
Abstract
Craniofacial trauma is difficult to repair and presents a significant burden to the healthcare system. The inflammatory response following bone trauma is critical to initiate healing, serving to recruit inflammatory and progenitor cells and to promote angiogenesis. A role for inflammation in graft-induced bone regeneration has been suggested, but is still not well understood. The current study assessed the impact of Toll-like receptor (TLR4) signaling on calvarial repair in the presence of morselized bone components. Calvarial defects in wild-type and global TLR4−/− knockout mouse strains were treated with fractionated bone components in the presence or absence of a TLR4 neutralizing peptide. Defect healing was subsequently evaluated over 28 days by microcomputed tomography and histology. The matrix-enriched fraction of morselized bone stimulated calvarial bone repair comparably with intact bone graft, although the capacity for grafts to induce calvarial bone repair was significantly diminished by inhibition or genetic ablation of TLR4. Overall, our findings suggest that the matrix component of bone graft stimulates calvarial bone repair in a TLR4-dependent manner. These results support the need to better understand the role of inflammation in the design and implementation of strategies to improve bone healing.
Introduction
L
Autologous bone graft consists of cellular and matrix components, which are presumed to facilitate integration with host tissue following transplantation. The cellular component of autologous bone is associated with its superior osteogenic properties and includes osteoblasts, osteoclasts, osteocytes, and supporting stromal cells. The acellular component of bone is derived primarily from the bone extracellular matrix (ECM) and its associated biochemical cues, which include growth factors and other signaling molecules secreted by bone cells. The acellular component of bone has osteoconductive and osteoinductive properties, providing architectural support and creating a distinct biochemical environment that supports the formation and maintenance of bone.2,3 The cellular and acellular components of bone graft possess immunologic properties, which are incompletely understood.4,5
Recent studies have demonstrated that a potent inflammatory response to damaged ECM and necrotic or stressed cellular components is also involved in tissue regeneration, highlighting a potential role for graft-induced inflammation in facilitating fracture healing.6,7 Although considerable controversy exists regarding the exact role of inflammation during bone healing, inflammation is well recognized as an integral component of the injury response and has received increasing attention for its role in host tissue regeneration and repair.8,9 The inflammatory response following bone injury is critical for the initiation of healing cascades, serving to recruit inflammatory and progenitor cells and to promote angiogenesis during the remodeling of damaged tissues. 10 A role for inflammation in graft-induced bone regeneration has been suggested, but is still not well understood.
Toll-like receptors (TLRs) play an essential role in innate recognition of microbial products and are critical activators of the innate immune response. Another unique role of TLRs is to sense cellular stress or tissue damage by responding to endogenous ligands released from necrotic cells and damaged tissues in many settings, including musculoskeletal trauma.8,11 TLR expression is primarily detected within macrophages, neutrophils, dendritic cells, and bone cells.12–15 TLR4 signaling is of particular interest in regenerative biology due to its pronounced impact on healing in diverse models of injury and sterile inflammatory disease.13,16
The impact of TLR4 activation on tissue homeostasis and regeneration partially depends upon organ setting and mode of injury. TLR4 signaling has been suggested to mediate bone destruction by inducing inflammation and osteoclastogenesis.17–20 In a previous study, we showed that calvarial bone healing is accelerated in TLR4 knockout (TLR4−/−) mice and is associated with elevated expression of cytokines and osteoclast differentiation markers. 21 Furthermore, our team has previously presented data suggesting that morselized bone graft induces a TLR4-dependent systemic inflammatory response in a bilateral femoral fracture model. 22 Based upon these observations, we hypothesized that TLR4 signaling is necessary for graft-induced bone formation by mediating the remodeling and integration of bone graft within regenerating calvarial defects. To test this hypothesis, we assessed calvarial bone healing using radiography, live computerized tomography (CT), and histological analyses in wild-type (WT) and TLR4−/− mice engrafted with different fractions of morselized bone-enriched implants.
Materials and Methods
Animal care and experimental design
WT C57BL-6J mice (Jackson) and TLR4−/− mice (from an ongoing breeding colony housed at the University of Pittsburgh), between 10 and 12 weeks of age (20–30 g), were utilized in this study. All mice were maintained in the Rangos Research Center Animal Facility, Children's Hospital of University of Pittsburgh, with a 12-h light:12-h dark cycle and free access to standard laboratory food and water. All procedures were carried out in accordance with the regulations, regarding the care and use of experimental animals, published by the National Institutes of Health and were approved by the Institutional Animal Use and Care Committee of the University of Pittsburgh.
Preparation of bone component implants
In the current study, four different types of implants derived from morselized bone and phosphate-buffered saline (PBS) solution were implanted into the WT and TLR4−/− mice. The four implants included (1) BC group (morselized bone components), (2) BC+TLR4 inhibitor group and BC+TLR4 inhibitor peptide control group (morselized bone component mixing with TLR4 peptide inhibitor or with its peptide control), (3) CE group (fractionated cell-enriched portion of morselized bone component), and (4) ME group (fractionated matrix-enriched portion of morselized bone component).
To generate implants, two femora and two tibiae were harvested from one donor WT mouse and were crushed with a mortar and pestle under sterile conditions, while bone marrow was not removed before crushing. The BC group was generated by resuspending the morselized bone in 1 mL of PBS. The BC+TLR4 inhibitor group was generated by resuspending the morselized bone in 1 mL of VIPER TLR4 neutralizing peptide (0.3 mg/mL; Imgenex). To fractionate the BC group into its CE and ME groups, morselized bone resuspended in 1 mL of PBS was flushed through a 70-μm cell strainer (BD Biosciences) and centrifuged at 1200 rpm for 10 min. The resulting cellular pellets were resuspended in 1 mL of PBS to obtain the CE group. The remaining bone cortices were resuspended in 1 mL of PBS to derive the ME group. Cell counts of the CE group per animal showed ∼6–8×105 nucleated cells per mL. The density of the ME group was 200–300 mg/mL. Fractionated bone suspensions (300 μL) were mixed with 100 μL fibrinogen (10 mg/mL) and 2 μL thrombin (Sigma-Aldrich) and were incubated at 37°C for 1 h before being implanted in the calvarial defect model (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). Sham controls received no graft following surgery, and vehicle controls were prepared in the absence of bone components.
