
Abstract
Background:
Decellularized tissue-engineered heart valves (TEHVs) are under investigation as alternative for current heart valve prostheses with the potential to rapidly repopulate with cells within the body. Ideally, these valves are stented for transapical or minimally invasive delivery. It is unclear if and how the matrix of these valves remodels under in vivo hemodynamic loading conditions and in the presence of a stent. Here, we study the evolution of collagen orientation and tissue maturation in the wall of stented decellularized TEHVs with time after implantation.
Methods and Results:
In a previous study, stented TEHVs based on rapidly degrading scaffolds were cultured in bioreactors, decellularized, and transapically implanted as pulmonary valve replacement in sheep. In the present study, collagen (re)orientation in the initially isotropic valvular wall was assessed using a fluorescent collagen probe combined with confocal imaging and image analysis of explanted tissue at 8, 16, and 24 weeks following implantation. Collagen tortuosity or waviness in the explants, as a measure of matrix maturity, was quantified using a Gabor wavelet method and compared with tortuosity in native sheep vascular wall tissue. Results indicate that on the luminal side of the valvular wall, fibers became aligned in circumferential direction, while tortuosity increased with implantation time, showing striking similarities with the native collagen structure after 24 weeks. On the outside of the wall, where the engineered tissue touches the stent, collagen fibers in the vicinity of the struts aligned along the struts, whereas collagen fibers in between struts were randomly oriented. Immunohistochemistry was performed to evaluate the presence of elastin and collagen type I and III. After 8 weeks, collagen types I and III were mostly present at the luminal side of the wall, whereas at 16 and 24 weeks, a homogenous distribution of collagen I and III was observed throughout the wall. Elastin was mostly expressed at the luminal side after 24 weeks. Biochemical assays showed that the amount of DNA (as a measure of cell number) increased significantly after 8 and 24 weeks, glycosaminoglycans increased significantly after 8, 16, and 24 weeks, and hydroxyproline, as a measure of collagen amount, increased significantly after 24 weeks compared to the controls.
Conclusions:
The collagen matrix in the wall of decellularized TEHVs shows clear structural remodeling and maturation with time. While collagen orientation rapidly remodels toward a native anisotropic architecture on the luminal side of the engineered valvular wall, it is dominated and guided by stent geometry on the outer side of the wall. Collagen tortuosity was increased with implantation time and was accompanied by an increase in elastin, especially on the luminal side of the vessel.
Introduction
C
Collagen is the main load-bearing component of cardiovascular tissues and provides them with the ability to withstand continuous hemodynamic loading. Rather than the amount of collagen, the mutual organization of collagen fibers into (anisotropic) load-bearing networks determines the mechanical functionality of the tissue. 3 Thus, methods are being explored to engineer tissues that mimic the structural collagen organization of their native counterpart by using principles like strain-induced remodeling, cell-traction, or contact guidance.4,5 To this end, in vitro loading protocols mimicking the in vivo hemodynamic loading conditions or structural blueprints to guide collagen organization have been used to achieve the desired structure and function of the engineered tissue. Another option is to implant a homogenous isotropic tissue with sufficient strength that remodels in vivo toward the desired organization.
With our current interest in decellularized engineered tissues, it is relevant to know if and how these matrices remodel in vivo. For decellularized stented TEHVs, structural remodeling in the valvular wall may be influenced by hemodynamic loading conditions and the presence of the stent. In addition, this specific case provides an interesting situation to study different aspects of collagen remodeling in vivo. Contact guidance (by the stent struts) will induce collagen fiber orientation near the struts,5,6 whereas physiological mechanical stresses will guide collagen remodeling in the regions where the effect of contact guidance is negligible.
The in vivo structural remodeling of the stented decellularized valvular wall was investigated by monitoring the evolution of collagen orientation and tortuosity or waviness up to 24 weeks following implantation and in comparison to nonimplanted controls. To allow for remodeling, cell infiltration into the implanted matrix must suffice to produce autologous collagen and elastin, which are the major fibrillar components of cardiovascular tissues, and remodel the matrix during development.
