
Abstract
The bone morphogenetic proteins (BMPs) are potent osteogenic molecules that are used for bone repair in delivery systems and in regenerative medicine. We studied the responses of murine MC3T3-E1 preosteoblasts to doses of recombinant human (rh)BMP-9 with and without fetal bovine serum (FBS). rhBMP-2 was used as a control since it is currently approved by the Food and Drug Administration for bone application. We analyzed the major cell signaling pathways and the expression of osteogenic markers. Without FBS, BMP-9 had a similar effect on MC3T3-E1 preosteoblast differentiation in comparison to BMP-2. In contrast, FBS reduced the EC50 of BMP-9 fourfold to sixfold, as determined by osterix gene expression and alkaline phosphatase (ALP) activity, while it had no influence on EC50 of BMP-2. As suggested by MAPK inhibitor assays, FBS could induce an intracellular signaling environment that favors cell response to BMP-9 by inhibiting ERK1/2 activation and increasing p38 phosphorylation. Finally, IGF-2 (100 ng/mL) could mimic the effect of FBS on BMP-9 cell response in terms of MAPK signaling and ALP activity. Thus, the action of BMP-9 on preosteoblast differentiation can be greatly improved by IGF-2. This finding may well be critical for developing optimal growth factor delivery systems and bone tissue engineering strategies.
Introduction
B
Large bone defects that cannot self-repair are among the greatest challenges. Several strategies have been investigated over the past two decades to overcome the drawbacks of autografts such as the risks of infection and morbidity at both the donor and receiver sites.2,3 Of these, growth factor (GF) delivery systems and tissue engineering have attracted broad interest. GF delivery systems are usually composed of a scaffold and a carrier that delivers osteogenic GFs such as the bone morphogenetic proteins (BMPs). They are designed to trigger and sustain bone healing directly at the defect site. 4 Tissue engineering uses synthetic highly porous bone grafts containing bone-forming cells (usually from the patient). These are expanded in sterile reactors to create bioengineered bone tissue that can then be transplanted to the defect site. 5 This requires a complex mixture of mitogenic elements, vitamins, and other GFs. These factors are usually provided by serum such as fetal bovine serum (FBS).6,7 However, the use of FBS and other animal-derived sera, which could contain harmful animal-derived elements, is highly restricted and regulated by agencies such as the Food and Drug Administration (FDA) for tissue engineering applications. 8 It is therefore important to mimic the serum environment as closely as possible and use more defined media. The most important mitogenic GFs in FBS are IGF-1 and IGF-2. 9 IGF-2 could be important for bone regeneration as it can promote osteogenic differentiation.10,11
BMPs are also promising candidates for delivery systems and tissue engineering strategies. These homodimeric proteins are potent osteoinducers that are involved in natural endochondral bone healing. 12 They were first identified by Urist and Strates 13 in the early 1970s and more than 20 BMPs are presently known. 14 The BMPs become bound to the serine/threonine kinase membrane receptors (type I and type II) on cells through their knuckle and wrist epitopes.14–16 BMP-receptor interactions trigger the phosphorylation of Smad 1/5/8 by type I receptors. The phosphorylated Smad 1/5/8 form complexes with Co-Smad that are translocated to the nucleus, where they initiate and regulate the expression of the genes encoding osteogenic markers. 17 MAPK, especially p38 and ERK1/2, and Wnt pathways are also involved in BMP signaling.18,19 Several of the 20 BMPs (BMP-2, BMP-4, BMP-6, BMP-7, and BMP-9) are potent stimulators of mesenchymal stem cell osteogenic differentiation.20–22 BMP-2 and BMP-7 have also been approved by the FDA for use in bone repair applications such as spinal fusion and the treatment of tibia bone shaft fractures.4,14,22
However, Kang et al. 21 used adenovirus encoding BMP (AdBMP) to show that BMP-9 has a greater osteogenic effect than BMP-2. BMP-9 is unlike all the other BMPs. It is mostly synthesized in the liver and its concentration in the blood is about 2–12 ng/mL.23–25 BMP-9 is also found in platelets, possibly in their granules. 26 This versatile protein could be the most important in GF delivery systems and tissue engineering, but little is known about its mechanism of action or its role in bone healing.
We therefore established the dose–response curves for the in vitro effects of BMP-9 on murine preosteoblast cells (MC3T3-E1 subclone 14), since it is a well-known cell model that behaves like human preosteoblasts in terms of their differentiation and osteogenic marker profiles. 27 BMP-2 was used as a control. We evaluated the capacity of MC3T3-E1 preosteoblasts to undergo osteoblastic differentiation in the presence of BMP-9 by measuring the activation of signaling pathways and the synthesis of late osteogenic markers with and without FBS. We used specific inhibitors to verify the involvement of MAPK signaling in the responses to BMP-9. Since FBS had an enormous positive effect on BMP-9-stimulated MC3T3-E1 preosteoblast differentiation, we then investigated the impact of IGF-2 on the responses of cells to BMP-9 in a controlled environment and found interesting behaviors that have not been shown yet with preosteoblasts.
Materials and Methods
Materials
Recombinant carrier-free human BMP-2 and BMP-9 (from Chinese hamster ovarian cells) were purchased from R&D Systems (Minneapolis, MN). MEK1/2 (PD98059), p38 (SB202190), and JNK (SP600125) inhibitors were purchased from Millipore EMD (Calbiochem® Inhibitors & Research Essentials Biochemicals; EMD Millipore, Billerica, MA). Lyophilized IGF-2 was recovered as specified by the manufacturer's instructions (PeproTech, Oak Park, CA). Rabbit primary antibodies directed against ERK1/2 (p44/42 MAPK ERK1/2 137F5), p38 (p38 MAPK), or their phosphorylated forms pERK1/2 (pp44/42 MAPK T202/Y204) and pp38 (pp38 MAPK T180/Y182) were purchased from Cell Signaling Technologies (Danvers, MA). The rabbit primary antibodies against the phosphorylated form of Smad 1/5/8 (pSmad 1/5S463/465/8S426/428) were also purchased from Cell Signaling Technologies, while those against Smad 1/5/8 (Smad 1/5/8 (N-18)-12 IgG) and the phosphorylated form of JNK (pJNKG-7) were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). Finally, secondary antibodies (anti-rabbit IgG) coupled to horseradish peroxidase (ECL™ horseradish peroxidase-linked anti-rabbit IgG, whole antibody from donkey) were purchased from GE Healthcare (Buckinghamshire, United Kingdom).
