
Abstract
Chondrogenic priming (CP) of mesenchymal stem cells (MSCs) and coculture of MSCs with human umbilical vein endothelial stem cells (HUVECs) both have been shown to significantly increase the potential for MSCs to undergo osteogenic differentiation and mineralization in vitro and in vivo. Such strategies mimic cartilage template formation or vascularization that occur during endochondral ossification during early fetal development. However, although both chondrogenesis and vascularization are crucial precursors for bone formation by endochondral ossification, no in vitro bone tissue regeneration strategy has sought to incorporate both events simultaneously. The objective of this study is to develop an in vitro bone regeneration strategy that mimics critical aspects of the endochondral ossification process, specifically (1) the formation of a cartilage template and (2) subsequent vascularization of this template. We initially prime the MSCs with chondrogenic growth factors, to ensure the production of a cartilage template, and subsequently implement a coculture strategy involving MSC and HUVECs. Three experimental groups were compared; (1) CP for 21 days with no addition of cells; (2) CP for 21 days followed by coculture of HUVECs (250,000 cells); (3) CP for 21 days followed by coculture of HUVECs and MSCs (250,000 cells) at a ratio of 1:1. Each group was cultured for a further 21 days in osteogenic media after the initial CP period. Biochemical (DNA, Alkaline Phosphatase Activity, Calcium, and Vessel Endothelial Growth Factor) and histological analyses (Alcian blue, alizarin red, CD31+, and collagen type X) were performed 1, 2, and 3 weeks after the media switch. The results of this study show that CP provides a cartilage-like template that provides a suitable platform for HUVEC and MSC cells to attach, proliferate, and infiltrate for up to 3 weeks. More importantly we show that the use of the coculture methodology, rudimentary vessels are formed within this cartilage template and enhanced the mineralization potential of MSCs. Taken together these results indicate for the first time that the application of both chondrogenic and vascular priming of MSCs enhances the mineralization potential of MSCs in vitro while also allowing the formation of immature vessels.
Introduction
B
Recently, stem cell research has opened up the possibility of using bone marrow mesenchymal stem cells (MSCs) to regenerate bone tissue in vitro, which can be implanted to replace damaged bone tissue2–4 and might provide an effective alternative to treat bone defects arising due to disease or injury. Previous studies have found that MSCs can produce a bone-like matrix when seeded on ceramic scaffolds and implanted in rodents.5,6 However, when MSC-seeded ceramic scaffolds were implanted in a human jaw defect only one construct out of six showed bone matrix formation. 7 Others have investigated the use of cell-seeded collagenous scaffolds, but it has been shown that MSCs seeded onto collagen sulphate glycosaminoglycan (sGAG) scaffolds can act as a barrier to healing in rodent cranial defects. 8 Once implanted, the scaffolds become encapsulated and host vasculature is inhibited, which leads to core necrosis and ultimately failure of the implant construct. 8 To date no approach has been able to fully regenerate bone tissue in vitro that can be clinically used to replace degenerated bone in load bearing locations. 9
Recent studies have suggested that in vitro approaches, which mimic certain aspects of the endochondral ossification process, can enhance bone tissue regeneration both in vitro and in vivo. Chondrogenic priming (CP) of MSCs in vitro, before implantation in vivo, has been shown to overcome issues with poor oxygen and nutrient supply in tissue engineered constructs,10–12 and in particular promoted healing in an osteochondral defect model. 11 Other studies have investigated the implantation of chondrogenically primed MSC in both aggregate and scaffold form to heal a large segmental defect.13,14 These studies found that the implantation of MSCs, which were predifferentiated toward the chondrogenic lineage, lead to bone regeneration through endochondral ossification within the defect site when compared to undifferentiated MSCs. However, none of these studies investigated prevascularization of the constructs and both studies only examined rat MSCs rather than human MSCs. Other studies have also shown that chondrogenically primed constructs seeded with human MSCs subsequently mineralized following subcutaneous implantation.10,13–16 Therefore, in vivo, chondrogenically primed human MSCs can form bone through endochondral ossification. However, each of these studies observed core degradation and an uneven distribution of bone mineral throughout the construct.10,12,16 In a recent study we found that CP of Bagg Albino (BALBc) mice MSCs and human MSCs in vitro for specific durations (14, 21 days) can have optimum influence on their mineralization capacity and can produce a construct that is mineralized throughout the core. 17
Another method to overcoming the core degradation challenges of current bone tissue engineering strategies is through the formation of vasculature within the biomaterial-based bone tissue constructs. In vivo vessel formation is an integral part of the bone formation process that occurs during early fetal development, known as endochondral ossification. During endochondral ossification MSCs condense to become clusters of cells,18–20 differentiate to become chondroblast cells, and ultimately form a cartilage template. Vessel invasion occurs once the cartilage template has formed by a process known as quiescent angiogenesis, which involves endothelial cells invading through the cartilage canals already present in the developing bone tissue,18–21 and this process typically occurs between 14 and 18 days of embryogenesis.21,22 For in vitro bone regeneration approaches it is believed that bone vasculature is necessary to provide appropriate growth factors, hormones, chemokines, and nutrients, and without a vascular supply, cells within tissue-engineered constructs do not get the necessary requirements to regenerate the damaged bone tissue and ultimately die when implanted in vivo.23,24
Coculture studies of MSCs with endothelial stem cells have been shown to upregulate the osteogenic potential of the MSCs in both 2D and 3D culture in vitro.25–30 Other studies have investigated whether prevascularizing tissue engineering constructs in vitro would allow faster host integration postimplantation.31–36 Prevascular networks can be formed in vivo as early as 7 days in coculture of HUVECs31–35 and MSCs,34,36 however, they are only sustained in the presence of the MSCs.34,36 Other studies have looked into direct cell–cell contact coculture approaches through the formation of cellular aggregates,25,26,37–39 and prevascular networks have been observed in certain cellular aggregates. Moreover, the presence of both MSCs and HUVECs promotes the formation of vascular networks and upregulates early osteogenic markers like alkaline phosphatase (ALP) activity.25,38
While in vitro bone regeneration strategies have sought to incorporate either the production of the cartilage template or the vascularization of the construct, no strategy has sought to incorporate both events simultaneously, although both are crucial precursors for bone formation in vivo during endochondral ossification. In this study we test the hypothesis that a tissue regeneration approach that incorporates both CP of MSCs, to first form a cartilage template, and subsequent prevascularization of the cartilage constructs, through the coculture of HUVECs in vitro, will serve as an effective in vitro bone regeneration approach. The specific objective of this study is to compare the regenerative potential of (1) CP of MSCs in pellet culture and (2) addition of HUVECs to chondrogenic MSC pellets, to (3) a novel methodology involving both CP and the coculture of HUVECs and MSCs. The regenerative potential is assessed by means of biochemical and histological analysis for DNA content, sGAG production, ALP Production, calcium content, vascular endothelial growth factor (VEGF) production, CD31+, and collagen type X content.
Methods
Cell isolation and characterization
Human donor MSC: isolation and characterization
Human bone marrow-derived MSCs were extracted from bone marrow aspirates. The bone marrow aspirates were obtained from the iliac crest of normal human donors under ethical approval and informed consent, as approved by the Research Ethics Committee of the National University of Ireland Galway (NUI Galway, Ireland) and the Clinical Research Ethical Committee at University College Hospital, Galway, Ireland. Bone marrow aspirates were obtained from 4 donors; 45, 48, 56, and 59 years of age. The MSCs were isolated on the basis of plastic adherence from bone marrow aspirates as previously described. 16 The MSCs were expanded in standard tissue culture flasks using Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS; EU Thermo Scientific), 100 U/mL penicillin (Sigma Aldrich), 100 g/mL streptomycin (Sigma Aldrich), and 2 mM L-glutamine (Sigma Aldrich). After approximately 4 days, large colonies had formed, and the culture medium was changed and cultured until confluent. These cells were further cultured to passage 4. Once suitable colonies had formed the chondrogenic, osteogenic, and adipogenic potential of these MSCs were confirmed as outlined below.