As bone marrow was not removed before crushing the donor bones, the CE suspension was considered to be rich in bone marrow lineage cells, including hematopoietic cells, myelopoietic cells, erythropoietic cells, and mesenchymal stem cells, while the ME suspension included mesenchymal stem cells, bone, adipose, and muscle residency cells.23–26 To further characterize the CE and ME suspensions, Syto® green-fluorescent nucleic acid staining (Life Technologies), propidium iodide staining (PI staining; Sigma-Aldrich), PicoGreen® dsDNA Reagent and Kits (Life Technologies), and the near-infrared fluorescent probe image system (far-red fluorescent pamidronate, FRFP680, Osteosense; VisEn Medical) were applied according to the manufacturer's instructions.
CE and ME suspensions from three donor WT mice were separately diluted with PBS to 1 million cells per mL and washed with sterilized PBS thrice before following with staining. Suspensions were incubated with 5 μM Syto green dye and 3 μM PI dye in the dark at 37°C for half an hour before being observed under fluorescent microscopy. All specimens were imaged at 100× and 200× magnifications. FRFP680 is a fluorescently labeled bisphosphonate that binds to mineral substrates and has been utilized as a tool for monitoring bone remodeling and skeletal calcification in vivo. 27 To demonstrate the extent of the matrix portion in CE and ME portions, three WT mice, between 10 and 12 weeks of age, were administrated FRFP680 at 10 nmol/100 μL by tail vein injection. Baseline tissue autofluorescence was examined in two WT mice with control saline injection. Mice were euthanized 24 h after FRFP680 administration. Femora and tibiae were harvested and imaged by the IVIS imaging system (IVIS® lumina; Xenogen). CE and ME suspensions were later collected as previously described and imaged by the IVIS imaging system for fluorescence efficiency. To characterize the extent of dsDNA concentration in CE and ME portions, CE and ME solutions from three donor mice were suspended in TE buffer and stained with PicoGreen dsDNA reagent at room temperature in the dark for 10 min. Fluorescence intensity was later measured using the Synergy H1 Hybrid microplate reader (BioTek).
Calvarial defect model
A circular parietal defect was made using a 1.8-mm outer diameter trephine (Fine Science Tools) as previously described 21 to model craniofacial bone defects that result from trauma, craniofacial reconstruction, or management of tumors. Implants were created using a 5-mm outer diameter trephine (Fine Science Tools). Circular implants in the groups outlined above were placed on top of the 1.8-mm calvarial defect. After implant placement, the scalp was closed with 4-0 Vicryl resorbable sutures. During surgery, sterile PBS was used for continuous irrigation at the defect site, and 1 mg/kg ketoprofen (Fort Dodge Animal Health) was administered immediately postsurgery as an analgesic.
Live microcomputed tomographic and radiographic analyses
For characterization of the three-dimensional (3D) calvarial defect healing process, bone healing was analyzed using a high-resolution microcomputed tomographic (μCT) system (Inveon microCT; Siemens). At postoperative days 0, 7, 14, and 28, BC-implanted WT and TLR4−/− groups and the BC+TLR4 inhibitor-implanted WT group (average seven each) were scanned using live μCT with a fixed isotropic voxel size of 63.8 μm. 3D images were reconstructed using Osirix software, and region of interest (ROI) was defined as 4.0 mm2×2.09 mm. Qualitative and quantitative data were analyzed by Osirix software with a global fixed threshold of −330. Standard μCT measurements (bone volume fraction, BV/TV within the ROI; regenerated BV: BV-day 0 BV) were calculated for each sample using Osirix software. CE- and ME-implanted WT and TLR4−/− mice were also sacrificed on postoperative day 28. Calvariae were harvested and fixed in 4% paraformaldehyde for 24 h. Radiographs were obtained using Faxitron MX-20 (Faxitron X-Ray) with a 35 kV exposure and a 45-s exposure time to analyze calvarial healing.
Histological analyses
Hematoxylin and eosin (H&E) and pentachrome staining were performed as described in our previous study. 21 Calvarial samples obtained from each group were harvested, fixed in 4% paraformaldehyde, decalcified in EDTA, dehydrated in a series of graded ethanol (EtOH) solutions, and subsequently embedded in paraffin. Five- to six-micrometer-thick sections were obtained from paraffin-embedded specimens by sectioning through the coronal plane. Three regions of each circular defect, 50 μm apart, were cut and placed on slides (for a total of ∼30 slides per animal). Slides were stained with Harris' H&E (Surgipath Medical Industries) and Russell-movat pentachrome staining (American MasterTech) for conventional, qualitative bright-field light microscopy. All specimens were imaged at 50×, 100×, 200×, and 400× magnifications.
Immunohistochemical staining was performed as previously described. 21 Sections were deparaffinized with xylene solution and rehydrated through serially graded EtOH solutions. Antibodies were diluted in 2% normal horse serum (Vector Laboratories) and incubated with sections according to the manufacturer's instructions. Slides were incubated in goat polyclonal anti-F4/80 (1:250 dilution; Santa Cruz Biotechnology), goat polyclonal anti-osteopontin (anti-OPN, 1:250 dilution; Santa Cruz Biotechnology), and rabbit polyclonal anti-cathepsin K (1:250 dilution; Abcam) for 30 min at room temperature. Slides were then incubated in biotinylated horse anti-goat secondary antibody (BA-9500, 1:250 dilution; Vector Laboratories, Inc.) or biotinylated goat anti-rabbit secondary antibody (1:250 dilution; Abcam), followed by Streptavidin-HRP (1:500 dilution; R&D systems). Color was developed by application of the DAB kit (Vector Laboratories). Sections were dehydrated and mounted before examination at 50×, 100×, 200×, and 400× magnifications.
Histomorphometric analysis was used to quantify two-dimensional bone formation using three to five slides per animal. New areas of bone formation were visually identified under 100× magnifications and measured using Northern Eclipse software (Empix Imagine, Inc.). Regions of new bone formation included areas at the defect as well as on both endo- and ectocranial surfaces of the calvaria within ROI (Supplementary Fig. S2). The sum totals of newly formed bone areas were averaged by the number of slides per animal.
Anti-cathepsin K staining was used for visualizing the osteoclast infiltration. Cathepsin K-positive cells attaching the bone surface were counted under 200×. Numbers of osteoclasts per total bone surface area (N.OC/BS; n/mm) were calculated. Data are expressed as mean area±SEM.
