
Abstract
We investigated the effects of CD14 macrophages and proinflammatory cytokines on chondrogenic differentiation of osteoarthritic synovium-derived stem cells (SDSCs). Osteoarthritic synovial fluid was analyzed for interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and IL-6. Levels of stem cell surface markers in osteoarthritic SDSCs were evaluated using flow cytometry. CD14-negative cells were obtained using magnetically activated cell sorting. We compared chondrogenic potentials between whole cells and CD14-negative cells in CD14low cells and CD14high cells, respectively. To assess whether nuclear factor-κB (NF-κB) and CCAAT/enhancer-binding protein β (C/EBPβ) modulate IL-1β-induced alterations in chondrogenic potential, we performed small interfering RNA transfection. We observed a significant correlation between the CD14 ratio in osteoarthritic SDSCs and IL-1β and TNF-α in osteoarthritic synovial fluid. Phenotypic characterization of whole cells and CD14-negative cells showed no significant differences in levels of stem cell markers. mRNA expression of type II collagen was higher in CD14-negative cell pellets than in whole cell pellets. Immunohistochemical staining indicated higher levels of type II collagen in the CD14-negative cell pellets of CD14high cells than in whole cell pellets of CD14high cells. As expected, IL-1β and TNF-α significantly inhibited the expression of chondrogenic-related genes in SDSCs, an effect which was antagonized by knockdown of NF-κB and C/EBPβ. Our results suggest that depletion of CD14+ synovial macrophages leads to improved chondrogenic potential in CD14high cell populations in osteoarthritic SDSCs, and that NF-κB (RelA) and C/EBPβ are critical factors mediating IL-1β-induced suppression of the chondrogenic potential of human SDSCs.
Introduction
O
Synovial membrane inflammation can occur at a lower grade during the early stages of OA than that observed in rheumatoid arthritis (RA). Histologically, hyperplasia in the OA synovium is associated with an increased number of synovial lining cells, accompanied by infiltration of the inflammatory cells mainly consisting of macrophages. 10 The cellularity of the synovial lining layer has been reported to be correlated with the number of CD14+ macrophages infiltrating the sublining areas. 11 In addition, a variety of proinflammatory and anti-inflammatory cytokines—interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and IL-10, as well as the major matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs)—are present in OA synovial cells. 12 It has been previously shown that targeted depletion of synovial macrophages by using anti-CD14-conjugated magnetic beads in RA synovial cells improves inflammation.13,14 For OA, it has also been shown that OA synovial macrophages likely play an important role in activating the fibroblasts and perpetuating the production of proinflammatory cytokines and destructive enzymes. 15 CD14-negative porcine SDSCs have been reported to possess high chondrogenic potential. 16
Although the levels of proinflammatory cytokines are generally lower in OA than those are in RA, IL-1β and TNF-α have been suggested as the key players in OA synovial inflammation. 12 IL-1β and TNF-α are activated cartilage catabolic factors that induce enzymes that degrade the extracellular matrix, in addition to reducing the synthesis of primary cartilage components, such as type II collagen and aggrecan. 17 IL-1β and TNF-α exert their proinflammatory and catabolic effects through the activation of transcriptional nuclear factor-κB (NF-κB) pathway. 18
NF-κB represents a family of inducible dimeric transcription factors, including Rel (c-Rel), RelA (p65), RelB, NF-κB1 (p50/p105), and NF-κB2 (p52/p100). NF-κBp65 is a key active subunit in NF-κB-mediated transcription in several cell types and its small interfering RNA (siRNA)–mediated knockdown has been documented to inhibit the expression of MMP-2, nitric oxide synthase-2 (NOS-2), and cyclooxygenase-2 (COX-2) in IL-1β-induced chondrocytes.19,20 In addition to NF-κB, IL-1β and TNF-α also induce or activate CCAAT/enhancer-binding protein (C/EBP) family in chondrocytes. The C/EBP family of basic leucine-zipper transcription factors includes the following six known family members: C/EBPα, β, δ, ɛ, γ, and ζ. Activation of C/EBPβ and δ by IL-1β and TNF-α results in the induction of various genes related to inflammation, such as phospholipase A2 and COX-2.21,22 C/EBPs also regulate the expression of matrix proteins, such as pro-α1 and α2 type I collagen, matrix Gla protein, and osteocalcin.23–26
Previously published information is scarce regarding the influence of inflammatory mediators on the chondrogenic differentiation of SDSCs. It has been reported that TNF-α inhibits the chondrogenic differentiation of synovial fibroblasts through p38 mitogen-activated protein kinase pathways; NF-κB-dependent inhibition by IL-1β and TNF-α of the chondrogenic differentiation of human bone marrow (BM) MSCs has also been reported.27,28 To date, however, the direct function of these factors with respect to repression of chondrogenic differentiation has not been clearly elucidated.
We hypothesized that OA synovial macrophages could adversely affect the chondrogenic potential of SDSCs. To test this hypothesis, we compared the chondrogenic potential of isolated CD14-negative SDSCs with nonisolated osteoarthritic SDSCs, and examined the effects of IL-1β and TNF-α on the chondrogenic differentiation of osteoarthritic SDSCs. In addition, we evaluated whether NF-κB (RelA/p65) and C/EBPβ mediate IL-1β-induced suppression of the chondrogenic potential of osteoarthritic SDSCs.
Materials and Methods
Enzyme-linked immunosorbent assay
Synovial fluid levels of IL-1β, TNF-α, and IL-6 were measured by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's protocol (R&D Systems, Minneapolis, MN). About 100 μL of undiluted tissue extract or synovial fluid or culture media, diluted according to the manufacturer's recommendations, were incubated with biotinylated monoclonal antibodies specific for IL-1β, TNF-α, and IL-6. After washing, streptavidin-horseradish peroxidase was added, and the 3,3′,5,5′-tetramethylbenzidine (TMB) chromogen was used for the colorimetric reaction. Absorbance was measured by ELISA reader using 450 nm as the primary wavelength.
