
Abstract
In vitro differentiation of stem cells into specific cell lineages provides a stable cell supply for cell therapy and tissue engineering. Therefore, understanding the mechanisms underlying such differentiation processes is critical for generating committed lineage-specific cell progenies effectively. We previously developed a two-step protocol to differentiate mesenchymal stromal cells (MSCs) into hepatocyte-like cells. Since hepatic differentiation involves mesenchymal–epithelial transition (MET), we hypothesize that promoting MET could further accelerate the differentiation process. Ras-related C3 botulinum toxin substrate 1 (Rac1) is involved in actin polymerization and its role in MET was investigated in the study. Our results showed that inhibition of Rac1 activation by Rac1-specific inhibitor, NSC23766, led to cells favoring epithelial morphology and being more packed during hepatic differentiation. In addition, Rac1 inhibition accelerated the upregulation of hepatic marker genes accompanied by more mature hepatic functions. Taken together, promotion of MET by inhibiting Rac1 accelerates the hepatic differentiation of MSCs. Our findings open a new prospect of directing the commitment of MSCs by manipulating cell morphology and cytoskeleton arrangement through small molecules. The results provide further insight into scaffold design for rapid production of MSC-differentiated hepatocytes.
Introduction
S
Liver development or hepatogenesis from hepatic stem cells is a process that involves mesenchymal–epithelial transition (MET). 8 Comparative transcriptome analysis demonstrated that expression of mesenchymal markers, Twist and Snail, decreases when MSCs differentiate into hepatocyte-like cells, indicating that MET is involved during hepatic differentiation of MSCs. 9 However, hepatic growth factor (HGF) and fibroblast growth factor-2 (FGF2) are two growth factors added in the induction medium of a two-step protocol for hepatic differentiation of MSCs 1 ; both of them are potent modulators for epithelial (endothelial)–mesenchymal transition (EMT)10,11 and may oppose MET and attenuate the hepatic differentiation of MSCs. HGF and FGF2 are key factors associated with specification of the endoderm, which is the developmental origin of hepatocytes.12,13 HGF also plays an important role in hepatogenesis after partial hepatectomy. 14 Given that, these two factors should be beneficial for hepatic differentiation of MSCs. Hence, our strategy to accelerate hepatic differentiation based on the existing two-step protocol is to investigate HGF- and FGF2-mediated downstream pathways and specifically block the one responsible for epithelial–mesenchymal transition.
Ras-related C3 botulinum toxin substrate 1 (rho family, small GTP-binding protein Rac1) activity is required for FGF2-induced endothelial–mesenchymal transformation of corneal endothelial cells. 15 Rac1 activation is also required for HGF-stimulated mesenchymal phenotypes, such as loss of cell–cell adhesion, increase of cell motility, and formation of dynamic protrusions in MDCK epithelial cells.16,17 Moreover, Rac1 activity is essential for HGF-induced migration and stress fiber formation of MSCs; once Rac1 activity is inhibited, MSCs lose HGF-induced motility and exhibit a more rectangular shape. 18 Therefore, Rac1 is a potential target for the purpose of blocking HGF- or FGF2-induced mesenchymal phenotypes. We hypothesize that Rac1 inhibition can accelerate mesenchymal–epithelial transition through opposing HGF- or FGF2-induced mesenchymal phenotypes and thus promote hepatic differentiation of MSCs.
In addition to chemical induction, physical cues such as substrate stiffness,19,20 applied strain, 21 and cell shape (via micropatterning)22,23 can regulate stem cell commitment as well. These physical stimuli pass from the extracellular environment to intracellular cytoskeleton systems and thus the rearranged cytoskeleton provides spatial and temporal cues to switch on and off the downstream signal cascades. 24 Since there is correlation between hepatic differentiation and MET, we speculated that directly manipulating MET through cytoskeleton remodeling can affect the differentiation process as well. In addition to being downstream signaling of HGF and FGF2 pathways, Rac1 also plays roles in several cellular processes, including cell–cell adhesion and motility through actin reorganization. 25 For example, an elongated mesenchymal-like cell migration, different from a rounded ameboid-like one, is driven by Rac1 activation. 26 In the study, we demonstrated that Rac1 inhibition can lead to the accelerated presence of epithelial phenotypes and promotes hepatic differentiation of MSCs due to cytoskeleton remodeling partially, if not wholly. Our study provides an alternative approach to manipulate MET or EMT through actin remodeling using small molecules other than physical methods and it may be more practical for clinical application.
Materials and Methods
Cell maintenance and expansion
Mouse mesenchymal stromal cells (mMSCs) were derived from the bone marrow of a postnatal 7–8-week-old BALB/c mouse. The isolated cells were assessed by the differentiation assays and characterized by surface markers, CD11b, CD29, CD31, CD34, CD44, CD45, CD73, CD105, CD106, and Sca1, using flow cytometry before being further used for the study. Human mesenchymal stromal cells (hMSCs) were purchased from Cell Applications, Inc., which were collected from a healthy, young female donor. For maintenance and expansion, mMSCs were cultured in low-glucose Dulbecco's Modified Eagle's Medium (Sigma-Aldrich) containing 10% fetal bovine serum (Invitrogen) and 100 U penicillin, 100 μg/mL streptomycin, and 2 mM L-glutamine (1% PSG; Sigma-Aldrich). Cells of 10th to 14th passage were selectively used for the experiments. hMSCs were cultured in Iscove's modified Dulbecco's medium (IMDM; Invitrogen) containing 10% fetal bovine serum, 1% PSG, 10 ng/mL FGF2 (Sigma-Aldrich), and 10 ng/mL epidermal growth factor (EGF). Both kinds of cells were seeded at the confluency of 30% to 40%. Cells were replated under the same culture conditions once they reached confluency around 70%, ∼15,000 cells/cm2.
Hepatic differentiation
Hepatic differentiation was based on the two-step protocol established by our group described previously. 1 Briefly, cells at 90% confluency or higher, which were ∼60,000–70,000 cells/cm2, were washed with phosphate-buffered saline (PBS) twice and incubated in step1 induction medium, which is IMDM supplied with 20 ng/mL HGF (R&D system), 10 ng/mL FGF2, 0.61 g/L nicotinamide (Sigma-Aldrich), and 1% PSG, for 7 days. After 7 days of induction, cells were washed twice with PBS and changed to a step2 maturation medium consisting of IMDM supplemented with 20 ng/mL oncostatin M (Prospec), 1 μM dexamethasone (Sigma-Aldrich), 1× ITS+ premix (BD Biosciences), and 1% PSG for 2 weeks. The differentiation protocol of hMSCs is similar to that of mMSCs, with differences in step 2 maturation medium consisting of 0.2% fetal bovine serum and 2 ng/mL HGF, and the duration of step 2 treatment was prolonged to 3 weeks. The medium were changed twice a week.