Statistical analysis
Statistical analyses were performed using SPSS v. 20.0 software (SPSS, Inc.). Mean areas of newly formed bone by time points, numbers of osteoblasts per bone surface area, and numbers of osteoclasts per bone surface area were compared among groups using one-way ANOVA, followed by post hoc least significant difference test. A p-value less than or equal to 0.05 was considered significant.
Results
Characterization of BC, CE, and ME implants
Characterization of CE and ME suspensions indicated good separation of grounded bone graft (BC group) into CE and ME portions (Supplementary Fig. S3). Both CE and ME suspensions contained similar percentages of live cells with around 44.5% live cells in CE suspension and 46.6% live cells in ME suspension. The CE suspension contained 22 million cells per mL, while the ME suspension contained 0.32 million cells per mL (Supplementary Fig. S3A). IVIS imaging results showed that majority of FRFP fluorescent labeling was detected in the ME portion. Fluorescent efficiency in the CE portion was similar compared with the baseline control group without FRFP administration (Supplementary Fig. S3B). Picogreen analysis demonstrated that there were, on average, 38,038.30±817.95 ng/mL dsDNA in the CE portion and, on average, 5590.11±2114.96 ng/mL dsDNA in the ME portion (Supplementary Fig. S3C).
Delayed calvarial healing in the absence of TLR4 signaling after bone graft implantation
To understand the role of TLR4 signaling in bone graft-induced calvarial defect healing, BC (morselized bone component) implants, with and without a TLR4 inhibitor, were placed within calvarial defects of WT mice. BC was implanted in TLR4−/− mice as well. Vehicle controls were included for both WT and TLR4−/− mouse strains. Representative 3D reconstruction images of μCT of three implant groups are shown (Fig. 1A). Qualitative and quantitative analyses of the calvarial defects revealed more mineralized tissue in the BC-implanted WT group than either the BC-implanted TLR4−/− group or the BC+TLR4 inhibitor-implanted WT group. Larger areas of mineralized tissue were visible within the defect and adjacent to the implants in the BC-implanted WT group at designated time points compared with the other two groups. Morselized bone graft, however, remained relatively unchanged in TLR4−/− mice or in WT mice when mixed with TLR4 inhibitor (Fig. 1A).
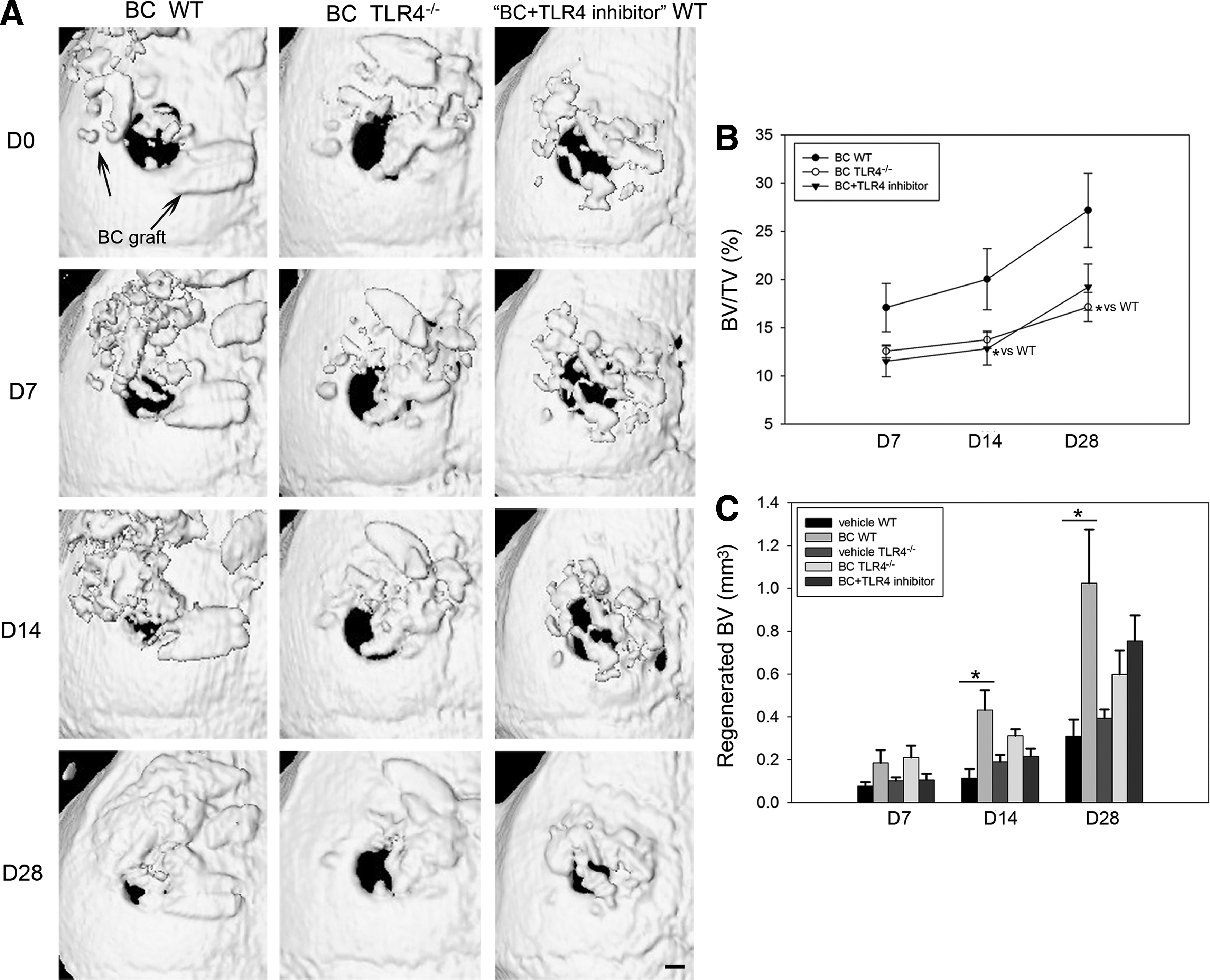
Live μCT analysis of WT and TLR4−/− graft recipients. WT recipients were engrafted with BC in the presence or absence of TLR4 inhibitor, and TLR4−/− recipients were engrafted with BC only. Calvarial bone repair was measured over 28 days using μCT.