To confirm adipogenic potential of the MSCs used, cells were plated at 2×104 cells/cm2 and incubated in expansion medium until they reached 80% confluency. Then, adipogenic induction medium was added containing DMEM with 10% FBS, 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, 1 μM dexamethasone, 10 μg/mL insulin, 200 μM indomethacin, and 500 μM isobutyl-1-methyl-xanthine (All Sigma Aldrich). After 3 days, maintenance medium was added containing DMEM, 10% FBS, 2 mM L-glutamine, 100 U/mL penicillin, 100 g/mL streptomycin, and 10 μg/mL insulin (All Sigma Aldrich). This cycle was repeated for a 21-day cell culture period, after which cells were fixed with 10% formalin and stained with 0.5% Oil Red O in methanol (All Sigma Aldrich). A negative control group was also stained for comparison, in which MSCs had been incubated in expansion medium for the same duration.
To confirm osteogenic differentiation cells were plated at 2×104 cells/cm2 and incubated in expansion medium until they reached 80% confluency. Osteogenic medium was added containing DMEM supplemented with 10% FBS, 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, 20 mM β-glycerol phosphate, 100 nM dexamethasone, and 50 μM ascorbic acid (All Sigma Aldrich). Media was changed every 3 days and cells were cultured for 21 days. After 21 days, cells were fixed in 10% formalin and stained with 10% alizarin red (All Sigma Aldrich). A negative control group was also stained for comparison, in which MSCs had been incubated in expansion medium for the same duration.
To confirm the chondrogenic potential of the MSCs, cells were seeded in pellet culture with 2.5×105 cells per pellet and incubated in 500 μL of complete chondrogenic medium, which consisted of a chemically defined medium containing high glucose DMEM GlutaMAXTM (Gibco, Life Sciences), 10 ng/mL TGF-β3 (ProSpec-Tany TechnoGene Ltd.), 50 μg/mL ascorbic acid, 4.7 μg/mL linoleic acid, 100 nM dexamethasone (All Sigma Aldrich), and 1x insulin-transferrin-selenium (ITS; BD Biosciences). The medium was changed three times a week. After 21 days, pellets were harvested, embedded in paraffin, sectioned, and stained with Alcian Blue (Sigma Aldrich). A negative control group was also stained for comparison, in which MSCs had been incubated in expansion medium for the same duration.
To obtain sufficient stocks of MSCs, human MSCs (24-year-old donor) were also purchased from Lonza, Braine-l'Alleud, Belgium and cultured in using DMEM supplemented with 10% fetal bovine serum (FBS; EU Thermo Scientific), 100 U/mL penicillin (Sigma Aldrich), 100 g/mL streptomycin (Sigma Aldrich), and 2 M L-glutamine (Sigma Aldrich). Media were replaced every 3 days and upon reaching 80–90% confluency, cells were passaged using trypsin-EDTA solution. MSCs were further cultured to passage 4.
HUVECs culture
Human umbilical vein endothelial cells (HUVECs) were purchased from PromoCell, Heidelberg Germany and cultured in endothelial growth medium (EGM) (Promocell). Media were replaced every 3 days and upon reaching 80–90% confluency, cells were passaged using trypsin-EDTA solution. HUVECs were further cultured to passage 4.
Pellet formation
Once the human MSCs reached a confluency of ∼90% the cells were trypsinized, counted, and centrifuged at 650 g, at a temperature of 22°C for 5 min. For this study all pellets were formed from individual donors, cells were not pooled. The cells were then resuspended in expansion media at a density of 0.25×106 cells/mL. This cell suspension was divided into 1.5 mL tubes so that there were 250,000 cells in each tube, and then centrifuged for 5 min (Eppendorf Centrifuge 5430R) at 400 g to create cell pellets. Carefully avoiding the newly formed pellet, the media were removed from each of the pellets and 0.5 mL of either chondrogenic media or endothelial growth media plus osteogenic growth factors was added depending on experimental conditions (described in detail below). Chondrogenic medium consisted of a chemically defined medium, which contained high glucose DMEM GlutaMAXTM (Gibco, Life Sciences), 10 ng/mL TGF-β3 (ProSpec-Tany TechnoGene Ltd.), 50 μg/mL ascorbic acid (Sigma Aldrich), 4.7 μg/mL linoleic acid (Sigma Aldrich), 100 nM dexamethasone (Sigma Aldrich), and 1x insulin-transferrin-selenium (ITS; BD Biosciences). EGM plus osteogenic growth factors consisted of EGM media (Promocell) supplemented with 8% FBS, 100 nM of dexamethasone, 50 μg/mL ascorbic acid, and 10 mM β-glycerol phosphate (Sigma Aldrich). For all experiments pellet cultures were fed twice per week by performing a 50% medium exchange. During each feed the pellets were agitated so as to prevent them from adhering to the micro-tube. This was achieved through aspirating the media beneath the pellet with a micro-pipette.
These cells were further cultured under the following conditions: (1) CP21–HUVECs: CP (Chemically Defined Medium plus TGF-β1, linoleic acid, ascorbic acid, and dexamethasone) for 21 days; (2) CP21+HUVECs: CP for 21 days and addition of HUVECs (250,000 cells); and (3) CP21+HUVECs:MSCs: CP for 21 days and coculture of HUVECs and MSCs at a ratio of 1:1 (125,000:125,000 cells).
For the coculture groups, confluent layers of HUVECs/MSCs were trypsinized and counted. Cells were suspended depending on experimental conditions so that there were 0.5×106 cells/mL. In the case of the CP21+HUVECs:MSCs, the ratio of cells was 1:1 HUVECs:MSCs. The cells were suspended in EGM media containing osteogenic growth factors and 20% methocel from a stock solution that was generated by dissolving 6 g of carboxymethylcellulose (Sigma Aldrich) in 500 mL of DMEM as previously described. 40 The medium was removed from the chondrogenically primed group and EGM media containing osteogenic growth factors was added. In the case of the group that contained HUVECs, the media added also contained suspended HUVECs alone and in the case of the group that contained both HUVECs and MSCs, the media added also contained suspended MSCs and HUVECs. After 24 h the medium that contained methocel was removed and was replaced with EGM media containing osteogenic growth factors and the pellets were cultured for a further 21 days.
Histochemical analysis and biochemical analysis
Pellets were examined at day 0, 1 week, 2 weeks, and 3 weeks after the start of culture in their respective conditions and were prepared for either histochemical analysis or biochemical analysis. At each of the time points, the culture medium from the pellets was collected, snap frozen, and stored at −80°C until biochemical assays could be performed. The remaining pellets were washed with PBS and then treated in one of the following two ways; (1) snap frozen and stored at −80°C for biochemical analysis or (2) fixed overnight in paraformaldehyde before being placed in PBS and refrigerated for histochemical analysis. For the above study, two independent experiments were carried out with at least two repeats in each experiment (n=4 for histological analysis and n=6 for biochemical analysis).
Quantitative biochemical analysis
DNA content
To assess DNA content, 500 μL of papain digest (100 mM sodium phosphate buffer containing 10 mM L-cysteine (Sigma Aldrich), 125 μg/mL papain (Sigma Aldrich), and 5 mM Na2EDTA (Sigma Aldrich) in ddH2O at pH 6.5) was added to the pellets and pellets were placed in an oven at 60°C overnight, as previously described. 41 Once the pellets were digested the biochemical assays were performed straight away, or stored at −80°C until the assays could be performed. DNA content was performed using Hoechst 33258 DNA assay with calf thymus DNA (Sigma Aldrich) as a standard, following a previously published protocol.17,41–43 Briefly, in minimal light 40 μL of papain digest of the sample/standard was added to a 96-well plate in triplicate. To this 200 μL of working solution (assay buffer and 1 mg/mL Hoechst Dye solution (Sigma Aldrich)) was added. The plate was incubated away from light for 10 min and then read on a microplate reader (Synergy HT BioTek Multi-Mode Microplate Reader) at an excitation of 350 nm and emission of 450 nm as previously described.17,41,43,44
ALP production
Extracellular ALP production was determined using a colorimetric assay of enzyme activity (SIGMAFAST p-NPP Kit; Sigma Aldrich), which uses p-nitrophenyl phosphate (pNPP) as a phosphatase substrate with ALP enzyme (Sigma Aldrich) as a standard. Next, 40 μL of the medium was added to a 96-well plate in triplicate with a 50 μL of pNPP solution, which contains both pNPP and assay buffer. The samples were shielded from direct light at room temperature for 1 h. After this, 20 μL of Stop Solution (3N NaOH) was added to the wells and the plate was read at 405 nm in a microplate reader as previously described.17,43,44
Calcium content
Calcium deposition within the pellets was measured using the Calcium Liquicolor kit (Stanbio Laboratories) according to the manufacturer's protocol. Briefly, pellets were digested by adding 1 mL of 0.5 M HCL and rotating the solution overnight in a cold room. Next, 10 μL of each of the digested samples and assay standard was added to a 96-well plate and 200 μL of the working solution was added. The plate was analyzed on a microplate reader at an absorbance of 550 nm as previously described.17,45
Enzyme-linked immunosorbent assay for vascular growth factor
An enzyme-linked immunosorbent assay (ELISA; R&D Systems) was used to quantify the levels of vascular endothelial growth factor (VEGF) expressed by the pellets. The cell culture media was collected and analyzed at the specific time points. Assays were carried out according to the manufacturer's protocol (R&D Systems) and analyzed on a microplate reader at a wavelength of 450 nm.
