
Abstract
Background:
Adipose tissue represents an abundant and easily accessible source of multipotent cells that may serve as an excellent building block for tissue engineering. However, adipose-derived stromal cells (ASCs) are a heterogeneous group and subpopulations may be identified with enhanced osteogenic potential.
Methods:
Human ASC subpopulations were prospectively isolated based on expression of bone morphogenetic protein receptor type-IB (BMPR-IB). Unsorted, BMPR-IB(+), and BMPR-IB(−) cells were analyzed for their osteogenic capacity through histological staining and gene expression. To evaluate their in vivo osteogenic potential, critical-sized calvarial defects were created in immunocompromised mice and treated with unsorted, BMPR-IB(+), or BMPR-IB(−) cells. Healing was assessed using microcomputed tomography and pentachrome staining of specimens at 8 weeks.
Results:
Increased osteogenic differentiation was noted in the BMPR-IB(+) subpopulation, as demonstrated by alkaline phosphatase staining at day 7 and extracellular matrix mineralization with Alizarin red staining at day 14. This was also associated with increased expression for osteocalcin, a late marker of osteogenesis. Radiographic analysis demonstrated significantly enhanced healing of critical-sized calvarial defects treated with BMPR-IB(+) ASCs compared with unsorted or BMPR-IB(−) cells. This was confirmed through pentachrome staining, which revealed more robust bone regeneration in the BMPR-IB(+) group.
Conclusion:
BMPR-IB(+) human ASCs have an enhanced ability to form bone both in vitro and in vivo. These data suggest that positive selection for BMPR-IB(+) and manipulation of the BMP pathway in these cells may yield a highly osteogenic subpopulation of cells for bone tissue engineering.
Introduction
I
Recent investigations have focused on the use of adipose-derived stromal cells (ASCs) for skeletal tissue engineering given their well-described ability to form bone.3–5 However, the stromal vascular fraction, from which ASCs are harvested, is composed of a heterogeneous population that also includes circulating blood cells, fibroblasts, pericytes, endothelial cells, and adipocyte progenitor cells. 6 To further enhance bone regeneration in cell-based strategies, cell sorting has been employed to enrich for subsets of cells with enhanced osteogenic potential. Our laboratory has already demonstrated that isolation of cellular subsets based on cell-surface marker expression, such as CD105 and CD90, may yield populations of ASCs with increased bone-forming capacity.7,8
CD105 is an accessory receptor for transforming growth factor (TGF)-β, and it has been purported that the enhanced osteogenic potential of CD105-depleted ASCs may be mediated through a reduction in TGF-β1/Smad2 signaling. 7 Isolation of CD90(high) cell populations from ASCs also yields a highly osteogenic subpopulation with studies demonstrating that expression of CD90 in stromal cells occurs once the cells begin to express an osteoblastic phenotype.8–11 However, the exact mechanism responsible for osteogenesis in CD90(high) cells is not fully understood. CD105 and CD90 are widely expressed in multipotent stromal cells, and these markers have also been demonstrated to play a role in other nonosteogenic processes such as angiogenesis. 10
Importantly, however, significant phenotypic drift has been observed for many cell surface markers, including both CD105 and CD90.7,10,12 In addition, conflicting data exist regarding the correlation of CD105 and CD90 expression with osteogenic differentiation capacity, and these data have complicated use of these markers for prospective isolation of freshly harvested ASCs.7,10,13–15 In contrast, we have demonstrated that osteogenesis in ASCs is associated with enhanced expression of the bone morphogenetic protein receptor type IB (BMPR-IB) isoform.16,17 BMP signaling through this receptor pathway has an important role in directing bone formation and differentiation.18,19 After BMP ligand binding to the BMP receptors and the formation of hetero-oligomeric complexes of type-I and type-II receptors on the cell surface, the type-II receptor phosphorylates the type-I receptor.20–22 Activated type-I receptors, in turn, phosphorylate Smad1/5/8, which then assemble into complexes with Smad4 and translocate to the nucleus to regulate osteogenic gene transcription through Runx2. 23 Since osteogenic differentiation in ASCs has been shown to be dependent on signaling through BMPR-IB, this cell surface protein may thus provide an alternative marker to identify a more highly osteogenic subpopulation among the larger heterogeneous mix of cells.
Given these findings, we thus evaluated the ability to isolate human ASCs based on expression of BMPR-IB and then determined the capacity for these fractions, BMPR-IB(+) and BMPR-IB(−), along with unsorted ASCs, to undergo in vitro osteogenesis. Gene expression analysis was also performed to compare transcript levels for various osteogenic markers. Finally, the bone regenerative capacity for unsorted, BMPR-IB(+), and BMPR-IB(−) ASCs was evaluated in critical-sized calvarial defects in immunocompromised mice.
Materials and Methods
Cell isolation
Human lipoaspirate specimens were obtained after acquiring informed consent from six female patients, in accordance with the Stanford University Institutional Review Board guidelines. ASCs were harvested from the adipose tissue of patients (aged between 31 and 35 years) without any medical co-morbidities, who were undergoing elective liposuction of the abdominal region. 3 Briefly, raw lipoaspirates were washed and treated with 0.075% collagenase type I (Sigma-Aldrich) in Hank's balanced salt solution (Cellgro) for 1 h at 37°C with gentle agitation. The cellular pellet was resuspended in ACK lysis buffer (Gibco) to remove red blood cells. Cell counting was performed before cell sorting.
