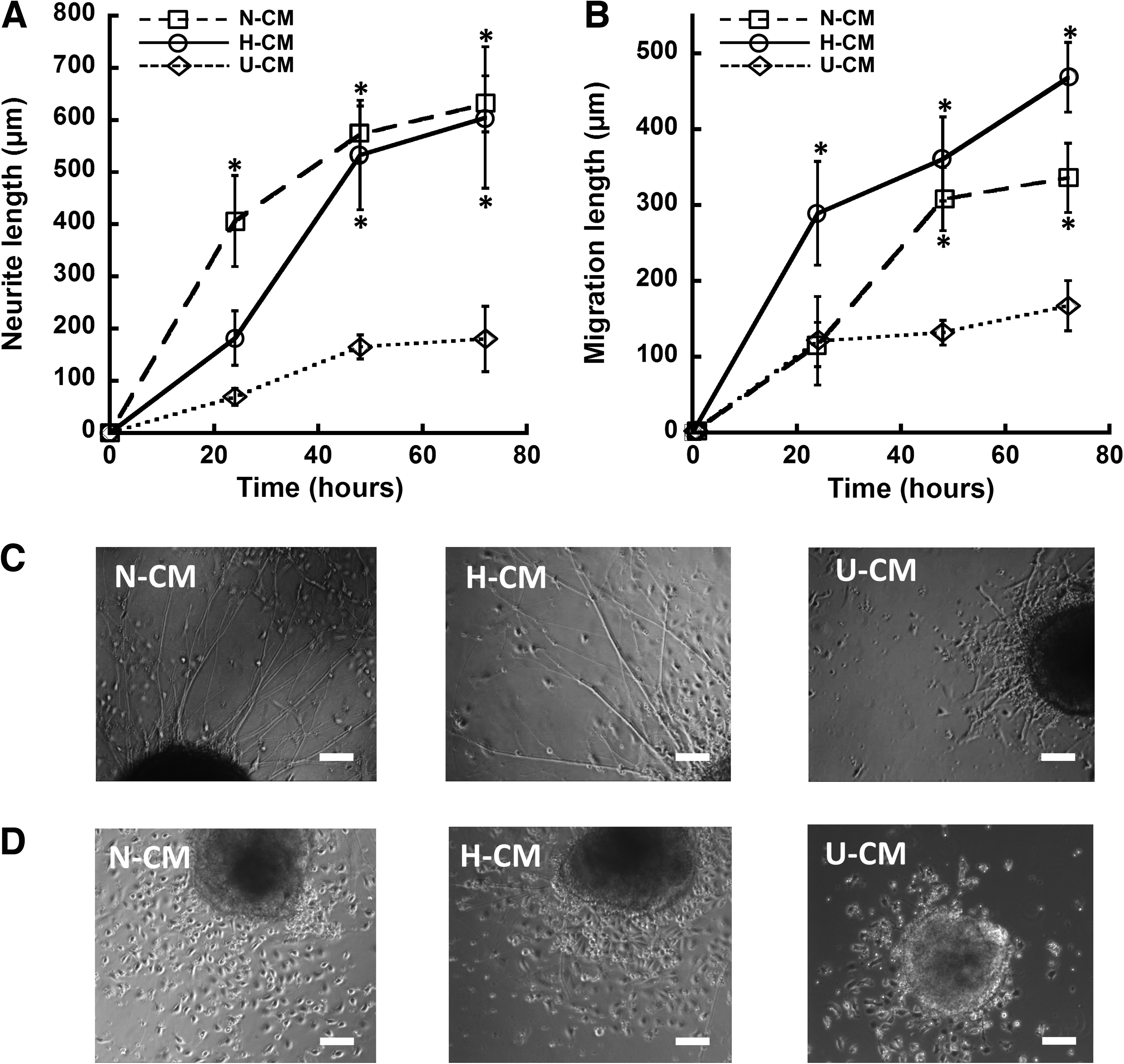
Neurite extension and cell migration of ESC-derived NPC aggregates in N-CM, H-CM, and U-CM.