Cell culture
Murine calvarial MC3T3-E1 preosteoblast subclone 14 (CRL-2594™; ATCC®, Manassas, VA) was used between passage 4 and 11 and seeded at 5000–10,000 cells/cm2 onto tissue culture-treated polystyrene flasks. Cells were grown at 37°C (humidified atmosphere containing 5% CO2) in the minimum essential medium-alpha without ascorbic acid (Gibco®, Grand Island, NY) supplemented with 10% (v/v) heat-inactivated FBS (Wisent Bioproduct, St-Bruno, Canada). Cells at 80% confluence were starved of FBS for 1 day to synchronize the cell cycle before stimulation with GFs. The same lot of FBS was used for the entire study.
Western blotting and densitometry
MC3T3-E1 preosteoblasts were seeded and cultured as described above. They were then incubated with BMP-2 or BMP-9 (0, 0.1, 0.2, 0.4, 1, 4, 8 nM) with or without 10% (v/v) FBS for 1 h. Cells used for MAPK inhibitor assays were first incubated with 20 μM of MEK1/2 inhibitor (PD98059; PD), p38 inhibitor (SB202190; SB), or JNK inhibitor (SP600125; SP) for 1 h before stimulation with 1 nM BMP-2 or BMP-9 with or without 10% (v/v) FBS. Western blot analyses were performed as previously described. 28 Revealed bands were scanned using the FluorChem™ 5500 (Alpha Innotech, San Leandron, CA) imaging system and the densitometry of the bands was analyzed using the image analysis software coupled to the imaging system (AlphaEase® v4.0 Image Analysis software). The integrated densities of the phosphorylated forms of signaling proteins (pSmad 1/5/8, pERK, pp38, and JNK) were normalized to their overall amount (Smad 1/5/8, Total ERK, p38, and actin) for each experiment.
Gene expressions
We measured the mRNA levels encoding for osterix (Osx) and osteocalcin (OC) in cells that had been incubated for 1 or 6 days with BMP-2 or BMP-9 (0, 0.04, 0.1, 0.2, 0.4, 1, 4, 8 nM) with or without 10% (v/v) FBS. The cells used to measure Bglap (OC) were stimulated twice on day 0 with BMP-9 or BMP-2 (1 nM), with or without 10% (v/v) FBS and on day 3, for a total of 6 days of stimulation. The mRNA was then extracted using the TRIzol® reagent (Invitrogen, Carlsbad, CA) following the manufacturer's instructions and precipitated with isopropanol (Fisher, Mississauga, Ontario, Canada). The precipitated mRNA samples were centrifuged (24,550 g), washed twice with ethyl alcohol (75% v/v; Fisher), and partially dried in a vacuum centrifuge (DNA120 SpeedVac concentrator; Thermo Electron Corporation, Madison, WI). Finally, the RNA concentration was quantified spectrophotometrically at 260/280 nm using GeneQuant pro (Biochrom, Cambridge, United Kingdom). Aliquots of mRNA (1 μg) were treated with DNAse I (Invitrogen) and used to synthesize first-strand cDNA using Oligo(DT) primers and Superscript™ Reverse transcriptase II (Invitrogen). Quantitative polymerase chain reactions (qPCR) were performed as previously described using specific primers that amplify gene encoding for murine Osx (Mm_Sp7_1_SG, 69 bp, QuantiTec Primers Assays), for murine OC (Mm_Bglap_1_SG, 102 bp, QuantiTec Primers Assays), and for the reference gene Gapdh (Mm_Gapdh_3_SG, 144 bp, QuantiTec Primers Assays). 28
Alkaline phosphatase activity
MC3T3-E1 preosteoblasts were seeded and cultured as described above. Cells used for dose–response experiments were stimulated for 72 h with BMP-2 or BMP-9 (0, 0.04, 0.1, 0.2, 0.4, 1, 2, 4, 8 nM) with or without 10% (v/v) FBS. Cells to be treated with specific inhibitors of MAPK MEK1/2 (PD98059; PD), JNK (SP600125; SP), or p38 (SB202190; SB) were first incubated with the inhibitor (20 μM) for 1 h and then stimulated with BMP-9 or BMP-2 (1 nM) for 3 days with or without FBS (10% v/v). MC3T3-E1 preosteoblasts were also stimulated with IGF-2 (100 ng/mL) for 3 days and with IGF-2-depleted FBS.
Alkaline phosphatase (ALP) activity assay (SensoLyte® pNPP Alkaline Phosphatase Assay Kit; Anaspec, San Jose, CA) was performed according to the manufacturer's instructions. The hydrolysis of the substrate para-nitrophenyl phosphate (pNPP) was monitored by measuring the absorbance at 405 nm using a microplate reader Synergy TM HT (Biotek, Winnosli, VT). The ALP activity was then evaluated from a standard curve (0–100 ng/mL pNPP) and the concentrations normalized to the total number of cells used for each experimental condition using an image analysis program developed with MatLab software R2007b V. 7.5.0.342 (MathWorks, Natick, MA). Briefly, three to four representatives pictures were taken per well before cell lysis (∼0.02 cm2/picture) using an Eclipse TE200-S microscope coupled to a 10× objective and a CCD camera (Retiga 1300R; Nikon, Mississauga, Canada) to estimate the total number of cells (9.62 cm2/condition). Finally, the ALP activities for each independent experiment were normalized to their control.
Dose–response modeling and determination of effective doses
The dose–response relationships for pSmad 1/5/8 relative density, Osx gene expressions, and ALP activity (y) were modeled with respect to the concentrations of BMPs ([BMP]) as a sigmoid function (Eq. 1). Dose–response profiles usually have three steps: (i) basal activation/expression, (ii) exponential increase when the concentration exceeds the threshold, and (iii) a plateau when the number of BMP molecules is far greater than the number of cell membrane receptors (BMPRI and BMPRII).
The model parameters (ymax, B, and C) were estimated using the Levenberg-Marquardt algorithm programmed with Matlab software R2007b V. 7.5.0.342 (MathWorks). This optimization algorithm is based on the minimization of the sum of square between the experimental and model data. The effective concentration (EC50) is defined as the concentration at which the response is half of the maximal possible response (
Depletion of IGF-2 in FBS
FBS was depleted of IGF-2 by a modified immunoprecipitation method. Briefly, FBS samples were incubated with protein G-agarose microspheres (Protein G Immunoprecipitation Kit; Sigma, St. Louis, MO) overnight at 4°C under agitation to remove proteins that could be nonspecifically adsorbed. Samples were then centrifuged at 12,000 g. The resulting supernatant was collected, mixed with a fresh solution containing protein G-agarose microspheres and 20 μg/mL IGF-2 polyclonal antibodies (MJS BioLynxinc, Ontario, Canada), and incubated at 4°C for 120 min with agitation. The protein G-agarose microsphere-antibody-IGF-2 complex was removed by centrifugation at 12,000 g for 15–30 s and the resulting low-IGF-2 supernatant was collected and store under sterile conditions. The microspheres were collected and washed with sterile PBS (two to three times), mixed with the Laemmli buffer, and the mixture was boiled for 5 min. The efficiency of IGF-2 removal was assessed by western blotting following the same procedure as previously developed using fibronectin as a reference protein.