Flow cytometry and magnetic cell sorting
Freshly isolated ASCs were incubated in cytometry buffer (phosphate buffered saline, 2% fetal bovine serum [FBS]) containing Human BMPR-IB/ALK-6 Biotinylated Affinity Purified Polyclonal Antibody (R&D Systems) at a 1:50 dilution. Cells were then washed and centrifuged before a 30-min incubation in cytometry buffer containing Streptavidin PE-Cy7 (eBiosceince) in a 1:100 dilution. After this, cells were again washed, centrifuged, and resuspended in cytometry buffer, then passed through a 40-micron cell strainer before analysis using fluorescence-activated cell sorting (FACS). FACS analysis was performed on an FACS Aria II instrument (BD Biosciences) with the use of a 100-μm nozzle. Gates were drawn as shown in Figure 1.
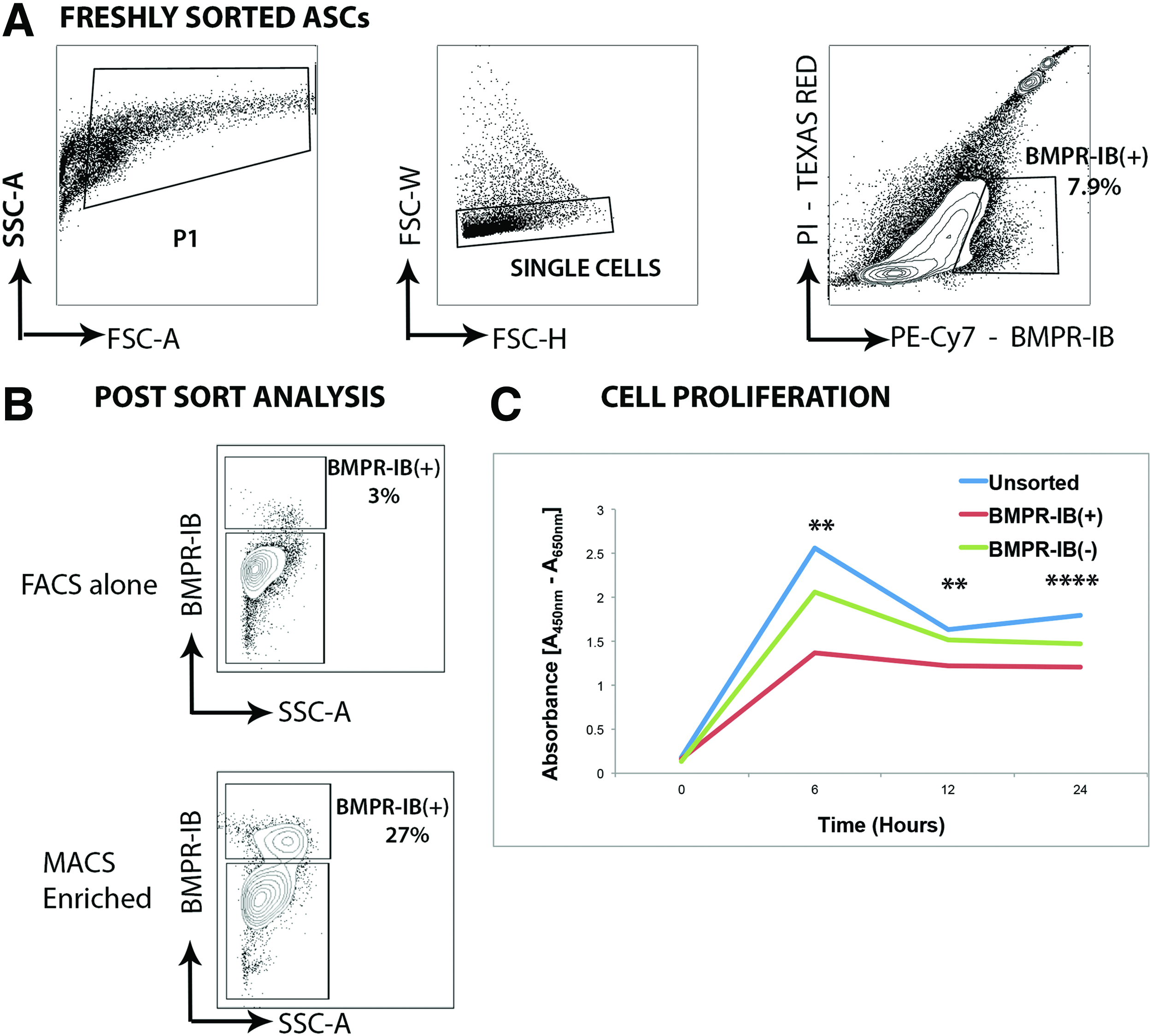
Cell sorting of ASCs for expression of BMPR-IB. Gating scheme for BMPR-IB expression in ASCs. Preliminary cell selection based on size and complexity to exclude debris (P1 gate). Doublets were excluded through selective gating to yield single cells.
For subsequent isolation of BMPR-IB(+) and BMPR-IB(−) cell populations, magnetic-activated cell sorting (MACS) was employed to utilize a more clinically translatable cell isolation approach in accordance with the manufacturer's instructions (Miltenyi Biotec, Inc.). Briefly, freshly isolated ASCs were incubated for 30 min in MACS buffer (Miltenyi Biotec, Inc.) containing Human BMPR-IB/ALK-6 Biotinylated Affinity Purified Polyclonal Antibody (R&D Systems) at a 1:50 dilution. After this, cells were washed with MACS buffer, centrifuged, and the supernatant was aspirated before resuspension of the cell pellet in MACS buffer containing Streptavidin Microbeads (Miltenyi Biotec) at a 1:100 dilution. Cells were then washed again, resuspended in MACS buffer, and the suspension was passed through a 40-micron cell strainer before running on an MACS LS column (Miltenyi Biotec, Inc.). Post–MACS analysis using FACS was performed to determine MACS sorting efficiency.