Biochemical assays (DNA, hydroxyproline [HYP], glycosaminoglycans [GAG]), collagen architecture analysis, and immunohistochemistry were performed to assess tissue development and maturation. Collagen orientation at different locations in the valvular wall was quantified using confocal laser scanning microscopy (CLSM) in combination with a fluorescent probe (CNA35-OG488),7,8 followed by image analysis using a Mathematica algorithm. To quantify collagen fiber tortuosity, the images were analyzed by a Gabor wavelet function9–12 programmed in MATLAB. Collagen fiber tortuosity is of importance because fiber growth and remodeling toward their natural wavy configuration are an indication of tissue maturation. 13 Finally, immunohistochemistry was performed to provide qualitative insight in the presence of elastin and collagen type I and type III as a measure of maturation of the engineered cardiovascular tissue in vivo. 14
Materials and Methods
Tissue culture, implantation, and explanation procedure
Decellularized TEHVs, consisting of a tubular wall and three leaflets, were engineered as previously described. 15 In short, trileaflet heart valve scaffolds (n=14) were fabricated from nonwoven polyglycolic acid meshes (thickness 1.0 mm; specific gravity 70 mg/cm3, Cellon; Bereldange Luxembourg) and coated with 1.75% poly-4-hydroxybutyrate (MW:1×106; TEPHA, Inc.). Scaffolds were integrated into radially self-expandable nitinol stents (length 27 mm, OD 30 mm when fully expanded at 37°C; pfm medical) and seeded with ovine vascular-derived cells using fibrin as cell carrier. 16 The ovine vascular-derived cells were derived from a piece of vein isolated using the plating (outgrowth) method. They were previously referred to as myofibroblasts, but are likely to comprise a mixture of fibroblasts, myofibroblasts, and smooth muscle cells. After seeding, scaffolds were cultured for 4 weeks in diastolic pulse duplicator systems 17 and decellularized. 15 Decellularized TEHVs were sterilized by immersion in 70% EtOH and stored in a fresh M-199 medium at 4°C until implantation. 2 To evaluate in vivo functionality and host repopulation capacity, decellularized TEHVs (n=10) were crimped 1 and transapically implanted as pulmonary valve replacement in adult sheep (age: 2.8±0.1 years; weight: 69±2 kg), as described previously. 2 Decellularized TEHVs were delivered into the pulmonary valve position (Fig. 1A) using an antero-latero-thoracic access and antegrade approach, as previously described.1,2 Valves were explanted after 8 weeks (n=2), 16 weeks (n=4), and 24 weeks (n=4) and compared to nonimplanted in vitro control valves (0 weeks, n=4, referred to as “controls”). The wall of TEHVs located inside the stent (Fig. 1B, C) was analyzed. For imaging, representative samples from different regions of the wall (luminal side, outer side next to the stent struts and outer side far away from the stent struts, but in-between struts), as shown in Figure 2, were used. The tortuosity index of all the 8-, 16-, and 24-week explants was compared with the tortuosity index of native pulmonary arteries outside (n=3) located next to the stent. For immunohistochemistry, one valvular wall at each time point (0, 8, 16, and 24 weeks) was analyzed. For reorientation and biochemical analysis, all controls (n=4) and explants (n=10) were used.

Schematic view of a stented decellularized tissue-engineered heart valve (TEHV) as pulmonary valve replacement
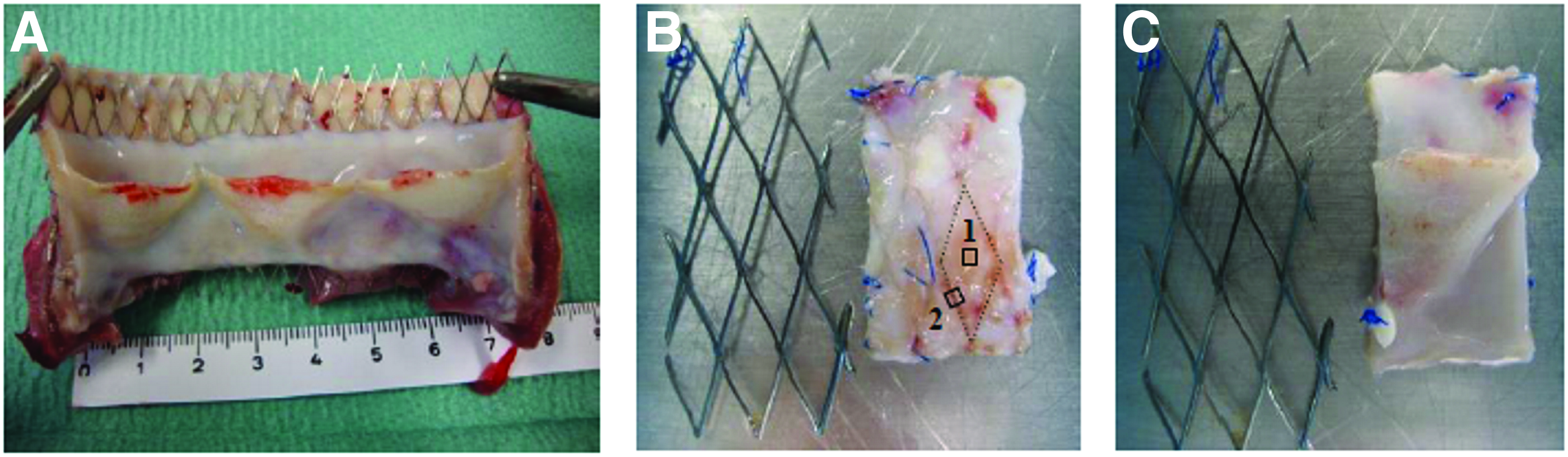
Macroscopic overview of the stented TEHV explant
Collagen orientation visualization