Harvesting of synovial tissue and isolation of synovium-derived cells
Patients were recruited from orthopedic clinics at Seoul National University Hospital (Seoul) under Institutional Review Board (IRB)–approved protocols, and all patients gave informed consent. The patient characteristics are shown in Table 1. Human synovial tissues were obtained from 40 patients who were undergoing total knee replacement arthroplasty for OA. A portion of the tissue specimens were frozen and stored at −70°C. Synovial fluid was obtained by aspiration just prior to surgical intervention. After centrifugation to remove cells and debris, aliquots were frozen and stored at −70°C.
BMI, body mass index; CRP, C-reactive protein; ESP, erythrocyte sedimentation rate; OA, osteoarthritis.
A portion of the harvested synovium was rinsed with Dulbecco's phosphate-buffered saline (DPBS; Gibco, Paisley, United Kingdom) and digested overnight with 0.2% collagenase (Sigma, Saint Louis, MO) in low-glucose Dulbecco's modified Eagle's medium (DMEM; Gibco) containing 10% fetal bovine serum (FBS), in a 5% CO2 humidified atmosphere. Cells were collected by centrifugation, washed twice, resuspended in low-glucose DMEM supplemented with 10% FBS and antibiotics (growth medium), and plated at a density of 2×104 cells per 150-mm culture dish (Nunc, Inc., Naperville, IL) at 37°C in a 5% humidified CO2 atmosphere. The medium was replaced every 2–3 days.
Flow cytometric analysis
Selection of positive and negative stem cell markers was based on the minimal surface marker proposed by the International Society of Cellular Therapy. 29 The following mouse anti-human monoclonal antibodies were used directly conjugated to fluorochromes: CD14-phycoerythrin (PE), CD31-PE, CD34-PE, CD44-PE, CD90-PE, and CD105-PE (BD Biosciences, San Diego, CA). Nonspecific monoclonal antibodies, selected after a literature review of specific surface markers of BM-MSCs, were included as controls. Briefly, after harvesting cells upon detachment, cells were washed twice with PBS, centrifuged, and washed in a flow cytometry buffer [1% bovine serum albumin (BSA; Sigma) in PBS]. Aliquots of 2×105 cells were then incubated with directly conjugated monoclonal antibodies and washed twice with a flow cytometry buffer. Nonspecific fluorescence was determined using equal aliquots of cell preparations that had been incubated with anti-mouse isotype monoclonal antibodies (BD Biosciences). After washing, cells were fixed in 1% paraformaldehyde. Data were obtained by analyzing 10,000 events on an FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA) using CellQuest software. Gates were set based on staining with combinations of specific and nonspecific monoclonal antibodies, such that no more than 1% of cells were positive using the nonspecific monoclonal antibody.
Magnetic-activated cell sorting
Selection of CD14 as a reliable macrophage/monocyte marker was based on previous studies.13,15,16,30 For cell sorting, an MSCS method (Miltenyi Biotec, Auburn, CA) was used. Briefly, isolated cells were counted and suspended at a concentration of 107 cells per 100 μL of FCM buffer, followed by incubation with primary antibodies at 4°C for 20 min. Primary antibodies were PE-conjugated monoclonal antibodies (BD Pharmingen, San Diego, CA) for the CD14 markers. Cells were washed, resuspended at 107 cells per 80 μL in magnetic-activated cell sorting (MACS) buffer [PBS with 0.5% BSA and 2 mM EDTA (pH 7.2)] and incubated with 20 μL of anti-PE Ig-microbeads for 15 min at 4°C, before being washed and resuspended with 500 μL of MACS buffer. Cells were separated using a magnetic column (LS+; Miltenyi Biotec) according to the manufacturer's recommendations. Four different experimental groups were established: whole cells of CD14high cells; CD14-negative cells of CD14high cells; whole cells of the CD14low cells, and CD14-negative cells of the CD14low cells. After isolation of the SDSCs by enzymatic digestion, the proportion of CD14 cell surface marker in cells from each donor was assessed using flow cytometry. CD14low cells were defined as <2% of the cells expressing CD14, while CD14high cells were defined as >4% of the cells expressing CD14. Whole cells were defined as the cells collected before magnetic selection and the CD14-negative cells were defined as cells that were collected after column elution while the positive cells remained attached to the column. After magnetic selection, the proportions of CD14 cell surface marker in each group were assessed using flow cytometry. We confirmed that CD14 was rarely expressed in CD14-negative cells. Pellet cultures were then prepared to evaluate the chondrogenic potential after obtaining both whole cells and CD14-negative cells from the same donor.
Chondrogenic differentiation
Chondrogenic differentiation in cell pellets was performed as previously described. 31 Briefly, 5×105 cells were centrifuged at 1500 rpm for 5 min in 15-mL polypropylene tubes and resuspended in 500 μL of a chemically defined chondrogenic medium consisting of low-glucose DMEM supplemented with 1 mM pyruvate, 100 nM dexamethasone (Sigma), 40 μL/mL proline (Sigma), 100 μM ascorbate-2-phosphate (Sigma), and 10 μL/mL ITS premix universal culture supplement (BD Biosciences, Bedford, MA) including 6.25 μg/mL insulin, 6.25 μg/mL transferrin, 6.25 μg/mL selenious acid, 5.35 μg/mL linoleic acid, 1.25 μg/mL BSA, 10 ng/mL TGF-β1 (R&D Systems), and 100 ng/mL BMP2 (R&D Systems). Cell pellets were cultured at 37°C with 5% CO2 for 21 days, and medium was changed every 2 days. Chondrogenic pellets were harvested at 21 days for gene expression analysis and histological evaluation, and glycosaminoglycan (GAG) quantification.
