
Abstract
Dedifferentiated fat cells (DFAT cells) isolated from adipose tissue have been demonstrated to differentiate into chondrogenic cells in vitro. Nevertheless, an efficient method to facilitate its chondrogenic differentiation is still unexplored, hampering the extensive application of these cells in cartilage regeneration therapies. Here we provide the evidence that supplementation of strontium ions (Sr) in a chondrogenic medium (CM) significantly promotes early chondrogenic differentiation of DFAT cells. Human DFAT cells and the mesenchymal stem cell line (RCB2153) were subjected to the CM supplemented with/without Sr. After 14 days, alcian blue staining intensity significantly increased in DFAT cells, but not in RCB2153, subjected to CM with Sr. mRNA expression analysis revealed that the CM with 1.5 mM Sr increased the expression of chondrogenic marker, collagen type 2 alpha 1, whereas there was no significant change in osteogenic markers, collagen type 1 alpha 1, runt-related transcription factor 2, and osteocalcin, and hypertrophic chondrogenic marker, collagen type 10 alpha 1. Inhibitors for extracellular signal-regulated kinase 1/2 (ERK1/2), Akt, and calcium-sensing receptor (CaSR) pathways significantly diminished the alcian blue staining intensity, providing the first evidence that these signal pathways are associated with chondrogenic differentiation of DFAT cells. CaSR and ERK1/2 pathways independently induced Sr-mediated early chondrogenic differentiation. These results suggest that Sr supplementation into the CM may provide a powerful platform for preparing chondrogenically differentiated DFAT cells for cartilage regeneration.
Introduction
A
Adipocyte-derived dedifferentiated fat cells (DFAT cells) were first established using a ceiling culture technique by Sugihara et al.6,7 The DFAT cells have recently been shown to differentiate into multiple mesodermal lineages, including osteoblastic,8–17 chondrogenic,9,17 adipogenic,9,18 and myogenic cells.17,19,20 Bone formation has been enhanced by the use of DFAT cells in rabbit 21 and rat 15 models. DFAT cells are similar to MSCs and ASCs in terms of cell markers and proliferative capacity, but are more homogenous7,9 and can differentiate into osteoblasts much earlier.11,22 Despite numerous reports of the utility of DFAT cells for bone tissue engineering,8–17,21,22 there remains no effective method to induce chondrogenic differentiation. Given the abundance and accessibility of adipose tissue compared with bone marrow, chondrogenically differentiated DFAT cells are likely to be an ideal alternative cell source for broadening cartilage regeneration therapies.
Strontium ion (Sr), an essential element for the human body, has been widely applied as a pharmacological drug strontium ranelate in osteoporotic structural damage. 23 Numerous studies have demonstrated the safety and efficacy of orally administered strontium ranelate. Sr exhibits a dual effect on bone metabolism 24 hindering the osteoclastic differentiation 24 and promoting the osteogenic differentiation 25 and the osteoblastic cell replication. 26 Although Sr is known to enhance the extracellular matrix formation of isolated rat 27 and human chondrocytes 28 in vitro, there has been no study that tried to use this element to chondrogenic differentiation on DFAT cell.
This study was designed to investigate whether a chondrogenic medium supplemented with Sr [CM+Sr(N), N=concentration of Sr] facilitates chondrogenic differentiation of primary human DFAT cells isolated from subcutaneous adipose tissue and the buccal fat pad. Using inhibitors for four pathways (extracellular signal-regulated kinase 1/2 [ERK1/2], Akt, p38-mitogen-activated protein kinase [p38-MAPK], and calcium sensing receptor [CaSR]), we verified how these signal transduction pathways are involved in CM+Sr-induced chondrogenic differentiation of DFAT cells.
Materials and Methods
Isolation and culture of primary human DFAT cells
DFAT cells were isolated from the adipose tissue of a healthy male (donor A) and a healthy female (donor B) patient. The donors had no systemic disease and underwent oral and maxillofacial surgery. The ages of the donors and the origin of the cells were as follows: DFAT-A cells, donor A, age 63 years, subcutaneous adipose tissue; DFAT-B cells, donor B, age 34 years, buccal fat pad. This study conformed to the tenets of the declaration of Helsinki, and the protocol was approved by the Ethics Committee of Osaka Dental University and Amagasaki Chuo Hospital (approval number: 110760). DFAT cells were prepared using the ceiling culture method as described previously (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).11,22 In brief, adipose tissue was filtered through a nylon mesh to remove undesirable cells, including stromal cells, vessel cells, and connective tissues. Mature adipocytes were seeded into the culture flask with the adhesive surface facing upward so that floating adipocytes containing lipid droplets attached themselves to the inner ceiling of the flask. After 7 days, the flask was inverted to remove the mature adipocytes. Although residual mature adipocytes with lipid droplets were still slightly contaminated right after the inversion, these cells gradually disappeared after several passages, and uniformed fibroblastic cells, designated DFAT cells, covered the bottom of the flask (Fig. 1A). DFAT cells at passages 3–5 were used for each assay. DFAT cells were identified by flow cytometric analysis (Supplementary Fig. S2).

Cell culture
The human MSC line (RCB2153) isolated from normal human bone marrow was gifted from Riken Cell bank (Tsukuba, Japan). DFAT cells and RCB2153 were maintained in Dulbecco's modified essential medium supplemented with 10% fetal bovine serum and 1% antibiotics in a 5% CO2 incubator at 37°C.