Scratch wound-healing assay
Cells were cultured in dishes and grown to confluence. A uniform scratch was made by a 200-μL sterile pipette tip, and detached cells were removed by PBS wash. Images were taken after wound scratching using phase-contrast microscopy at the same sites. The recovery rate of gap represents the migration ability of cells and is in a reverse correlation with the percent of the original wound size.
Real-time polymerase chain reaction
Total RNA was extracted by the RNeasy Mini Kit (QIAGEN). Two micrograms of total RNA was reverse transcribed to complementary DNA by MMLV high-performance reverse transcriptase according to the manufacturer's instructions (EPICENTRE). Quantitative real-time polymerase chain reaction (RT-PCR) was performed with Fast SYBR® Green Master Mix (2×) (Applied Biosystems) for mMSCs and with TaqMan® Fast Universal PCR Master Mix (2×) (Applied Biosystems) for hMSCs to determine the relative hepatic gene expression in differentiating cells. The following primers were used for quantifying mouse cDNA: GGGA AGCCCATCACCATCT/CGGCCTCACCCCATTTG for GAPDH as internal control, CACACCCGCTTCCCTCAT/CAAACTCATTTTCGTGCAATGC for AFP, CGAGCCA TTGTGGACAACAT/GGTCCCCAATTGACAGAGAGAT for TAT, CCATGGACTCCGCAAGGT/CTTCCTTGAGT GCCTCGATTTCT for CK18, GAGGCTCCCCTGAGAAT AGACA/TGTTTGGTGTGAAGGTCATGATTAG for HNF4A, GCCTCCGGAAGTATTGTCTCATC/CACCCCT AGCCCTTTTAGTAGCA for G6PC, and CGGTTTGA CTTGGTATCCCTTT/CCGGAAGGACCGGATGTC for EpCAM. The following primers were used for quantifying human cDNA: TGCGTAATCGATAAGGAAACC/TGGTA GGCACGCTCAGTCT for RPS11 as internal control, AATGTTGCCAAGCTGCTGA/CTTCCCTTCATCCCGA AGTT for ALB, AGCAACGGACAGATGTGTGA/TCAG ACCCTGAGCCACCT for NF4A, CGATGACAGCCTTG GACTTC/CGGAATTGCAAACTCTGGA for TDO2, TCT CGCGGCCTAGCTTTAC/AGTGGGAACTTGCTGAGGTG for GLUL, and AGAACCCCAAAGGCTTCTTC/CTTGGC TTTTCCTTCATGGT for ALPL.
siRNA transfection of Rac1
Fifty nanomolars of siRNA targeting for mouse Rac1 (GGC GAA AGA GAU CGG UGC UGU CAA A) or nonspecific siRNA control was transfected with RNAiMAX transfection reagent (Invitrogen) into mMSCs 1 day before the hepatic induction for 24 h according to the manufacturer's instructions (Invitrogen), and the transfection was performed again on the 6th day after the induction.
Periodic acid-Schiff stain for glycogen
Cells in culture dishes were washed twice with PBS and fixed with 3.7% formaldehyde (Sigma-Aldrich) for 20 min at room temperature. After washing twice with PBS, cells were permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 10 min. Samples were then washed twice with PBS and oxidized with 1% periodic acid (Sigma-Aldrich) for 5 min. Next, samples were rinsed thrice with deionized distilled water and treated with Schiff's reagent (Sigma-Aldrich) for 20 min in the dark. After washing with deionized distilled water thrice, samples were ready for observation.
CYP3A4, urea, and albumin functional assays
Enzyme activity of cytochrome P450, family 3, subfamily A, polypeptide 4 (CYP3A4) was measured using P450-Glo™ CYP3A4 Assay (Luciferin-PFBE) (Promega). The secreted urea level was determined using colorimetric assay (BioVision). The secreted albumin level was determined by immunoperoxidase assay (Immunology Consultants Laboratory). The assays were performed according to the manufacturer's instructions.
Statistical analysis
All data for statistical analysis were derived from at least three independent experiments and are presented as mean±SEM. Student's t-test was used to compare data from two groups. A p-value less than 0.05 was considered as statistical significance. The * indicates p<0.05, ** indicates p<0.01, and *** indicates p<0.001.
