
Abstract
Chronic low-frequency stimulation (CLFS) has long been used to induce a fast-to-slow phenotype shift in skeletal muscle. In this study, we explore the role of frequency (10 and 20 Hz), active time (15–60%), and streptomycin in inducing a fast-to-slow shift in engineered muscle. We found that C2C12 engineered muscle could respond to CLFS with an adult-like active time of 60% and found that a constant 10 Hz train of 0.6 s, followed by 0.4 s rest, induced a partial fast-to-slow phenotype shift. Following 2 weeks of CLFS, time-to-peak tension (TPT) (control [CTL]=40.9±0.2 ms; 10 Hz=58.5±3.5 ms; 20 Hz=48.2±2.7 ms) and half-relaxation time (1/2RT) (CTL=50.4±0.6 ms; 10 Hz=76.1±3.3 ms; 20 Hz=66.6±2.3 ms) slowed significantly in frequency, but not in an active time-dependent manner. Streptomycin significantly blunted the slowing of TPT and 1/2RT induced by CLFS by minimizing the fast-to-slow shift in SERCA isoform. Streptomycin (Nonstim=−42.8%±2.5%; Stim=−38.1%±3.6%) significantly prevented the improvement in fatigue resistance seen in CTL constructs (Nonstim=−58.4%±3.6%; Stim=−27.8%±1.7%). Streptomycin reduced the increase seen in GLUT4 protein following CLFS (CTL=89.4%±6.7%; STREP=41.0%±4.3%) and prevented increases in the mitochondrial proteins succinate dehydrogenase (SDH) and ATP synthase. These data demonstrate that streptomycin significantly blunts the fast-to-slow shift induced by CLFS. In the absence of streptomycin, CLFS induced slowing of contractile dynamics and improved fatigue resistance and suggests that this model can be used to study the mechanisms underlying CLFS-induced adaptations in muscle phenotype.
Introduction
S
Electromyography (EMG) recordings have shown that slow muscles have a natural tonic firing frequency of 10–20 Hz and can fire at least up to 300,000 impulses per day.12,13 Chronic low-frequency stimulation (CLFS) protocols that mimic the natural tonic firing pattern of slow phenotype muscles have been shown to induce fast-to-slow fiber-type transitions both in vivo 7 and in vitro. 11 Despite CLFS patterns attempting to mimic the natural neural input to slow fiber-type muscles (e.g., Soleus), no standardized CLFS protocol has been developed. This is due, in part, to the differential response to CLFS in different muscle groups10,14 and animal species,7,10,15 showing that no single CLFS pattern exists to transform all muscles into a slow muscle fiber type.
CLFS is typically performed by attaching small programmable stimulator units either on the backs of animals 16 or subcutaneously in animals17,18 and humans. 19 External stimulators can be damaged or dislodged by the animals and implantable devices can have limited reuse capability, leading to an increase in experimental cost. Furthermore, studying the mechanisms underlying CLFS-induced shifts through inhibitor studies is vastly more expensive and more difficult in vivo compared with in vitro. CLFS studies have been performed in vitro and have been shown to induce changes in myofibrillar and metabolic protein expression. 11 However, performing CLFS in two-dimensional (2D) cell culture is hindered by the fact that prolonged continuous stimulation results in rapid myotube detachment, and cell cultures typically detach within 7 days without electrical stimulation. 20 Animal studies have shown that changes in myosin isoform can take up to 60 days to appear 21 and can require constant electrical stimulation for 24 h per day, 22 which are technically impossible to perform with 2D cultures due to myotube detachment. Tissue-engineered skeletal muscle offers an alternative approach by permitting long-term culture (up to 5 weeks) 23 and allowing stimulation times far greater than what 2D cultures permit.14,20,24 In support of this, muscles engineered from primary rat muscle have been shown to adapt to different electrical patterns following 2 weeks of electrical stimulation in vitro. 14
Although engineered muscle is a highly promising model to study muscle physiology, 25 several factors, including electrical stimulation parameters,20,24 biomaterial choice,26–28 and nutrient supply, 29 need to be optimized to better recreate the in vivo milieu. One such factor is the choice of antibiotics used to minimize bacterial and fungal contamination. Streptomycin is a widely used antibiotic in cell culture, but it decreases protein synthesis and maturation in muscle cultures. 30 Streptomycin also acts as a nonspecific calcium blocker, including stretch-activated channels (SACs), which can allow conductance of Na+, K+, Mg2+, and Ca2+ ions 31 in response to both mechanical and electrical signals. Streptomycin has been shown to block the increase in force induced by the stretch in engineered cardiac tissue in vitro 32 and adult skeletal muscle in vivo. 33 In this article, we first sought to determine a CLFS protocol for muscle engineered from the C2C12 cell line, and second, we sought to determine whether streptomycin has any effect on the response of engineered skeletal muscle to CLFS.
Materials and Methods
Two-dimensional cell culture
The C2C12 myoblast cell line (ATCC) was grown in growth media consisting of high glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin until 70% confluent. C2C12 cells were used between passages 6 and 10.
Three-dimensional cell culture
Muscles were engineered using fibrin casting as reported previously. 23 Briefly, the muscle constructs were engineered between two 6 mm long silk sutures set 12 mm apart on Sylgard (PDMS)-coated dishes. Five hundred microliters of growth media containing 10 U/mL thrombin, 0.2 μg/mL genipin, and 0.5 μg/mL aprotinin was added to the plate and agitated until it covered the entire surface. Two hundred microliters of 20 mg/mL fibrinogen was added dropwise, and the gels were left to polymerize for 1 h before addition of 100,000 cells. Two days after plating cells, the constructs were switched to differentiation media consisting of high-glucose DMEM supplemented with 10% horse serum and penicillin (100 U/mL) for 2 days. Following the second day in differentiation media, the constructs were moved to high-glucose DMEM with 7% FBS and penicillin (100 U/mL) for the remainder of the experiment as previously found to be optimal. 23 For experiments containing streptomycin, streptomycin was added to the media at a concentration of 100 μg/mL.