Analysis of the chondrogenic potential by chondrogenic gene expression
After 3 weeks of culture, total RNA was isolated using a Qiagen (Mississauga, Ontario, Canada) RNeasy kit, following the manufacturer's instructions. Ten pellets were cultured to assess the chondrocytic differentiation of synovium-derived cells. cDNA was synthesized using first-strand cDNA synthesis kits (Fermentas, Hanover, MD). Quantitative assessment of the mRNA expression levels of type I, type II, and type X collagen; Sox9; aggrecan; RELA; C/EBPβ; p300; and cAMP-response element-binding protein-binding protein (CREBBP) was performed by real-time polymerase chain reaction (PCR) using TaqMan probes (ABI PRISM 7300 Sequence Detection System; Applied Biosystems, Foster City, CA). TaqMan probe was designed by Assays-by-Design SM service (Applied Biosystems) as follows: type I collagen 5′-[FAM] AAGACGAAGACATCCCACCAATCAC [TAMRA]-3′, type II collagen 5′-[FAM] GGGCCAGGATGTCCAGGAGGCTGGC [TAMRA]-3′, aggrecan 5′-[FAM] ACTGTAACATAGACCAGGAGGTATG [TAMRA]-3′, Sox9 5′-[FAM] CGAGCACTCGGGGCAATCCCAGGGC [TAMRA]-3′, type X collagen 5′-[FAM] CACCATAAAGAGTAAAGGTATAGCA [TAMRA]-3′, C/EBPβ 5′-[FAM] CTACGGGGGCAAGAACTGCAAGAAG [TAMRA]-3′, RELA 5′-[FAM] TGTCCCCAAGCCAGCACCCCAGCCC [TAMRA]-3′, p300 5′-[FAM] CACCATGGAGAAGCATAAAGAGGTC [TAMRA]-3′, CREBBP 5′-[FAM] GTGCTAAGAGGCTGCAGACCACAAG [TAMRA]-3′, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 5′-[FAM] GACTCATGACCACAGTCCATGCCAT [TAMRA]-3′. Expression levels were normalized by subtracting the number of copies of the endogenous GAPDH reference gene from the number of copies of the target gene (ΔCt=Ct of target gene−Ct of GAPDH). Expression of the specific gene was calculated using the formula 2−(ΔCt).
Analysis of chondrogenic potentials by GAG evaluation
The dimethylmethylene blue (DMB) method was used for the detection of total GAG contents in pellets. The media were harvested at each changing of the media during the pellet culture. After 1, 2, and 3 weeks of pellet culture, three pellets per group were harvested. The media and pellets were digested overnight in papain/buffer solution (0.1 M sodium formate, 200 μg/mL papain in 5 mM EDTA, and 5 mM L-cysteine; pH 3.0) at 55°C. After centrifugation, 50 μL aliquots of the papain-digested extracts were pipetted into each well of 96-well plates and 250 μL of DMB dye solution was added. The absorbance of these extracts was determined using an ELISA reader at 530 and 590 nm. Chondroitin-6-sulfate was dissolved to be used as the standard, and the GAG levels were then normalized to total cellular DNA, which was measured by the Indole assay. Briefly, 100 μL of 1 N NaOH and 100 μL of Indole reagent were added to the cell pellets, heated for 10 min at 100°C, and plated in 96-well plates. Total DNA levels were quantified with an ELISA reader at 490 nm.
Analysis of chondrogenic potential by histological evaluation
For histological evaluation, pellets were harvested and fixed in 4% paraformaldehyde for 4 h at room temperature. The fixed pellets were placed into Tissue-Tek embedding medium (Sakura Finetek, Torrance, CA) and frozen at −70°C. Sections of 5-μm thickness were cut on a cryostat at −20°C and mounted onto slides. Sections were air dried and immediately fixed in absolute acetone at −20°C for 15 min. The sections were stained with fast-green solution and Safranin-O solution in the normal manner.
For immunofluorescence, the specimens were digested for 30 min with 50 U/mL chondroitinase ABC (Sigma) in 100 mM Tris-acetate. Nonspecific binding of antibodies was eliminated by incubating the specimens with blocking solution (1×PBS, 5% normal goat serum, and 0.3% Triton X-100) for 10 min at room temperature. Mouse monoclonal anti-collagen 2Ab (NeoMarkers, Fremont, CA) was applied for 1 h at room temperature, followed by incubation with 2 g/mL of FITC-goat anti-mouse Ig Ab for 1 h. Finally, slides were washed three times in PBS and mounted using mounting medium. Staining was observed using a confocal microscope (LSM 510; Carl Zeiss, Jena, Germany).
For immunohistochemistry, endogenous peroxidase activity was blocked by 8–10-h incubation in 0.3% H2O2 in methanol. Sections were then pretreated with 0.2% hyaluronidase in 0.1 M PBS for 30 min at 37°C. The residual enzyme was removed with three 3-min washes with PBS, and blocking solution was applied to the section for 30 min at room temperature. Mouse monoclonal anti-collagen 2Ab (NeoMarkers) was incubated with the sections for 16–24 h at 4°C. After washing with PBS to remove the residual primary antibody, reactivity was detected using biotinylated secondary antibody and streptavidin-peroxidase. Peroxidase activity was visualized using diaminobenzidine as a substrate. The sections were counterstained with hematoxylin.
siRNA transfection into the SDSCs
ON-TARGETplus SMARTpool siRNA, containing a mixture of four SMART selection-designed siRNAs targeting NF-κB, C/EBPβ, and nontargeting siRNA, was purchased from Dharmacon (Lafayette, CO). Lyophilized RNAi (5 nmol) was dissolved in a 250 mL siRNA buffer to yield a concentration of 20 mM. Cells were seeded at least 1 day before siRNA transfection. Cells were transfected using 2 μL/mL DharmaFECT 1 transfection reagent (Dharmacon, Lafayette, CO) and a 100 nM pool of siRNA targeting NF-κB and C/EBPβ, or a 100 nM pool of nontargeting negative control siRNA. After 48 h of incubation at 37°C in 5% CO2, the cells were subjected to further experiments.