In vitro differentiation assays
Osteogenic differentiation was performed on freshly isolated unsorted, BMPR-IB(+), and BMPR-IB(−) cell populations to compare bone-forming capacity in vitro. ASCs were plated in six-well plates at a density of 100,000 cells/well. At 80% confluence, cells were treated with osteogenic differentiation media (Dulbecco's modified Eagle's medium, High Glucose, GlutaMAX, HEPES supplemented with 10% FBS, 1% penicillin, 100 μg/mL ascorbic acid, and 10 mM B-glycerophosphate). Alkaline phosphatase staining and quantification was performed at day 7 of differentiation. Alizarin red staining was performed at 14 days to assess extracellular mineralization. For quantification, an alizarin red extraction solution (20% methanol/10% acetic acid in ddH2O) was added to each well. After 15 min, photometric quantification was performed on a spectrophotometer at a wavelength of 450 nm. All experiments were performed in triplicate to facilitate statistical analysis.
Cell proliferation assay in vitro
Unsorted, BMPR-IB(+), and BMPR-IB(−) cells were isolated and cell proliferation was measured with an XTT-based assay (Cell Proliferation Kit II XTT; Roche Applied Science) according to the manufacturer's instructions. Briefly, cells were seeded in 96-well plates at a density of 20,000 cells per well in culture medium. Plated were incubated overnight at 37°C and 5% CO2, and proliferation was evaluated over a 24-h period. The absorbance of each well was determined using a microplate reader at 450 and 650 nm (SpectraMAX 384 Plus; Molecular Devices Ltd.). All assays were performed in triplicate.
Reverse transcription and quantitative real-time polymerase chain reaction
RNA was isolated from cells using the RNeasy Mini kit (Qiagen) as described by the manufacturer's protocol. Reverse transcription was performed, and osteogenic gene expression was examined by quantitative real-time polymerase chain reaction (qRT-PCR) using the Applied biosystems Prism 7900HT sequence detection system (Applied Biosystems) and SYBR Green PCR Master Mix (Applied Biosystems). All values were normalized to GAPDH expression in the corresponding samples. Specific primer sequences for genes examined were obtained from PrimerBank and shown in Table 1. 24
In vivo bone formation
To evaluate osteogenesis in vivo, nonhealing, critical-sized (4-mm) calvarial defects were created in the right parietal bone of 60-day-old male Crl:CD-1-Foxn1nu athymic nude mice (Charles River Laboratories), as previously described. 25 All research involving vertebrate animals was performed in accordance with protocols approved by the Stanford Administrative Panel on Laboratory Animal Care (APLAC). Hydroxyapatite (HA)-coated poly(lactic-co-glycolic acid) (PLGA) scaffolds were fabricated from 85/15 PLGA by solvent casting and a particulate leaching process as previously described (inherent viscosity=0.61 dL/g, Birmingham Polymers). 5 For apatite coating, the obtained PLGA scaffolds were incubated in simulated body fluid solution for 12 h as previously described.26–28 In preparation for cell engraftment, scaffolds were seeded with ASCs at 24 h before implantation. Each scaffold was seeded with 200,000 cells in 125 μL of medium in 96-well culture plates and incubated overnight. Animals were treated with the HA-PLGA scaffold seeded with the following cell types: unsorted ASCs (n=5), BMPR-IB(+) ASCs (n=5), or BMPR-IB(−) ASCs (n=5).
For microcomputed tomography (micro-CT) scans, the mice were anesthetized with isoflurane. Imaging was performed using a Siemens Inveon MicroPET/CT scanner (Siemens Medical Solutions, Inc.). Using our scan protocol parameters, each high-resolution 100-μm image was acquired in a total scan time of 10 min. Baseline CT scans were obtained on postoperative day 1 (week 0), and subsequently at 2, 4, 6, and 8 weeks after surgery. Data were reconstructed into three-dimensional surfaces using the GEMS Microview software (GE Medical Systems). The three-dimensional reconstructed images were then analyzed by quantifying pixels in the defect using Adobe PhotoShop. Percentage healing was determined by dividing the defect area by the immediate postoperative defect size.
Histological analysis
At 8 weeks postoperatively, calvariae were harvested, immediately fixed in 10% formalin overnight, decalcified in 19% EDTA, dehydrated through an ethanol series, and embedded in paraffin, as previously described. 29 Deparaffinized sections were stained with Movat's pentachrome to detect bone matrix formation. Bright field images were obtained with a 20×objective at room temperature using a Leica DM5000 microscope (Leica Microsystems, Inc.) equipped with a DFC300FX camera. The images were analyzed using Leica IM1000 Version 4.0 Image Acquisition Software (Leica Microsystems).
Data analysis and statistics
Means and standard deviations were calculated, and statistical analysis was performed using a one-way analysis of variance (ANOVA) when more than two groups were compared. Post-hoc analysis was performed using Bonferroni's multiple-comparisons test for a comparison between groups. A *p-value <0.05 was considered statistically significant.