Statistical analysis
Statistical analyses were performed using Excel (Excel 2010®). Analyses of variance (ANOVA) were performed using the Excel Data Analysis Toolbox with a level of confidence of 95%. Only differences with a p<0.05 were considered significant. Since dose–response experiments frequently show heteroscedasicity,29,30 we used Box-Cox transformations when necessary before ANOVA to obtain uniform variances.
Results and Discussion
Influence of BMP-2 and BMP-9 concentrations on Smad 1/5/8 and MAPK phosphorylation
As BMPs activate the Smad 1/5/8 pathway and MAPK, both of which are involved in osteogenic differentiation,17,18,31,32 we investigated the effects of several doses of BMP-9 on these signaling cascades to identify the mechanisms involved in the responses of MC3T3-E1 preosteoblasts in the presence or absence of FBS. BMP-2 was used as a control since it is currently applied in commercial delivery systems and tissue engineering strategies to favor bone formation.2,3 The phosphorylated states of Smad 1/5/8 and p38, ERK1/2 and JNK in cells incubated with BMP-2 or BMP-9 (0.1–8 nM) with or without 10% (v/v) FBS were analyzed by western blotting (Fig. 1A, B).
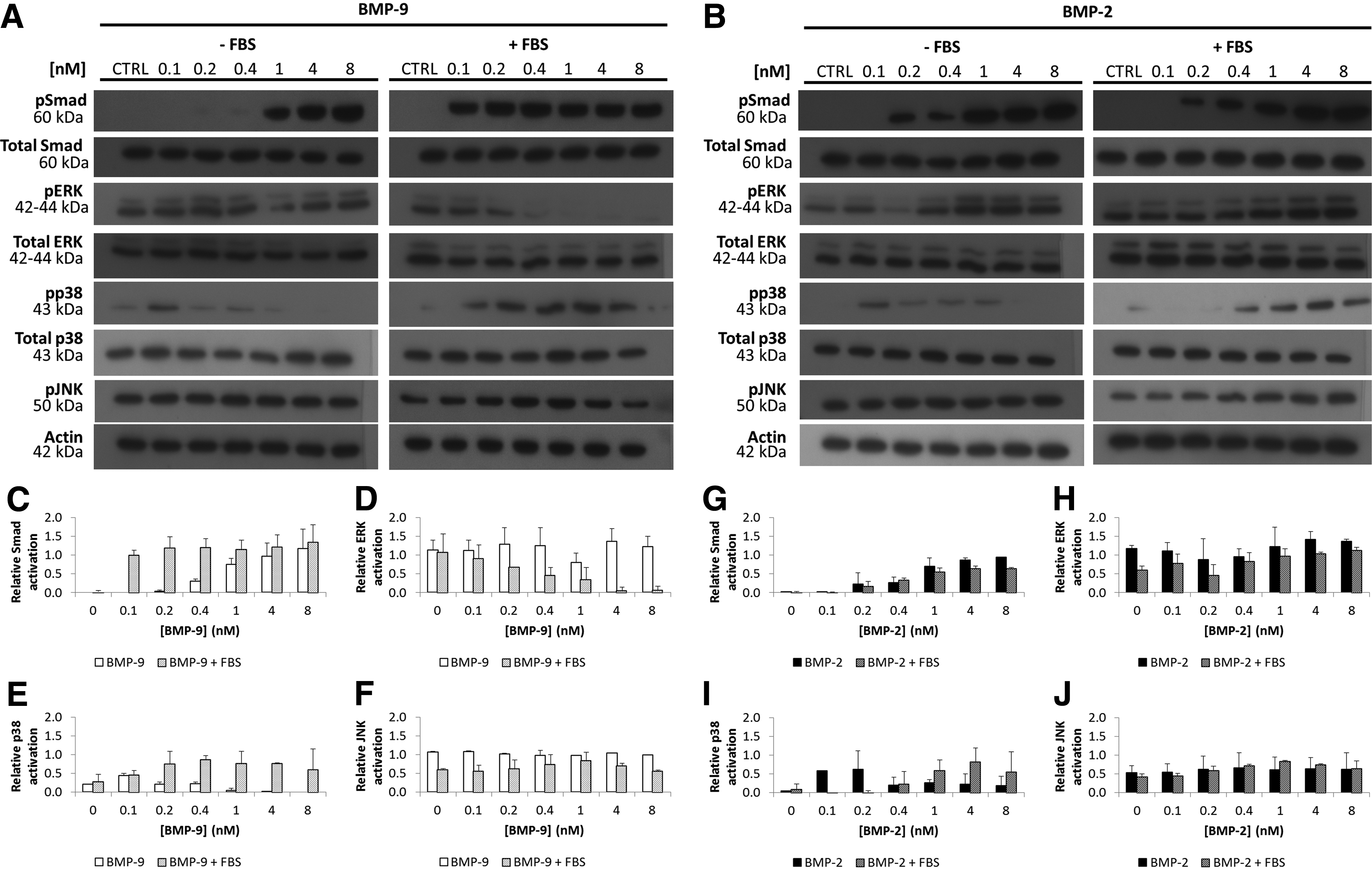
Western blots showing dose-dependent effects of bone morphogenetic proteins (BMPs) on the phosphorylation of Smad 1/5/8, ERK1/2, p38, and JNK in MC3T3-E1 preosteoblasts stimulated for 1 h
The intensity of the bands corresponding to the phosphorylated Smad 1/5/8 in cells stimulated with BMP-2 with and without FBS was not significantly different. Both conditions (+or−FBS) resulted in similar sigmoid-shaped dose–response profiles (Fig. 1G). The phosphorylation of Smad 1/5/8 in cells incubated with various BMP-9 concentrations, but without FBS, was also sigmoid (Fig. 1C). Bergeron et al. 33 previously found that MC3T3-E1 preosteoblasts stimulated without serum behaved similarly in response to 1.92 nM BMP-2 and BMP-9 in terms of Smad 1/5/8 phosphorylation (1 h of stimulation). However, incubation with FBS+BMP-9 produced a different profile; Smad 1/5/8 were activated at a lower concentration (Fig. 1C). Mathematical modeling (Table 1) gave an EC50 for BMP-9 in the presence of FBS on Smad phosphorylation that was only 20% that of cells incubated with BMP-9 alone.