Chondrogenic differentiation
Both cells were seeded at 25,000 or 30,000/well in a 12-well plate and cultured till day 14 after treatment with each condition medium. Three preincubation conditions after the cell seeding were prepared to evaluate the distinct effect of confluency (Fig. 1B). At prescribed dates, both cells were subjected to (1) control medium (Cont), (2) CM similar to the previous report, 9 (3) CM+Sr containing 0.15–1.5 mM Sr (Table 1) to induce chondrogenic differentiation. In the pilot study, compared with CM+Sr(1.5), the higher dose of Sr attenuated the cell viability and chondrogenic differentiation of DFAT cells (Supplementary Figs. S3 and S4).
CM, chondrogenic medium; DMEM, Dulbecco's modified Eagle's medium; FBS, fetal bovine serum.
Alcian blue staining and its quantification analysis
Chondrogenic differentiation was partially assessed by alcian blue staining. Cells were fixed with 0.1% glutaraldehyde in phosphate-buffered saline (PBS) and then rinsed with PBS. The cell plates were stained with 1% alcian blue 8-GX (Sigma Chemical Co, St. Louis, MO) in 0.1 M HCl for 2 h, staining the highly sulfated glycosaminoglycans (GAGs) representative of cartilaginous matrix. After staining, the cells were washed with 0.1 M HCl and PBS. As for quantification data, 6 M guanidine hydrochloride solution was added to the stained cells. After gently shaking for 5 h, the absorbance of supernatants was measured at 620 nm. The alcian blue staining level was calculated as follows: Relative alcian blue staining level=absorbance of sample/absorbance of benchmark. Cells treated with Cont and CM+Sr(1.5) were used as this benchmark.
Inhibition of cell signaling pathways
Cell singling pathways associated with CM+Sr-induced chondrogenic differentiation of DFAT cells were investigated using various inhibitors, CaSR (NPS 2143), ERK1/2 (PD98059), Akt (API-2), and p38-MAPK (SB203580), obtained from Sigma Chemical Co. These inhibitors were mixed with CM+Sr(1.5) at the following concentrations: 2.5, 25, and 50 μM for PD98059; 1, 10, and 20 μM for API-2; 1, 10, and 20 μM for SB203580; 5, 50, and 500 nM for NPS2143.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Inc., Valencia, CA). Reverse transcription was performed with 1 μg RNA using the Transcriptor Universal cDNA Master (Roche Diagnostics, Mannheim, Germany). mRNA levels were determined for Sox9 (SOX9), collagen type 2 alpha 1 (COL2A1), collagen type 1 alpha 1 (COL1A1), runt-related transcription factor 2 (RUNX2), collagen type X alpha 1 (COL10A1), osteocalcin (OCN), and calcium-sensing receptor (CASR) using Universal Probelibrary set human and a FastStart Universal Probe Master (Roche Diagnostics). Optimal primer and probe sets were designed using the Universal Probe Finder in Roche Applied Science website (Table 2). The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was evaluated using gene expression assays (#4310884E; ThermoFisher Scientific, Inc., Waltham, MA). Quantitative real-time polymerase chain reaction (qPCR) was performed using a Step one plus (ThermoFisher Scientific, Inc.). The cycling conditions were as follows: 10 min at 95°C, 45 cycles of 15 s at 95°C, 60 s at 60°C. All signals were normalized to the GAPDH signal. mRNA expression levels were calculated using the comparative CT method.
Cell morphology and immunofluorescence staining
DFAT cells and RCB2153 were cultured on cell culture plates and chamber strips. As for immunofluorescence staining, cells were fixed in 4% paraformaldehyde in PBS and incubated with the primary antibody (rabbit anti-human CaSR: GTX101348; GeneTex, Inc., Irvine, CA). Following incubation with a secondary antibody (FITC goat anti-rabbit IgG, #111-095-045; Jackson ImmunoResearch Laboratories, Inc., West Grove, PA), cells were mounted in Prolong Gold antifade regents with DAPI (Thermo Fisher Scientific, Inc.). All immunofluorescence images were captured using a confocal laser scanning microscope Zeiss LSM700 (Carl Zeiss Microscopy, Jena, Germany).
Western blotting
After a 30-min stimulation, cell lysates were obtained using a cocktail, including the Bolt LDS sample buffer and Bolt Sample Reducing Agent (B0007 and B0009), following Invitrogen's protocol. The proteins were separated by SDS-PAGE and transferred onto PVDF membranes using the iBlot Dry Blotting system. The iBind western system performed hands-free blocking, antibody binding, and washes. Signals were detected using the SuperSignal West Pico Chemiluminescent Substrate or West Femto Maximum Sensitivity Substrate (Thermo Fisher Scientific, Inc.). The rabbit anti-phospho-ERK1/2 (#9101), rabbit anti-phospho Akt (#4060), and rabbit anti-phopho-p38 MAPK (#4511) antibodies were obtained from Cell Signaling Technology (Beverly, MA). Goat anti-rabbit IgG-HRP (#170-6515) was purchased from Thermo Fisher Scientific, Inc. The mouse anti-GAPDH antibody (H00002597-M01) from Abnova (Taipei City, Taiwan). The Novex® AP Mouse Chemiluminescent Detection Kit from Thermo Fisher Scientific, Inc. was used for the chemiluminescent detection of GAPDH.
In vivo experiment using cell sheet technology
The cell sheet technology 29 was used for obtaining cell sheets from the DFAT cells treated with CM and CM+Sr(1.5) (Supplementary Fig. S5A–D). Cell culture dishes were purchased from CellSeed, Inc. (Tokyo, Japan). The cells were cultured at condition 2 (Fig. 1B); the cell sheets were obtained according to the manufacturer's instruction manual. The sheets were collected, as gel, into a 1.0-mL syringe equipped with an injection needle. An appropriate volume of the gel was subcutaneously injected into male, 8-week-old, C.B-17/IcrHsd-Prkdc scid (SCID) mice for 3 months. SCID mice were purchased from Shimizu Laboratory Supplies Co., Ltd (Kyoto, Japan). All animal experiments strictly followed the guidelines approved by the Local Ethics Committee of Osaka Dental University (approval number: 14-02022).