Histology
Pellets were obtained at day 0, 1 week, 2 weeks, and 3 weeks post media switch for histological analyses. After each sample had been fixed overnight in 4% paraformaldehyde, samples were dehydrated and embedded in paraffin using an automatic tissue processor (Leica ASP300; Leica). All samples were sectioned with a thickness of 8 μm using a rotary microtome (Leica microtome, Leica). Sections were stained with 1% Alcian Blue 8GX solution for sGAG, as previously described, 41 and finally 2% alizarin red solution for mineralization (all Sigma Aldrich) as previously described. 17
CD31 and collagen type X immunohistochemical analysis
Immunohistochemical analysis was used to detect CD31 and collagen type X. Sections were deparaffinized overnight before a series of rehydration steps. The samples were then treated with 40 μg/mL of proteinase K for 20 min at 37°C (Sigma Aldrich), rinsed with PBS-Tween, and blocked with PBS with 1% w/v bovine serum albumine (BSA) and 3% w/v normal goat serum (NGS) (Sigma Aldrich) for 60 min. Sections were then incubated overnight at 4°C with either rabbit polyclonal anti-CD31 (ab28364 Abcam, 1:50) or rabbit polyclonal anti-collagen type X (ab58632 Abcam, 1:200). After three washing steps with PBS with 1% w/v BSA the sections were incubated with Dylight488 goat anti-rabbit secondary antibody (Jackson Immunoresearch, 115-485-209, 1/200), for 1 h at room temperature in the dark. The samples were washed thrice in PBS with 1% w/v BSA, and the slides were mounted using propidium iodide (PI) mounting media (Sigma Aldrich). Serum containing blocking solution (PBS with 1% w/v BSA and 3% w/v NGS) was used to replace primary antibodies as a negative control for both CD31 and collagen type X staining. HUVECs provided a positive control for CD31 staining. Chondrocytes within the hypertrophic zone of neonatal mice provided a positive control for collagen type X staining.
Statistical analysis
Results are expressed as mean±standard deviation. For all the biochemical analysis two-way analysis of variance with time and media type as the independent factors followed by a pairwise multiple comparison procedure (Tukey's HSD test) was used to test for significance. All analyses were performed with Minitab. For all comparisons, the level of significance was p≤0.05.
Results
Characterization of human donor MSCs
The differentiation assays confirmed the multipotency of the isolated MSCs that displayed positive markers of (1) osteogenic differentiation; as seen by the positive alizarin red staining by MSCs cultured in osteogenic differentiation media (Fig. 1A) and lack of staining in the negative control group (expansion media) (Fig. 1B), (2) adipogenic differentiation; as seen by the positive Oil Red O stained lipid nodules with MSCs cultured in adipogenic induction medium (Fig. 1C) and lack of lipid staining in the negative control group (expansion media) (Fig. 1D), and (3) chondrogenic differentiation, as seen by the positive sGAG staining (Fig. 1E) compared to a lack of sGAG staining in the negative control group (expansion media) (Fig. 1F).
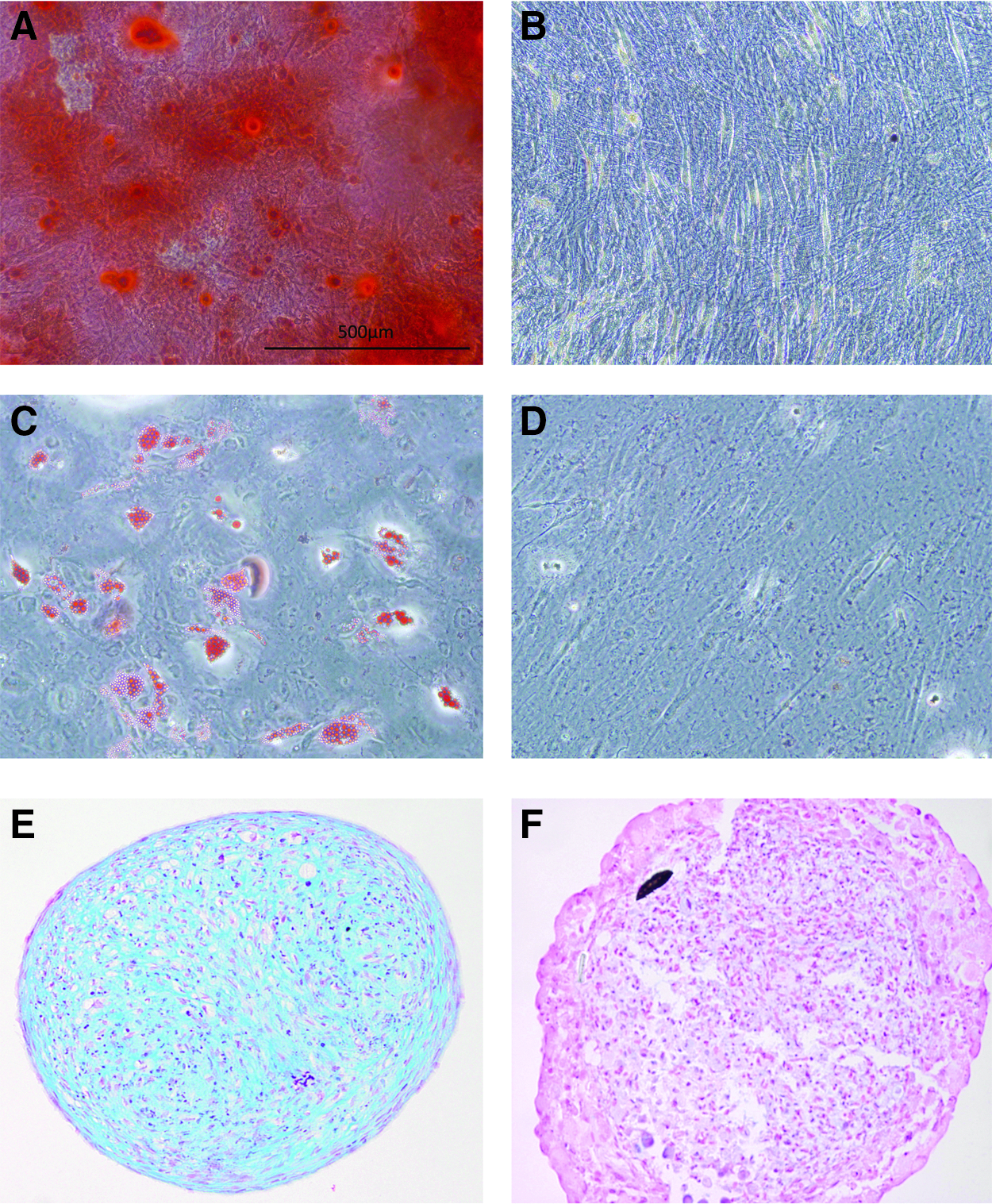
Characterization of the multipotency of isolated human mesenchymal stem cells (MSCs). Calcium deposition is confirmed by positive alizarin red staining by MSCs cultured in osteogenic differentiation media
Cell number
There was no statistical difference in cell number for the CP21-HUVECs group over the course of the experiment. However, there was a significant increase (p<0.05) in DNA content in both the CP21+HUVECs and CP21+HUVECs:MSCs groups from 1 to 3 weeks into culture. The CP21+HUVECs:MSCs group had significantly higher DNA content than both the CP21–HUVECs (p<0.001) and the CP21+HUVECs group (p<0.05) at both 1 and 2 weeks into culture. Both the CP21+HUVECs and CP21+HUVECs:MSCs groups had significantly higher DNA content than the CP21-HUVECs (p<0.001) after 3 weeks (Fig. 2).