Results
Fluorescence-activated cell sorting
The ability to sort for a single surface marker ensures the largest capture of cells in the shortest amount of time. Given previous work by our laboratory demonstrating the importance of BMPR-IB in osteogenic commitment of ASCs, we used flow cytometry to evaluate ASCs based on BMPR-IB expression. Through FACS analysis, we identified that 3–8% of freshly isolated ASCs were BMPR-IB(+) (Fig. 1). However, despite the advantages of using FACS for cell sorting, it is a labor-intensive procedure that requires a skilled operator. To utilize a method that could be more easily translated clinically, subsequent studies employed magnetic cell sorting to isolate populations of BMPR-IB(+) cells. 30 We demonstrated that MACS sorting resulted in an almost 10-fold enrichment of BMPR-IB(+) ASCs when compared with FACS sorting alone (Fig. 1). Assessment of the proliferative potential of unsorted, BMPR-IB(+), and BMPR-IB(−) cells in vitro using an XTT assay demonstrated that the proliferative potential of the BMPR-IB(+) cell population was significantly lower than the unsorted and BMPR-IB(−) cell populations over a 24 h period of in vitro culture (Fig. 1) (**p<0.01, ****p<0.0001, ANOVA).
Enhanced in vitro osteogenesis of BMPR-IB(+) ASCs
To characterize the osteogenic potential of MACS-sorted ASCs, cells were cultured with osteogenic differentiation medium. At 7 days of culture, staining for Alkaline Phosphatase activity, an early marker of bone formation, was significantly increased in the BMPR-IB(+) group compared with the unsorted ASCs (*p<0.05) and BMPR-IB(−) ASCs (**p<0.01) (Fig. 2). After 14 days in osteogenic differentiation medium, Alizarin