BMP, bone morphogenetic protein; FBS, fetal bovine serum.
There were also noticeable differences in MAPK activation that depended on the type of BMP used and the presence or absence of FBS. The intensity of the bands corresponding to phosphorylated ERK1/2 in cells stimulated with BMP-2 with or without FBS was similar to each other (Fig. 1H) and to those of controls. BMP-9 had a completely different effect in the presence of FBS. The amounts of phosphorylated ERK1/2 in cells stimulated with BMP-9 (0.1–8 nM) without FBS did not change and were similar to the basal level (control), while the phosphorylation of ERK1/2 decreased drastically after incubation with BMP-9+FBS. This was confirmed by the densitometry analysis graphs (Fig. 1D). Furthermore, the phosphorylated p38 in BMP-9- or BMP-2-stimulated cells decreased as the BMP concentration increased with a maximum intensity at 0.1 nM. On the contrary, the presence of FBS produced an increasing dose–response profile with a threshold concentration around 1 nM (25 ng/mL) for BMP-2-stimulated cells. Guicheux et al. 19 also observed that 100 ng/mL BMP-2+1% (v/v) fetal calf serum stimulated the phosphorylation of p38 in MC3T3-E1 preosteoblasts between 1 and 6 h of incubation time. Adding serum also increased and stabilized the phosphorylated state of p38 over the BMP-9 concentration range tested. Finally, FBS had no effect on JNK phosphorylation in the presence of BMP-9 or BMP-2 in comparison to the control.
Osx and OC mRNA responses to BMP-2 and BMP-9 with or without FBS
Activation of the canonical Smad pathway leads to a complex (R-Smad/Co-smad) that moves to the nucleus, where it interacts with promoters that regulate the expression of early osteogenic genes such as Runx2, Dlx5, and Osx. 17 We used qPCR to investigate the effect of several doses of BMP-2 and BMP-9 with or without FBS on the expression of Osx gene in cells incubated for 1 day.
The levels of Osx transcripts in the control with or without FBS were essentially the same, indicating that FBS alone did not trigger the activation of the Osx gene. However, both BMP-2 and BMP-9 stimulated cells showed dose-dependent increases in Osx transcripts in the absence of FBS (Fig. 2A) with a similar sigmoid-shaped profile. This was also confirmed by the similar EC50 for BMP-2 and BMP-9 estimated from the mathematical modeling (Table 2). Bergeron et al. 33 also showed similar Osx mRNA expression levels for MC3T3-E1 preosteoblasts stimulated with BMP-2 and BMP-9 (1.92 nM).

Dose–response effect of BMP (0–8 nM) on osterix (Osx) mRNA.
Osx, osterix.
FBS completely modified the dose-dependent response of cells to BMP-9, but had no great influence on the response to BMP-2 (Fig. 2B, D). The EC50 of BMP-2 with (∼0.5 nM) and without (0.6 nM) FBS was similar (Table 2). BMP-9+FBS strongly stimulated the production of Osx transcripts, even at BMP-9 concentrations as low as 0.1 nM (2.12 ng/mL). The EC50 of BMP-9 when incubated with and without FBS was different (it was about fourfold lower with FBS) (Table 2). Moreover, cells stimulated with BMP-9+FBS contained more mRNA (20 at 8 nM) than cells treated with BMP-9 alone (15 at 8 nM).
As the activation of osteogenic genes by BMP-9 seemed to be strongly promoted by FBS, we evaluated the influence of FBS on the activation of a late osteogenic marker by measuring the concentration of OC mRNA in cells stimulated for 6 days with the BMPs (1 nM) with and without FBS (Fig. 2E). This concentration is slightly above the EC50 of BMPs for activating Smad without FBS and corresponds to a physiologic concentration.24,26
FBS alone slightly increased the production of OC transcripts over that of controls (CTRL, Fig. 2C). This may have been due to cell restimulation as the culture medium was replaced after 3 days. Cells stimulated with BMP-2+FBS had significantly more (twofold) OC transcripts than those stimulated with BMP-2 alone. The serum drastically increased the responses of cells to BMP-9. Cells incubated with BMP-9+FBS had six times more OC mRNA than cells incubated with BMP-9 alone.
Alinejad et al. 34 recently reported that incubating MC3T3-E1 preosteoblasts (subclone 14) with BMP-9 in combination with single-wall carbon nanotubes and 10% (v/v) FBS stimulated osteogenic differentiation more than did BMP-2 (2 nM), as assessed by the expressions of the Osx and OC genes and ALP activity. Kang et al. 21 also found that, when combined with 10% (v/v) fetal calf serum, AdBMP-9 was more osteoinductive than AdBMP-2 for C2C12 cells, but the concentrations of the synthesized BMPs were not given. For example, Li et al. 35 showed that C2C12 cells infected with the same PFU density produced significantly different amounts of BMP-2, -4, -6, -7, and -9. The concentration of BMP-9 was six times more compared with BMP-2. 35
BMP-2 and BMP-9 dose–response curves for ALP activity with and without FBS
The concentration of ALP increases as the cells undergo osteogenic differentiation, since it is essential for bone mineralization. 36 We therefore evaluated the effects of stimulating MC3T3-E1 preosteoblasts for 3 days with BMP-2 or BMP-9 with and without FBS on their ALP activity.
Adding FBS to the cell culture (0, Fig. 3B) did not significantly increase the ALP activity over that of the control without FBS (CTRL, Fig. 3B) (p>0.05). Cells stimulated with both BMP-2 and BMP-9 without FBS gave sigmoid-shaped profiles with a plateau at 2–4 nM.