Effects of FGF-2, TGF-β1, and BDNF signaling
To further delineate the effects of hMSC secretomes, the presence of FGF-2 and TGF-β1 in N-CM and H-CM were analyzed (Table 1). H-CM contained higher concentration of FGF-2 than N-CM but has similar level of TGF-β1. To illustrate the effects of FGF-2 and TGF-β1, inhibitors for FGF-2 and TGF-β1 were tested in the cultures. As BDNF was another important growth factor in MSC-CM, 42 the blocking effect of anti-BDNF was also evaluated. After PD173074 (a FGFR inhibitor) treatment, the adherent cell numbers significantly decreased (8–10×105 vs. 2–4×105) in N-CM/H-CM or modestly (3–4×105 vs. 0.5–1×105) in U-CM (Fig. 7A). The decrease in U-CM condition indicated the inhibition of endogenous FGF signaling from NPCs. 43 No difference in neurite length was observed with PD173074 treatment (Fig. 7B), while migration length decreased (400–500 μm vs. 200–300 μm) for N-CM and H-CM (Fig. 7C). Because FGF-2 binding triggers the phosphorylation of ERK activity, 35 pERK expression was also evaluated. A decrease in pERK activity was observed after PD173074 treatment in all three conditions, further confirming the presence of FGF signaling (Fig. 7D).

Effect of fibroblast growth factor-2 signaling on ESC-derived NPC aggregates in N-CM, H-CM, and U-CM. The cells with or without treatment of PD173074 were compared.
Indicates average concentrations from three collections of conditioned media.