Statistical analysis
Statistical significance was assessed with one-way analysis of variance with Dunnett's and Tukey–Kramer multiple comparisons tests. Microsoft Excel software statistic package was used for the calculation.
Results
Sr enhance chondrogenic differentiation of DFAT cells
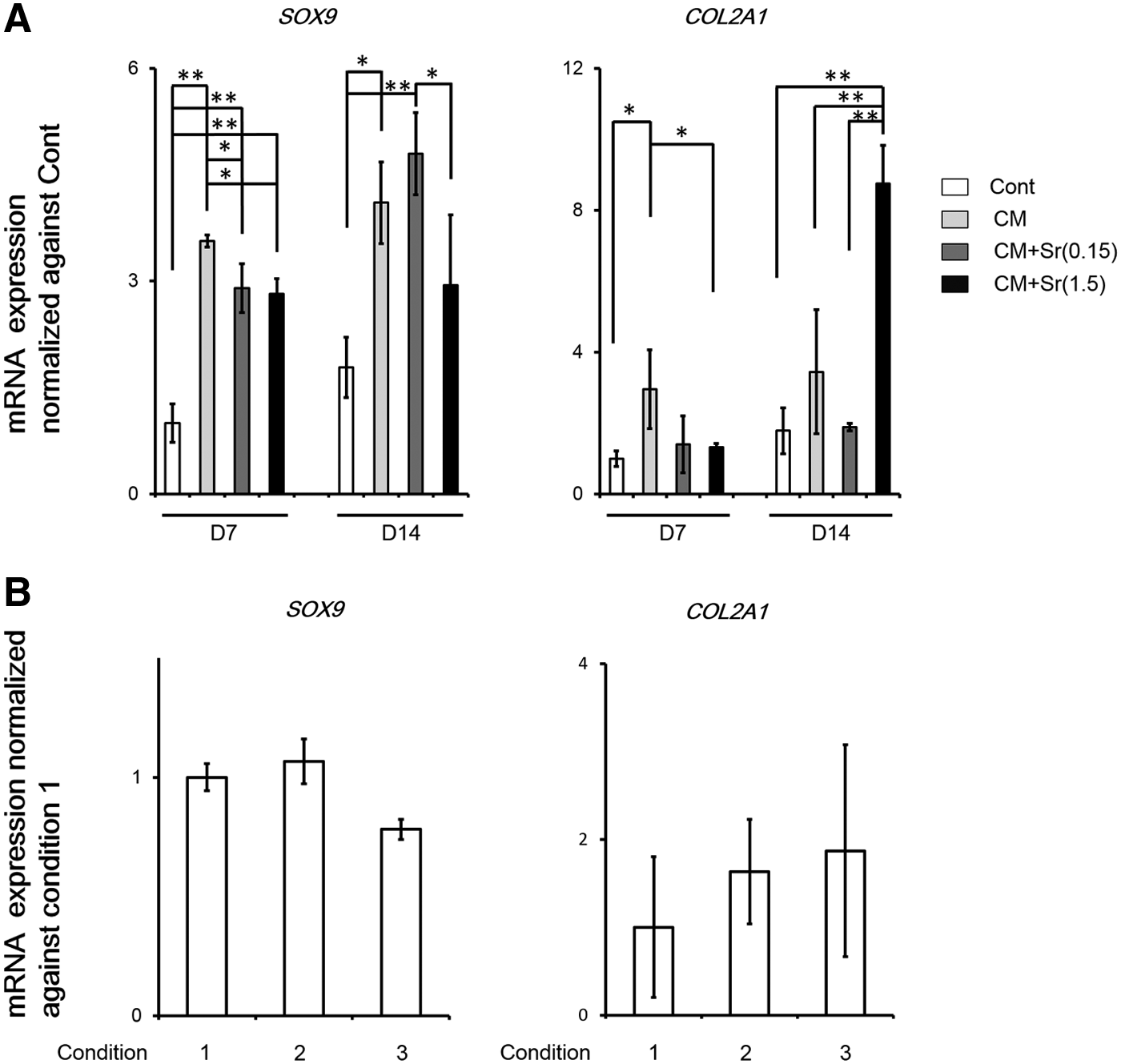
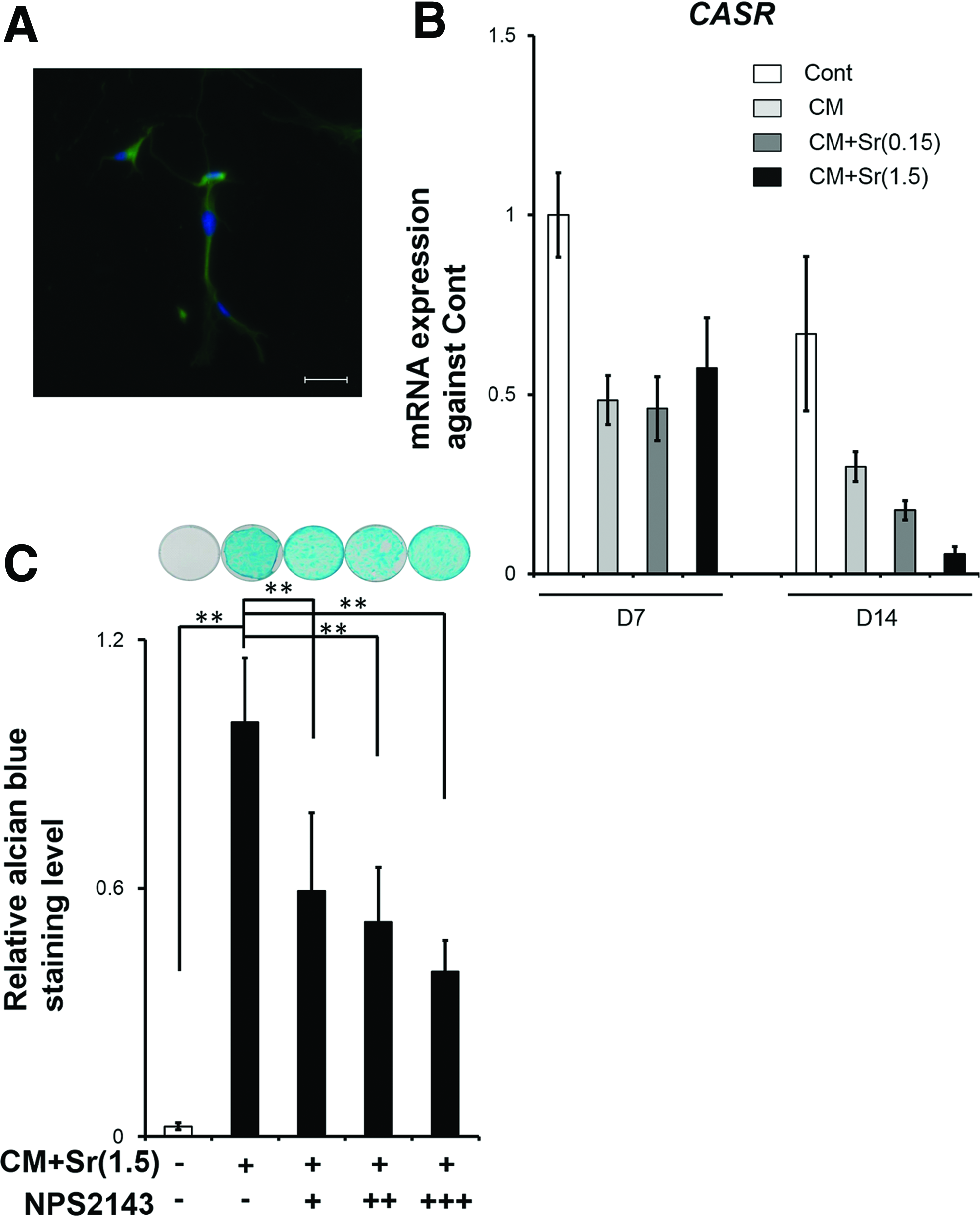
Using alcian blue staining and qPCR, we evaluated whether Sr supplementation of CM facilitated chondrogenic differentiation of DFAT cells and RCB2153. CM+Sr(1.5) significantly and dose dependently enhanced alcian blue staining intensity of stimulated DFAT cells, but not RCB2153. CM+Sr(1.5) resulted in a higher alcian blue staining intensity at day 14 compared to Cont and CM (Fig. 2). Similarly, mRNA expression of COL2A1, a marker of chondrogenic differentiation, increased robustly in DFAT cells treated with