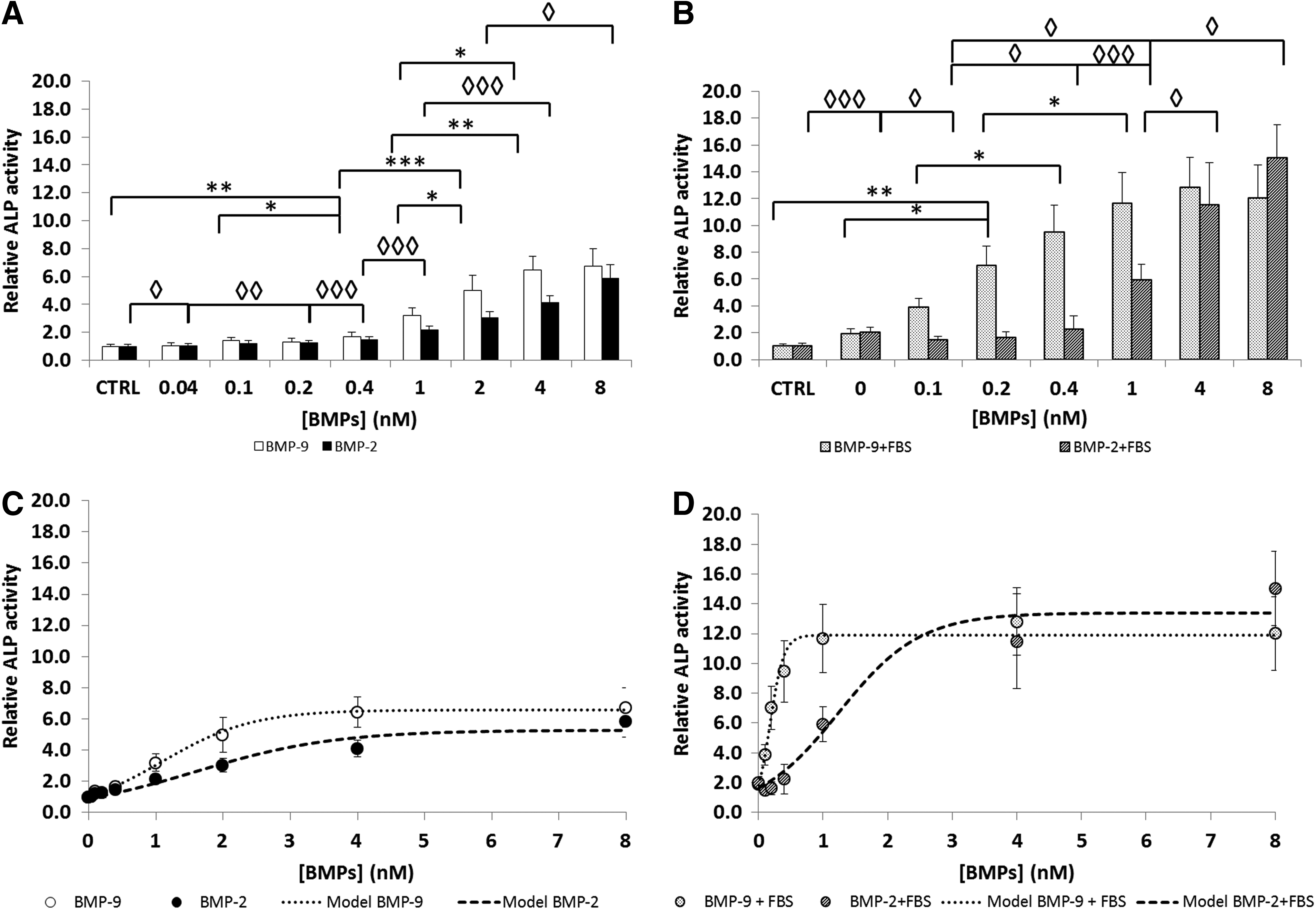
Alkaline phosphatase (ALP) activity in MC3T3-E1 preosteoblasts stimulated with BMP-9 and BMP-2 (0–8 nM)
However, in the presence of FBS, the ALP activity induced by BMP-2 was increased twofold in comparison to BMP-2 alone, but the EC50 of BMP-2+FBS was not significantly different. In contrast, adding FBS strongly affected the dose–response profile of BMP-9 (Fig. 3B), decreasing the EC50 for BMP-9 sixfold (Table 3) and increasing twofold the ALP activity response to high BMP-9 concentrations (1, 4, and 8 nM). Other studies using mouse myoblasts (C2C12) 37 and mesenchymal stem C3H10T1/2 cells 38 have found for ALP activity higher BMP-2 EC50 of 120 ng/mL and 2.8 nM, respectively. This difference reflects the fact that various cell types may not respond exactly the same to BMPs due to the amount and the availability of receptors.
ALP, alkaline phosphatase.
Since MC3T3-E1 preosteoblasts can produce BMP-4 early in their differentiation and this could act as an autocrine and/or paracrine stimulus, 39 we verified that MC3T3-E1 preosteoblasts did not contain BMP-2 or BMP-4 transcripts after 1 day of stimulation with BMP-2 or BMP-9 (1 nM) (data not shown). In addition, the combined concentration of BMP-4, BMP-6, and BMP-9 in FBS is generally about 12–25 ng/mL, with BMP-9 accounting for ∼50%. 24 The BMP concentrations should be about 1.2–2.5 ng/mL (0.06–0.12 nM) in our culture medium since we used 10% (v/v) FBS. The potent effect of FBS on BMP-9 might not be due to circulating BMPs because cells stimulated with FBS alone showed no Smad activation at 1 h, no difference in terms of MAPK signaling compared with control (without FBS), no osteogenic gene expression at 1 day or ALP activity at 3 days, and FBS had also no significant effect on the EC50 of BMP-2.
Effect of MEK1/2 and JNK inhibitors on signaling pathways and ALP activity
As the MAPK pathway seemed to play an important role in the transduction of BMP signals, 40 we investigated the effect of the inhibitors PD98059 (MEK1/2), SP600125 (JNK), and SB202190 (p38) on the activation of the Smad pathway and ALP activity in MC3T3-E1 preosteoblasts stimulated with BMP-2 or BMP-9 with or without 10% (v/v) FBS.
We first verified the ability of each MAPK inhibitor (PD98059, SB202190, and SP600125) to prevent the phosphorylation of its target protein in cells pretreated for 1 h and then stimulated by BMP-9 or BMP-2 (1 nM) (Fig. 4A, B). Inhibiting MEK1/2 had no effect on Smad 1/5/8 phosphorylation in cells stimulated with BMP-9 or BMP-2 without FBS. In contrast, adding FBS produced a net increase in Smad 1/5/8 phosphorylation in MC3T3-E1 preosteoblasts pretreated with the MEK1/2 inhibitor and stimulated with BMP-9. BMP-2+FBS had no such effect. The JNK inhibitor (SP600125) seemed to slightly increase the phosphorylation of Smad 1/5/8 in response to stimulation with BMP-9+FBS. This suggests that inhibiting MEK1/2 and JNK increases the activation of the Smad pathway induced by BMP-9+FBS. In contrast, the p38 inhibitor (SB202190) had the opposite effect on phosphorylated Smad 1/5/8 induced by BMP-9. Unlike BMP-2, cells pretreated with SB202190 and then stimulated with BMP-9+FBS had very much less phosphorylated Smad 1/5/8 than BMP-9 alone without SB202190 pretreatment.