Indicates significant difference (p<0.05).
BDNF, brain-derived neurotrophic factor; FGF, fibroblast growth factor; H-CM, conditioned media under hypoxia; N-CM, conditioned media under normoxia; TGF, transforming growth factor.
TGF-β1 inhibitor SB431542 significantly reduced the adherent cell numbers (3.5–4.2×105 vs. 0.5–1.8×105) in N-CM and H-CM (Fig. 8A). The cell number in U-CM was also reduced with SB431542 treatment, indicating the presence of endogenous TGF-β1 signaling from NPCs. In addition, reduced neurite length (630–690 μm vs. 160–190 μm) and migration length (570–600 μm vs. 160–200 μm) were observed in the presence of SB431542, indicating the impact of TGF-β1 signaling on neural cell functions (Fig. 8B, C).
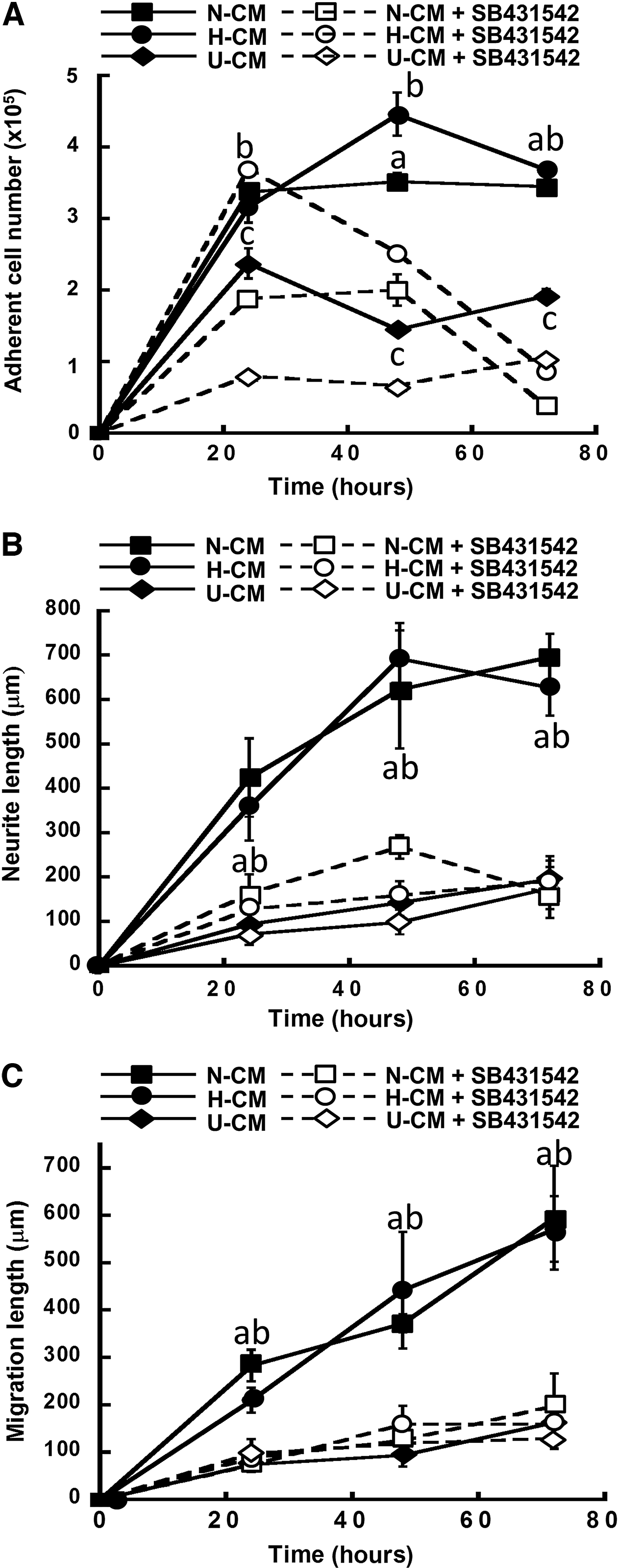
Effect of transforming growth factor-β1 signaling on ESC-derived NPC aggregates in N-CM, H-CM, and U-CM. The cells with or without treatment of SB431542 were compared.
Inhibition of BDNF signaling with anti-BDNF significantly reduced the adherent cell numbers (3.2–3.6×105 vs. 1.1–1.5×105) in N-CM/H-CM and in U-CM (1.7×105 vs. 0.8×105) (Fig. 9A). The decrease of adherent cell number in U-CM indicated endogenous BDNF secretion by NPCs. The neurite extension was reduced after treating with BDNF antibody (480–650 μm vs. 300–450 μm), but no significant difference was observed for migration length (Fig. 9B, C). The inhibition effects of these three molecules were summarized in Table 2. Taken together, N-CM/H-CM conditions enhanced FGF-2, TGF-β1, and BDNF signaling and differentially promoted NPC proliferation, migration, and neurite extension.