Measurements from μCT analysis revealed that bone volume fraction was significantly larger in the BC WT group (BV/TV: 28.46%±6.88%) than in the BC TLR4−/− group (BV/TV: 17.15%±1.47%, p<0.05) on day 28. No significant difference in bone healing was observed between the BC+TLR4 inhibitor WT group and the BC TLR4−/− group on postoperative days 7, 14, or 28 (Fig. 1B). The BC WT group showed a significantly larger volume of regenerated mineralized tissue compared with the WT vehicle control group at postoperative days 14 and 28. No significant differences in regenerated bone volume were observed between BC TLR4−/−, BC+TLR4 inhibitor WT, and TLR4−/− vehicle control groups (Fig. 1C). No significant difference in bone volume fraction was observed between the BC WT group and BC+TLR4 inhibitor control peptide group (Supplementary Fig. S4).
Accelerated intramembranous bone formation and active graft remodeling in WT, but not in TLR4−/− mice, after BC implantation
Histological characterization revealed early evidence of intramembranous bone formation accompanied by greater osteoblast and macrophage infiltration and more intense periosteal and dural cellularity in WT mice compared with TLR4−/− mice after BC implantation (Figs. 2 and 3). On day 7, BC WT mice exhibited more pronounced evidence of newly formed, highly cellularized woven bone along the endocortical (dural) side of the calvarial bone and along the defect perimeter than observed in control WT mice. Newly regenerated woven bone was visible in the TLR4−/− control group, while BC TLR4−/− mice showed little to no obvious mineralization on day 7 (Fig. 2A). Histomorphometric analysis on day 7 showed significantly more bone formation in the BC WT group compared with the WT sham control group and the BC TLR4−/− group and also the WT control group compared with the TLR4−/− sham group. No significant difference was found between the BC TLR4−/− group and the TLR4−/− sham control group (Fig. 2B).
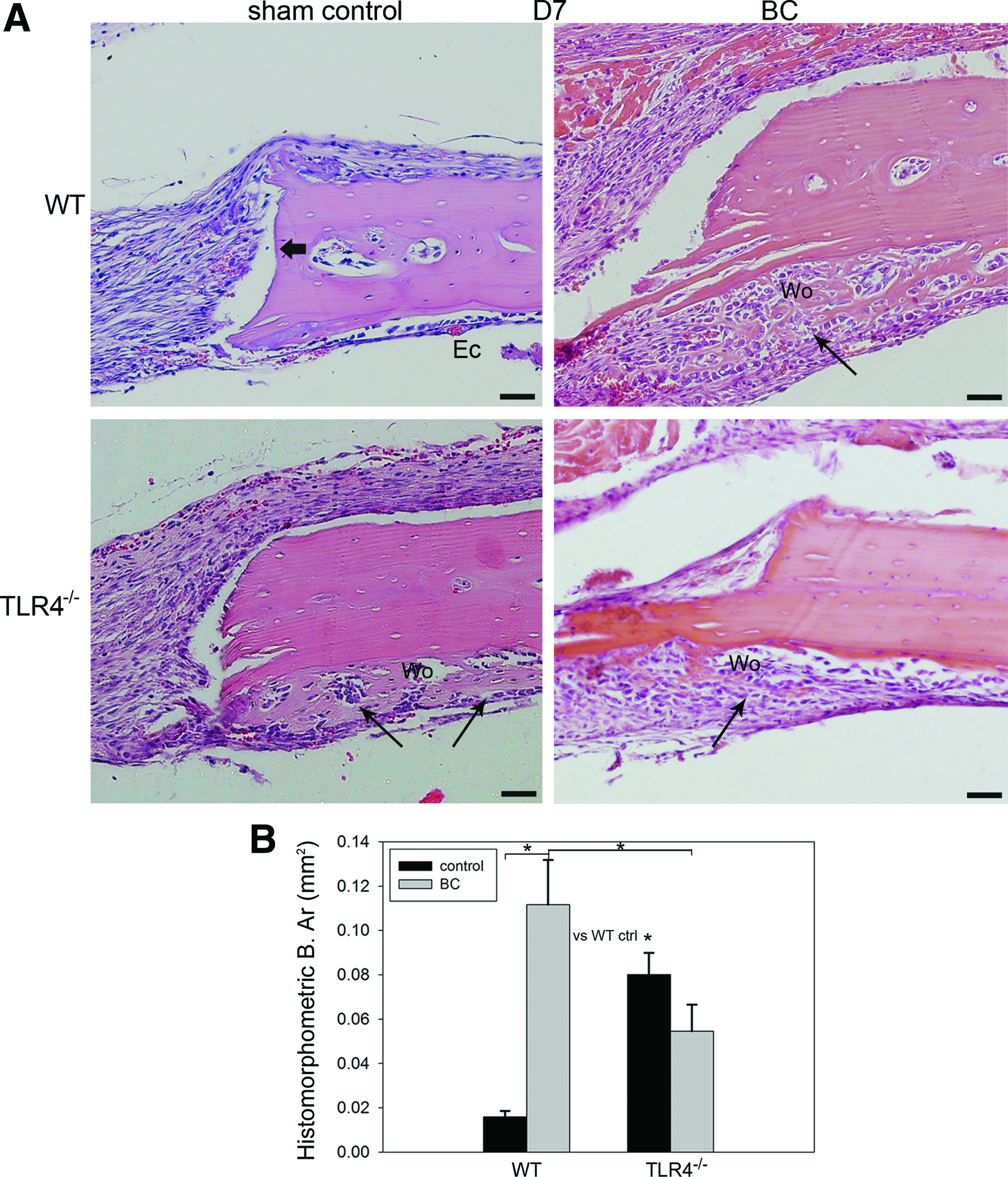
Histology and histomorphometric analysis of calvarial bone repair in WT and TLR4−/− graft recipients at postoperative day 7.