Results
MSCs demonstrated morphological transition and decreased in motility during hepatic differentiation
In our hepatic differentiation platform, a dramatic morphological transition was observed both in hMSCs and mMSCs. For hMSCs, upon induction with step 1 medium, undifferentiated fibroblast-like cells started to elongate and maintained the spindle-like shape throughout the step 1 stage. During the period of step 1 induction, there were fewer cell–cell contacts. However, after they were transferred to the step 2 medium, cells started to have contacts with their neighboring cells and demonstrated an epithelial polygonal morphology with ordered arrangement (Fig. 1A). For mMSCs, the phenomena of morphological transition were very similar to those of hMSCs, except faster epithelia-like maturation. In the step 1 medium, mMSCs started to shrink their cell bodies with less adhesive contact to the dish bottom as well as demonstrated long and thin stretched protrusions. Few days after step 1 induction, cells retracted their protrusions and presented a polygon-like shape. Once they were transferred to the step 2 medium, cells began to contact each other compactly and became much more arranged as time proceeded (Fig. 1B). In summary, a transition from mesenchymal-like to epithelial-like morphology occurred during hepatic differentiation for both hMSCs and mMSCs. In addition, loss of cell motility during hepatic differentiating is another indication of MET. Scratch wound-healing assay revealed that cell motility was decreasing as differentiation proceeded, with the highest mobility in undifferentiated MSCs, while almost immobilization for terminally differentiated hepatocyte-like cells (Fig. 1C). Taken together, our results were consistent with the previous findings that hepatic differentiation of MSCs is a process associated with MET. 9
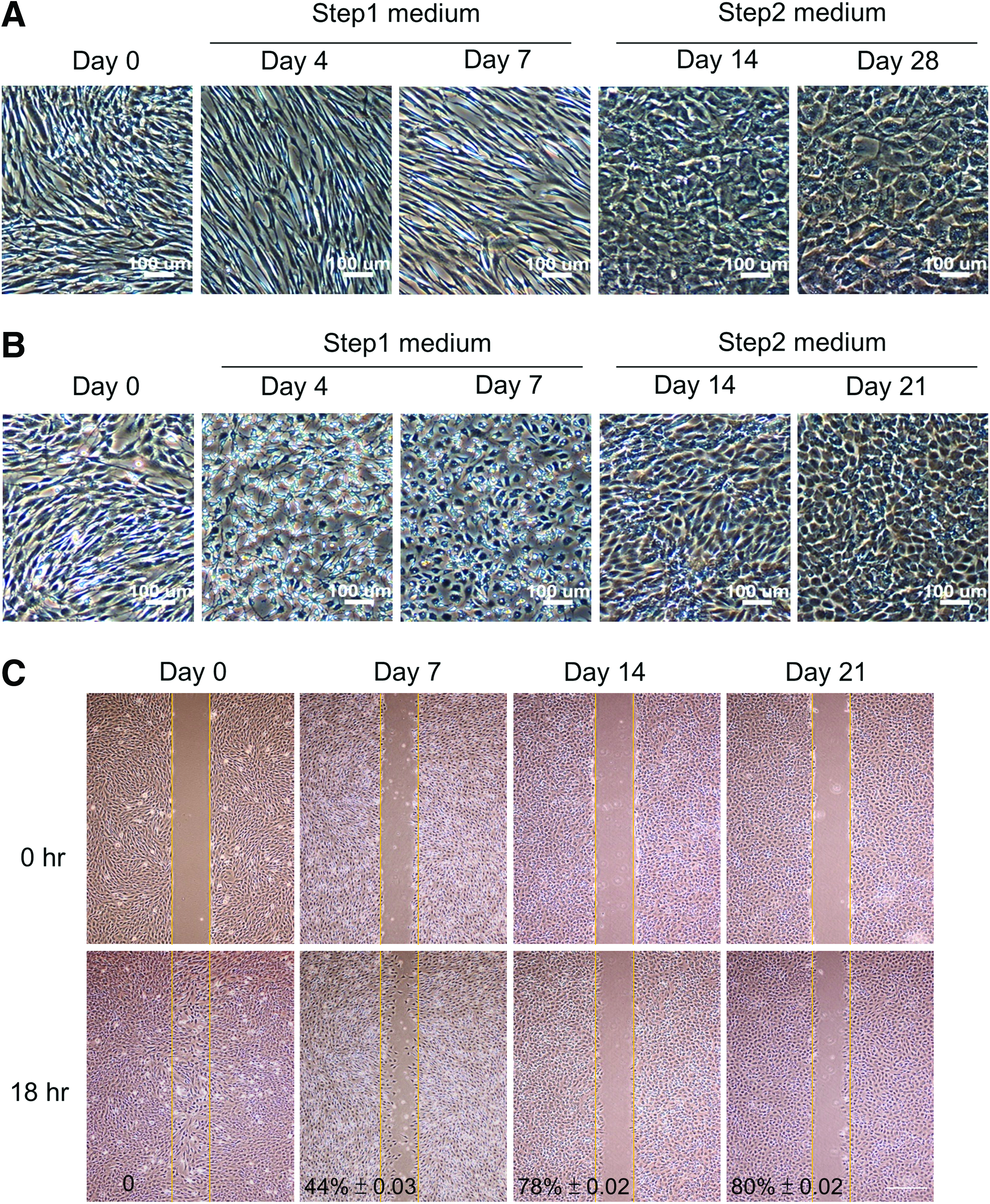
Morphological transition and decreased cell motility during hepatic differentiation of MSCs.
Rac1 inhibition accelerated mesenchymal-to-epithelial morphological transition in the early stage of hepatic differentiation of MSCs
NSC23766, a pyrimidine compound that specifically inhibits Rac1 GDP/GTP exchange activity, but has no effects on Cdc42 or RhoA activities, was used as Rac1 inhibitor in the study to test if Rac1 inhibition promotes hepatic differentiation of MSCs. NSC23766 showed no toxicity to mMSCs undergoing hepatic differentiation up to 20 μM (Fig. 2A). Upon treatment of the Rac1 inhibitor, mouse cells exhibited a more epithelial morphology with fewer stretching protrusions in step 1 induction medium compared with the control group, and the difference in morphology gradually disappeared after entering the step 2 maturation stage (Fig. 2B). The decrease of stretching protrusions is in an NSC23766 dose-dependent manner (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). To quantify the morphological differences between the two groups, we counted the cells with one or more protrusions longer than their cell bodies, which is considered as a typical characteristic of the mesenchymal phenotype. The result showed that there were about 70% cells with protrusions in the control group on day 2, while the percentage was significantly lower in the Rac1 inhibitor-treated group with just 30%. On day 7, the percentage in the Rac1 inhibitor-treated group was further decreased to 15%, while it still remained up to 55% in the control group. The morphological difference disappeared on day 14 (Fig. 2C). Similar phenomena were observed in hMSCs undergoing hepatic differentiation, with a morphological difference on day 14 and no difference in the late stage of differentiation (Fig. 2D). These results suggest that Rac1 inhibition can accelerate differentiating cells into a more epithelial morphological state.