CM+Sr(1.5) (Fig. 3A). These results indicate that Sr supplementation to CM upregulates chondrogenic differentiation of DFAT cells. Although we hypothesized that stimulation timing relative to confluency would affect the enhancement of chondrogenic differentiation, there were no significant differences among the three conditions (Fig. 3B).
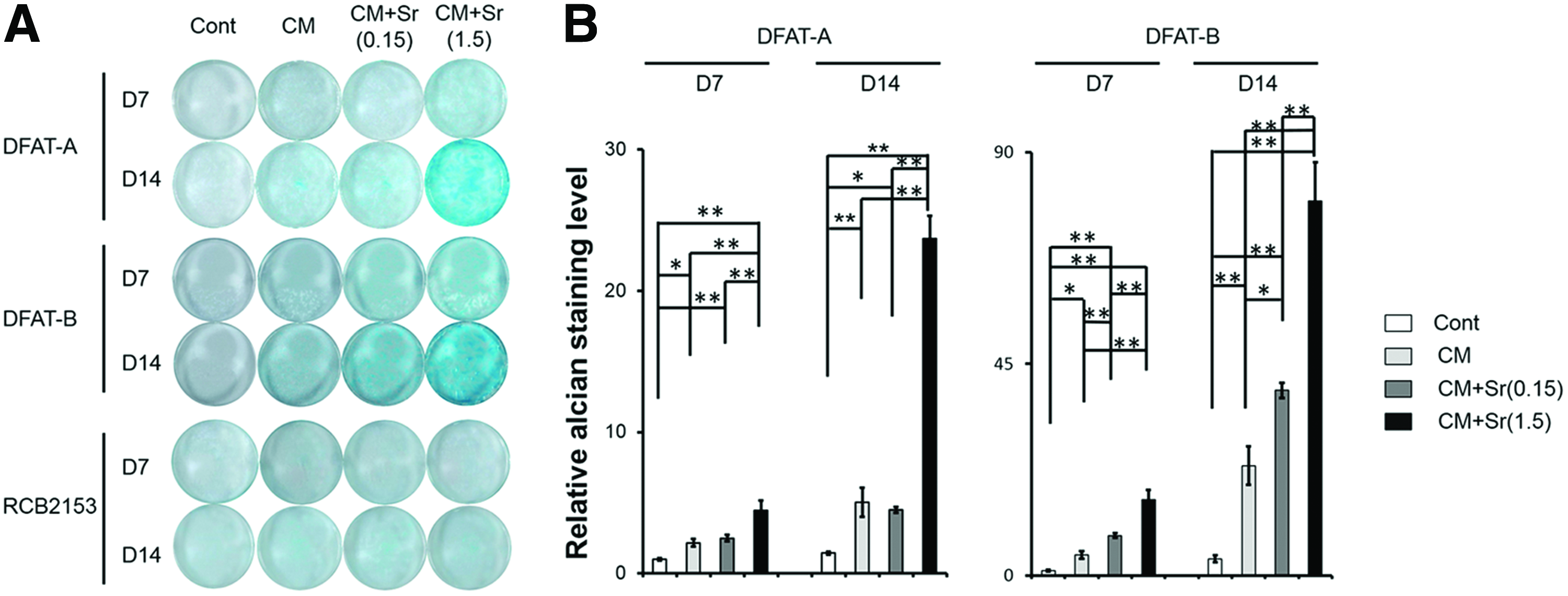
Effect of Sr supplementation in CM for chondrogenic differentiation of DFAT cells and RCB2153 by alcian blue staining
Osteogenic and hypertrophic chondrocyte differentiation
To verify that CM+Sr did not induce undesirable osteoblastic and hypertrophic chondrocyte differentiation of DFAT cells, we evaluated mRNA expression of osteoblastic markers, COL1A1, RUNX2, and OCN, and the hypertrophic chondrocyte marker COL10A1 (Fig. 4). There was no change in the expression of any of the markers with CM+Sr.