DNA content of the coculture groups (CP21+HUVECs, CP21+HUVECs/MSCs) compared with 21 days priming group alone (CP21-HUVECs) pre and 1, 2, and 3 weeks post the addition of HUVECs/MSCs (n=6 samples per group per time point). ^p<0.05 versus CP21+HUVECs group and *p<0.05 versus CP21-HUVECs. Error bars denote standard deviation. HUVECs, human umbilical vein endothelial stem cells.
Production of cartilage template
Alcian blue staining
All three culture groups stained positive blue for sGAG 1 week into culture. HUVECs were still present around the periphery of the pellet in the CP21+HUVECs group after 3 weeks in culture. However, an increase in size of the pellet from 2 (105,914.426 μm2) to 3 weeks (218,870.303 μm2) suggests that the HUVECs/MSCs were incorporated into the pellet after 3 weeks in culture in the CP21+HUVECs:MSCs group (Fig. 3F, I).
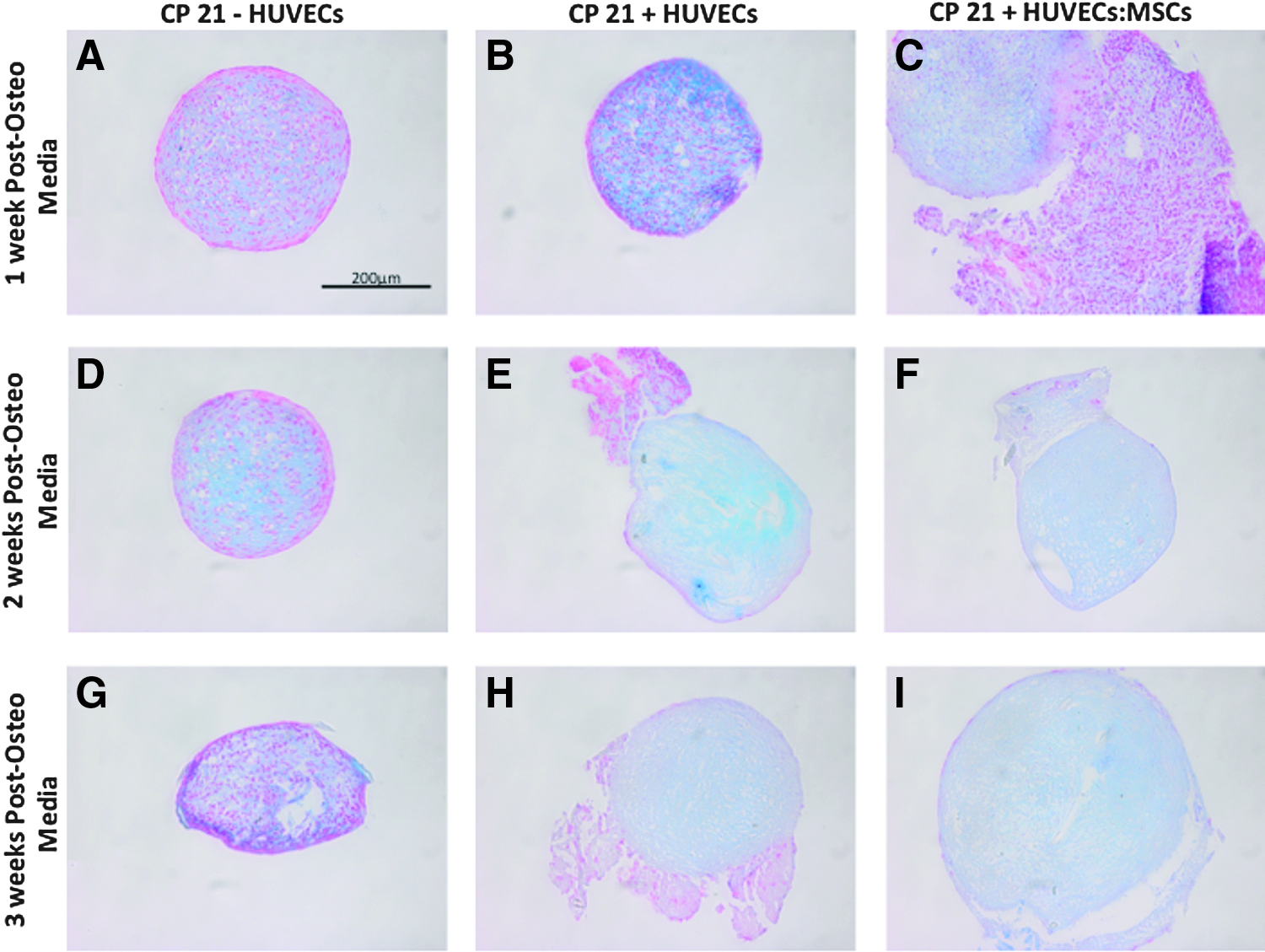
Alcian Blue staining of all coculture groups (CP21+HUVECs
Mineralization of the cartilage template
ALP production
There was no statistical difference between the groups at any time point (Fig. 4).
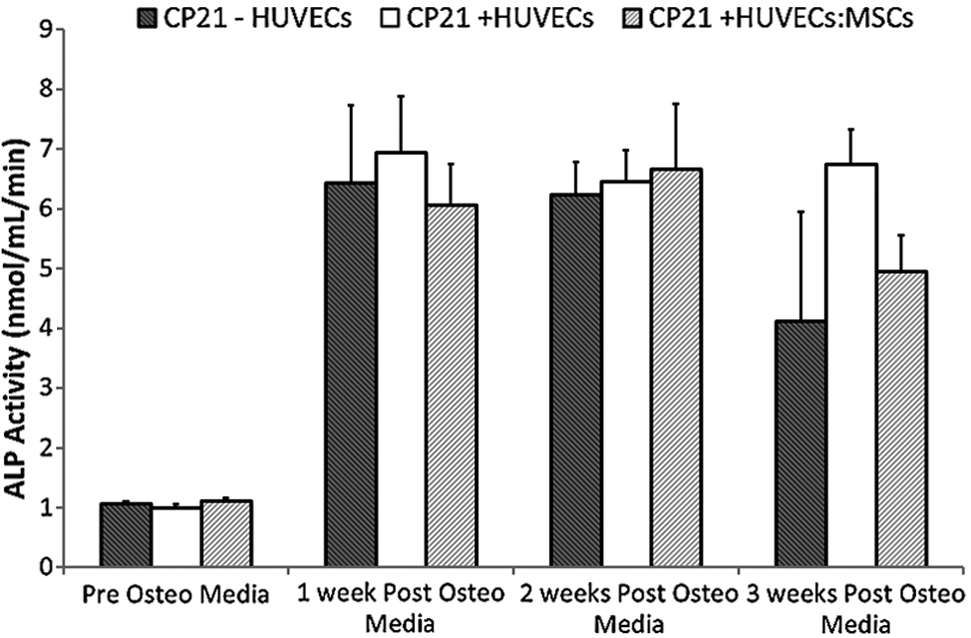
ALP activity of the coculture groups (CP21+HUVECs, CP21+HUVECs/MSCs) compared with 21 days priming group alone (CP21-HUVECs) pre and 1, 2, and 3 weeks post the addition of HUVECs/MSCs (n=6 samples per group per time point). Error bars denote standard deviation. ALP, alkaline phosphatase.
Calcium content
The CP21+HUVECs:MSCS group increased calcium production up to 2 weeks into culture, after which the calcium content began to plateau (Fig. 5). The CP21+HUVECs:MSCS group had significantly higher calcium content (p<0.05) than both the CP21–HUVECs and CP21+HUVECs groups at both 1 and 2 weeks into culture. By 3 weeks this significance was lost, however, there was still a trend toward higher calcium content in both the CP21+HUVECs and CP21+HUVECs:MSCs groups compared to the control, although not statistically different (p-value=0.2) (Fig. 5).