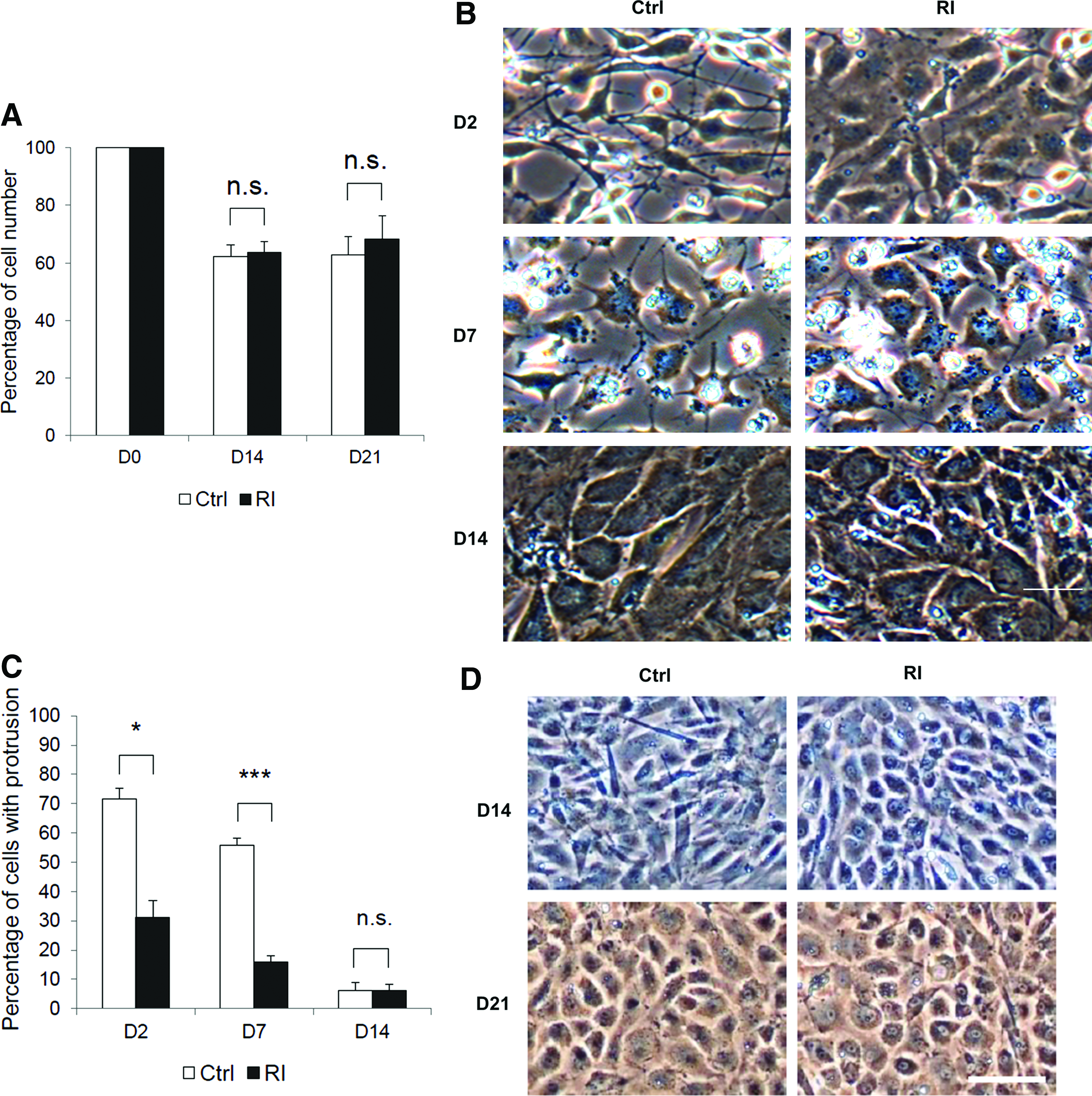
Rac1 inhibition accelerated morphological transition from mesenchymal phenotype to epithelial phenotype.
Rac1 inhibition upregulated the expression of hepatic cell-specific marker genes during hepatic differentiation of MSCs
Since hepatic differentiation of MSCs is a process associated with MET, we proposed that Rac1 inhibition-accelerated MET would be beneficial to the differentiation process. We quantified the expression of hepatic genes in the control group and Rac1 inhibitor-treated group during hepatic differentiation of mMSCs by quantitative real-time PCR. The result showed that the expression of hepatic cell-specific genes, including alpha-fetoprotein (AFP), tyrosine aminotransferase (TAT), cytokeratin 18 (CK18), hepatocyte nuclear factor 4 alpha (HNF4A), glucose-6-phosphatase catalytic subunit (G6PC), and epithelial cell adhesion molecule (EpCAM), in the NSC23766-treated group was two- to three-fold higher than that in the control group on day 14. Moreover, the level of gene expression in the Rac1-inhibited group on day 14 was comparable with that in the control group on day 21, which indicated an early hepatic maturation of NSC23766-treated cells (Fig. 3A). Consistent with the results of Rac1 inhibition, RT-PCR results of Rac1 gene knockdown showed that the expression of hepatic cell-specific genes was significantly higher compared with the control group on day 14 (Fig. 3B). Rac1 inhibition also promoted the gene expression of albumin (ALB) and accelerated the upregulation of HNF4A gene expression in hepatic differentiation of hMSCs (Fig. 4).

Both Rac1 inhibition and knockdown accelerated the expression of hepatic marker genes during hepatic differentiation of mMSCs.
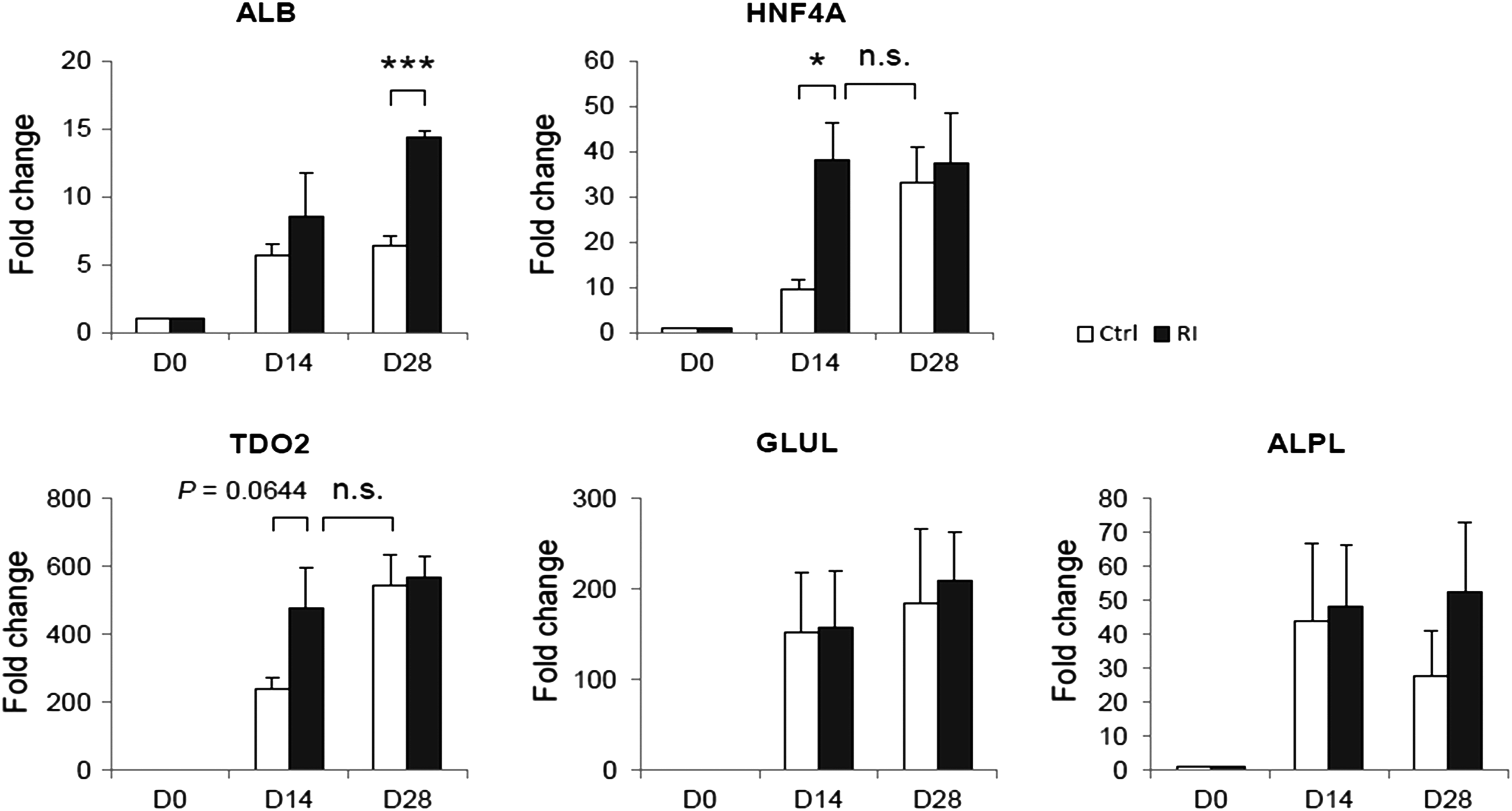
Rac1 inhibition accelerated the expression of hepatic marker genes during hepatic differentiation of hMSCs. Relative expression of hepatic marker genes, ALB, HNF4A, TDO2, GLUL, and ALPL without (white bar) or with the treatment of 20 μM Rac1 inhibitor (dark bar) on days 0, 14, and 28 during hepatic differentiation of hMSCs. * Indicates p<0.05 and *** indicates p<0.001. p-value less than 0.05 was considered as statistical significance. n.s., not significant.