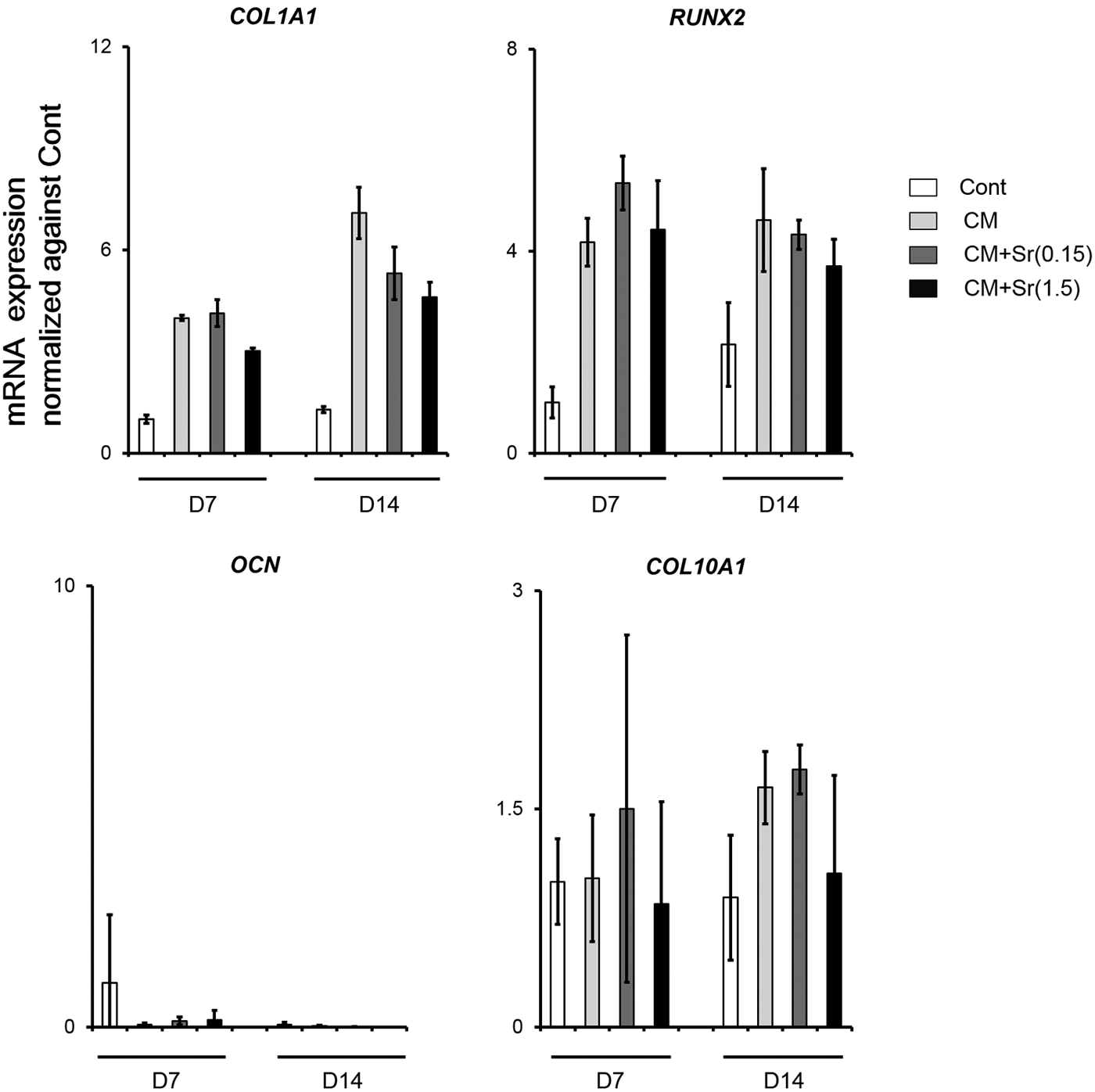
mRNA expression profile of osteoblastic and hypertrophic chondrocyte makers. DFAT-A cells were treated with each medium for 7 and 14 days at cell-seeding condition 2. Compared with CM, CM+Sr(1.5) did not induce marked increment of osteoblastic and hypertrophic chondrocyte differentiation of DFAT cells. The bar graphs show the mean with SD of three independent determinations.