red staining was performed to assess for extracellular matrix mineralization. Similar to that observed with alkaline phosphatase staining, the BMPR-IB(+) population demonstrated significantly greater staining for extracellular mineralization compared with the unsorted population (*p<0.05) and BMPR-IB(−) population (**p<0.01) (Fig. 3).

Alkaline phosphatase staining of ASC subpopulations. Alkaline phosphatase staining of
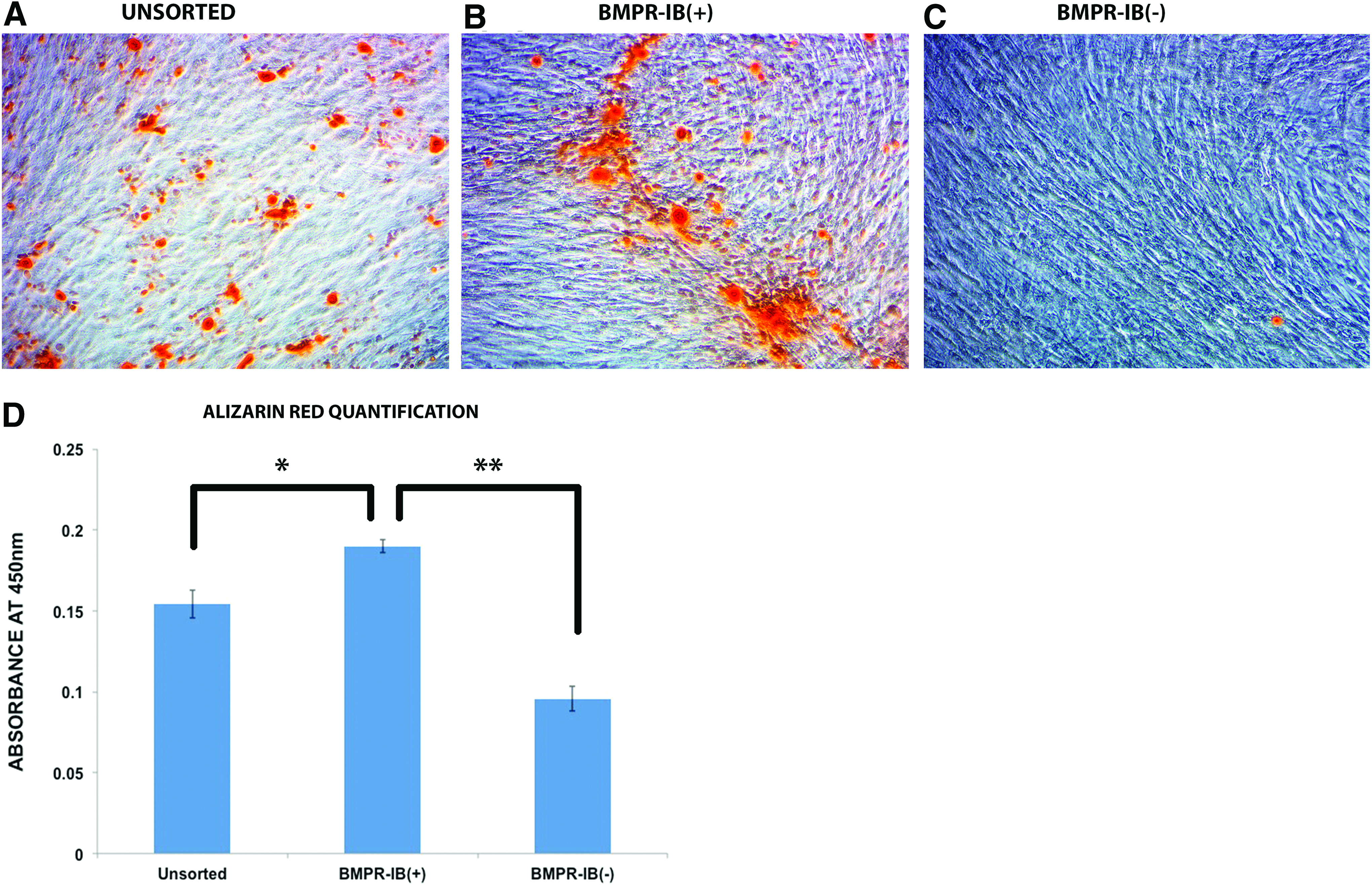
Alizarin red staining of ASC subpopulations. Alizarin red staining of ASCs after 14 days of osteogenic differentiation for
Increased osteogenic marker expression in BMPR-IB(+) ASCs
Given the enhanced osteogenic capacity of BMPR-IB(+) ASCs in vitro, as shown by histological staining, we compared the baseline expression of osteogenic markers in these cells relative to unsorted ASCs and BMPR-IB(−) ASCs. Interestingly, BMPR-IB(+) cells demonstrated significantly less expression for the early marker Runx-2 compared with unsorted ASCs and BMPR-IB(−) ASCs (***p<0.001) (Fig. 4A). Transcript levels for osteopontin, an intermediate marker, were not significantly different between any of the groups (Fig. 4B). Finally, freshly isolated ASCs sorted for BMPR-IB(+) demonstrated significantly increased expression levels of the late osteogenic marker osteocalcin (*p<0.05) (Fig. 4C). Collectively, gene expression analyses, thus, suggest BMPR-IB(+) ASCs to be a cell population already more committed along an osteogenic lineage. 31
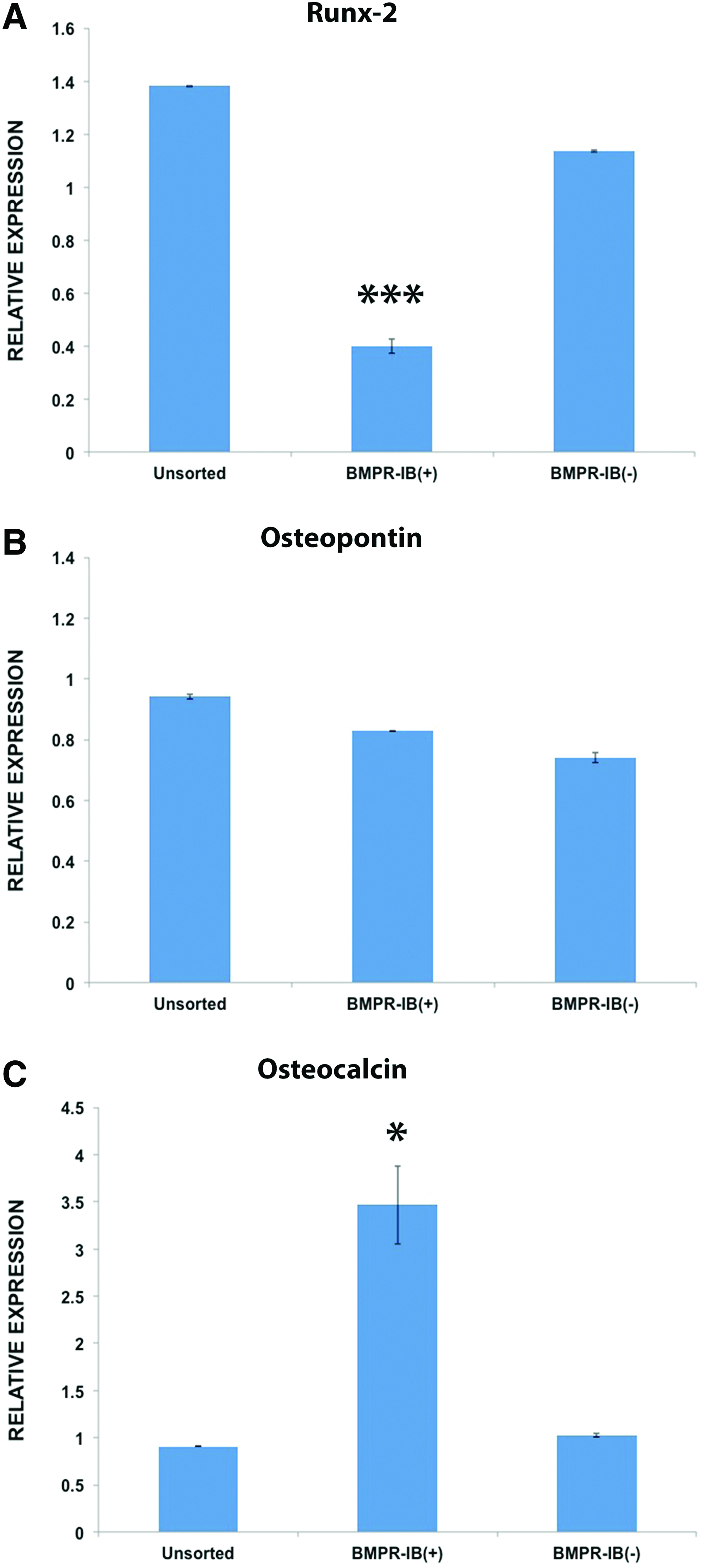
Osteogenic gene expression in ASC subpopulations. Gene expression for markers of osteogenic differentiation in unsorted, BMPR-IB(+), and BMPR-IB(−) ASCs.