To image collagen fibers in both controls and explants, the CNA35-OG488 fluorescent probe was used. 7 Samples were incubated overnight with a 3 μM CNA probe. Subsequently, samples were washed thrice with PBS and placed in chamber slides for imaging with a Zeiss LSM 510 Meta laser scanning microscope (Carl Zeiss) coupled to an inverted Axiovert 200 microscope (Carl Zeiss). To evaluate the orientation in different regions of the vascular wall, tile scans in the x-y plane were made at different locations along the z direction (=along the radial direction of the vascular wall and up to maximum 200 μm from both the luminal and outer side) by programming the computerized x-y stage. The x-direction and y-direction were defined parallel to the circumferential and the axial direction of the wall, respectively, which were marked upon excision of the samples. Tiles were stitched together in MATLAB (Mathwork) to reconstruct x-y surfaces. 18
Collagen orientation quantification
A previously developed algorithm (Mathematica) was used to quantify fiber orientation in the microscopy images.8,19 For each image, the orientation of all collagen fibers in view was calculated based on the principal curvature directions, which were determined from the eigenvectors of Hessian's matrix. Next, a histogram representing the distribution of fiber orientation was obtained for each image. Circular statistics was used to calculate the mean angle (α) and dispersity (r) of the fiber distribution. 8 If r is zero, fibers are randomly oriented and if r is 1, fibers are completely aligned. For tile scan images, the mean angle and dispersity of the fiber orientation in each tile were depicted as ellipses. It means that if the fibers are completely isotropic, the dispersity is depicted as a circle and if the fibers are completely aligned, it is shown as a line. Furthermore, the angle between the major axis of each ellipse and the horizontal axis shows the mean angle of the fibers in each tile.
Collagen tortuosity quantification
First, the images were preprocessed by image enhancement operation. If the number of adjacent white pixels was smaller than a predetermined threshold value, these pixels were excluded from the image and considered as noise. Thereafter, the remaining white pixels kept their value, and gray pixel values were set to 255. The Gabor wavelet, as a local edge detector, was convolved with the image with different wavelengths and orientations. By changing the wavelength, edges with different widths were detected. The Gabor wavelet used in this study is given by Equation (1). The procedure of obtaining the Gabor function is explained thoroughly by Arivazhagan et al. and Sigari et al.12,20
x′ and y′ were calculated by
The parameters of the Gabor function are the orientation of the wavelet (θ), which is the orientation of the edges that Gabor wavelet detects; the frequency of the wavelet (λ) reflects the sensitivity of the function to a specific edge; phase of cosine signal (φ) is 0 or π/2; Gaussian radius (σ) indicates the magnitude of the region to which the function is applied; and finally, the aspect ratio of the Gaussian function (γ) was set to 1.
Different Gabor wavelets with varying orientation angles (0, π/4, π/2, π) and wavelengths (2, 3, 4, 5) were applied to the images and the corresponding histograms were obtained. A tortuosity index was calculated by deducting the maximum number in the Gabor histogram from 1. Therefore, the final index was between 0 and 1. If the tortuosity index equals 1, tortuosity is very high and if the index equals 0, there is no tortuosity. The index was calculated for collagen fibers in explants with time and in native arterial tissue located behind the stent. To validate the algorithm, various predefined images with known tortuosity were analyzed. We could not compare the waviness of the explants with the controls because the fibers in the controls were randomly distributed.