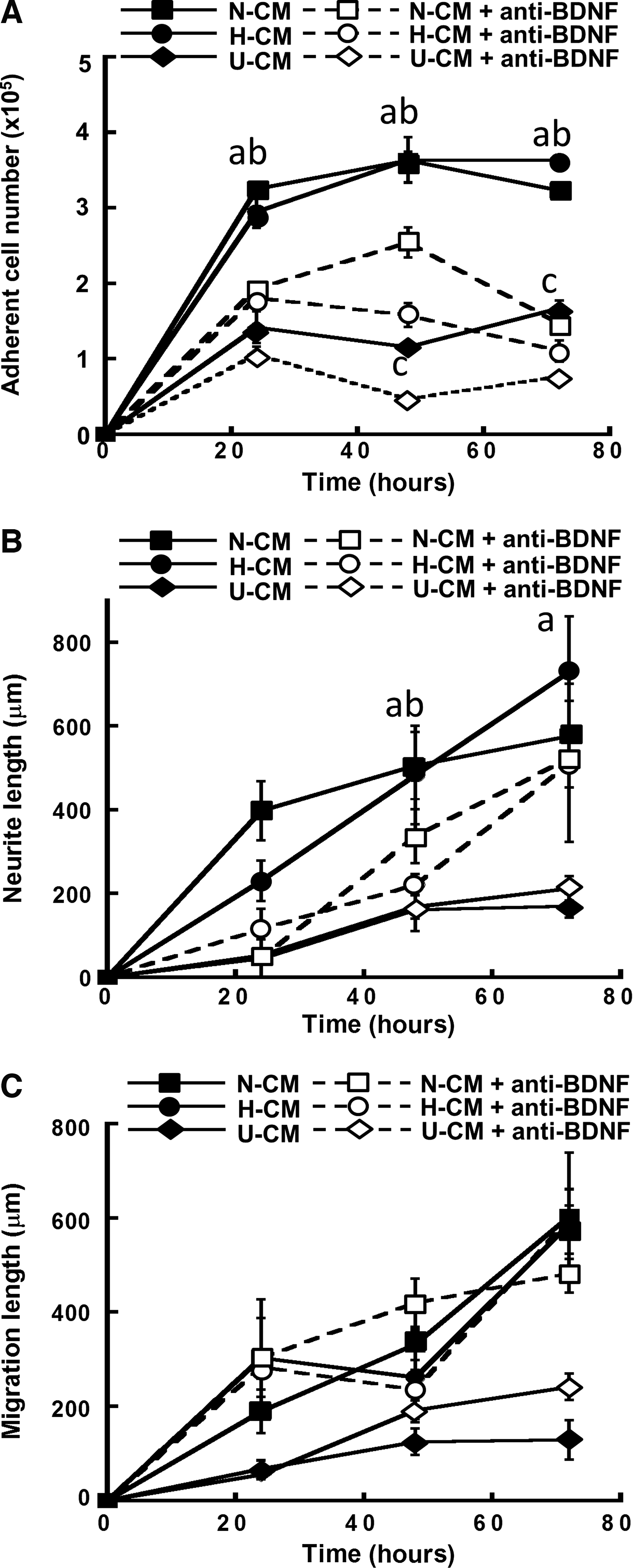
Effect of brain-derived neurotrophic factor (BDNF) signaling on ESC-derived NPC aggregates in N-CM, H-CM, and U-CM. The cells with or without treatment of anti-BDNF antibody were compared.
Percentage of inhibition was calculated using the average values of 48 and 72 h after replating (from Figs. 7–9) as follows:
Indicates significant difference (p<0.05) between N-CM/H-CM and U-CM groups.
Indicates significant difference (p<0.05) between N-CM and H-CM groups.
U-CM, unconditioned media.
Discussion
NPC derivation from PSCs is a promising approach to obtain renewable neural cells but the derivation process is less than robust and the NPC properties remain to be improved. Methods such as niche-dependent and ECM-supported differentiations have been tested to improve derivation efficiency and promote NPC maturation.26,44 hMSC secretome contains a diverse range of bioactive molecules that regulate the cellular events underpinning NPC differentiation and maturation. The results of the current study revealed that hMSC secretome enhanced ESC-derived NPC proliferation, migration, and neurite extension primarily through regulating the molecular milieu in NPC aggregates including ECM proteins and growth factors.