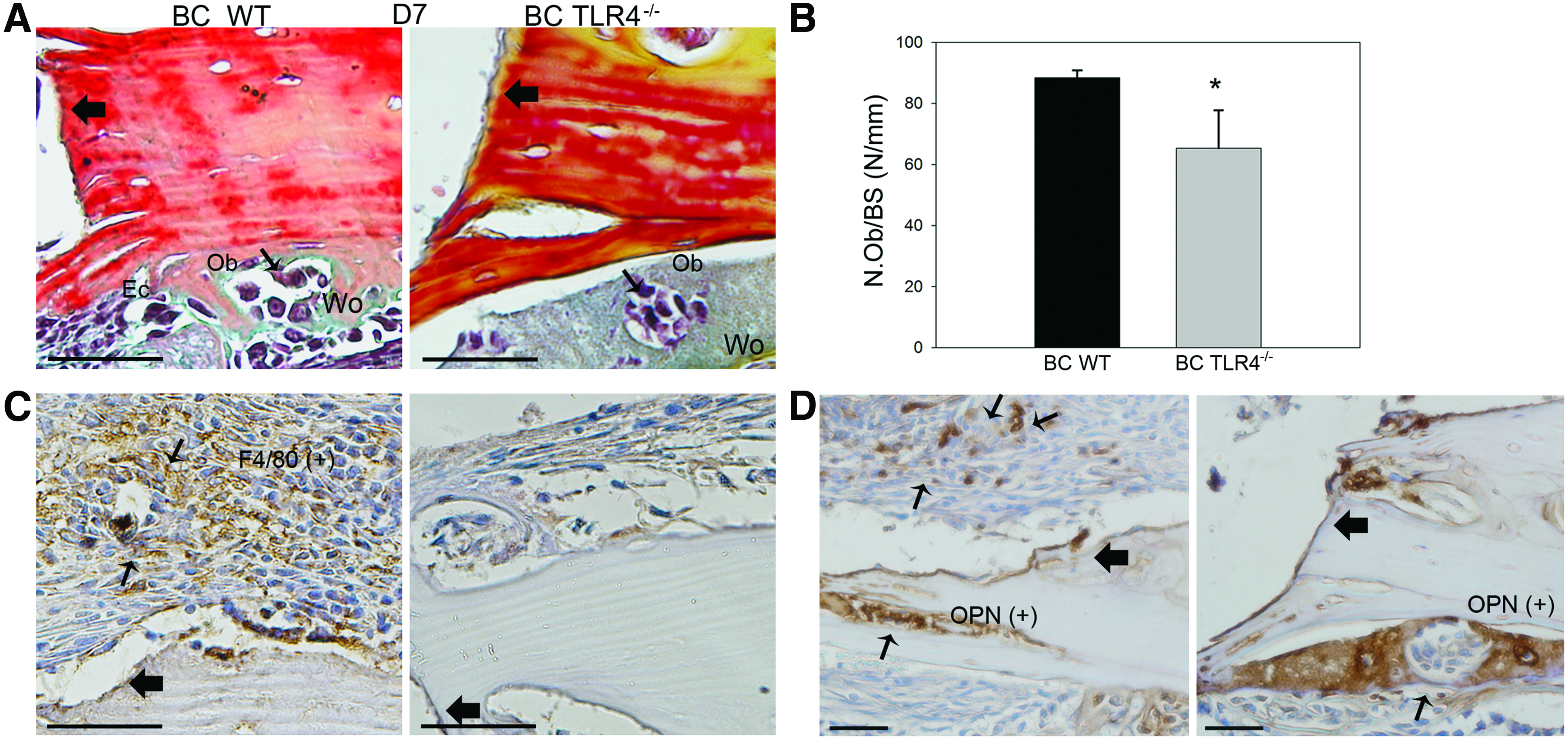
Cell population dynamics in WT and TLR4−/− graft recipients. BC-implanted WT mice had more osteoblast and macrophage infiltration versus TLR4−/−mice.
To detect osteoblast and macrophage lineage cell infiltration, pentachrome staining, OPN, and F4/80 immunohistochemistry staining were performed on day 7 slides in BC WT and BC TLR4−/− mouse groups. On day 7, more bone surface lining mononucleated osteoblasts and woven bone were evident in the BC WT group than in the BC TLR4−/− group (Fig. 3A). These observations were consistent with the osteoblast count based on pentachrome staining analysis (Fig. 3A, B). Positive F4/80 macrophage lineage cells were observed in the BC WT group, especially in the regenerated periosteal region, whereas no obvious F4/80-positive cells were evident in the BC TLR4−/− group on day 7 (Fig. 3C). Intense OPN staining was observed in both the BC-implanted WT and TLR4−/− groups (Fig. 3D).
On day 28, remodeled woven bone matrix and thickened periosteum were observed in all three experimental groups. In addition, Howship's lacunae at the edge of the graft implants and bony union of grafts and recipients were observed in the WT group. In contrast, fibrous encapsulation of morselized bone components was observed in TLR4−/− graft recipients (Fig. 4A). Histomorphometric measurements of newly regenerated bone area indicated no significant difference among the three groups (Fig. 4B; BC WT group: 0.14±0.01 mm2; BC TLR4−/− group: 0.19±0.04 mm2; BC+TLR4 inhibitor WT group: 0.15±0.02 mm2). More intense OPN staining showing evidence of bone resorption near scalloped edges of the BC grafts (Fig. 5A), and significantly more intense cathepsin K-stained osteoclasts were observed in the WT group than in the TLR4−/− group (Fig. 5B, C).