Immunohistochemistry
The presence and location of the fibrillar matrix proteins collagen type I, collagen type III, and elastin were analyzed qualitatively by immunohistochemistry. Representative pieces of the controls and explants were fixed in 3.7% formaldehyde and subsequently embedded in paraffin. Transverse valvular wall sections of 10 μm were cut, deparaffinized, and prepared for histochemical analysis. First, antigen retrieval was performed with 10 mM sodium citrate and samples were washed with PBS afterward. To inhibit endogenous peroxidases, samples were incubated in a solution of 0.3% hydrogen peroxide in 0.1% sodium azide. To block nonspecific interaction, 10% horse serum in PBS was applied. Then, samples were incubated with rabbit polyclonal anti-collagen I (ab34710; Abcam), anti-collagen III (ab7778; Abcam) and anti-elastin (ab21610; Abcam) primary antibodies. Thereafter, samples were washed with PBS and PBS-Tween and incubated with the goat anti-rabbit IgG biotinylated secondary antibody (Vector Labs, Inc.) accompanied by incubation with the VECTASTAIN ABC Reagent (Vector Labs, Inc.). 21 Slides were washed with PBS and tris/maleate and incubated with 3,3′-diaminobenzidine tetrahydrochloride (D5637; Sigma-Aldrich). Finally, all slides were dehydrated and mounted with Entellan (Merck) onto microscope slides. The stained sections were visualized using a Zeiss light microscope (Axio observer; Zeiss).
Biochemical tissue analysis
The total amount of DNA, GAG, and collagen (HYP) in the wall of controls and 8-, 16-, and 24-week explants were analyzed using biochemical assays. For each valvular wall, two to three samples from the wall were analyzed and represented as one averaged value per valve. To assess the DNA, GAG, and collagen content, sample parts were lyophilized and digested in papain solution (100 mM phosphate buffer, 5 mM L-cysteine, 5 mM ethylenediaminetetraacetic acid, and 125–140 mg papain per mL) at 60°C for 16 h. As the amount of DNA in the decellularized control tissues was assumed to be minimal, it was determined with a very sensitive commercially available Quant-iTTM PicoGreen® dsDNA kit (Molecular Probes, Invitrogen). For the explanted samples, DNA content was determined by using the Hoechst dye method. 22 In short, samples were diluted in the TE buffer (10 mM TRIS, 1 mM EDTA, pH 7.4) and 100 μL was put into a black 96-well plate (Corning) in duplo. A similar amount containing the Hoechst dye (10 mM TRIS, 1 mM EDTA, 2M NaCl, and 2.5 μg Hoechst dye per mL) was added to each well. After 10 min of incubation at room temperature and being protected from light, the fluorescence was measured (excitation: 355 nm, emission: 460 nm). The amount of DNA was computed using a standard curve prepared from calf thymus DNA (Sigma). The amount of GAG was assessed according to a modified version of the protocol described by Farndale et al. 23 and a standard curve prepared from chondroitin sulfate from shark cartilage (Sigma). In brief, 150 μL dimethyl methylene blue (46 μM dimethyl methylene blue, 40.5 mM glycin, 40.5 mM NaCl, pH 3.0) was added to 40 μL of diluted sample and absorbance was measured at 540 nm. The amount of HYP as a measure for collagen content was determined according to the protocol described by Huszar et al. 24 and a standard curve prepared from trans-4-hydroxyproline. The values for DNA, GAG, and collagen are expressed in μg per mg dry weight.
Statistics
Fiber tortuosity and biochemical content data are presented as mean±standard error of mean. One-way ANOVA, followed by Tukey's multiple comparison post hoc test, was used to determine the difference between different time points. Statistical analyses were performed using GraphPad Prism software (GraphPad Software, Inc.). A p-value <0.05 was considered significant.
Results
Collagen fiber orientation
All samples demonstrated similar regional changes in fiber orientation at each time point. Therefore, a representative image of a sample at each time point and its corresponding histogram are shown in Figure 3. Collagen fibers on the luminal side and the outside of the valvular wall were randomly distributed in the controls (Fig. 3A, B). On the outside, small regions in the vicinity of the struts showed alignment of collagen fibers in the direction of the struts (Fig. 3C). After implantation, collagen fibers mainly and quickly aligned in the circumferential direction at the luminal side of the valvular wall (Fig. 3D, G, J). Already after 8 weeks, a clear anisotropic organization was observed that was maintained with time. On the outer side of the wall, the collagen fiber orientation was random in the regions in between and far away from the struts (Fig. 3E, H, K), while the fibers aligned toward the direction of the struts in areas where the tissue was in contact with the struts (Fig. 3F, I, and L). Interestingly, the area of alignment along the struts, where collagen alignment was dominated and guided by the strut direction, became larger with implantation time (Fig. 4).