The presence of hMSC secretome influenced the ratio of adherent to nonadherent NPC aggregates derived from ESCs. The heterogeneity of ESC-derived NPC aggregates is a result of differential cell adhesion properties, while treatment with hMSC secretome appeared to increase NPC adhesion due to the upregulation of ECM proteins such as LN and FN especially in floating aggregates. In vitro, ECM proteins have been shown to regulate the survival, proliferation, and neurite outgrowth of human ESC-derived NPCs in a dose-dependent manner through integrin-ECM signaling.41,45 In vivo, ECM proteins are involved in neural development and have shown to support neurogenesis from the transplanted NPCs or endogenous progenitors by promoting cell survival and providing a neuro-permissive environment.28,46 The mechanism for the enhanced ECM secretion in ESC-derived NPC aggregates may involve multiple factors and regulatory pathways emanating from hMSC secretome. FGF signaling is known to induce the transcription of LN-1 and COL IV in EBs, affecting the ECM microenvironment. 47 The presence of tissue inhibitor of metalloproteinase-1 in hMSC secretome may also play a role because it could inhibit ECM degradation by regulating MMP-9 activity. 48 Although the exact balance of the increased ECM secretion versus inhibition of ECM degradation requires further study, multiple factors in hMSC secretome and their concerted effects are likely to be involved.47–49
Increased ECM secretion in the 3D aggregates may serve as reservoirs and in turn enhance local enrichment of growth factors such as FGF-2 and TGF-β1.35,50 FGF-2 is known to bind heparin sulfate proteoglycans and influence cell fate through the activation of mitogen-activated protein kinase with consequent phosphorylation of ERK1/2. 35 FGF-2 is a mitogenic factor and has been commonly used to maintain NPCs at progenitor stage. 4 Indeed, blocking FGF-FGFR signaling using PD173074 abolished the positive effects of hMSC secretomes on cell adhesion and migration, but not neurite extension as shown in our study. TGF-β1 is also a matrix-associating protein and its sequestration and release from ECMs is an important mechanism that regulates its bioactivity.51,52 MSCs are significant source of TGF-β1,37,43,53 which has been shown to promote glial cell differentiation of adult NPCs. 37 TGF-β1 also exerted anti-apoptotic and growth/migration-enhancing activities on NPCs through SMAD2 (for growth and migration) and ERK1/2 (for anti-apoptotic behavior) signaling. 54 In our study, SB431542 treatment reduced NPC adhesion/proliferation, migration, and neurite extension, suggesting the involvement of TGF-β1 signaling in neural development. BDNF, secreted by hMSCs at a higher level than NPCs, is a primary trophic factor that supports the growth and differentiation of neuronal lineage.42,43 BNDF binding to collagen enhanced the neurotrophic effect and promoted axonal regeneration compared with BDNF without collagen-binding domain. 55 In the current study, inhibition of BDNF signaling resulted in the reduced cell number and neurite length but not migration, indicating its contribution in neuronal cell maturation. Together, the results suggested that multiple secretome-derived factors are involved in regulating NPC behaviors from initial adhesion to later neurite extension and that ECM may be an important facilitator for the growth factor-mediated enhancement.
Apart from the factors in the hMSC secretome, endogenous factors secreted from NPCs may also have autocrine effects that regulate NPC fate. NPC aggregates treated with inhibitors in U-CM also showed the reduced adherent cell number (50–68%) as in N-CM (42–63%) and H-CM (58–73%) groups (Table 2). However, the more significant decrease in adherent cell number, neurite length, and migration length following growth factor inhibition in the N-CM/H-CM groups compared with the U-CM group (Figs. 7–9) suggested more pronounced trophic effects originating from the hMSC secretomes.
hMSC secretomes significantly reduced caspase expression and promoted NPC aggregate survival from our results. Besides FGF-2, TGF-β1, and BDNF, MSC secretomes are reported to contain anti-apoptotic and antioxidant factors such as stanniocalcin-1, 14 VEGF, and stromal cell-derived factor-1. 56 These factors may together contribute to the protection of NPCs from ROS damage and caspase activation,14,54,57 thereby increasing cell viability.
In this study, hMSC secretomes did not affect the percentage of differentiating cells but rather increased the yield of neural cells, presumably due to the increased cell adhesion, survival, and proliferation. Whether the restricted effects are due to the inherent secretome properties or variations in secretome collection and treatment time remains to be investigated.58,59 The day 8 NPC aggregates used in this study may be at primitive stage, and a longer treatment with hMSC secretome may be required to influence mature neural populations. The collection time of hMSC secretome at 24 h may also limit its effect on neural differentiation. 58 Nevertheless, the results demonstrated the potential of hMSC secretome to enhance the functions of differentiated NPCs.