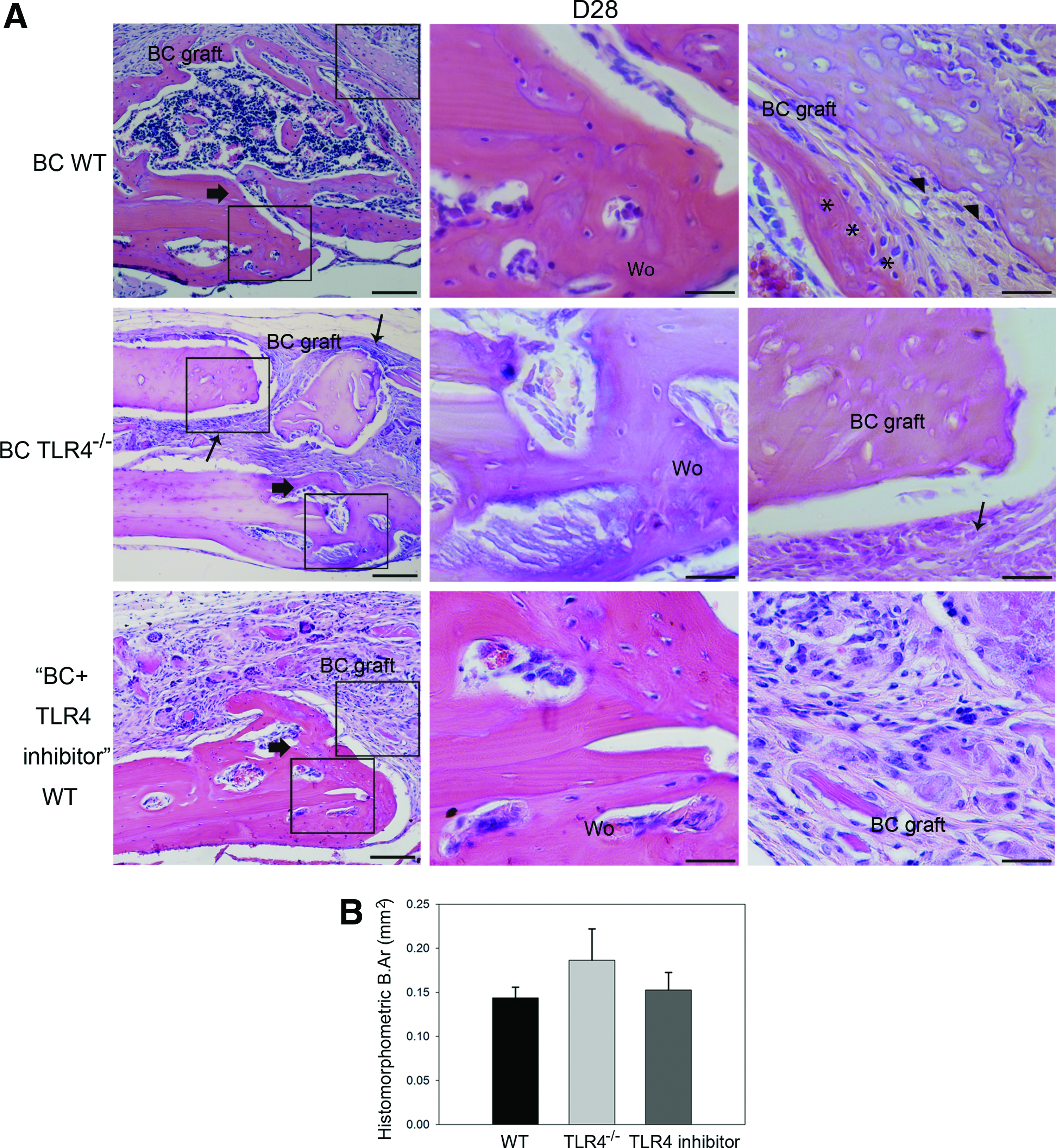
Histology and histomorphometric analysis of calvarial bone repair in WT and TLR4−/− graft recipients at postoperative day 28.
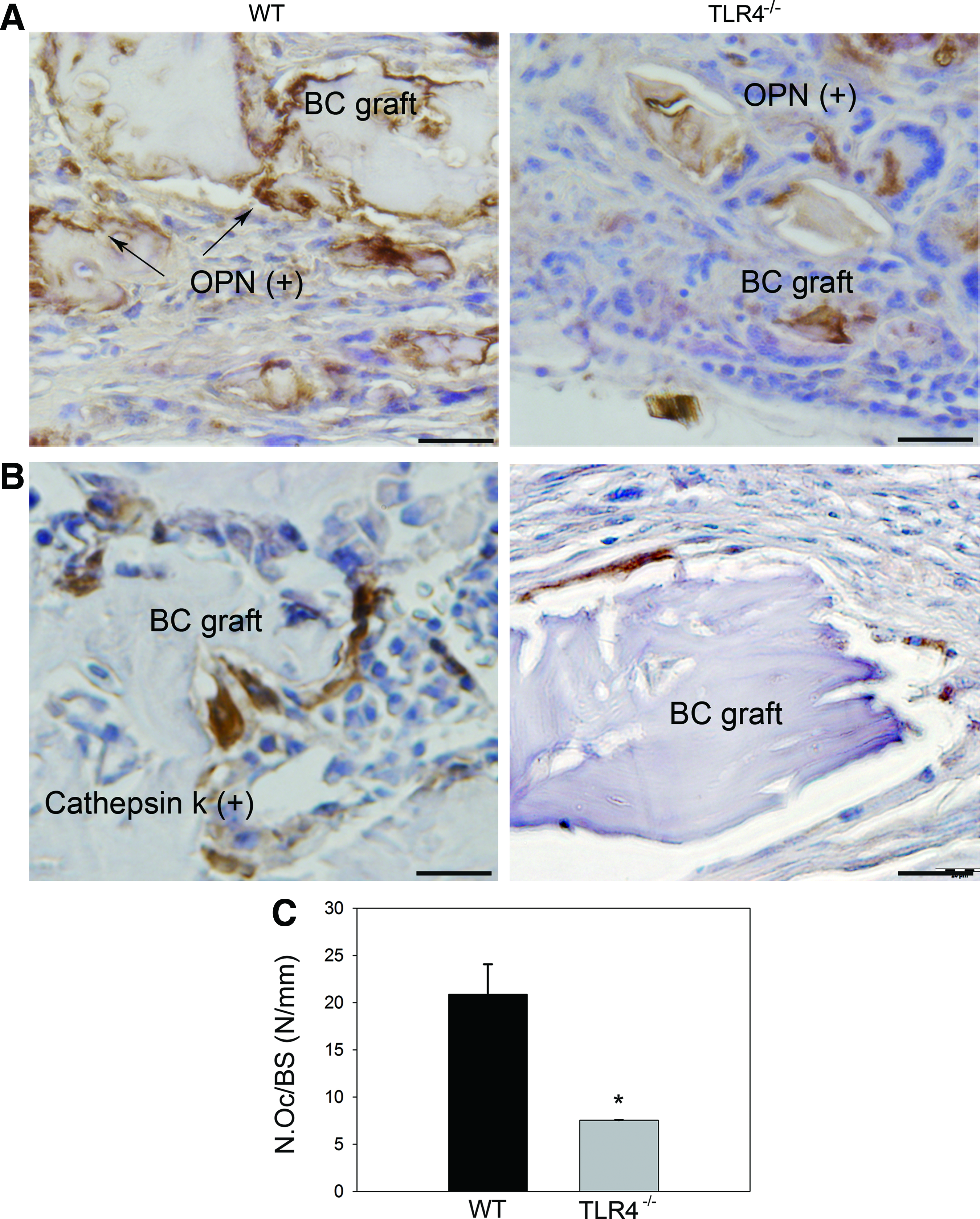
Representative images showing OPN and cathepsin K-stained slides of BC-implanted WT and TLR4−/− mice on day 28. More intense OPN staining
The matrix-enriched fraction of morselized bone enhanced calvarial healing in WT mice, not in TLR4−/− mice
To determine the impact of bone graft component on calvarial repair, cell-enriched (CE) and matrix-enriched (ME) bone fractions were implanted in the calvarial defects in WT mice. On day 28, an overall more intense reaction of periosteum was shown in the ME group compared with the CE group. Remodeling bone matrix was evident in the soft tissue covering the defect in the ME group (Fig. 6A). To determine if TLR4 signaling plays a role in matrix-induced calvarial defect healing, the CE and ME implants were similarly placed in calvarial defects in TLR4−/− mice. Histological characterization revealed that CE and ME implant groups presented a similar healing pattern in TLR4−/− mice on postoperative day 28. The majority of woven bone formation was evident on the endocortical side of the calvarial bone and adjacent to the original defect in both groups in TLR4−/− mice (Fig. 6B).