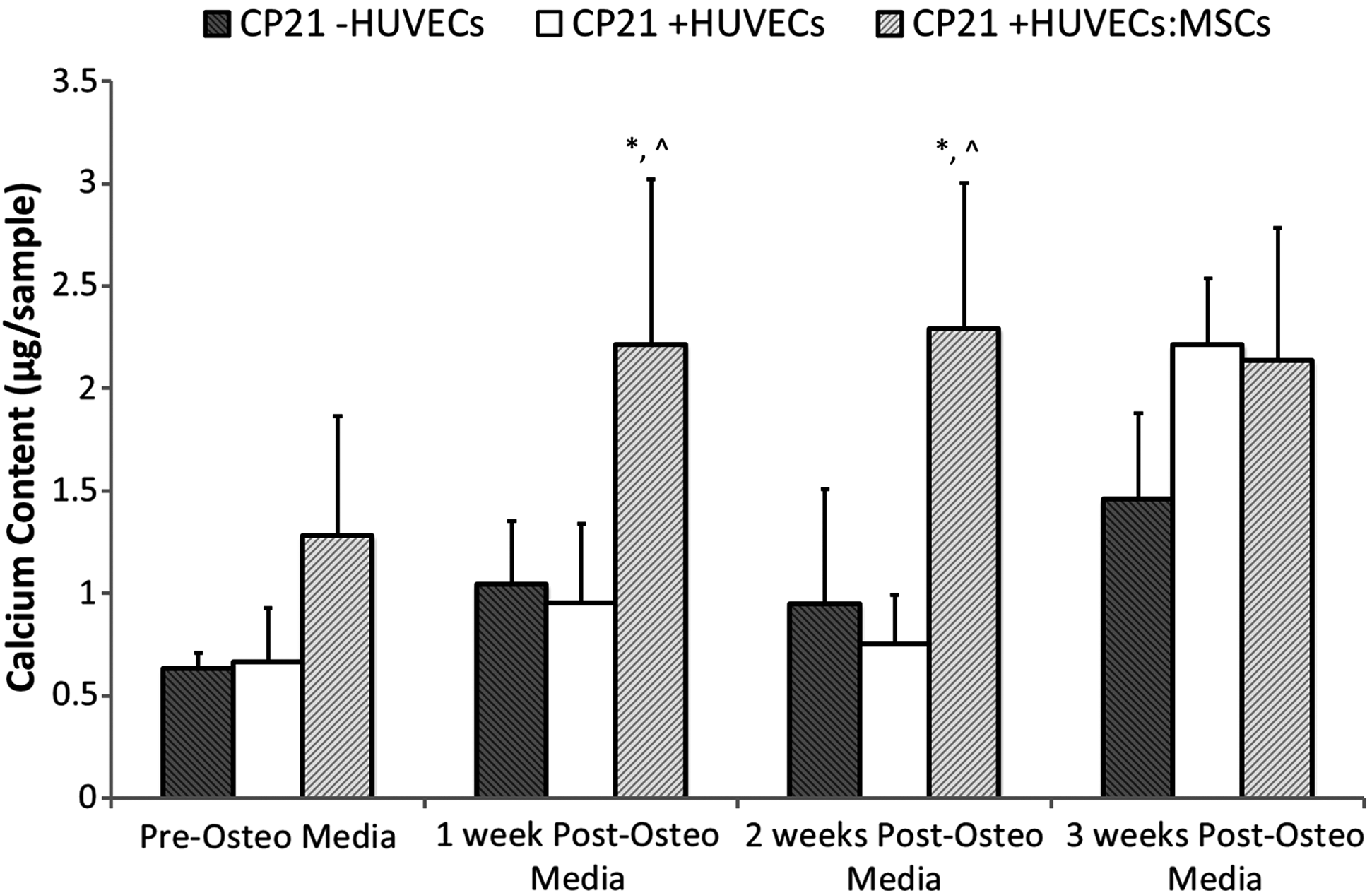
Calcium content for the coculture groups (CP21+HUVECs, CP21+HUVECs/MSCs) compared with 21 days priming group alone (CP21-HUVECs) pre and 1, 2, and 3 weeks post the addition of HUVECs/MSCs (n=6 samples per group per time point). ^p<0.05 versus CP21+HUVECs group; *p<0.05 versus CP21-HUVECs. Error bars denote standard deviation.
Alizarin red staining
The highest amount of positive staining was seen in the CP21+HUVECs:MSCs group after 1 week into culture (Fig. 6). By 3 weeks the highest amount of alizarin red staining was present in the CP21+HUVECs group. This positive staining was seen in the CP21+HUVECs group throughout the pellet, whereas in the CP21+HUVECs:MSCs group the mineralization was present in small discrete nodules throughout the construct. The least amount of alizarin red mineralization at each time point was in the CP21–HUVECs group.
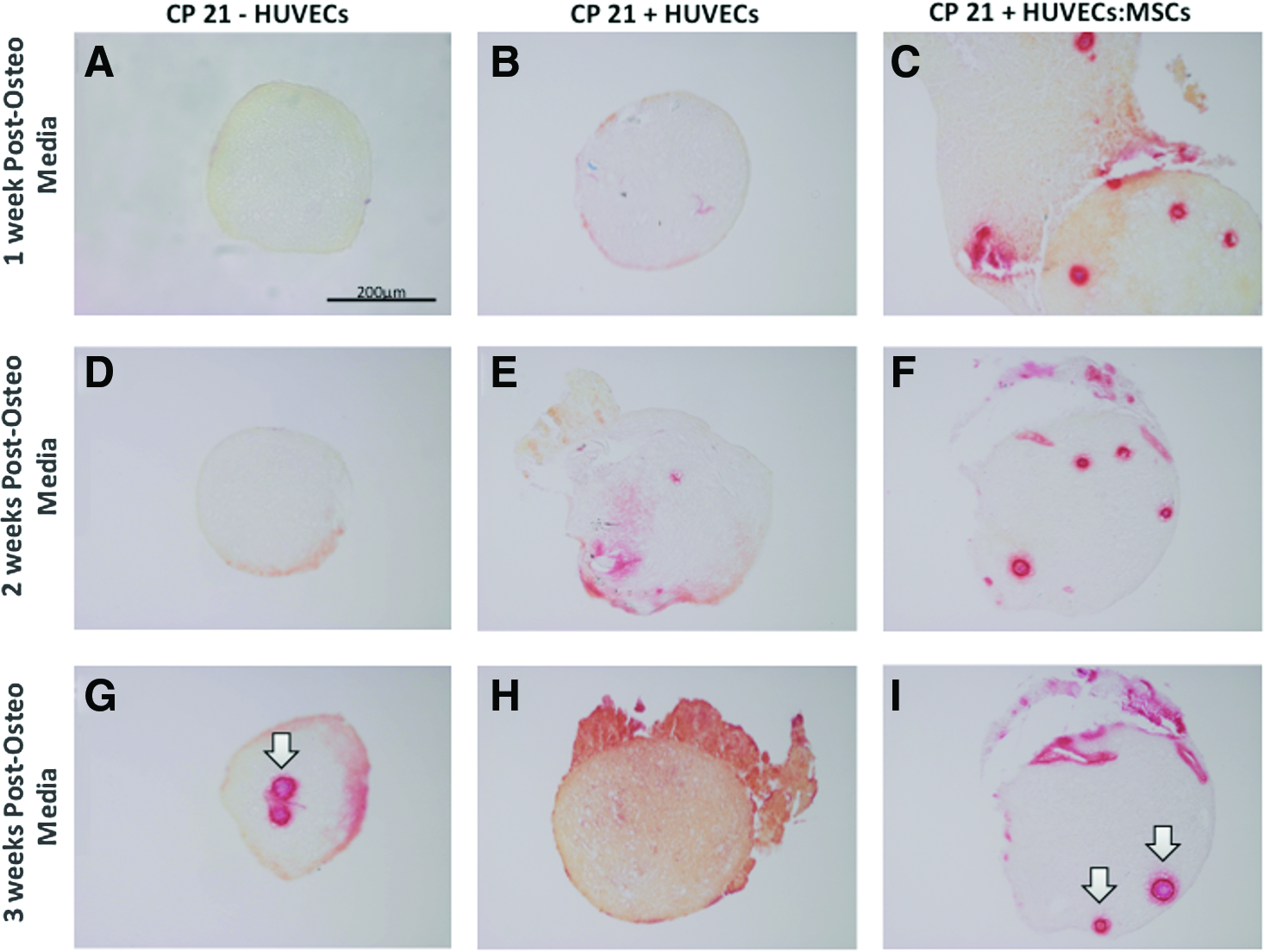
Alizarin red staining of coculture groups (CP21+HUVECs
Vascularization of the cartilage template
VEGF production
The CP21+HUVECs:MSCs group had the highest amount of VEGF production at each time point. At both 2 and 3 weeks post addition of cells there was significantly (p<0.05) higher VEGF content in the CP21+HUVECs:MSCs group compared to the CP21+HUVECs group. By 3 weeks the CP21+HUVECs:MSCs group had significantly higher (p<0.01) VEGF content compared to both the other groups (Fig. 7).