BMPR-IB(+) ASCs accelerate in vivo bone regeneration
With enhanced in vitro osteogenesis noted in BMPR-IB(+) ASCs, we next determined whether this same subpopulation could similarly accelerate in vivo bone regeneration. Critical-sized (4-mm) calvarial defects in the parietal bone of male athymic, nude mice were created into which HA-PLGA scaffolds seeded with unsorted, BMPR-IB(+), or BMPR-IB(−) ASCs were placed. Defects treated with BMPR-IB(+) ASCs demonstrated accelerated bone regeneration compared with defects treated with unsorted or BMPR-IB(−) ASCs. Micro-CT scans revealed significantly greater bone formation as early as 2 weeks after repair (**p<0.01) (Fig. 5A). By 8 weeks, BMPR-IB(+) ASCs resulted in 92% regeneration compared with 46% by BMPR-IB(−) ASCs and 58% in the unsorted ASCs (**p<0.01) (Fig. 5B). Histological staining was performed to confirm radiographic findings, and at 8 weeks, staining with Movat's pentachrome revealed more robust bone formation across the region of the defect with BMPR-IB(+) ASCs compared with unsorted and BMPR-IB(−) ASCs (Fig. 6).
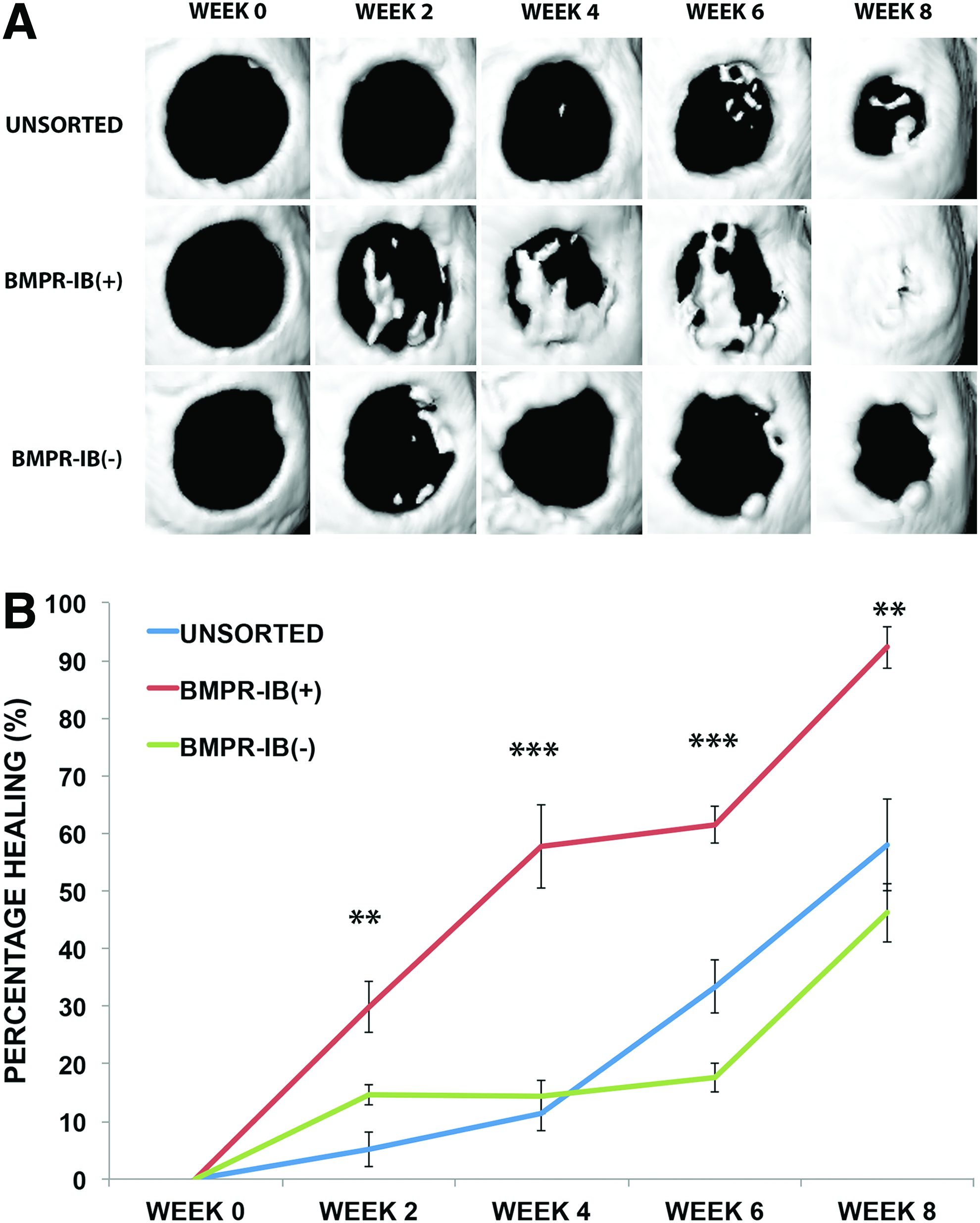
Healing of critical-sized calvarial defects with different ASC subpopulations.