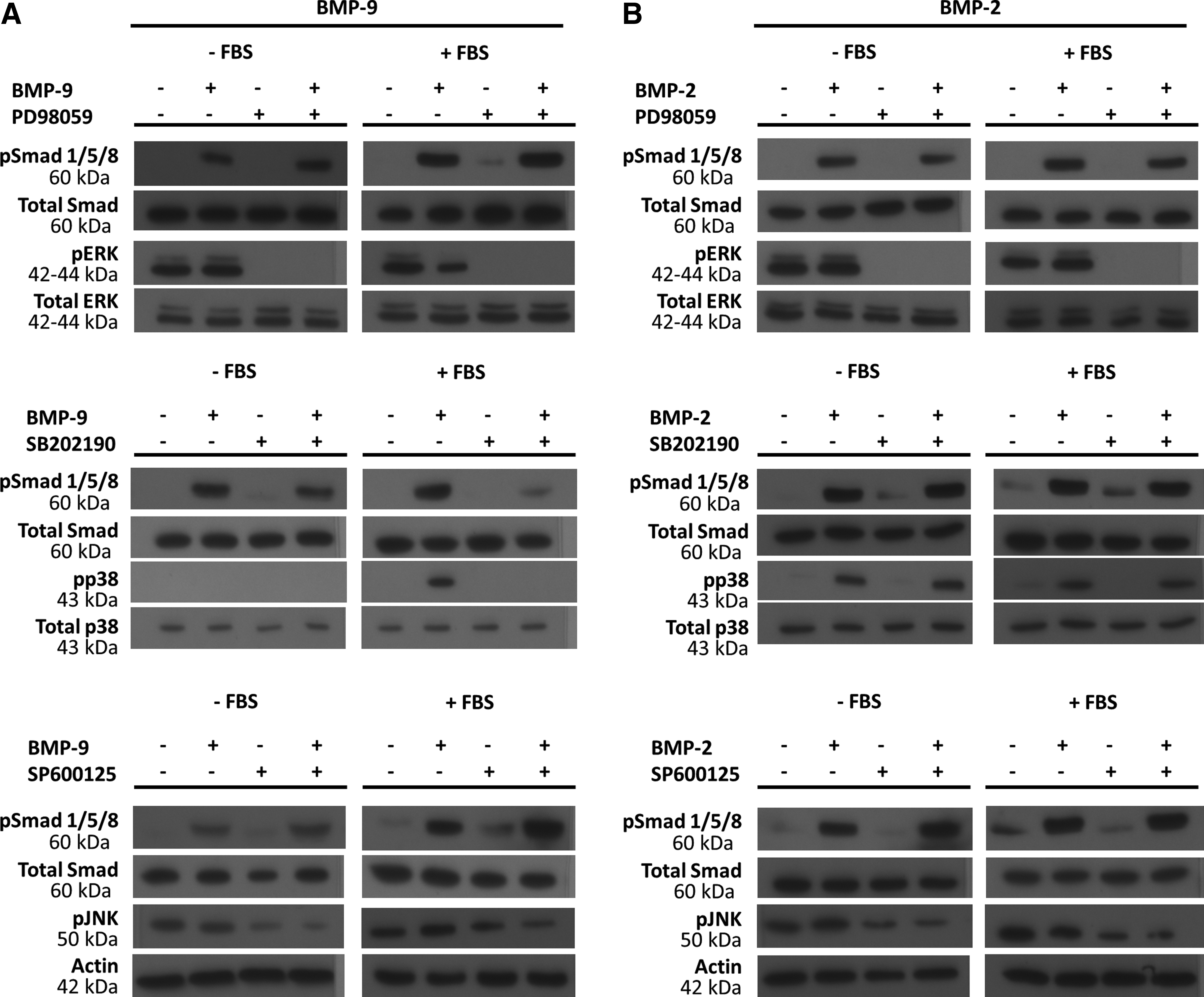
Western blots showing the effect of inhibitors of MEK1/2, p38, and JNK on the phosphorylation of Smad 1/5/8 and ERK1/2, p38 or JNK stimulated for 1 h by
We evaluated the effects of these inhibitors on the ALP activity in cells stimulated with BMPs for 3 days (Fig. 5M). We checked that adding the MEK1/2 inhibitor (PD98059) did not significantly affect the morphology of the cells (Fig. 5B, F, and J). Cells incubated with SP600125 and SB202190 were more elongated than controls (Fig. 5C, D, G, H, K, and L). Cell counting (Fig. 5N) indicated that there were significantly fewer of them than in control wells, and the cells stimulated in the presence of PD98059.
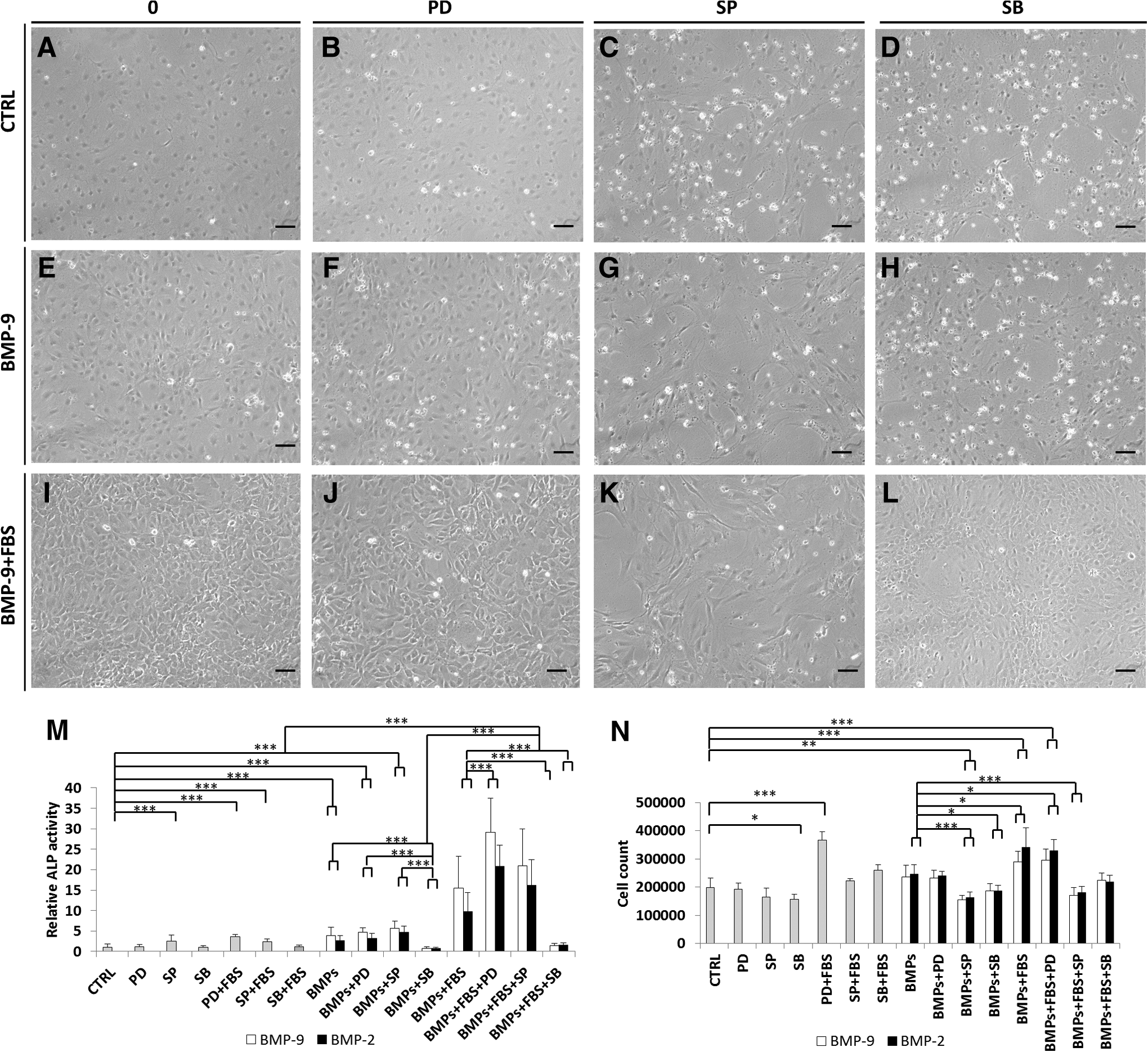
Images of four pictures from three independent experiments performed in quadruplicate of MC3T3-E1 preosteoblasts pretreated with 20 μM MAPK inhibitors (PD98059, SP600125, and SB202190) and then stimulated with BMP-2 or BMP-9 with or without 10% (v/v) FBS
The ALP activity measurements (Fig. 5M) revealed that MC3T3-E1 preosteoblasts pretreated with the p38 inhibitor SB202190 and then stimulated with BMP-9 or BMP-2 without FBS had ALP activities similar to that of the control (p>0.05). In contrast, cells pretreated with the MEK1/2 inhibitor and then stimulated with BMP-2 or BMP-9 without FBS had ALP activities similar to those of cells incubated with BMP-2 or BMP-9 alone. Cells pretreated with PD98059 or SP600125 and then stimulated with either BMP-2 or BMP-9+FBS had about twice the ALP activity induced by BMP-2 or BMP-9+FBS without inhibitor pretreatment. In contrast, SB202190 totally inhibited the effect of BMP-9+FBS; the activity obtained was similar to that of the control (CTRL, Fig. 5M). Other studies also showed that pretreating C3H10T1/2 with PD98059 cells infected by AdBMP-9 or incubated with a conditioned medium collected 48 h after infection of HCT116 cells by AdBMP-9 had much more ALP activity. In contrast, p38 inhibitor SB203580 blocks their osteogenic differentiation.32,40,41