Effect of signaling pathways on chondrogenic differentiation of DFAT cells
Both CM and CM+Sr included abundant growth factors and amino acids. Given that these extrinsic factors could modulate various signaling pathways synergistically, we applied three distinct inhibitors (inhibitors for ERK1/2, Akt, and p38-MAPK signaling pathway) with CM+Sr(1.5) to elucidate the contribution of these pathways to the chondrogenic differentiation of DFAT cells (Fig. 5). Akt and ERK1/2 inhibitors significantly decreased alcian blue staining intensity, whereas inhibition due to the p38-MAPK inhibitor was not significant. These results suggest that the signaling pathways, particularly ERK1/2 and Akt, are involved in chondrogenic differentiation of DFAT cells. Further investigation using western blotting indicated that CM+Sr increased phosphorylation of ERK1/2 (Fig. 7).
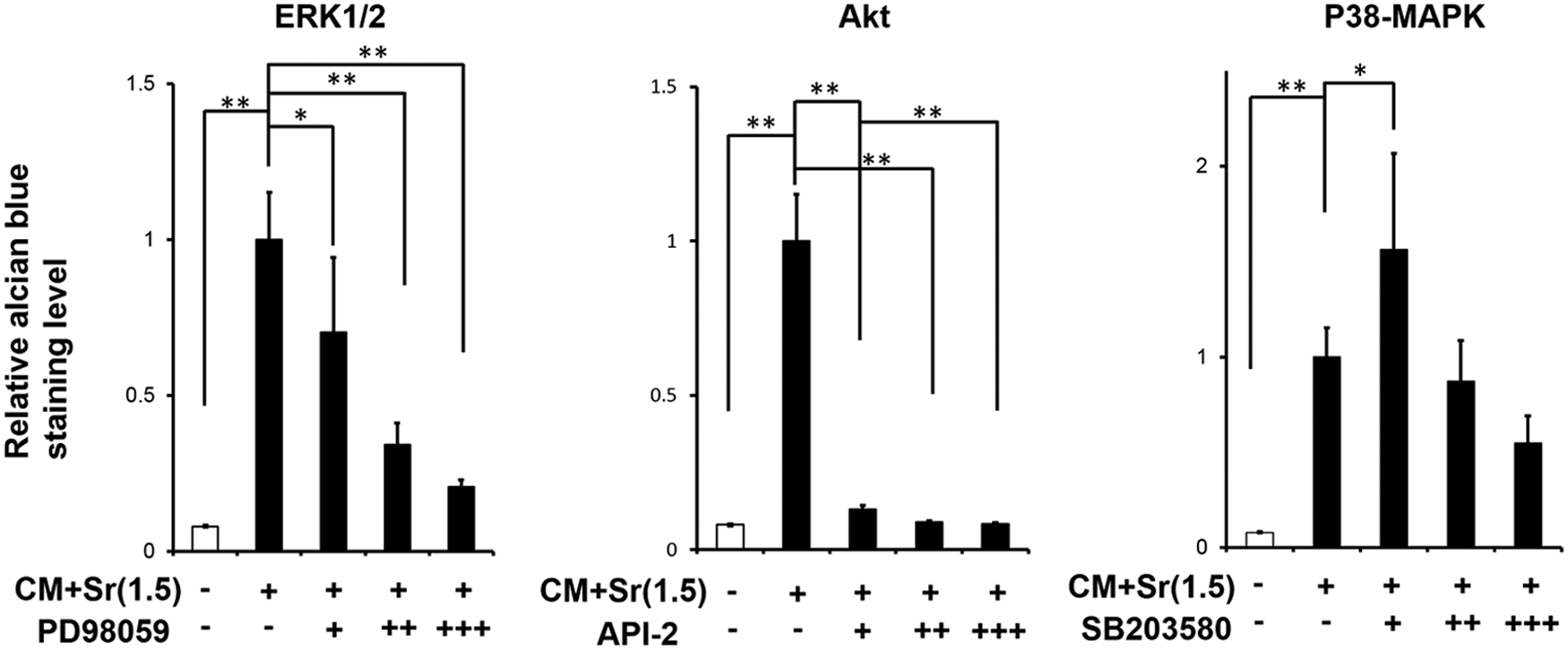
Inhibitory effect of ERK1/2, Akt, and p38-MAPK signaling pathways on CM+Sr-induced chondrogenic differentiation of DFAT-A cells. The concentrations of inhibitors (designated as +, ++, and +++): 2.5, 25, and 50 μM for ERK1/2 inhibitor PD98059; 1, 10, and 20 μM for Akt inhibitor API-2; 1, 10, and 20 μM for p38-MAPK inhibitor SD203580. DFAT-A cells were subjected to CM+Sr(1.5) with/without inhibitors for 14 days at cell-seeding condition 2. All alcian blue staining levels of the samples were normalized against that of DFAT cells subjected to CM+Sr(1.5) without inhibitors. ERK1/2 and Akt inhibitors significantly inhibited CM+Sr(1.5)-induced chondrogenic differentiation, whereas the effect of p38-MAPK inhibitor was subtle. *p<0.05, **p<0.01 (ANOVA with Dunnett's test). The bar graphs show the mean with SD of three independent determinations. ERK1/2, extracellular signal-regulated kinase 1/2.