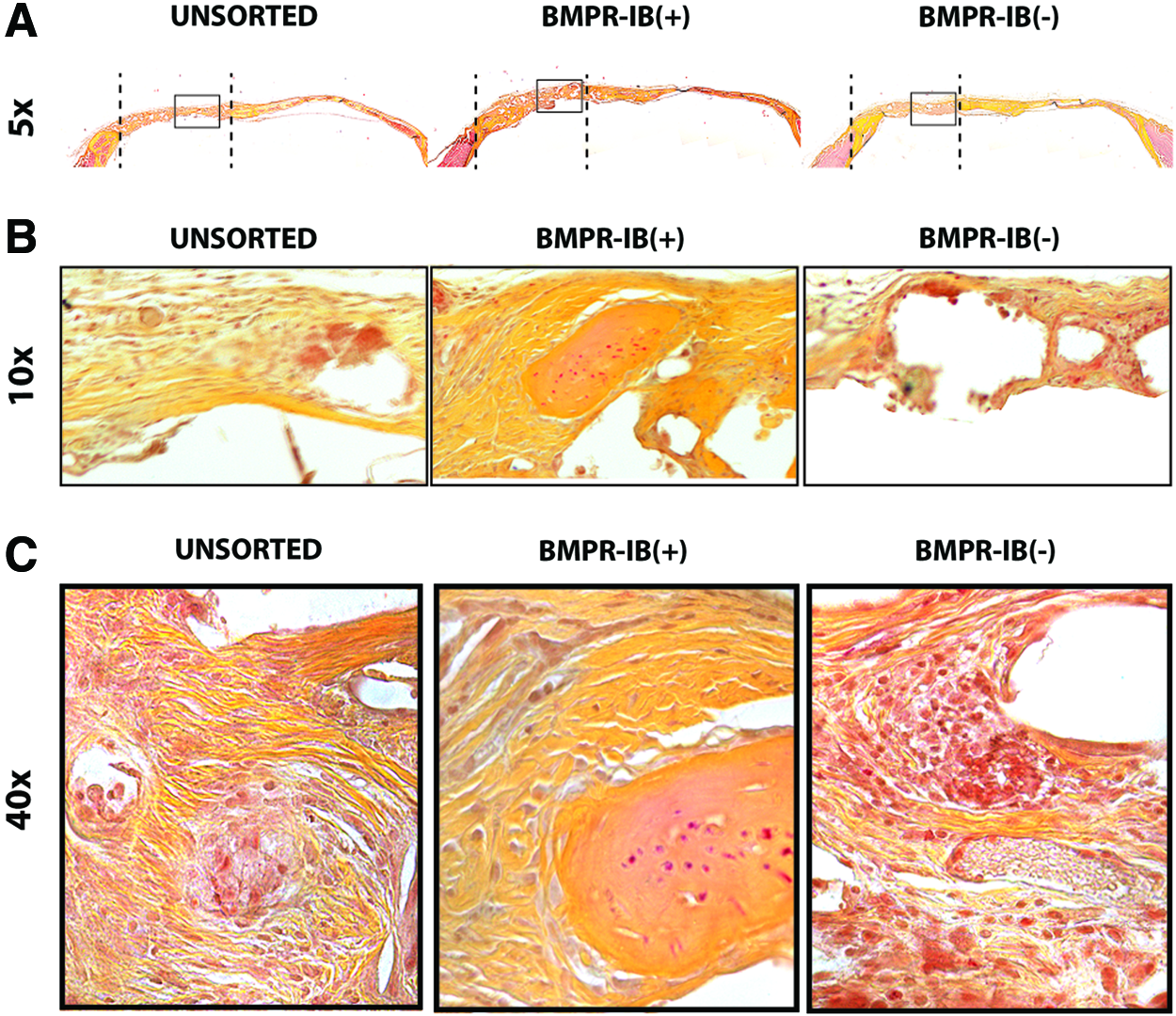
Histological staining of bone regenerate.
Discussion
Reconstruction after large-scale bone loss frequently employs autografts, allografts, or xenografts. Their use, however, is associated with multiple complications, including limited availability, donor site morbidity, disease transmission, and immunological rejection.32,33 To overcome this problem, novel cell-based strategies to regenerate de novo skeletal tissue have been proposed. As a potential building block for skeletal tissue engineering, a wide range of osteogenic cell types have been identified, including ASCs and bone marrow-derived mesenchymal cells.5,34,35 Importantly, the type of cell chosen will likely be critical for clinical translation and the ultimate success of bone repair. Our laboratory has extensively studied the role of ASCs in healing critical-sized calvarial defects in mice.5,29 However, the heterogeneity of ASCs has been well described, and this remains a major impediment to the development of therapeutic strategies employing these cells. 8
An informatics approach utilizing single-cell transcriptional analysis has previously identified cell surface markers such as CD105 and CD90 to be associated with bone-forming transcriptional profiles. 36 Importantly, however, ASCs have been shown to demonstrate a significant phenotypic drift for numerous surface markers, thus making the prospective isolation of cell populations difficult.7,8,10 For example, we have demonstrated this to be the case for CD105, in which cell sorting for CD105low yields a highly osteogenic subset of ASCs. 7 We have previously demonstrated that 90% of freshly sorted ASCs are CD105low, but this is reduced to 50% after 36 h of in vitro culture. 7 While isolation of CD105low fractions at this time point yields a more osteogenic subpopulation, these progressively become CD105high, making any direct contribution of the CD105low phenotype to enhanced bone-forming capacity dubious.