The JNK inhibitor SP600125 increased the ALP activity of cells stimulated with BMP-2+FBS over that obtained with BMP-2 alone. It had no effect on BMP-9 with FBS.
The inhibition of the ERK1/2 pathway and increased p38 phosphorylation in MC3T3-E1 preosteoblasts stimulated with BMP-9 in the presence of FBS may explain the effect of serum on preosteoblast response to BMP-9.
Effect of IGF-2 on signaling pathways and ALP activity with and without BMP-2 or BMP-9
FBS lowers the EC50 of BMP-9 required to activate the Smad 1/5/8 signaling, Osx gene expression, and the ALP activity in MC3T3-E1 preosteoblasts. We attempted to identify the GF(s) that mimicked this effect. The main candidates were IGF-2 and bFGF. It was shown that bFGF can increase the response of primary calvarial osteoblasts to BMP-2, in terms of Smad signaling and Runx2 gene expression,42,43 while IGF-2 is one of the most abundant GFs in FBS and seems to promote the osteoinductive effect of AdBMP-9. 6 Cells were first screened by incubating them for 3 days with 10 ng/mL of bFGF. 6 The ALP activity in cells treated with bFGF±BMPs was similar to that of the control (data not shown).
We determined the effect of IGF-2 on the preosteoblasts by measuring the effect of several recombinant human (rh)IGF-2 doses (from 0 to 100 ng/mL) on the responses of the Smad 1/5/8 and MAPK pathways, since this high dose (100 ng/mL) is about 10% of the highest concentration in the FBS (150–950 ng/mL) (Fig. 6A). Increasing the IGF-2 concentration increased dose dependently the phosphorylation of Smad 1/5/8 (Fig. 6A). IGF-2 also had a dose-dependent effect on the MAPKs. The phosphorylated ERK1/2 decreased at high doses of IGF-2 (50 and 100 ng/mL). This effect was more noticeable at 100 ng/mL. Phosphorylation of p38 was also more important at 100 ng/mL. Cells stimulated with IGF-2+BMP-9 also had more highly phosphorylated Smad 1/5/8 than cells stimulated with BMP-9 alone (Fig. 6B). The phosphorylation of ERK1/2 in these cells was decreased, while the amount of phosphorylated p38 was increased in comparison to the control.