Hypoxic secretome H-CM did not enhance NPC adhesion and proliferation compared to N-CM. Although hypoxia could increase the secretion of FGF-2, BDNF, and pro-angiogenic factors such as VEGF compared with normoxia,35,60 the dose level (pg-ng/mL) may not be sufficient to elicit a difference in cell adhesion/proliferation compared to N-CM. However, the more significant decline in NPC numbers by BDNF inhibition in H-CM (63% vs. 42%, Table 1) indicates a higher contribution of hMSC-derived BDNF in H-CM. The reduced ROS expression by NPCs in H-CM compared with N-CM under oxidative conditions (i.e., using TBHP) also suggests an increased antioxidant effect in H-CM compared with N-CM.
Compared to the restricted effects by H-CM in NPC derivation, hypoxia directly influences multiple cellular events during neurogenic differentiation of ESCs. 61 Hypoxia (2% O2) was shown to accelerate early commitment of ground-state ESCs toward primitive neuroectoderm but appear to negatively impact cell viability after 6 days of neural commitment. 62 The effects of hypoxia on the neurosphere derivation are mediated by activation of hypoxia-inducible factors (HIF) and the target genes of erythropoietin, VEGF, and Bcl-2 family members, although HIF-independent pathways may also play a role.63,64 Hypoxia also affects the stage of lineage specification from NPCs. Mild hypoxia (2.5–5% O2) was shown to enhance human neural stem cell differentiation into neuronal and oligodendroglial cells, while under normoxia (20% O2) the cells had the preferential commitment to the astrocyte lineage, possibly due to anti-apoptotic effects indicated by the reduced caspase expression. 65 Compared to H-CM, oxygen tension directly influences neurogenic differentiation of ESCs via multiple mechanisms at molecular levels.
Further study is required to address the temporal effects of the secreted factors, their interactions, and inhibitory effects from the known or unidentified factors in the secretome. To this end, modulating hMSC culture condition is an important means to control the secretome properties. For example, negative factors such as MMP-2 endogenously secreted by hMSCs is known to mediate ECM degradation and negatively influence NPC derivation. 48 Pretreatment by tumor necrosis factor alpha or hypoxia, however, could inhibit the activity of hMSC-secreted MMP-2, highlighting the effects of culture conditions on hMSC secretome.48,66 Recent studies suggest that hMSC culture as 3D aggregates dramatically influences their secretory profiles and produces secretome with enhanced anti-inflammatory properties.67,68 Moreover, studies are also required to address the interplay between multiple factors that coexist in the secretome. It is known that the growth factors in hMSC secretome are able to exert their effects at concentration range well below those of exogenous growth factors typically used in in vitro studies.14,44 Whether the regulatory molecules in hMSC secretome have additive effects on cellular behaviors is yet to be investigated. Understanding these interactions may reveal novel regulatory pathways and lead to novel approach to enhance NPC derivation. To this end, system biology approach is desirable because of its potential to provide a global view of regulatory network during lineage-specific differentiation. 69
Conclusions
hMSC secretomes enhanced endogenous ECM secretion from NPCs and modulated adhesion, proliferation, survival, migration, and neurite extension of ESC-derived NPC aggregates. These regulatory functions were mediated in part by hMSC-derived FGF-2, TGF-β1, and BDNF and possibly their interactions with ECM proteins. H-CM treatment displayed similar cellular responses compared to N-CM condition but had better antioxidant effects. The results demonstrated the potential of hMSC secretome to regulate neural development of PSC-derived NPCs.
Footnotes
Acknowledgments
The authors would like to thank Ms. Ruth Didier of FSU Department of Biomedical Sciences for her help in flow cytometry analysis. This work is supported by FSU startup fund, FSU GAP award, and FSU student technology committee. Funding support in part from the James King Biomedical Research Program (4KB09) is also acknowledged.
Disclosure Statement
No competing financial interests exist.