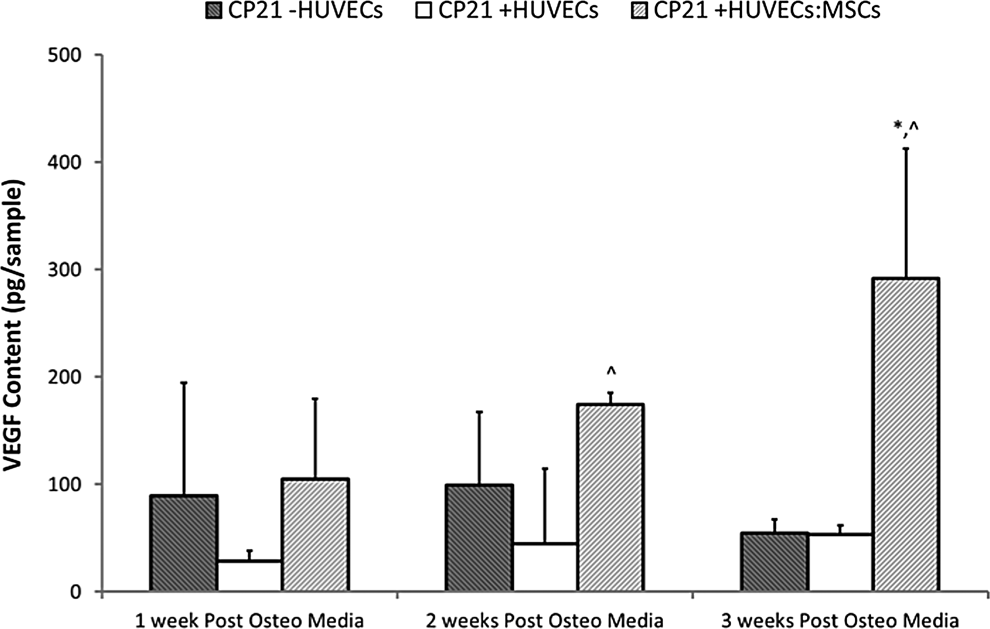
VEGF content the coculture groups (CP21+HUVECs, CP21+HUVECs/MSCs) compared with 21 days priming group alone (CP21-HUVECs) pre and 1, 2, and 3 weeks post the addition of HUVECs/MSCs (n=6 samples per group per time point). ^p<0.05 versus CP21+HUVECs group; *p<0.05 versus CP21+HUVECs: MSCs. Error bars denote standard deviation. VEGF, vascular endothelial growth factor.
CD31+ staining
At all time points there was a distinct lack of positive staining for CD31+ in the CP21-HUVECs group, as expected there were no HUVECs present. Positive staining for CD31+ was present around the periphery of the cartilage template in the CP21+HUVECs group at all time points (see arrows), but there was little infiltration into the cartilage template. However, with the CP21+HUVECs: MSCs group there was positive CD31+ staining on the periphery of the cartilage template 1 week into coculture. By 2 weeks there was positive CD31+ staining within the template, and by 3 weeks into coculture rudimentary vessels had formed within the template (Fig. 8). These rudimentary vessels were present in a series of sections throughout the pellets (Fig. 9A–D) with an average cross-sectional area of 220.89 μm2 and average diameter of 3.79 μm (Fig. 9E, F).
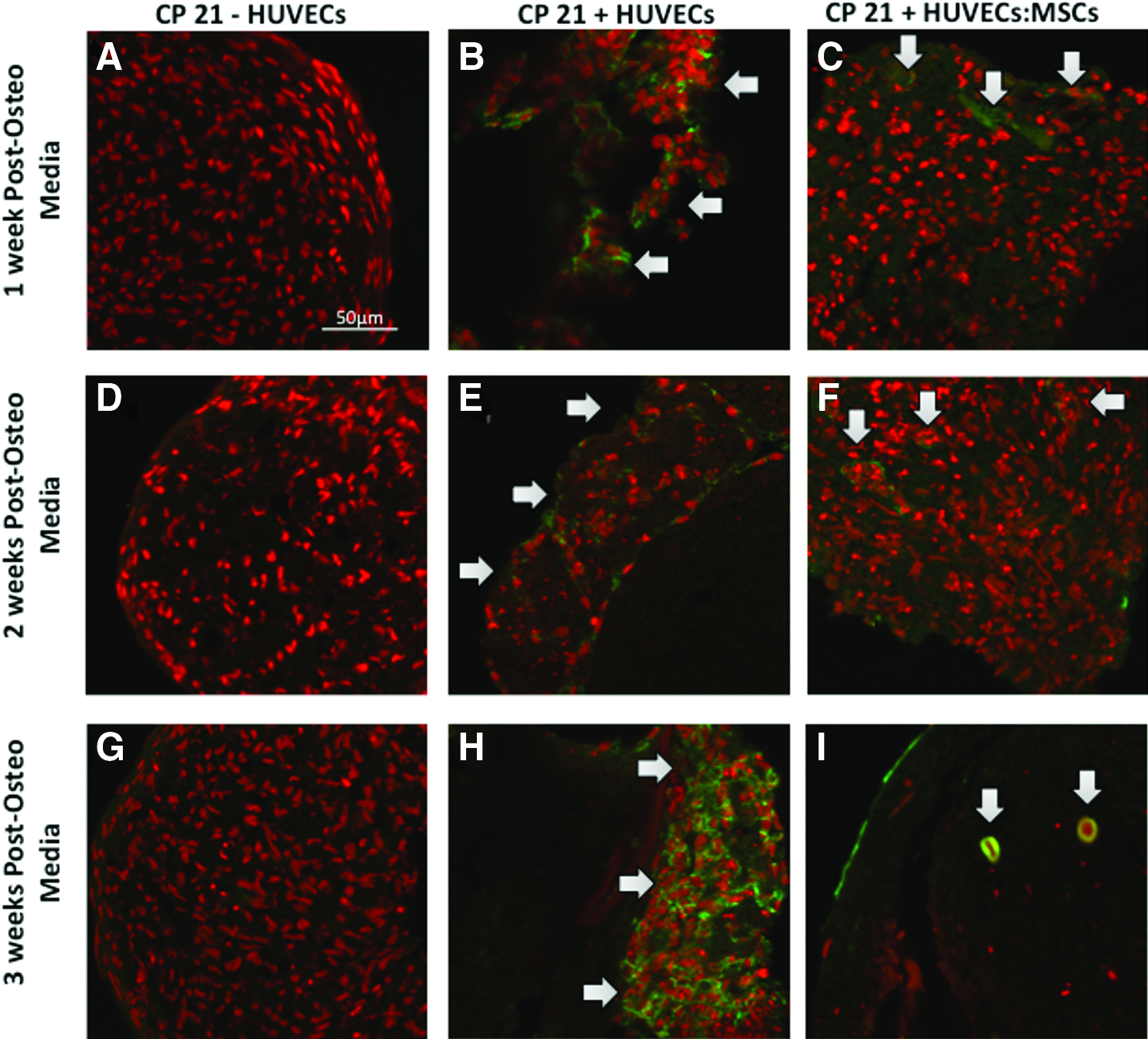
CD31 staining (green) of coculture groups (CP21+HUVECs
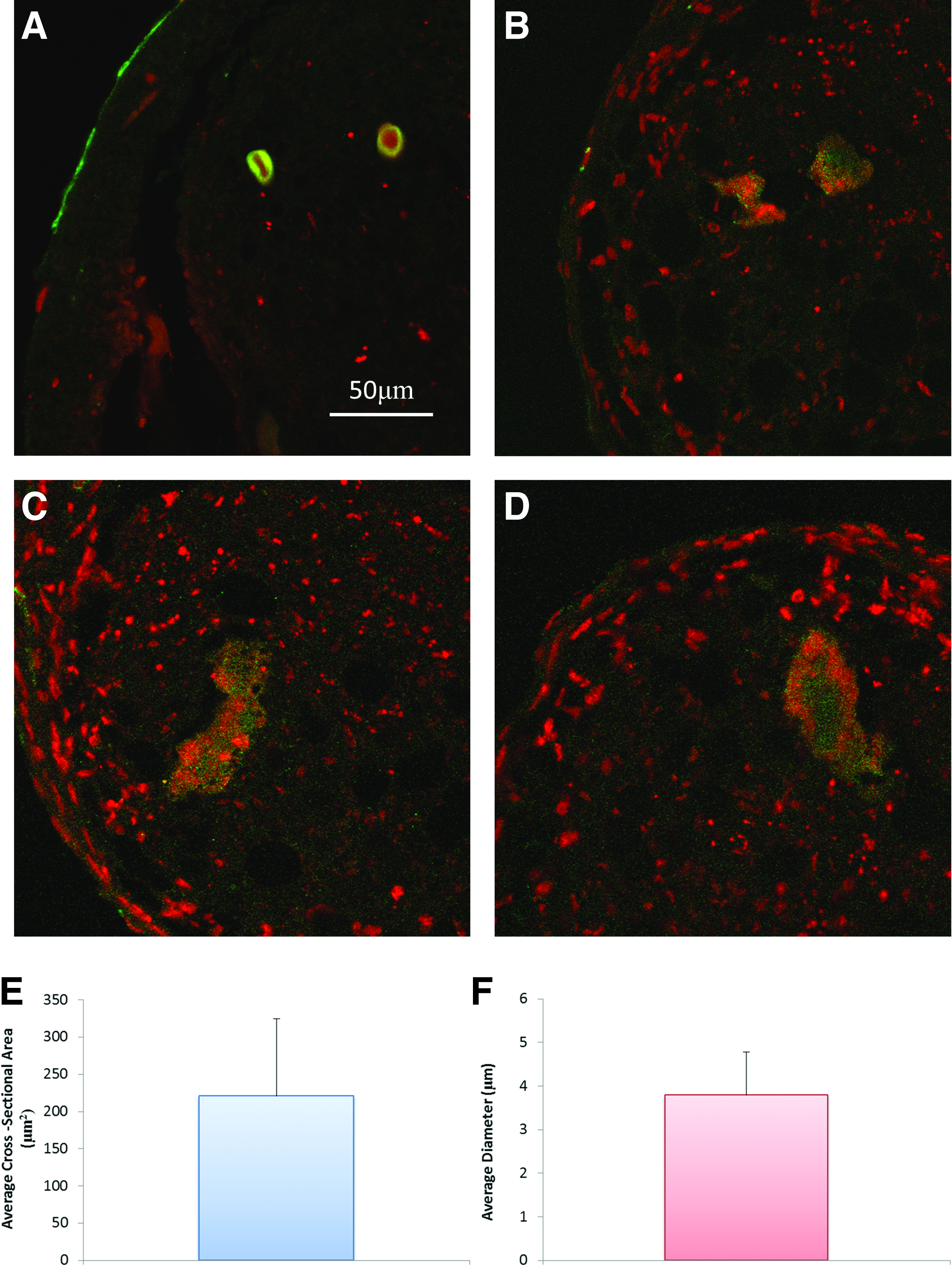
Representative images of CD31+ (green) stained sections showing the presence of rudimentary vessels
Production of hypertrophic cartilage template
Collagen type X staining