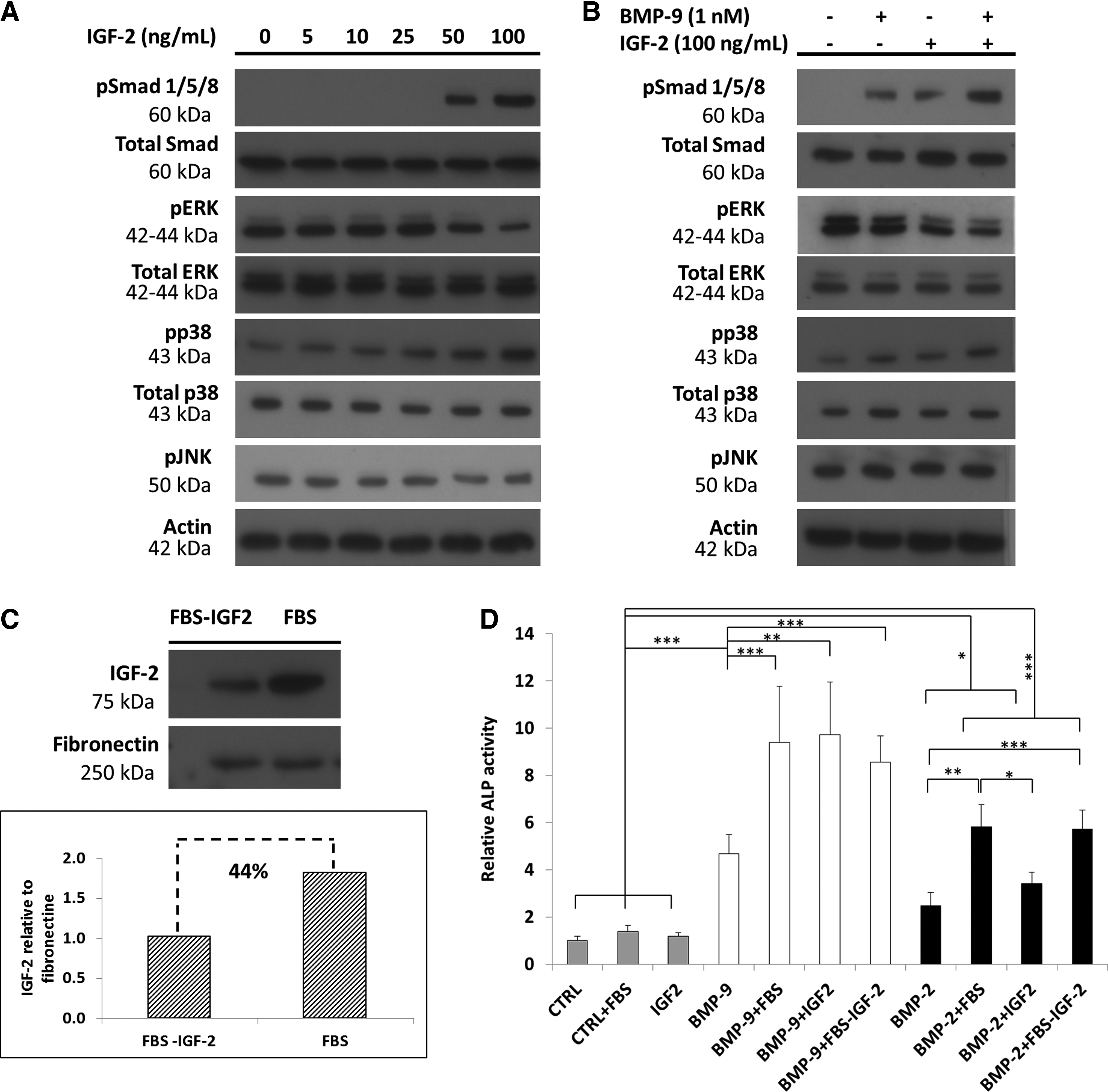
The IGF-2 (Fig. 6A, B) has an impact on the phosphorylated state of the MAPKs similar to that of FBS (10% v/v) on BMP-9-stimulated cells (Fig. 1), decreasing ERK1/2 phosphorylation at high dose (100 ng/mL), while increasing p38 phosphorylation. IGF-2 at high dose also induced an increase in the phosphorylation of Smad 1/5/8. Our inhibitor assays suggest that the loss of phosphorylated ERK1/2 and stabilization of the phosphorylated p38 are the keys to the osteogenic effect of BMP-9 on MC3T3-E1 preosteoblasts. IGF-2 at high dose could therefore induce an intracellular signaling that may favor the cell response to BMP-9 as shown in Figure 6B. However, further experiments are required to determine the effect of IGF-2 on other signaling cascades such as the canonical Wnt pathway that may play a role in the BMP-9 cell response. 28
We then investigated the effect of 100 ng/mL rhIGF-2 with BMP-2 or BMP-9 (1 nM) on ALP activity in MC3T3-E1 preosteoblasts (Fig. 6D). The ALP activity in cells stimulated with IGF-2 alone was similar to that of the control (p>0.05). IGF-2 combined with BMP-2 also produced an ALP activity that was not significantly different from that induced by BMP-2 alone (p>0.05). In contrast, adding IGF-2 to BMP-9 synergistically increased the ALP activity above that of cells stimulated with BMP-9 alone (p<0.001). Chen et al. 10 found that AdIGF-2 increased the differentiation of AdBMP-9-infected murine mesenchymal stem cells, but the amounts of GFs used were not specified. We found that 100 ng/mL IGF-2 mimics the synergistic effect of FBS on BMP-9 (1 nM)-stimulated cells.
Since IGF-2 may be one of the GFs in FBS involved in the observed synergistic effect, we used IGF-2-immunoprecipitated FBS to verify if depleting IGF-2 decreased the ALP activity. The IGF-2 immunoprecipitation efficiency (44%) was determined by western blots and densitometric analysis (Fig. 6C). The partial removal of IGF-2 from the FBS (FBS-IGF-2) had no significant effect on the ALP activity in cells stimulated with either BMP-9 or BMP-2 (Fig. 6D). FBS deprived of about 44% of its IGF-2 should have led to a significant decrease in the ALP activity as the IGF-2 concentration in our experimental conditions (FBS 10% v/v) was 15–95 ng/mL. 6 Furthermore, we checked the effect of IGF-2 (60 ng/mL) on BMP-9 (1 nM)-stimulated cells, but no significant differences were observed in comparison to cells stimulated with BMP-9 alone (data not shown). Thus, there must be one or more molecules in FBS that act in synergy with BMP-9. For instance, Liu et al. 44 have recently shown that AdEGF could also potentiate the effect of AdBMP-9 in murine mesenchymal stem cells and might be an interesting target. Thus, we used a subclone of murine MC3T3-E1 preosteoblasts well known to behave like human preosteoblasts to show the synergistic effect of both FBS and IGF-2 on the cell response to BMP-9. 27 However, a future study will focus on the influence of serum or GFs like IGF-2 on BMP-9 action using human mesenchymal stromal cells that can differentiate along an osteoblastic lineage.
Conclusion
Our findings shed new light on the dose-dependent effects on MC3T3-E1 preosteoblasts of BMP-9, with and without FBS. FBS markedly potentiates the effect of BMP-9, influencing cell signaling, osteogenic gene expression, and ALP activity. It also reduces the EC50 of BMP-9 about sixfold, while IGF-2 (100 ng/mL) mimics the effect of FBS on the response of cells to BMP-9. These findings are of particular interest as BMPs are very expensive to produce and purify. Perhaps, the amounts of GF required may be greatly reduced, leading to new cost-effective bone repair strategies. Last, this work showed that the combination of IGF-2 and BMP-9 is particularly relevant for bone tissue engineering applications.
Footnotes
Acknowledgments
The authors thank Dr. Owen Parkes for editing the English text. This research was supported by the Canada Research Chair in Cell-Biomaterial Biohybrid Systems held by Prof. Nathalie Faucheux and the Natural Sciences and Engineering Research Council of Canada (NSERC) program.
Disclosure Statement
No competing financial interests exist.