Western blot analysis of NF-κB and C/EBPβ
Protein lysates (20 μg) extracted from transfected cells were separated on an 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and electroblotted on polyvinylidene fluoride (PVDF) membranes (Immobilon-P; Millipore, Billerica, MA). After blocking, the blots were incubated with antibodies overnight. Primary antibodies used for this analysis were phospho-RelA (Cell Signaling, Beverly, MA), anti-C/EBPβ antibody (Cell Signaling), and anti-β-actin. Membranes were then incubated for 1 h with secondary antibody. Detection was performed by ECL (Amersham) and by chemiluminescence using Kodak X-Omat film.
Statistical analyses
Experimental data were analyzed using SPPS software (SPSS Predictive Analytics, Chicago, IL). Repeated measures and one-way analyses of variance were applied to assess significant differences and correlations. The Spearman's test was used for correlation analysis. Statistical significance was accepted at the 0.05.
Results
Synovial fluid IL-1β, TNF-α, and IL-6 protein abundance in OA patients
The demographic characteristics of all patients are shown in Table 1. Synovial fluid was collected on the day of surgery from 40 OA patients. The average synovial fluid IL-1β level in OA patients was 0.6±0.4 pg/mL (range: 0.06–1.6 pg/mL). The average synovial fluid TNF-α level in OA patients was 1.6±1.4 pg/mL (range: 0.2–6.01 pg/mL). The average synovial fluid IL-6 level in OA patients was 96.04±147.9 pg/mL (range: 0.2–482.8 pg/mL) (Fig. 1A).
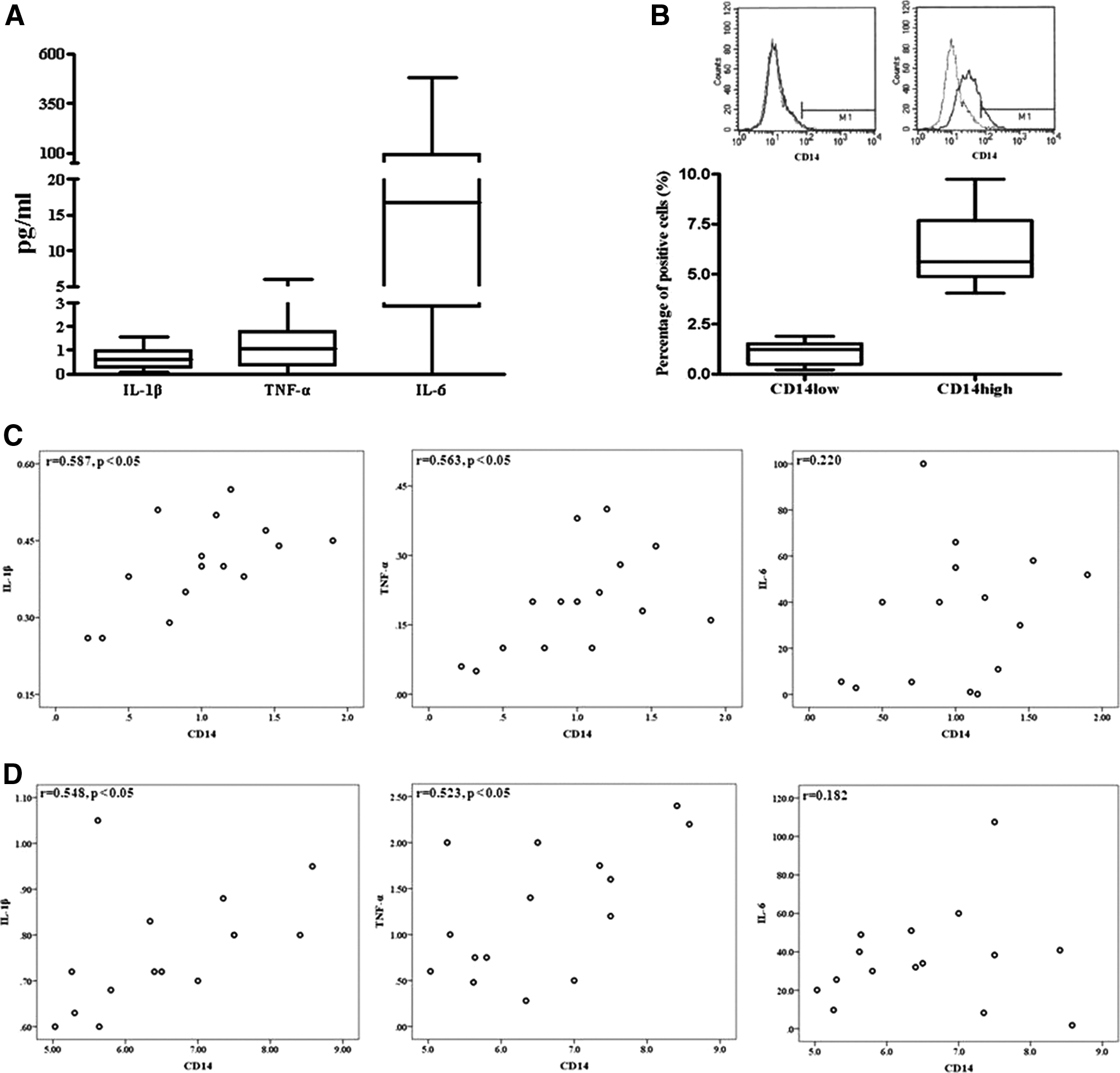
Levels of interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), IL-6, and CD14 in synovial fluid and correlation of IL-1β and TNF-α levels with CD14 levels of synovium-derived stem cells (SDSCs) in osteoarthritis patients.