There was a higher amount of positive staining for collagen type X in both CP21+HUVECs and CP21+HUVECs:MSCs groups compared to the CP21-HUVECs group after 3 weeks into culture (Fig. 10G–I, arrows). This collagen type X was predominately seen around the periphery of the constructs. The positive control stained chondrocytes were identified in the growth plate of long bones from a neonatal mouse model (Fig. 10K), and negative controls replacing primary antibody by normal serum were used (Fig. 10J) and did not show any specific staining.
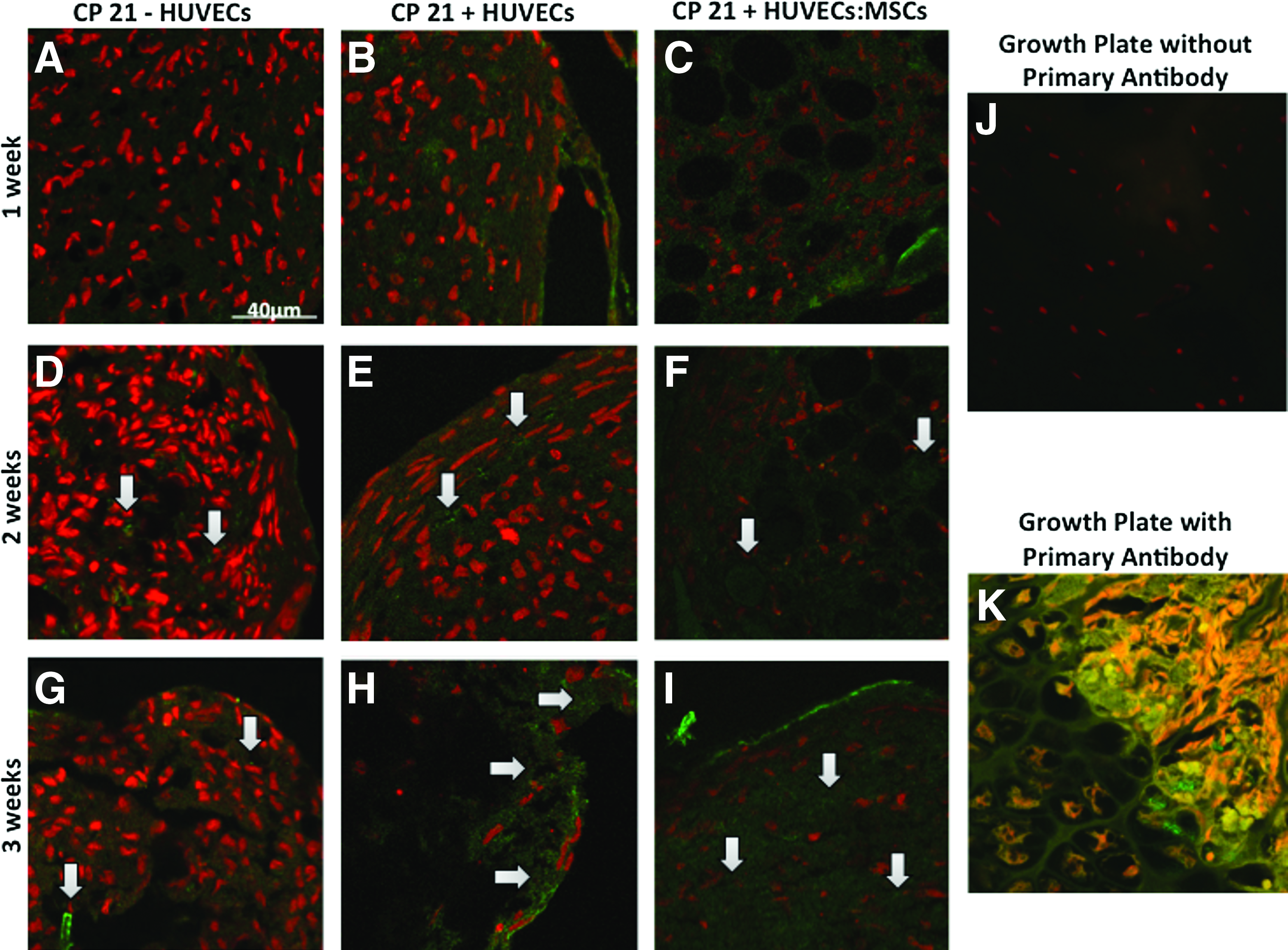
Collagen type X staining (green) of coculture groups (CP21+HUVECs
Discussion
In this study we show that both CP (for a period of 21 days) and subsequent vascular priming can enhance the mineralization potential of MSCs produced through CP alone. In particular we show that a cartilage template provides a suitable platform for HUVECs/MSCs to attach, proliferate, and infiltrate for up to 3 weeks in culture (as indicated by CD31+ staining). The results also show that an upregulation of VEGF expression occurs in the coculture group when both MSCs/HUVECs added compared to just HUVECs added and the non coculture group. This enhanced VEGF expression, seen with the MSCs/HUVECs added coculture group, was further supported by the fact that it was the only group to have the formation of immature vessels. These rudimentary vessels were present in a series of sections throughout the pellets (as seen through CD31+ staining). Interestingly, this study also shows that the highest amount of mineralization was seen when HUVECs alone were added to the cartilage template, whereas when both MSCs and HUVECs were added to the culture the mineralization was reduced, although vascularization and formation of rudimentary vessels were observed. Together these results suggest that there may be a trade-off between mineralization and vascularization for in vitro bone regeneration strategies for the durations investigated in this study.