Representative microscopy images (900×900 μm) of collagen orientation in the valvular wall of TEHV controls

Microscopic tile images (1.8×2.7 mm) of the outer surface of a TEHV control
Collagen tortuosity
To illustrate and validate our algorithm, artificial fibers and their corresponding tortuosity indices, as obtained by the developed algorithm, are shown in Figure 5. By increasing the fibers tortuosity from left to right (Fig. 5A, B, C, respectively), the tortuosity index was increased.

Artificial fibers with different tortuosity and their quantified tortuosity indexes. From left to right
The collagen fiber tortuosity in the wall of the explants was quantified from the microscopy images of 8-, 16-, and 24-week explants and compared to the tortuosity in native pulmonary artery samples collected from the tissue behind the stent (Fig. 5D). The degree of fiber tortuosity in the 8-week explants was significantly smaller than in the 24-week explants and native artery. No differences between the 16-, 24-week explants, and the native artery were observed, indicating a mature native-like waviness of the collagen at these time points.
Immunohistochemistry
Elastin and collagen type I and type III distribution in the wall of control TEHVs and in the 8-, 16-, and 24-week explants was examined by immunohistochemistry. No elastin was detected in the control, illustrating the absence of elastin in the decellularized engineered tissue (Fig. 6A). However, elastin was presented at 8 and 16 weeks after implantation (Fig. 6B, C). At 24 weeks, a dense layer of elastin was observed at the luminal side of the explanted valvular wall (Fig. 6D). Collagen type I in the controls and 8-week explants was predominantly present at the luminal side of the wall (Fig. 6E, F), while it was distributed homogeneously throughout the vascular wall of the 16- and 24-week explants (Fig. 6G, H). Collagen type III was also more dominant at the luminal side of the wall in controls and 8-week explants (Fig. 6I, J). In vivo remodeling of TEHVs coincides with thickening of the tissue, which then become thinner after 24 weeks.
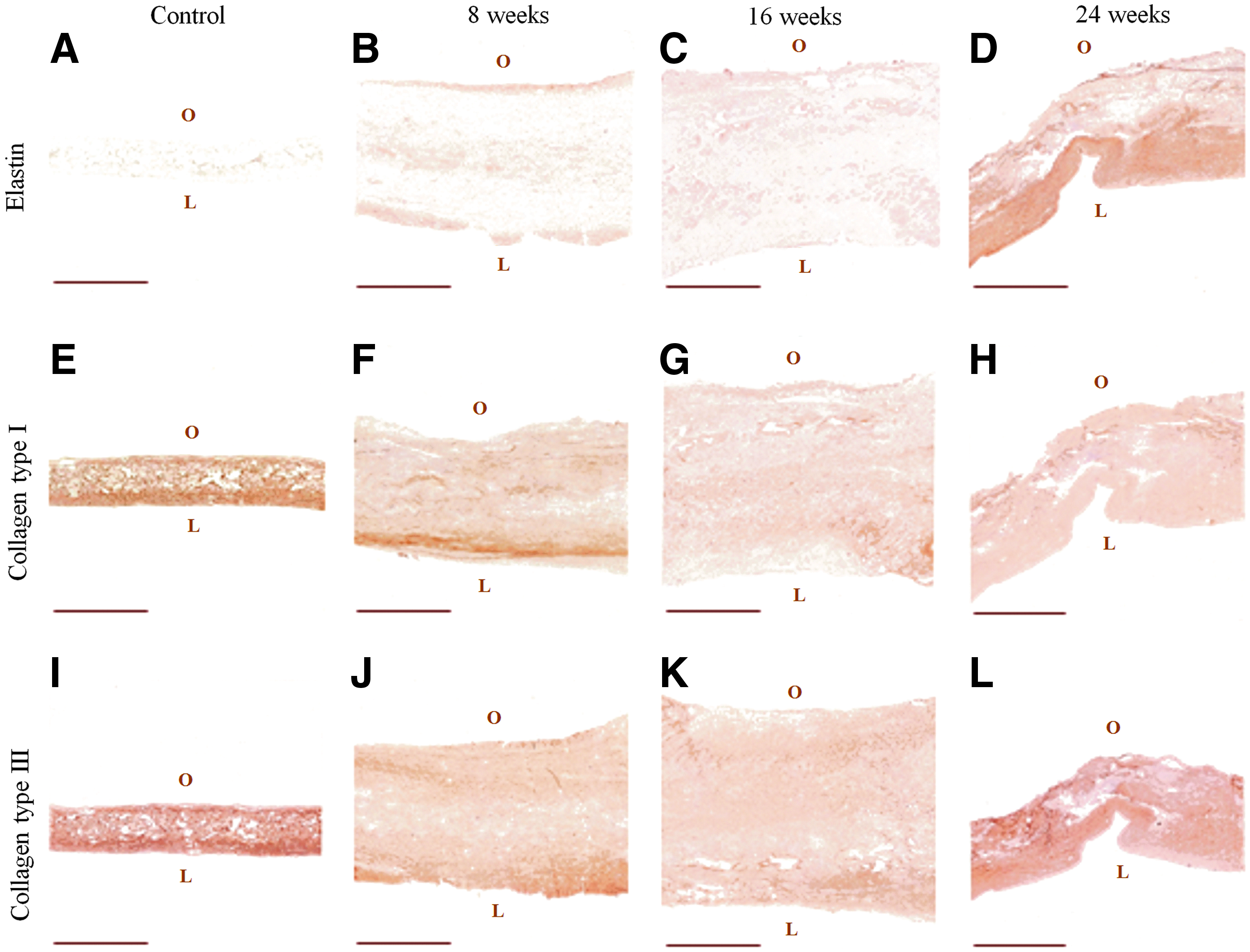
Representative images of immunohistochemical stainings for elastin
Biochemical content
The amount of DNA per dry weight was higher in 8- and 24-week explants compared to the controls, indicating recellularization in vivo (Fig. 7A). Furthermore, the amount of HYP per dry weight was significantly higher in 24-week explants, but not in 8- and 16-week explants, compared to the controls (Fig. 7B), marking a slow increase in the amount of collagen in vivo. The amount of GAG per dry weight, on the other hand, was significantly elevated at 8 weeks and remained stable thereafter (Fig. 7C).

DNA
Discussion