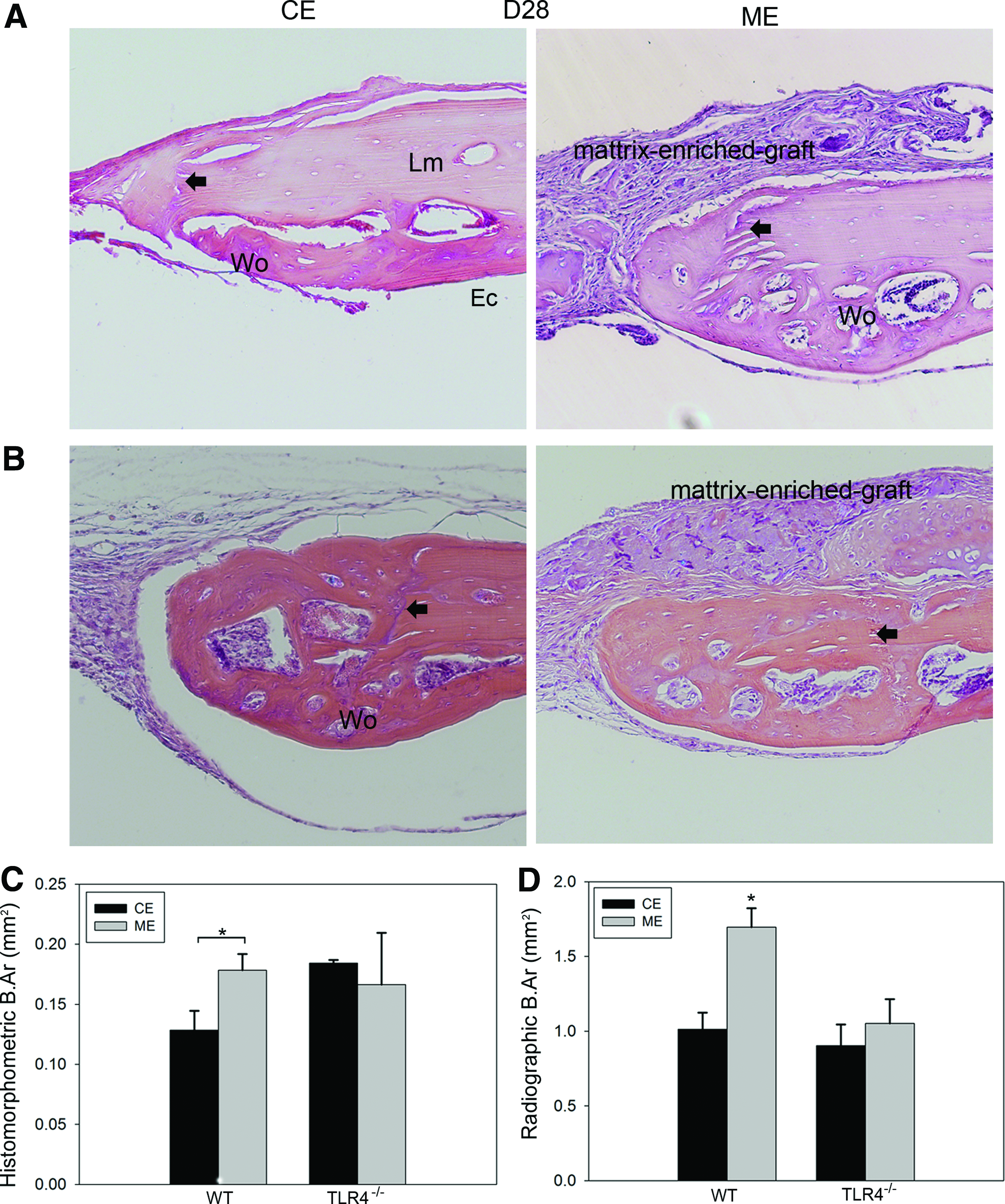
Histology and histomorphometric analysis of WT and TLR4−/− mice engrafted with fractionated bone components at day 28. Representative images showing H&E-stained slides of CE- and ME-implanted WT
Histomorphometric measurements showed significantly more newly formed bone in the ME group (0.18±0.01 mm2) compared with CE (0.13±0.02 mm2, p<0.05) and vehicle control (0.12±0.00 mm2, p<0.05, data not shown) groups in WT mice on postoperative day 28. No significant difference was shown between CE (0.18±0.00 mm2) and ME (0.17±0.04 mm2) groups in TLR4−/− mice on day 28 (Fig. 6C). Calculation of newly formed bone area based on radiographic images (data not shown) on postoperative day 28 revealed significantly more new bone formation in the ME group (1.70±0.13 mm2; 66.79% healing) than in the CE group (1.01±0.11 mm2, 39.91% healing, p<0.05) and the vehicle control group (0.65±0.12 mm2, 25.59% healing, p<0.001, data not shown) in WT mice. CE- and ME-implanted TLR4−/− mice exhibit comparable bone healing (CE: 0.90±0.14 mm2; 35.43% healing; ME: 1.05±0.16 mm2, 41.34% healing; Fig. 6D).