One limitation of this study was that osteogenic and chondrogenic factors (ascorbic acid, dexamethasone, β-glycerol, and TGF-β3) were introduced into the culture media of MSCs to encourage MSC differentiation down specific pathways. It was unlikely that these factors were present in the combinations used here during endochondral ossification in vivo, and some concern may exist regarding cell viability in long-term in vitro studies. However, previous research studies have exposed MSCs to both chondrogenic and osteogenic differentiating agents for long durations (14 days to 5 weeks) and have demonstrated long-term viability and matrix production by MSC's.10,16,41,46–53 Furthermore, the control groups were exposed to the same osteogenic or chondrogenic factors; so the differences observed in osteogenesis, due to the combined strategy of CP and coculture with HUVECs/MSCs, cannot be explained by differences in cell viability arising from growth factors. Another potential limitation is donor variability, specifically the age differences between the donors. While the age differences between the middle-aged donors and young donor may confound these results, it was not feasible to obtain a large enough stock of human bone marrow MSCs from bone marrow aspirates alone to conduct all of the necessary experiments, involving numerous treatment groups and time points. However, it should be noted that upon analysis of the data we found little variation between the pellets formed from MSCs of middle-aged donors and the young donor for any of the results (DNA content, calcium content, etc.), and as such our findings of the importance of a combination of CP and vascular priming are deemed to be applicable to both young and older donors. A limitation of this study is that the mineralization potential has been evaluated from the in vitro production of mineral through alizarin red staining, which gives a measure of mineral quantity and distribution rather than the quality and mechanical integrity of that mineralized matrix. Future studies are required to evaluate this platform further both in vitro and in vivo to investigate both mineral quantity and quality, ultimately to understand whether this matrix has the potential to fulfill the load-bearing functions of bone tissue in the body.
The first stage of endochondral ossification involves MSCs condensing and aligning to become clusters of cells, and subsequently these MSCs differentiate down the chondrogenic pathway.18–20 Our previous studies have shown that CP for 21 days lead to the formation of a cartilage-like tissue, 17 and this is also further confirmed through the histological staining of this study. However, in this study we also see that this cartilage template forms a suitable platform for HUVECs and MSCs to attach, proliferate, and invade. Previous studies have reported that CP of human MSCs can enhance mineralization both in vitro16,17 and once implanted subcutaneously can enhance ectopic bone formation,10,12,15–17 however, these studies did not provide tissue appropriate for clinical use, due to issues with core degradation and an uneven distribution of bone mineral throughout the construct.10,12,16 Other studies have shown the clinical potential of chondrogenically primed rat MSCs for the repair of large bone defects, by implanting chondrogenically primed MSCs in either cellular aggregate form or seeded upon biomaterial scaffolds, and comparing the amount of bone formation, to that of constructs seeded with undifferentiated rat MSCs alone.13,14 However, as these studies used animal stem cells, the results cannot be completely transferred to human cells and before this the response of human MSCs to chondrogenic and vascular priming was unknown. This study shows for the first time that CP can positively affect the mineralization potential of human MSCs. Moreover, this study demonstrates that vascular priming also plays an important role in the mineralization capacity of human MSCs. As such, the results of this study suggest that a combination of chondrogenic and vascular priming is an effective strategy for osteogenic differentiation of human MSC aggregates in vitro. Future in vivo studies are required to investigate the effect of CP and vascular priming in vivo. In particular it is necessary to establish whether such constructs will survive once implanted in vivo and whether they can be scaled to aggregates of sufficient size for clinical treatment of large bone defects.
Previous studies have shown that the coculture of MSCs and vascularization cells (HUVECs) alone can enhance the osteogenic potential of MSCs when immediately put into direct coculture.25,27, 38 However, these studies only looked at ALP activity, which is an early maker for bone formation and cannot be directly used to infer the likelihood of bone mineralization, which is necessary to provide a construct with sufficient mechanical integrity. Our study shows for the first time the beneficial effect of vascular priming for encouraging the production of calcium, a crucial indicator of the formation of bone mineral in the construct. Interestingly, our results also show that the addition of both HUVECs and MSCs, compared to HUVECs alone, has a significant effect on mineralization location within the constructs. The highest amount of mineralization is seen when HUVECs are added to the cartilage template alone. Additionally, it is the only construct to have mineralization throughout the construct. Previous CP studies have been limited by mineralization only occurring around the periphery or in the core alone, but not throughout the pellet.12,16 Interestingly, when HUVECs were added to the culture alone there was indeed mineralization throughout the construct rather than just around the periphery, as was shown in Figure 6. However, when both MSCs and HUVECs were added to the culture the mineralization was reduced, compared to the addition of HUVECs alone, and was characterized by the formation of discrete mineralized nodules rather than homogenous mineralization throughout the construct. It is important to note that even though the coculture group with MSCs and HUVECs had less mineralization than the group with HUVECs added alone, it still had higher mineralization and more nodules than the control. We propose that, during the process of vessel formation mineralization did not proceed and as a result mineralization did not initiate until later for this group. Therefore, it is possible that culturing this group in vivo after both the chondrogenic and vascular priming period in vitro will ultimately allow enhanced mineralization, but this cannot be verified from the results of this study and future in vivo investigations are required.
Hypertrophy and the formation of vasculature networks, by a process known as quiescent angiogenesis, is the next stage of endochondral ossification and both events proceed to bone formation by means of this process in vivo.18–21 In this study, we see that both coculture groups had begun to undergo hypertrophy as the highest amount of collagen type X is seen in these groups after 3 weeks in culture. Interestingly, the highest amount of collagen type X staining was present around the periphery of the construct and not in the center. In vivo, 21 the onset of angiogenesis leads to hypertrophy of the cartilage template, so the addition of HUVECs to the periphery of the cartilage template might attribute to the cells around the periphery to become hypertrophic. Previous studies 54 have also found that the coculture of human MSCs and articular chondrocytes reduces the potential of the chondrocytes to become hypertrophic. Therefore, the reduction of collagen type X in the coculture group with HUVECs and MSCs seen in this study may be due to the addition of undifferentiated MSCs. However, further studies are needed to verify this.
The addition of the HUVECs or HUVECs and MSCs had a significant effect not only on the mineralization potential of the MSCs but also on the VEGF expression. VEGF has been shown to be the marker to stimulate vascular cells to undergo the formation of early vessels16,21,55–58 and previous studies have postulated that vasculogenesis should be induced before osteogenesis in vitro to obtain functional bone tissue in vivo. 31 Therefore, even though the coculture group with MSCs/HUVECs had less mineralization than that with HUVECS alone, it was the only group to form immature vasculature structures with an average diameter 59 and structure60,61 similar to those seen in vivo of early arterioles. It also had the highest expression of VEGF, and as such might have the best potential for host integration and mineralization once implanted in vivo.
Conclusions
The results of this study show that both CP (for 21 days) and coculture of MSCs and HUVECs can significantly increase the osteogenic potential produced through CP alone. Moreover, these results show that the formation of a cartilage-like template provides a suitable platform for HUVECs/MSCs to attach, proliferate, and infiltrate for up to 3 weeks. More importantly, we show that both MSCs and HUVECs must be added to the formed cartilage template for the formation of immature vessels. Taken together, these results indicate for the first time that the application of both chondrogenic and vascular priming of MSCs enhances the mineralization potential of MSCs, compared to CP alone in vitro, while also allowing for vessel formation. This study provides a valid model to study the endochondral ossification process in vitro and will inform novel tissue regeneration strategies for large bone defects.
Footnotes
Acknowledgments
This project was supported by the European Research Council Grant 258992 (BONEMECHBIO).
Disclosure Statement
No competing financial interests exist.