Collagen and elastin are the major components of cardiovascular tissues and give the tissue the ability to withstand physiological loading and to recoil to its original configuration after stretching. We are not sure to what extent we need to mimic these features before implantation. What is important is that these features of a well-organized matrix with elastic fibers will develop in vivo to ensure life-long functionality. In the present study, tissue remodeling and collagen organization evolution of the stented wall of TEHVs were investigated with implantation time. In vivo, the valvular wall is exposed to mechanical loading and potential guiding by the stent structure. Both cues play an important role in the valvular wall extracellular matrix remodeling.
Collagen fibers in different regions of the valvular wall were stained with a collagen-specific probe and visualized with CLSM. Both collagen orientation and collagen tortuosity were analyzed and quantified by in-house developed algorithms. At the luminal side of the valvular wall, collagen fiber orientation changed from random in the controls to circumferentially aligned in all explants. Collagen fiber orientation was further dramatically influenced by the stent struts on the outer side of the valvular wall, where the tissue was in direct contact with the stent. Fibers were aligned in the direction of the stent struts next to the struts and aligned randomly in-between the struts. In another study, we showed the same pattern at the luminal side of the stented pulmonary artery, where the tissue was touched by the stent (unpublished data). However, it was shown that collagen fibers remained circumferentially aligned in the outer region of the pulmonary artery, where the tissue was not in direct contact with the stent. It is known that cells align in response to contact guidance (e.g., by the stent struts in this case) and along the direction of contact guidance. Neighboring cells follow the orientation of the first cells by cell–cell contact, 25 as was observed in the vicinity of the struts in our study. It is unlikely that the observed contact guidance—or collagen orientation in general—was induced by scaffold remnants. Earlier studies on tissue-engineered vascular walls, obtained using the same scaffold,2,15 indicated that such remnants are only present up to week 8 in vivo as small chaotic fragments that do not provide any structural or mechanical support. Moreover, we recently demonstrated that also in vitro, and already after 3 weeks of culture, scaffold remnants do not influence collagen orientation. 26 Together, these data indicate that the hemodynamic loading and presence of the stent are the main determining factors of collagen orientation in the present study.