Percentage of the CD14 marker in osteoarthritic synovium-derived cells
CD14 is a membrane antigen that is predominantly expressed on macrophages and monocytes. Phenotypic analysis of the expression of CD14 markers was conducted by flow cytometry in osteoarthritic SDSCs. The average percentage of CD14 in osteoarthritic SDSCs was 3.6%±2.5% (range: 0.2–8.6%) (Fig. 1B). There was a statistically significant correlation between the IL-1β level of synovial fluid and the percentage of CD14low cells (r=0.548, p<0.05) and CD14high cells (r=0.587, p<0.05). Similarly, a statistically significant correlation was found between the TNF-α level of synovial fluid and the percentage of CD14low cells (r=0.563, p<0.05) and CD14high cells (r=0.523, p<0.05) in 40 OA patients. There was not a statistically significant correlation between the IL-6 level of synovial fluid and the percentage of CD14low cells and CD14high cells (Fig. 1C, D).
Comparable expression of MSC-specific surface markers between whole cells and CD14-negative cells
Whole cells and CD14-negative SDSCs were characterized according to the expression levels of surface markers in BM-derived MSCs or SDSCs. Flow cytometric analysis showed that the expression (<1%) of CD14, CD31, and CD34 was slightly lower in CD14-negative cells than in the whole cells of CD14low cell group and CD14high cell group, whereas those (>99%) of CD44, CD90, and CD105 was unchanged between the whole cells and CD14-negative cells of CD14low cell group and CD14high cell group. The positive and negative levels of stem cell surface markers were unchanged between whole cells and CD14-negative cells of SDSCs (Fig. 2).

Flow cytometric analysis of whole cells and CD14-negative SDSCs. Cell surface marker expression of whole cells of CD14high SDSCs
Chondrogenic potential of osteoarthritic SDSCs by depletion of CD14-positive synovial macrophages
After 3 weeks of culture, pellets (n=10) were examined for expression of type I, II, and X collagen; Sox9; and aggrecan, which are known to be related to chondrogenic differentiation. Expression levels of type II and X collagen were higher in CD14-negative cell pellets compared with whole cell pellets in CD14high cells (Fig. 3A). The expression levels of chondrogenic-related genes were unchanged between whole cells and CD14-negative cells in CD14low cells (Fig. 3A).
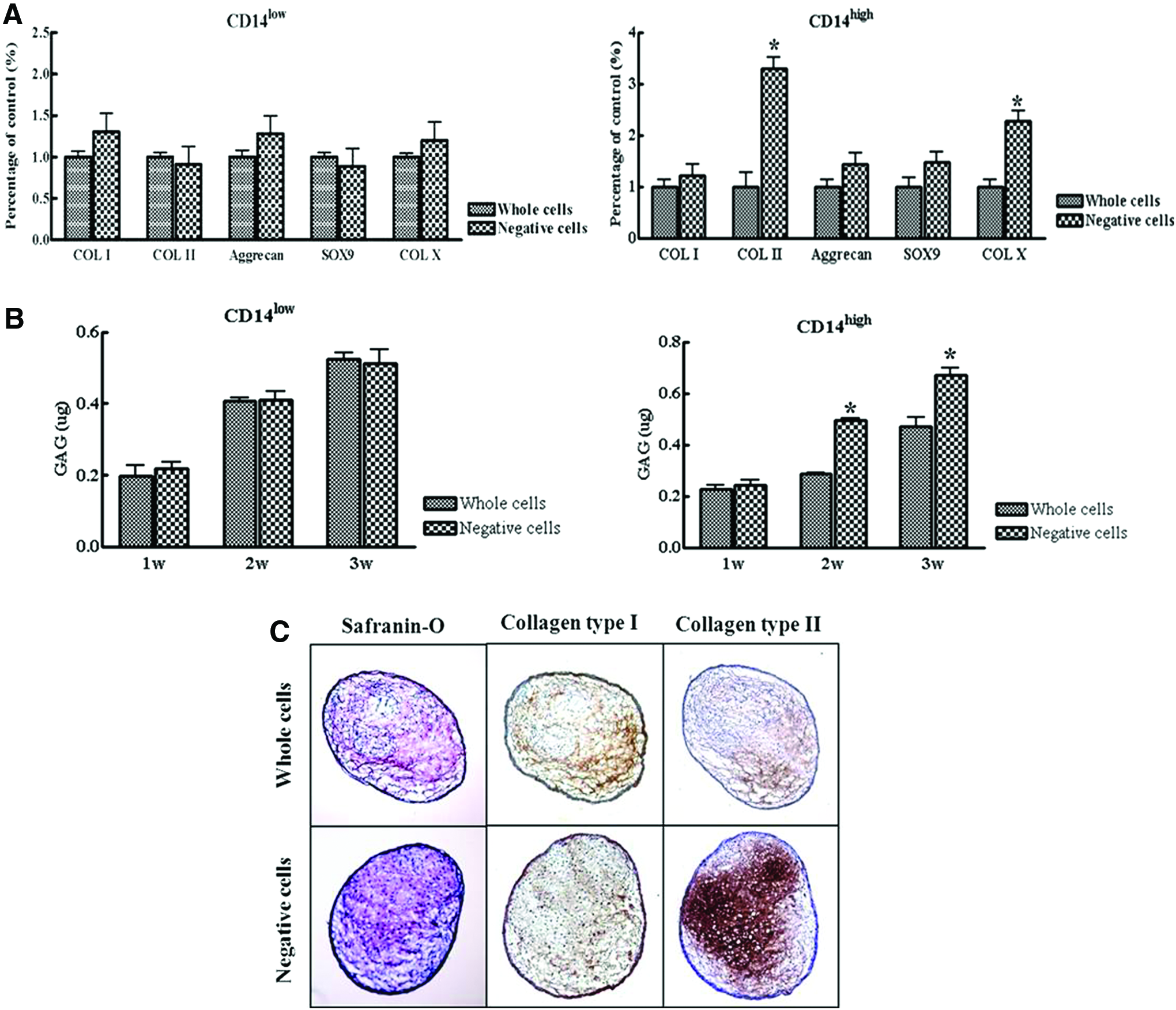
Effects of CD14-negative SDSCs on chondrogenesis.