Effect of CaSR for strontium-induced chondrogenic differentiation
CaSR is a known receptor for Sr ions. 30 To assess the involvement of CaSR-related signaling pathway in chondrogenic differentiation of DFAT cells induced by CM+Sr, we used the CaSR inhibitor NPS2143. We confirmed the presence of CaSR in DFAT cells by immunohistochemical staining and qPCR (Fig. 6A, B). mRNA expression of CaSR was decreased in response to CM+Sr-induced chondrogenic differentiation. Administration of NPS2143 significantly and dose dependently reduced alcian blue staining (Fig. 6C), indicating that chondrogenic differentiation of DFAT cells was partially enhanced by the CaSR pathway. However, the administration of CaSR inhibitor did not change phosphorylation of ERK1/2 (Fig. 7), suggesting that Sr-induced ERK1/2 activation is independent of the CaSR-related signaling pathway.
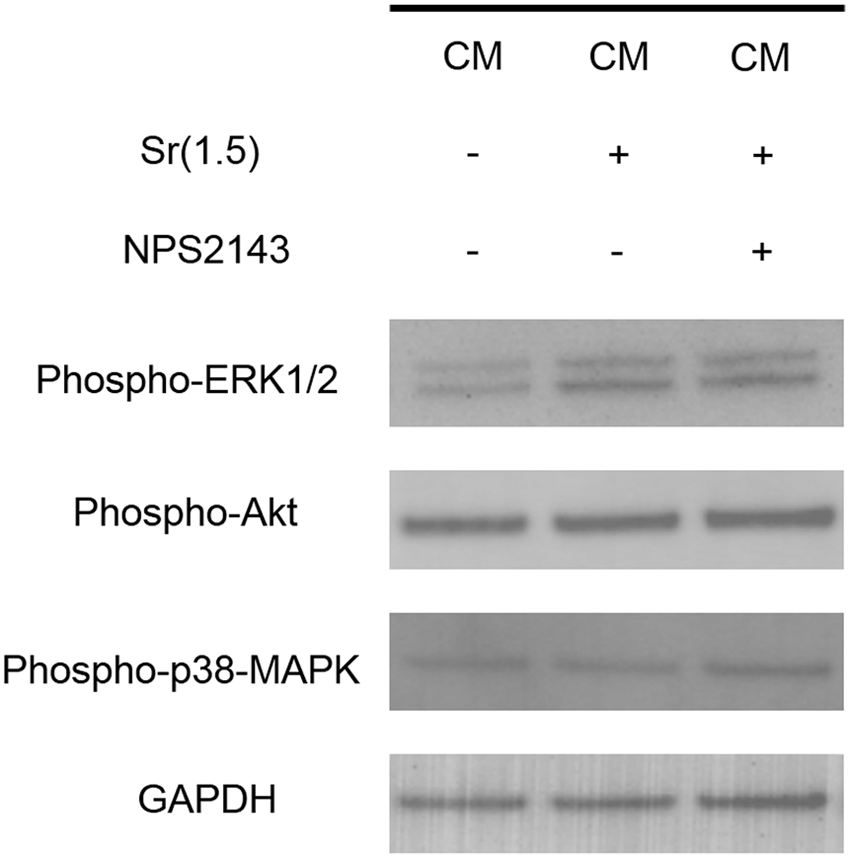
Western blotting of phospho-ERK1/2, phospho-Akt, and phospho-p38-MAPK level in DFAT-A cells treated with the CM with/without Sr and CaSR inhibitor NPS2143. Supplementation of Sr in the CM activated ERK1/2 pathway, whereas its effect was negligible for the other two pathways. No inhibitory effect for the phosphorylation of ERK1/2 could be observed by treating with NPS2143.
Tissue reconstruction capability of chondrogenically differentiated DFAT cells
Using the cell sheet technology and SCID mice, we evaluated the cell behavior of DFAT-derived chondrogenic cells implanted subcutaneously in vivo (Supplementary Fig. S5A–D). Chondrogenically differentiated DFAT cells induced by CM+Sr(1.5) represented stronger safranin O and alcian blue staining, than staining induced by the CM, even at 3 months. However, the tissue morphology showed a reticulated structure, rather than dense and large cartilage-like tissue. The results suggest that chondrogenically differentiated DFAT cells induced by CM+Sr(1.5) are likely to have a potential to fabricate cartilage tissue, even in vivo, although further improvement should be imperative, to explore the optimal implant procedure to utilize the cells in cartilage regeneration therapy.
Discussion
In this study, we provide evidence that Sr works as a stimulant for early chondrogenic differentiation of DFAT cells when used with an appropriate CM. The extent of the chondrogenic induction was markedly higher in CM+Sr(1.5) than that in CM similar to the previous report. 9 CaSR and ERK1/2 pathways were independently associated with the activation.