AFP is a plasma protein produced abundantly by fetal hepatocytes. In contrast to AFP, ALB is the main plasma protein produced by hepatocytes after birth. In addition, HNF4A is a transcription factor of HNF1A, which regulates the expression of several hepatic genes. It is also a key factor to maintain differentiated hepatocyte functions in 3D culture. 27 TAT and glucose-6-phosphatase are enzymes expressed actively in the liver. Therefore, the results suggest that Rac1 inhibition has positive effects on hepatic differentiation. Moreover, the upregulation of CK18 and EpCAM reconfirmed that Rac1 inhibition promotes MET in terms of intermediate filaments and cell–cell adhesion in addition to cell morphology.
Rac1 inhibition upregulated the expression of hepatic cell-specific proteins during hepatic differentiation of mMSCs
We further performed immunostaining and western blotting to investigate the effect of Rac1 inhibition on the expression of hepatic cell-specific proteins during hepatic differentiation of mMSCs. Protein expression of AFP was significantly higher in the Rac1-inhibited group than that in the control group on days 14 and 21. The expression of albumin was significantly higher in the Rac1-inhibited group than that in the control group on day 14. Both control and inhibited groups had a similar albumin expression level on day 21 (Fig. 5A, B). Results from western blot analysis were consistent with the results from immunostaining (Fig. 5C). Semiquantitative analysis of western blots showed that the albumin protein ratio after being normalized by GAPDH was 0.6: 1.2: 1.0: 1.0 as of the control group on day 14: inhibitor-treated group on day 14: control group on day 21: inhibitor-treated group on day 21.
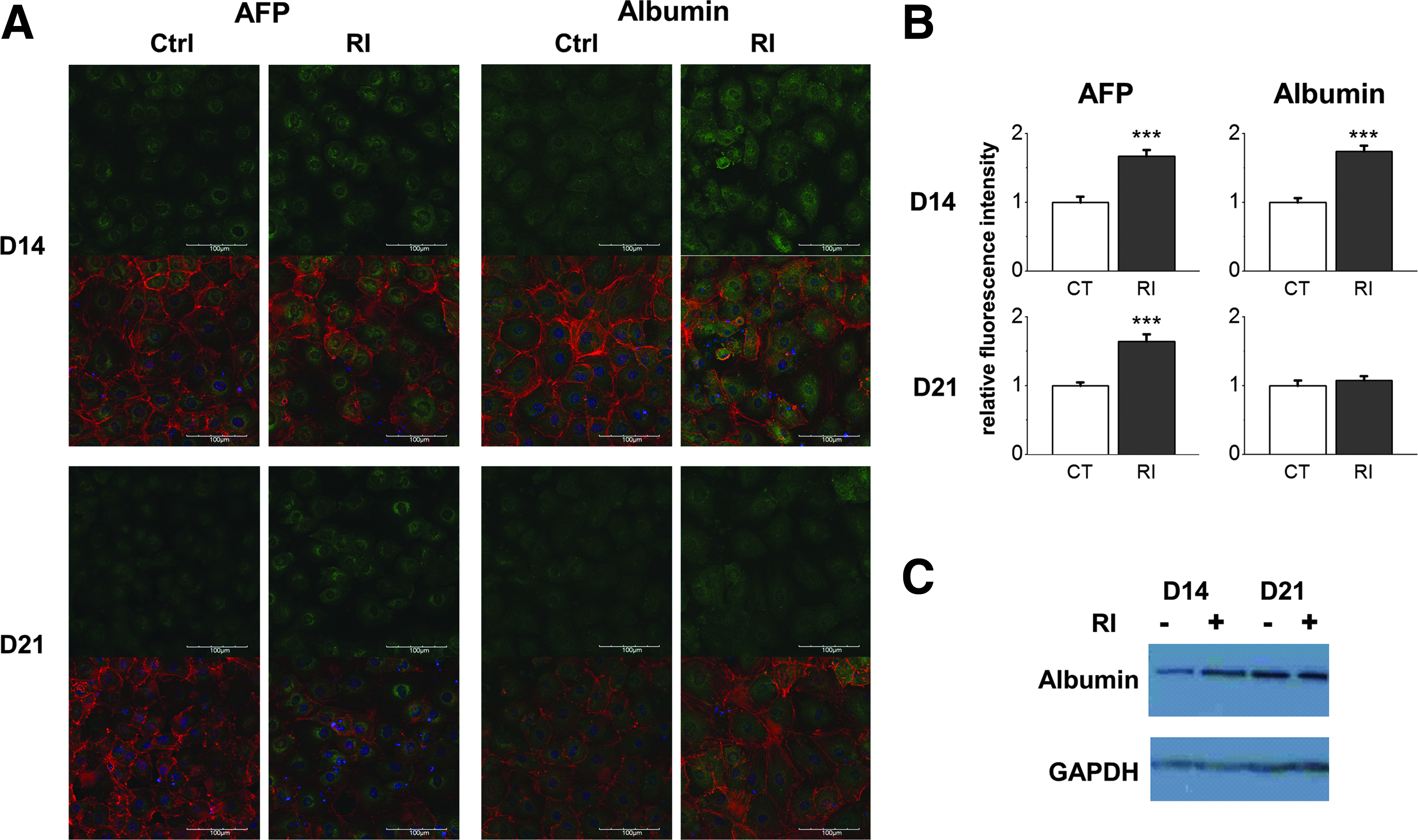
Rac1 inhibition upregulated protein expression of albumin and AFP during hepatic differentiation of mMSCs.