In terms of GAG synthesis by CD14-depleted cells cultured in pellets, CD14-negative cell pellets exhibited significantly greater GAG synthesis than whole cell pellets in CD14high cell group. No significant differences were observed between whole cells and CD14-negative cells in CD14low cell group (Fig. 3B). Safranin-O staining for proteoglycans and immunohistochemical staining for type II collagen indicated higher levels of type II collagen in CD14-negative cell pellets from CD14high cells than in whole cell pellets from CD14high cells, and that CD14-negative cell pellets from CD14high cells had well-formed matrices (Fig. 3C).
Suppression of the chondrogenic differentiation by IL-1β and TNF-α
The CD14-positive cells significantly inhibited lipopolysaccharide-stimulated release of IL-1β and TNF-α in the medium of SDSCs (Fig. 4A). The effects of IL-1β and TNF-α on chondrogenic gene expression in cell pellets were evaluated by quantitative real-time PCR. As expected, IL-1β and TNF-α significantly modulated chondrogenic gene expression in SDSCs (Fig. 4B). Expression levels of CREBBP and p300, a coactivator of Sox9, were decreased by IL-1β, while C/EBPβ expression was increased by IL-1β (Fig. 4C). Immunofluorescence of type II collagen showed that IL-1β significantly inhibited type II collagen levels (Fig. 4D). Safranin-O staining for proteoglycans showed that IL-1β suppressed uniformity and intensity of matrix staining (Fig. 4E).
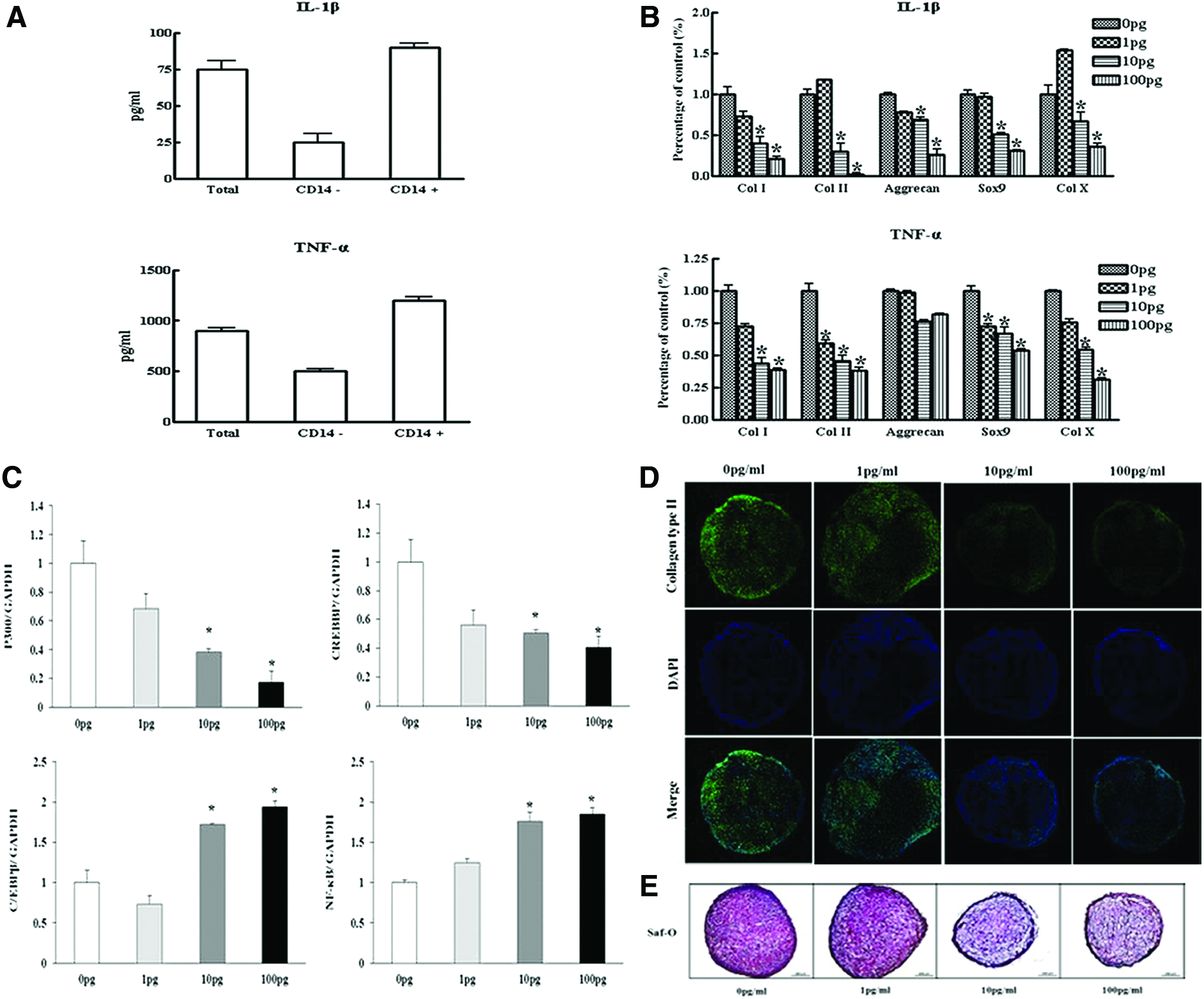
Effects of IL-1β or TNF-α on chondrogenesis.