Discussion
Although various engineered materials, including ceramics and metal implants, have been developed for bone repair, autologous and allogeneic bone implants remain the graft materials of choice for bone reconstruction. The successful integration of implant material with the host requires the implanted bone graft to undergo remodeling and the induction of the healing cascade. The TLRs play a unique role in tissue repair by helping to regulate the innate inflammatory response to damage. In this study, our data highlight a critical role for TLR4 signaling in graft-mediated calvarial repair.
The impact of inflammation on healing is complex and may be either beneficial or detrimental to repair.28,29 Recent advances in our understanding of the innate immune response have helped to distinguish between tissue-protective and tissue-destructive inflammatory signaling pathways. 30 In a previous study, accelerated bone healing within a calvarial defect model was observed in TLR4−/− mice, suggesting that TLR4 signaling is detrimental to the normal healing of calvarial bone defects. 21 In contrast to our previous findings, we report here that TLR4 is required to support graft-induced bone repair using a calvarial defect model. At day 7, after implantation, WT mice showed more F4/80-positive macrophage lineage cell infiltration, while macrophages were not detected in TLR4−/− mice after bone component implantation. Inflammation promotes cell proliferation and migration into the fracture site to trigger the repair and healing response within damaged bone.9,30 In addition, we showed that surgical implantation of morselized bone into damaged soft tissue induced a deleterious, TLR4-dependent, systemic inflammatory response. 22 Together, these data suggest that TLR4 functions differently in our two models of craniofacial bone injury and repair. Specifically, TLR4 function delays normal bone healing, but is essential for graft-induced bone formation. The results we present here suggest that certain molecular signals within bone graft require TLR4 signaling to induce bone healing. Whether allograft or synthetic graft material-assisted healing also needs TLR4 signaling, or manipulating TLR4-induced inflammatory response will assist in allograft or biomaterial-mediated bone healing, requires better investigation.
BC grafts were obtained from different mice of the same strain (WT, C57BL-6J). Thus, grafts were considered to be allogeneic in source.31–34 Allogeneic bone is frequently employed as a decellularized graft material, but is generally inferior to the autologous bone graft and exhibits inferior remodeling upon implantation. Coating of structural allografts with RANKL and VEGF resulted in marked remodeling, improved vascularization, and enhanced bone repair, however, suggesting an essential role for remodeling in forming bony unions between grafts and host bone tissue. 35 Our results support and extend this finding by suggesting that the modulator of the innate immune response (TLR4) is required for graft remodeling and for subsequent bone healing. In our study, engraftment of WT mice with BC grafts resulted in earlier mineralization, more pronounced infiltration of regenerating tissue with osteoblasts, and more F4/80-stained macrophage infiltration relative to TLR4−/− recipients at day 7 (Figs. 2 and 3). At day 28, implanted grafts in WT recipients showed irregular reversal lines and evidence of remodeling, whereas grafts in TLR4−/− recipients showed relatively smooth margins and no obvious signs of remodeling (Figs. 4 and. 5). Qualitative and quantitative μCT analyses revealed graft-enhanced repair in WT mice, but not in TLR4−/− mice. Moreover, inhibition of TLR4 suppressed healing in WT graft recipients, resulting in healing rates similar to those observed in TLR4−/− graft recipients between days 7 and 14. Healing was accelerated in the inhibitor group at later time points, presumably due to a decline in the local concentration of the inhibitor and restoration of TLR4 signaling over time. Because there was little or no evidence of graft remodeling (normally caused by osteoclastic destruction of the bone graft surface) in mice lacking the TLR4 gene, it is likely that TLR4 signaling is needed to induce graft remodeling.
We also sought to determine which fraction of bone graft triggers TLR4-dependent bone healing. It has been argued that the regenerative potential of bone graft is dependent upon the efficient transfer of progenitor cells to the defect site,36,37 although donor cells typically fail to survive beyond hours to weeks.38,39 In our previous work, 21 we used radiographic bone measurement to assess bone repair in a calvarial defect model and observed that 33.14% of the defect had regenerated by day 28 in WT mice, while the current study showed that BC and ME (matrix-enriched) groups achieved 74.80% and 66.79%, respectively (Fig. 6). The CE (cell-enriched) implant, in contrast, failed to stimulate bone repair beyond control values. Furthermore, comparable levels of healing were observed in both ME- and CE-engrafted TLR4−/− mice (Fig. 6). These combined observations suggest that the matrix-bearing fraction of morselized bone bears primary responsibility for graft-induced bone regeneration and this bone healing effect is also partially TLR4 dependent. Disparities between our findings and those reported in other studies40,41 may be due to the differences in graft fractionation procedures.
Several points of considerations should be noted in this study, including our use of a larger graft and approach for measuring bone formation. First, the graft size was designed to be larger than the defect size to facilitate stable placement of grafts on the top of the calvarial defect site, thereby ensuring consistency among all recipient mice. Second, we employ separate approaches for measuring bone formation (μCT and histomorphometric analyses). In our μCT analysis, we measured all mineralized tissue within the ROI, thus including both regenerated host new bone and remodeled bone graft. However, in our histomorphometric analysis, only newly regenerated host bone was measured as some bone grafts remained as tiny pieces, which made it challenging to accurately measure remodeled bone grafts (Supplementary Fig. S2). As a result, a slight discrepancy exists between our μCT and histomorphometric analysis (Figs. 1B and 6C). This discrepancy also explains the lack of significant differences among BC WT, BC TLR4−/−, and BC+TLR4 inhibitor WT groups in histomorphometric measurements (Fig. 4). However, this slight discrepancy in measurement does not alter the key finding of the study.
In summary, our results suggest that the healing effect of bone graft was inhibited without TLR4 signaling using a TLR4 gene knockout mouse model or the local delivery of TLR4 peptide inhibitor. These observations together establish an important and novel role of TLR4 in the healing effect driven by the bone graft, especially the matrix-enriched fraction. Future work will seek to identify the molecular signals released from bone graft or the ECM component that is recognized by TLR4 and capable of inducing bone remodeling and bone formation.
Footnotes
Acknowledgments
The authors are especially grateful to Dr. David Hackam (Department of Plastic Surgery, University of Pittsburgh), Sid Angle (Carnegie Mellon University), Gwen Taylor, Chris Hobson, Laurie Meszaros, Zoe Maclsaac (University of Pittsburgh), and Daifei Elmer Ker (Stanford University) for their assistance in this work. The authors thank the National Science Foundation for Young Scientists of China, No. 81201410, for the support of this study.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.