Rac1 inhibition promoted hepatocyte-related functions of differentiating cells
In addition to the upregulation of gene expression, we further compared the hepatocyte-related functions for evaluating the effect of Rac1 inhibition on hepatic functions. One of the signature functions of hepatocytes is glycogen storage. To detect the amount of glycogen in cells, Periodic acid-Schiff (PAS) stain was applied for the control group and the group with NSC23766 treatment on days 0, 14, and 21 during hepatic differentiation of mMSCs. As we expected, undifferentiated cells did not show any positive stain for glycogen, while almost all cells were positively stained for glycogen on day 14. Most importantly, the intensity of purple color in cells treated with the Rac1 inhibitor was much stronger compared with the control group on day 14 (Fig. 6A).
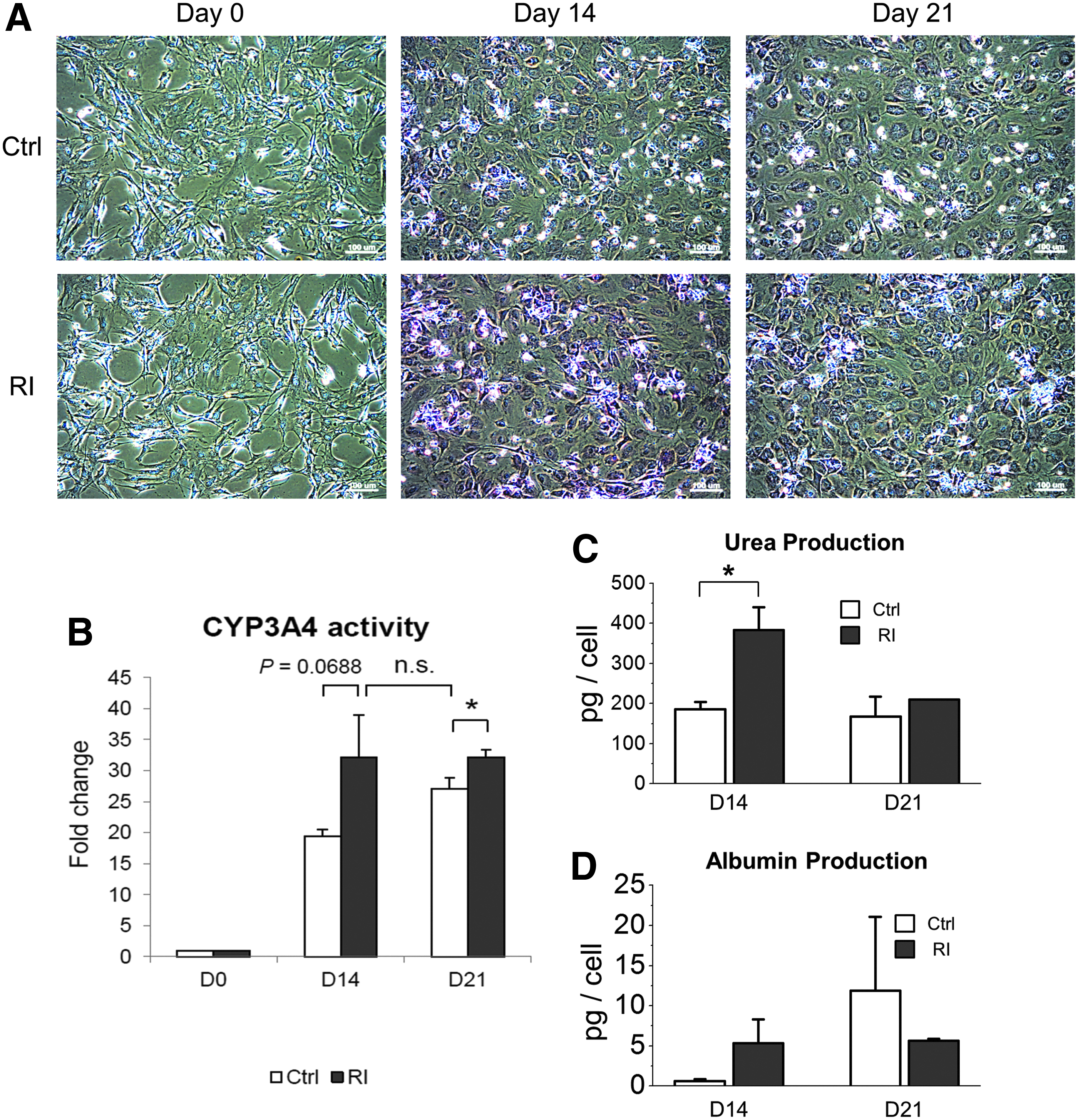
Rac1 inhibition benefited the acquirement of hepatic functions of MSCs undergoing hepatic differentiation.