Reversal of the IL-1β-induced alteration of chondrogenic potential by NF-κB or C/EBPβ knockdown
Expression levels and protein levels of NF-κB and C/EBPβ were effectively reduced using siRNAs targeting NF-κB and C/EBPβ, respectively (Fig. 5A, B). NF-κB siRNAs reduced the expression of C/EBPβ in SDSCs and, conversely, siRNAs targeting C/EBPβ reduced the expression of NF-κB in SDSCs (Fig. 5A). Exposure of cells to NF-κB or C/EBPβ siRNAs reversed the IL-1β-induced suppression of chondrogenic gene expression in SDSC pellets (Fig. 5C). Immunofluorescence analysis of type II collagen showed that NF-κB and C/EBPβ siRNAs enhanced the IL-1β-induced reduction of type II collagen levels (Fig. 5D). These data provide evidence of the functional role of NF-κB and C/EBPβ in IL-1β-induced suppression of chondrogenic potential.
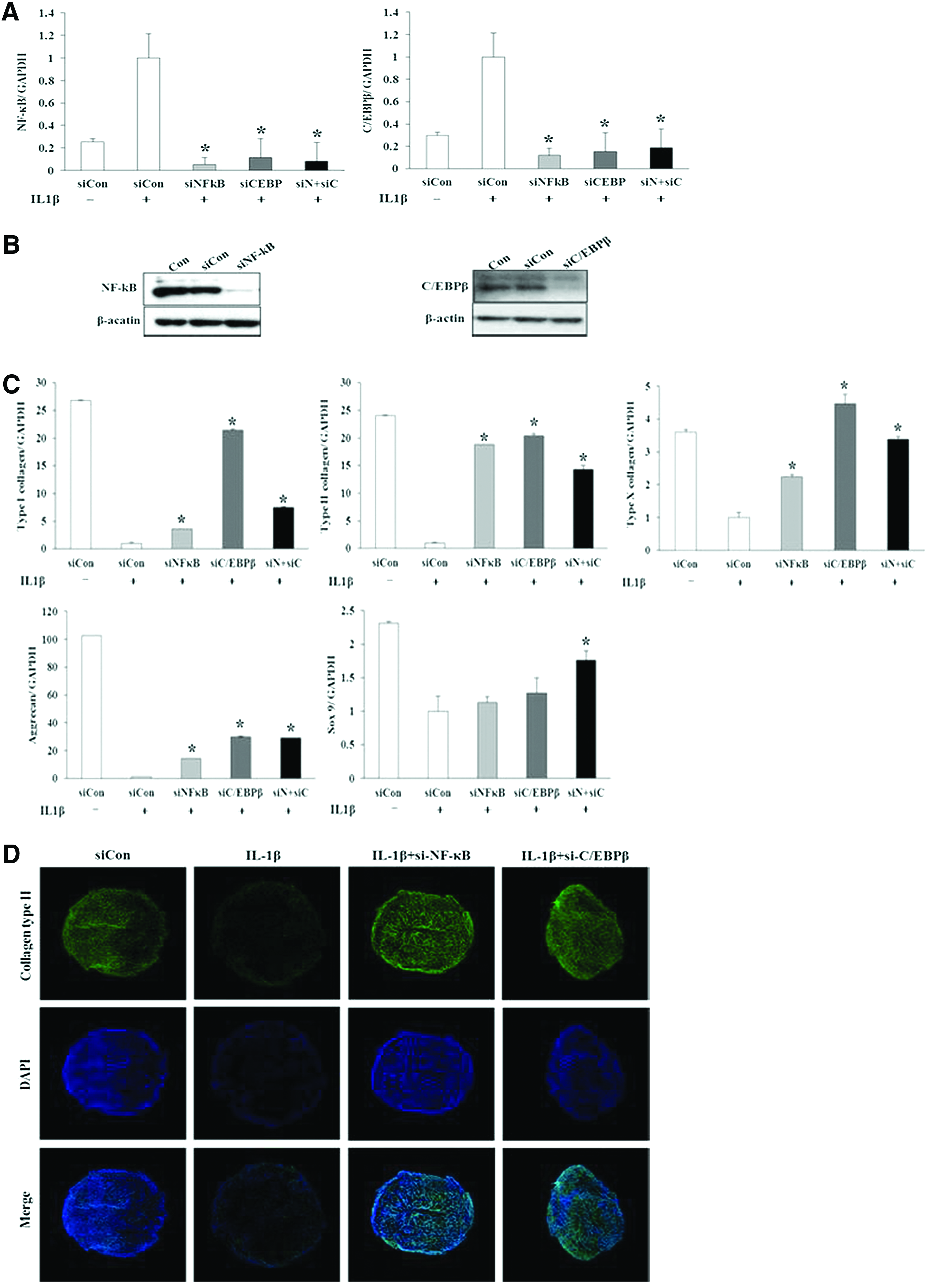
Effects of NF-κB and C/EBPβ small interfering RNAs (siRNAs) on chondrogenesis. Real-time PCR
Discussion
We have demonstrated that depletion of CD14+ synovial macrophages enhances the chondrogenic potential of CD14high osteoarthritic SDSCs, indicating that synovial macrophages interfere with the chondrogenic capacity of osteoarthritic SDSCs. Moreover, we have shown that NF-κB (RelA/p65) and C/EBPβ are pivotal factors mediating IL-1β-induced suppression of the chondrogenic potential of osteoarthritic SDSCs.