Most interestingly, collagen tortuosity or waviness was significantly higher in 24-week explants compared to the decellularized control tissues, in regions where collagen orientation was dominated by strain. It was also observed that tortuosity of stented parts of the valvular wall was lower than that of nonstented parts, distal to the stent, which might be due to the (static) radial expansion of the wall by the stent. We suggest that increasing the elastin formation with time is the main reason of increasing collagen waviness. Elastin—coinciding with collagen fibers—acts like a spring that helps the collagen fibers to recoil and retain a wavy configuration. 27 Similar to collagen tortuosity, the amount of elastin was higher in 24-week explants compared to controls. A dense layer of elastin was visualized on the luminal side of the valvular wall, comparable to the elastin distribution in a native arterial wall. It was suggested in a previous study that the tissue at the luminal side of an engineered valvular wall is more developed or matured, as there are no scaffold remnants near the surface and this part of the engineered tissue is best oxygenated. 15 This may explain the observed high content of elastin on the luminal side in the present study.
It is well known that the elastic matrix, a key structural component of the arterial wall that provides the tissue with elasticity, is not laid down by adult cells in vitro. The elastic matrix is therefore addressed as an absent component in in vitro tissue-engineered constructs.14,28 Production of tropoelastin was demonstrated by some studies in vitro, although this did not result in the formation of an elastic matrix. The formation of the elastic matrix clearly requires more than tropoelastin only. There were no signs of elastic matrix formation in the TEHV after in vitro culture. 1
Collagen type I and III were distributed evenly through the valvular wall. However, it has been shown that the concentration of collagen type I and III is increased from intima to adventitia in the native artery.29,30 DNA values revealed that cell repopulation, needed to initiate matrix remodeling of the implanted tissue, occurred after implantation. This impacts not only the collagen matrix (re) organization but also its amount and composition. In addition, it is possible that recruited cells at some regions degrade or synthesize more collagen compared to other regions in response to local stresses and strains.
Immunohistochemical observations revealed that the valvular wall was thicker than controls after 8 and 16 weeks of implantation, but became thinner afterward. It was assumed that this temporary in vivo tissue thickening of the valvular wall represents the functional remodeling and healing procedure of the implants, leading to a more native-like tissue.31,32 Schmidt et al. 1 suggested that this thickening might also be due to the transapical valve delivery method, but this hypothesis was later rejected by Dijkman et al. 15 In another study, it was hypothesized that remnants of scaffold material might be the reason behind tissue thickening as a local inflammation response was observed around scaffold remnants. 33 Although valve thickening can negatively affect valvular hemodynamic forces, which can cause further vascular complications, so far this phenomenon has been observed to be of temporary nature.
Biochemical assays showed that the amount of DNA was increased in the explants, which confirmed cell infiltration into the decellularized implants, required for remodeling. The amount of collagen (HYP) and GAG was also increased, pointing at matrix remodeling. The amount of HYP in 24-week explants was higher compared to the native pulmonary artery located behind the stent (unpublished data).
In conclusion, collagen organization evolution in stented tissue-engineered valvular wall under in vivo circumstances was evaluated. The observed rapid in vivo architectural reorganization, which supports mechanical functionality, suggests that it is sufficient to implant a homogeneous tissue—and not necessarily a structured one—since it will remodel in vivo. CLSM and image analyses revealed that collagen orientation and tortuosity changed up to 24 weeks after implantation. Tissue remodeling was accompanied by an increase in elastin, especially at the luminal side of the valvular wall. Moreover, biochemical content changes after implantation confirmed the remodeling process. This study provides new insights into the in vivo remodeling of tissue-engineered valvular wall constructs toward a native-like tissue architecture and demonstrates their potential to repair and remodel.
Footnotes
Acknowledgments
The authors gratefully acknowledge the support of the TeRM Smart Mix Program of the Netherlands Ministry of Economic Affairs and the Netherlands Ministry of Education, Culture and Science. The in vivo study leading to these results was performed with funding from the European Union's Seventh Framework Programme ([FP7/2007-2013]) under grant agreement no. 242008. The authors would like to thank Anna Vilanova for her great help in image analysis.
Disclosure Statement
No competing financial interests exist.