Another important characteristic function of hepatocytes is the ability to metabolize and detoxify compounds such as drugs or toxic chemicals. Cytochrome P450 superfamily of monooxygenases is a large group of enzymes responsible for catalyzing the oxidation of these substances. Given that, we examined the enzyme activity of CYP3A4, a member of the cytochrome P450 superfamily, which is expressed prominently in the liver. Similar to the result of the glycogen storage experiment, the CYP3A4 activity was upregulated along the differentiation, and the increase of enzyme activity was faster in the Rac1 inhibitor-treated group compared with that in the control group (Fig. 6B).
Urea is the main waste product in urine, produced primarily by the urea cycle during protein catabolism in the liver. The secreted urea level was significantly higher in the Rac1 inhibitor-treated group compared with that in the control group on day 14 during hepatic differentiation of mMSCs (Fig. 6C). We also found that secreted albumin was slightly higher in the Rac1 inhibitor-treated group than that in the control group on day 14 (Fig. 6D). Combining these functional data mentioned above, we concluded that Rac1 inhibition was beneficial for the maturation and functional performance of hepatocyte-like cells derived from MSCs.
Morphological change induced by Rac1 inhibition in the early stage of hepatic differentiation of MSCs was uncorrelated with E-cadherin
In most cases, an epithelial morphology is accompanied with an increase in E-cadherin and/or relocalization of E-cadherin to the cell surface. The accumulation of E-cadherin can strengthen the cell–cell contact and maintain cells in a mature epithelial shape. E-cadherin has been shown to be gradually upregulated during hepatic differentiation of hepatic progenitor cells. 8 Since Rac1 inhibitor, NSC23766, promoted MET in the early stage of hepatic differentiation of MSCs, we were wondering if Rac1 inhibition could also accelerate E-cadherin accumulation for stabilizing newly formed cell–cell contacts, as indicated by the upregulation of the EpCAM gene expression. Immunofluorescent staining was performed for detecting E-cadherin in hepatocellular carcinoma Hep3B (as a positive control), undifferentiated mMSCs, and differentiating mMSCs with or without 2-day treatment of Rac1 inhibitor, NSC23766 (Fig. 7). F-actin was also stained for observing the actin cytoskeleton arrangement. Hep3B showed a typical E-cadherin staining at the sites of cell–cell contact with actin forming a belt-like structure around the cells. For undifferentiated mMSCs, E-cadherin was weakly stained in the whole cell body with actin stress fiber emerged in the cytoplasm, just in agreement with the mesenchymal phenotype of mMSCs. The magnitude of E-cadherin staining was increased after step 1 induction, both in the control group and Rac1 inhibitor-treated group. However, even the distribution of E-cadherin was slightly more spreading in the whole cell body; no accumulation of E-cadherin was observed at contact sites in the epithelial-like cells of the Rac1 inhibitor-treated group. Although there was no significant difference in E-cadherin between the control and Rac1 inhibitor-treated groups, phalloidin staining showed slight differences in actin arrangement between the two groups (Fig. 7). We did not observe differences in E-cadherin and actin staining between the control and Rac1 inhibitor-treated groups at the mid-late maturation stage (Supplementary Fig. S3).
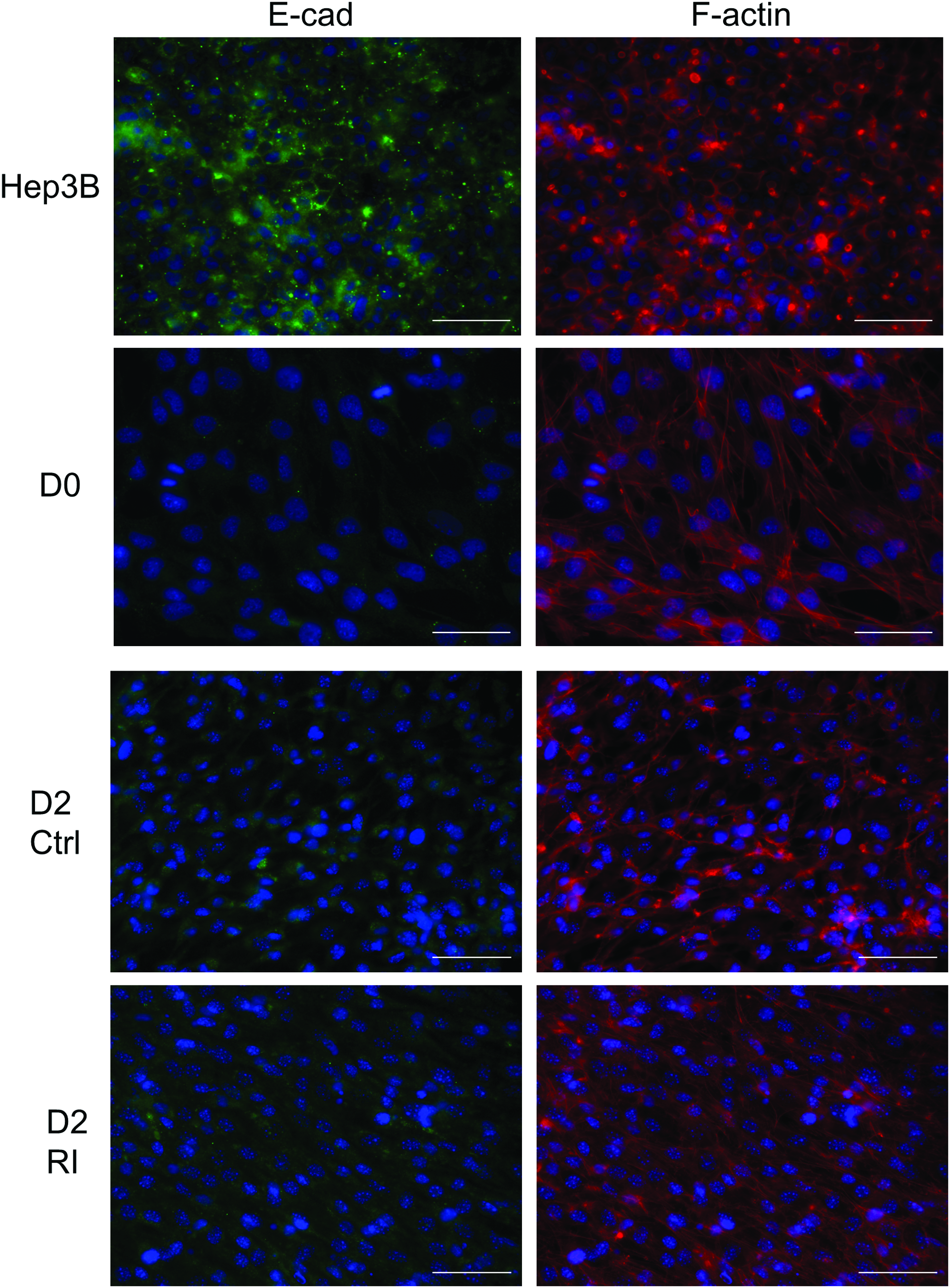
Rac1 inhibition affected actin remodeling, but did not induce E-cadherin localization at contact sites of mMSCs in step 1 medium. The expression and localization of E-cadherin (BD Biosciences), F-actin (Phalloidin), and nucleus (DAPI) were determined by immunofluorescence in Hep3B, undifferentiated MSCs (D0), and 2-day differentiating cells without (Ctrl) or with 20 μM Rac1 inhibitor (RI). Scale bar, 50 μm. Color images available online at www.liebertpub.com/tea
Discussion
The correlation between accelerated morphological transition and promoting effect on hepatic differentiation by Rac1 inhibition suggested that the premature morphological transition plays a role as a physical cue to promote the differentiation process. In fact, dramatic cell shape remodeling during hepatic differentiation of MSCs has been described previously in several articles.4,5,28,29 In our study, we further investigated whether the morphological remodeling can be a cause to affect hepatic differentiation. We found that Rac1 inhibitor, NSC23766, accelerated a mesenchymal-to-epithelial morphological transition in the induction and early maturation stages (day 7), but the morphological difference disappeared in the middle of maturation stage (day 14; Fig. 2). Even though a recent study indicated that NSC23766 may also act as an antagonist of muscarinic acetylcholine receptors, 30 our microarray data indicated that the gene expression of muscarinic acetylcholine receptors was very low in MSCs and did not change much during the hepatic differentiation (data not shown). Therefore, the major inhibition effect of NSC23766 was solely on Rac1. Promotion of MET by Rac1 inhibition was also observed in wound-healing