Sr is a common ingredient of oral drugs for osteoporosis due to its potency for promoting osteoblastogenesis of stem cells25,26 and bone cells. 24 Nevertheless, our results show that, when used with a CM, no significant enhancement of osteoblastogenesis was observed in the expression of osteoblastic markers, COL1A1, RUNX2, and OCN (Fig. 4). Meanwhile, CM+Sr(1.5) significantly elevated COL2A1 expression and GAG synthesis designated with alcian blue staining. This discrepancy may be explained by the use of combinatory CM+Sr containing multiple ingredients, including transforming growth factor-β3 (TGF-β3). TGF-β3 induced chondrogenic differentiation of MSC. 31 This induction is robustly retained even under stimulation with potent osteogenic factors, such as bone morphogenic protein 2. 32 The cross talk between TGF-β and other pathways, such as MAPK, Akt, Wnt, and Hedgehog, has been extensively investigated 33 ; the MAPK pathway, including ERK1/2, can synergistically regulate TGF-β-mediated cell responses, including cell differentiation.33,34 Sr activates ERK1/2 in various cells and our findings consistently suggest that the cross talk between Sr-induced ERK1/2 activation and TGF-β3-related pathway may synergistically lead to early chondrogenic differentiation of DFAT cells.25,26
CaSR is a G-protein-coupled receptor localized on the cell membrane that senses extracellular calcium and modulates intra- and extracellular function. 35 The activation of CaSR regulates the ERK1/2 pathway in numerous different cell types. 36 In this study, we demonstrated that CM+Sr activated both CaSR-related and ERK1/2 signaling pathways, resulting in chondrogenic differentiation of DFAT cells (Figs. 5–7). However, there does not appear to be a cross talk between the Sr-mediated activation of the CaSR pathway and ERK1/2 activation (Fig. 7). Although the detailed mechanism remains unknown, CaSR pathways modulate various other signaling pathways, such as cAMP, JNK, and IP3. 37 The CaSR-related and ERK1/2 pathways often independently regulate cell function. 38 Two different CaSR-related and ERK1/2 pathways activated in this study can be independently involved in early chondrogenic differentiation of DFAT cells.
The expression of SOX9, a key transcription factor in cartilage formation, 39 was less in the cells induced by CM+Sr(1.5) than the expression by CM, in the present study. Nevertheless, compared with CM, CM+Sr(1.5) apparently enhanced further chondrogenic differentiation for the DFAT cells, both in vitro and in vivo. Kypriotou et al. reported that Sox9 exerts a bifunctional effect on COL2A1 expression in dedifferentiated chondrocytes; low levels of SOX9 overexpression can enhance potent COL2A1 expression more than high overexpression levels. 40 More recently, epigenetic mechanisms have been shown to modulate the transcription activity of Sox9,41,42 resulting in the facilitation of chondrogenic differentiation. It is known that the cross talk of signaling pathways modulates the epigenetic mechanisms. 43 Unfortunately, we could not elucidate the complete mechanism for the chondrogenic capability of CM+Sr. These SOX9 levels and epigenetic mechanisms might be related to the potent chondrogenic capability of CM+Sr(1.5).
Similar to MSCs and ASCs, DFAT cells are multipotent for osteogenic, chondrogenic, and adipogenic differentiation. However, we have demonstrated that CM+Sr(1.5) enhanced the alcian blue staining intensity for both DFAT-A and -B cells isolated from the adipose tissues oriented from a different site, but not for RCB2153 (Fig. 2). We previously suggested that the isolated human DFAT cells represented significant early osteoblastogenesis superior to that of human MSCs and ASC.11,22 Furthermore, we recently reported that, despite using the same ingredients, the composition of the medium decisively alters the cell fate when using human MSCs, the osteoblastogenesis model, and ∼60 million candidates of media theoretically. 44 The series of experiments indicate that the characteristics of DFAT cells, while similar, differ from that of MSCs and ASCs, suggesting that specific and optimal media are essential for the efficient application of DFAT cells to tissue engineering and biological assays.
As mentioned above, DFAT cells and ASCs represent different cell characteristics, while both cells can be obtained from the same adipose tissue. ASCs are also a prospective cell source for chondrogenic regeneration therapy. However, its superiority for cartilage regeneration therapy is still controversial. Lee et al. have reported that there was no obvious superiority for cartilage therapy when using ASC-derived chondrocytes cultured in a different type of CM without Sr,45,46 suggesting that exploring alternative cell sources, instead of ASCs, would be worthwhile, to advance cartilage regenerative medicine.
So far, autologous chondrocytes are widely applied as the practical method of chondrogenic regenerative medicine 47 ; while a long expansion phase to secure an adequate cell population is likely to cause dedifferentiation of the cells. 48 Although MSCs obtained from various tissues seem to be an alternative cell source, the application of the cells remains controversial because chondrogenically differentiated MSCs often cause endochondral ossification rather that mature static cartilage tissue in vivo.3,48 Considering the high accessibility of adipose tissue, preparing abundant chondrogenic cells from DFAT cells may be feasible. Chondrogenically differentiated DFAT cells can be a promising cell source for cartilage regenerative medicine in orthopedic, oral, and maxillofacial regions, such as nasal deformity of cleft lip and palate. 47 In this study, we have proposed a method to advance the preparation of early chondrogenically differentiated DFAT cells. However, a further cautious investigation is essential to verify the fate of the cells in vitro and in vivo to fulfill safe and reliable cartilage regeneration therapy using DFAT cells.
Conclusion
While there have been extensive and worthwhile studies regarding osteoblastogenesis of DFAT cells, very few attempts have been made for chondrogenic differentiation. Our results demonstrate that Sr is a prospective stimulant to dose dependently induce early chondrogenic differentiation of DFAT cells if used with an appropriate CM. To the best of our knowledge, this study is the first trial to explore the signal pathways associated with chondrogenic differentiation of DFAT cells. CaSR-related and ERK1/2 signaling pathways were independently involved in this process. The supplementation of Sr in the CM can be a promising technique to provide chondrogenically differentiated DFAT cells for cartilage regenerative medicine.
Footnotes
Acknowledgments
This study was supported, in part, by a Grant-in-aid (25861904) from the Japan Science and Technology Agency. We thank H. Kawaki (Department of Oral Biochemistry, Asahi University School of Dentistry) for technical advice for western blotting, F. Sakamoto (Department of Orthodontics, Osaka Dental University) and A. Nishio (Second Department of Oral and Maxillofacial Surgery, Osaka Dental University) for technical support.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.