In contrast to CD105 and CD90, BMPR-IB has already been shown to play a critical role in the osteogenic commitment of ASCs.16,17 We have previously reported that ASCs respond to BMPs with enhanced bone formation, and BMPs are known to signal through a serine/threonine kinase heterotetrameric complex composed of BMPR-IA, -IB, and–II.31,37 Interestingly, Chen et al. found BMPR-IB to regulate osteogenic differentiation, while BMP signaling through BMPR-IA promoted adipogenesis. 22 BMPR-IB-deficient mice have also been described with various defects throughout the appendicular skeleton.22,38 Finally, in the absence of BMPR-IB, a significant reduction in the bone-forming capacity of ASCs has been observed. 16 Furthermore, growth factors such as TGF-B1, FGF-2, and PDGF-AB, which have been shown to play an important role in osteogenesis, may modulate the BMP/BMPR pathway by upregulating expression of the BMPR and enhance the osteogenic response of these cells to BMP. 39
With these considerations in mind, we prospectively sorted freshly harvested ASCs based on BMPR-IB expression into three subgroups and found the BMPR-IB(+) fraction to possess enhanced in vitro osteogenic potential compared with BMPR-IB(−) and unsorted ASCs. This was demonstrated by both histological and gene expression analysis. The greatest amount of staining for alkaline phosphatase and extracellular matrix mineralization with Alizarin red was noted with BMPR-IB(+) ASCs. Interestingly, this was associated with the lowest transcript levels for the early marker Runx-2 in BMPR-IB(+) ASCs, which is consistent with a cell population further committed along an osteogenic lineage. Also consistent with increased osteogenic commitment was the finding of significantly lower proliferation along with elevated levels of the late marker osteocalcin.40,41 Importantly, our in vitro findings translated to enhanced bone regeneration in vivo when BMPR-IB(+) cells were implanted into critical-sized mouse calvarial defects. This was demonstrated by both micro-CT imaging and histological analysis at 8 weeks which showed accelerated and more robust bone formation than that generated by BMPR-IB(−) cells.
An interesting observation in our study was that isolation of BMPR-IB(+) cells using MACS or FACS resulted in a low yield of this cell population and that the MACS-sorted population of cells contained a large number of negative cells on subsequent FACS analysis. Use of a low affinity of the antibody for the BMPR-IB receptor and an indirect method of staining to isolate these cells may have contributed to lower efficiency cell isolation. To overcome these issues, it would be ideal to use a primary-conjugated MACS antibody with a high affinity for the receptor.
Indeed, the choice of biomimetic scaffold plays an important role in driving appropriate tissue regeneration in vivo. Since the extracellular matrix (ECM) plays a pivotal role in guiding various biological processes, creating a scaffold that mimics the normal ECM should enhance tissue regeneration. HA, for example, is a major component of the skeletal ECM, and it has previously been demonstrated that coating of PLGA scaffolds with HA enhances osteogenesis in vivo. 5 It is postulated that the interaction of CD44 with HA results in enhanced chondrocyte proliferation, increased production of GAG, and, ultimately, enhanced type II collagen and aggrecan gene expression. 42 One of the limitations of using this scaffold, however, is that the cells remain in a hypoxic environment, and there is a requirement for angiogenesis for the cells to survive in the defect. Other studies have demonstrated methods to overcome these problems, which include the incorporation of pro-angiogenic cytokines into the scaffold.2,43 Of note, organic solvents used in the synthesis of these scaffolds can affect the bioactivity of the cytokines, and so novel techniques such as microencapsulation of cytokines, protein PEGylation, or gene therapy may be required to overcome these issues.44–46
While it has been extensively demonstrated that unsorted ASCs are capable of healing critical-sized calvarial defects, 29 we observed in this study that unsorted cell populations demonstrated a significantly reduced ability to repair defects compared with the BMPR-IB(+) sorted cell populations. Indeed, recent reports have concluded that cell sorting does not influence the osteogenic potential of ASCs. However, we have demonstrated that sorting for an osteogenic subset of ASCs results in enhanced calvarial bone healing compared with using unsorted ASCs. The variation in osteogenic potential of unsorted ASCs that is observed between studies may reflect variability between different patient samples. In addition, we have previously demonstrated the heterogeneous nature of ASCs using single-cell transcriptional analysis, which may explain the differences that we see. 36 Alternatively, interactions between the osteoinductive HA coating of the scaffold may interact specifically with the BMPR-IB(+) subset of ASCs to significantly enhance bone formation in this group by promoting BMP signaling.47,48
Our data, nonetheless, confirm that selection for human ASCs expressing the BMPR-IB isoform yields a highly osteogenic subpopulation of cells, as demonstrated by their enhanced capacity to generate bone both in vitro and in vivo. In addition, the ability to magnetically sort ASCs using a single marker with a direct role in osteogenesis offers the potential to quickly isolate a highly osteogenic subpopulation of cells from the larger heterogeneous pool, thus offering greater clinical translational potential by making it possible to easily isolate this osteogenic subpopulation in the operating room.
Footnotes
Acknowledgments
The authors gratefully acknowledge the following funding sources that supported this study: the National Institutes of Health research grants RO1 DE021683, RO1 DE019434, and UO1 HL09977 to M.T.L; the Tissue Engineering Fellowship from Lucile Packard Children's Hospital at Stanford to A.M.; The Hagey Laboratory for Pediatric Regenerative Medicine; and the Oak Foundation. D.C.W. was supported by the ACS Franklin H. Martin Faculty Research Fellowship, the Hagey Laboratory for Pediatric Regenerative Medicine, and the Stanford University Child Health Research Institute Faculty Scholar Award.
Disclosure Statement
No competing financial interests exist.