assays of hepatic-engaged MSCs (Supplementary Fig. S2). Rac1 inhibitor, NSC23766, slightly inhibited MSC motility in the early induction stage of hepatic differentiation, but had no effect on motility when the assay was performed in the maturation stage. However, in the mid-late maturation stage, even without the differences in morphology or motility, hepatocyte-related functions were still promoted in the NSC23766-treated group (Figs. 3–6). The temporal order of these incidents suggested that the accelerated morphological transition is the reason rather than just the result of the promotion of hepatic differentiation. In other words, the tension and cytoskeletal network in an epithelial-like morphology favor the epigenetic activation of hepatic genes, and Rac1 inhibition promotes hepatic differentiation through accelerating such epithelia morphology.
In addition to the direct impact on transcriptional regulation of hepatic differentiation by cytoskeletal remodeling through morphological manipulation, downstream MET events caused by Rac1 inhibition also contribute to positive effects on hepatic differentiation. In the early maturation stage of hepatic differentiation, cells began to contact each other compactly and Rac1 inhibition accelerated this phenomenon (Figs. 1 and 2). It is in agreement with the upregulation of EpCAM, indicating the formation of the premature adherens junction (Fig. 3). In fact, the inverse correlation between Rac1 activity and adherens junction or tight junction formation has been reported in many studies.31–33 Formation of these junctions can restrict cell proliferation and favor differentiation through sequestrating transcription factors such as β-catenin at cell contact sites and preventing them from nuclear translocation.34,35 A similar mechanism was also reported in three-dimensional hepatocyte aggregates. Once cell–cell interaction overrides cell–matrix interaction, the niche maintains expression of HNF4A and upregulates downstream hepatocyte-specific functional genes. 27 It is worth noting that lack of the ability to accumulate E-Cadherin at cell contact sites by Rac1 inhibition indicates the potential for further accelerating hepatic differentiation through promoting tight junction formation using other small molecules at the mid-late maturation stage (Fig. 7).
Other than actin remodeling and premature formation of cell–cell contacts, the circuit of Rac1 and mesenchymal markers, Twist and Snail, may be an additional factor to affect the MET acceleration. It has been reported that Rac1 can promote phosphorylation and nuclear localization of Snail through activating its downstream effector PAK-1. 36 Additionally, the reciprocal inhibition between Snail and HNF4A, which is the determining transcriptional factor for hepatic differentiation, was demonstrated previously.37,38 Therefore, the upregulation of HNF4A by Rac1 inhibition may be due to the decrease of phosphorylation and nuclear accumulation of Snail (Figs. 3 and 4). Recent studies also showed that Twist1 activates Rac1 through suppressing microRNA let-7i 39 and this may be through WAVE2 regulation by promoting actin assembly.40,41 Further investigation of the underlying mechanisms is worth pursuing in the future.
In this study, we demonstrated that Rac1 inhibitor, NSC23766, accelerated a morphological transition toward epithelial-like morphology in the early-mid stage of hepatic differentiation of MSCs. Accompanied with such accelerated morphological transition, we also showed that Rac1 inhibitor was able to accelerate the expression of hepatic marker genes and increase storage of glycogen as well as enzyme activity of CYP3A4. The positive correlation between accelerated morphological transition and promoting effect on hepatic differentiation suggests that Rac1 inhibition promotes hepatic differentiation of MSCs through accelerating MET and is related to actin remodeling. Our finding opens a new prospect of regulating stem cell fate commitment by manipulating cytoskeleton arrangement through small molecules other than conventional physical manipulation such as substrate stiffness or micropatterning. The study provides a practical clinical application of cell therapy where rapid production of mature differentiated hepatocytes is required to treat critically ill patients with liver problems.
Footnotes
Acknowledgments
This work was supported in part by the UST-UCSD International Center of Excellence in Advanced Bio-engineering sponsored by the Taiwan Ministry of Science and Technology I-RiCE Program under Grant Number NSC103-2911-I-009-101. The authors also acknowledge financial support from the Ministry of Science and Technology, Taiwan (MOST103-2314-B-010-053-MY3, MOST103-2120-M-010-001, MOST103-2321-B-010-023, NSC101-2314-B-038-022-MY3, NSC101-2120-M-010-002, and NSC100-2911-I-010-503), the Ministry of Economic Affairs, Taiwan (103-EC-17-A-17-S1-203), Wan Fang Hospital, Taipei Medical University (102swf02, 103swf04), and a research grant support from Taipei Medical University and Steminent Biotherapeutics (A-101-023). This study was also supported by a grant from Ministry of Education, Aiming for the Top University Plan.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.