OA is a degenerative joint disease characterized by cartilage breakdown, the formation of osteophytes, subchondral bone sclerosis, alterations to the joint capsule, and inflammation of the synovial membrane. 32 The inflammation of the synovium characteristic of both the early and late phases of OA results in synovitis, and is associated with alterations in the adjacent cartilage that are similar to those seen in RA. 33 Since SDSCs are used to facilitate cartilage tissue regeneration, it is critical to consider the effect of synovitis on chondrogenic differentiation of OA SDSCs since inflammatory cytokines have previously been shown to inhibit chondrogenesis.27,28
CD14 is a glycoprotein expressed on the surface of myelomonocytic cells as a GPI-anchored receptor or secreted in a soluble form. 34 We observed a correlation between IL-1β and TNF-α levels of synovial fluid and CD14 levels in osteoarthritic SDSCs. Bondeson et al. reported that depletion of CD14+ synovial macrophages results in decline of IL-1β and TNF-α, which may in turn inhibit the release of IL-6, chemokines, and MMPs in OA synovial cells. 15 This would indicate that CD14 plays a role in inflammatory and destructive responses from the synovial fibroblasts. Soluble CD14 is increased in synovial fluid in both early and advanced stages of human knee OA, and the synovial fluid of early OA patients has been shown to modulate inflammatory responses of synoviocytes to TLR2 and TLR4 ligands through soluble CD14. 35
IL-1β and TNF-α are considered to be major players in OA pathogenesis through suppression of matrix synthesis and promotion of cartilage catabolism. 36 We examined whether low concentrations of IL-1β and TNF-α would influence the chondrogenic differentiation potential. Studies have shown that synovial fluid levels of IL-1β and TNF-α vary widely in osteoarthritic patients, from 4.8 pg/mL 37 to 27.8 pg/mL 38 for IL-1β, and from 33.2 pg/mL 39 to 0.5 ng/mL 38 for TNF-α. Our results indicate that 10 pg/mL of IL-1β and 1 pg/mL of TNF-α are sufficient to modulate the expression of chondrogenic-related genes, suggesting that IL-1β and TNF-α in osteoarthritic synovial fluid might influence the chondrogenic differentiation potential.
While the superiority of SDSCs for cartilage formation has been demonstrated in multiple studies,4–8 the cytokine-induced inflammation of chondrogenic potential in SDSCs among other arthritides is still largely unknown. The chondrogenic potential of SDSCs in rheumatoid synovium has been shown to be affected by synovial inflammation and by the proportion of infiltrating monocytes, and cytokine production and Sox9 expression are comparable in RA-MSCs and OA-MSCs. 40 Moreover, Pei et al. have shown that a synovial fibroblast-enriched synoviocyte cell population undergoes more extensive chondrogenesis than a mixed cell population of synoviocytes. 16 Similarly, we have shown here that depletion of CD14+ synovial macrophages leads to improved chondrogenic potential in CD14high osteoarthritic SDSCs, reflected in type II collagen and cartilage matrix formation.
Our finding indicates that NF-κB and C/EBPβ are the major transcription factors mediating cytokine-induced inflammatory responses in chondrogenic differentiation in SDSCs. Specifically, transfection of NF-κB- and C/EBPβ-specific siRNAs enhanced the chondrogenic potential of human SDSCs. NF-κBp65-specific siRNA has been previously used in a variety of systems, such as vascular cells, 40 chondrocytes, 20 and an OA model, 41 and has been shown to inhibit the expression of MMP-9, NOS-2, and COX-2 in IL-1β-induced chondrocytes. 20 Consistent with these data, inhibition of NF-κB activation by overexpression of an IκB repressor improved chondrogenesis in human BM MSCs. 28 Moreover, IL-1β modulates levels of type II collagen, the expression of which has been shown to be regulated during ATDC5 differentiation by a balance between two transcription factors: the positive regulator Sox9 and the negative regulator C/EBPβ. 42 CREBBP and p300 interact independently with Sox9 and C/EBP, and function by distinct, but complementary, mechanisms to increase the expression of cartilage-related genes. 43 An improved understanding of chondrogenic differentiation will likely result from studies of regulation of chondrogenic gene expression by transcription factors and their interaction with coregulators.
We demonstrated that NF-κBp65-specific siRNA reduced the expression of C/EBPβ, and conversely, C/EBPβ-specific siRNA decreased the expression of NF-κB in SDSCs. This finding demonstrates that NF-κB and C/EBPβ mutually regulate IL-1β-induced suppression of the chondrogenic potential of SDSCs. Zhang et al. have demonstrated a functional relationship between these two transcription factors in chondrocytes in response to IL-1β, 44 and coregulation by NF-κB and C/EBPβ of genes in a variety of other cell types has been demonstrated.45–47 These data are consistent with the ability of NF-κB and C/EBPβ to block the expression of Sox9, 48 a transcription factor that is essential for chondrogenesis, and for repression of type II collagen expression. 49 Moreover, evidence suggests that C/EBPβ enhances NF-κB-associated signaling by negatively regulating the level of the NF-κB inhibitor IkB-α. 50
In conclusion, our study suggests that depletion of CD14+ synovial macrophages leads to improved chondrogenic potential in the CD14high osteoarthritic SDSCs, and that NF-κB (RelA) and C/EBPβ are critical factors mediating IL-1β-induced suppression of the chondrogenic potential of human SDSCs.
Footnotes
Acknowledgment
This study was supported by a grant of the Korea Healthcare technology R&D Project, Ministry for Health, Welfare & Family Affairs, South Korea (A100451).
Disclosure Statement
No competing financial interests exist.