Electrical stimulation
Electrical stimulation was performed using a custom-made electrical stimulator previously described in detail. 24 Constructs were differentiated for 7 days and then were initially electrically stimulated for 24 h with an electrical stimulation protocol consisting of a continuous 0.4 s 10 Hz train, followed by a 3.6 s rest. The constructs were then electrically stimulated for 14 days with an electrical stimulation protocol consisting of a continuous 0.6 s 10 Hz train, followed by a 0.4 s rest, unless stated otherwise. Both the nonstimulated and electrically stimulated constructs were fed daily once electrical stimulation was started.
Contractile testing
Functional testing of the C2C12 constructs was performed as described previously. 34 To determine the contractile properties of the engineered tissue, one of the anchors was freed from the Sylgard substrate and attached to a custom-made force transducer through one of the minutien pins. Rheobase (R50) and chronaxie (C50) were then determined as described previously. 34 Rheobase was calculated as the electric field strength (V/mm) eliciting 50% peak twitch force (Pt) with a 4 ms pulse width. Chronaxie was calculated as the pulse width required to elicit 50% peak force at twice the rheobase. Once excitability had been determined, all impulses were delivered with a 4 ms pulse width at 4× R50 as described previously. Fatigue was determined by stimulating for 0.75 s with 0.75 s rest at 50 Hz for 3 min at four times the rheobase with a 4 ms pulse width.
Cross-sectional area was calculated from the measured width of each construct (at its narrowest point) assuming a rectangular cross section and a depth of 500 μm. Specific force was calculated as kilonewtons per square meter as follows: the force generated by the construct (kN) divided by its cross-sectional area (m2).
Western blot
Tissues were washed in ice-cold PBS, and then blot-dried before freezing in liquid nitrogen and storing at −80°C. At the time of processing, samples were powdered in a 1.5-mL microcentrifuge tube on dry ice, suspended in 200 μL ice-cold sucrose lysis buffer (50 mM Tris pH 7.5, 250 mM sucrose, 1 mM EGTA, 1 mM EDTA, 1 mM sodium orthovanadate, 50 mM sodium fluoride, 5 mM Na2(PO4)2, and 0.1% DTT), and shaken at 1400 rpm for 1 h at 4°C in an Eppendorf thermomixer (Hauppauge). The samples were then centrifuged at 4°C for 1 min at 10,000 g to remove insoluble material. The supernatant was transferred to a new tube, and protein concentration was determined using the DC protein assay (Bio-Rad). Equal aliquots of protein in 1× Laemmli sample buffer were boiled for 5 min before separation on a 10% acrylamide gel by SDS-polyacrylamide gel electrophoresis. After electrophoresis, proteins were transferred to a nitrocellulose membrane (Protran, Whatman) at 100 V for 1 h. The membrane was blocked for 1 h in 3% milk in Tris-buffered saline+0.1% Tween (TBST). Membranes were incubated overnight at 4°C with the appropriate primary antibody in TBST at 1:1000. The membrane was then washed thrice in TBST before incubation for 1 h at room temperature with the appropriate peroxidase-coupled secondary antibody in TBST at 1:10,000 (Pierce). Antibody binding was detected using an enhanced chemiluminescence horseradish peroxidase substrate detection kit (Millipore). Imaging and band quantification were carried out using a Chemi Genius Bioimaging Gel Doc System (Syngene). The primary antibodies used in this study were MF20, F59, CaF2-5D2 (Hybridoma Bank), total eEF2, SERCA 2a (Cell Signaling), slow MHC (Novacastra), parvalbumin (GeneTex), total phosphofructokinase (PFK), GLUT4, succinate dehydrogenase (SDH), and ATP synthase (Santa Cruz Biotechnology).
Statistical analysis
Data are presented as mean±SEM. Differences in mean values were compared within groups and significant differences were determined by analysis of variance with the post hoc Tukey–Kramer HSD test using Brightstat.com. 35 The significance level was set at (p<0.05).
Results
Effect of frequency, active time, and impulse number on contractile properties
We first optimized a CLFS protocol by examining the effects of frequency, active time, and impulse number on the shift to a slower contractile phenotype following 2 weeks of electrical stimulation in the presence of streptomycin (Table 1). Overall, we found that time-to-peak tension (TPT) and half-relaxation time (1/2RT) were the contractile properties that displayed the greatest change following CLFS. We found that frequency was the dominant factor over both impulse number and active time in determining the change induced by CLFS. CLFS significantly slowed TPT (control [CTL]=40.9±0.2 ms; 10 Hz=58.5±3.5 ms; 20 Hz=48.2±2.7 ms) and 1/2RT (CTL=50.4±0.6 ms; 10 Hz=76.1±3.3 ms; 20 Hz=66.6±2.3 ms), but 10 Hz induced greater slowing than 20 Hz (Fig. 1). Despite the slowing of TPT and 1/2RT, we found no change in fatigue resistance, one of the classical changes following CLFS seen in vivo, and hypothesized that streptomycin in the media may prevent the metabolic changes required to improve fatigue resistance (Table 1).
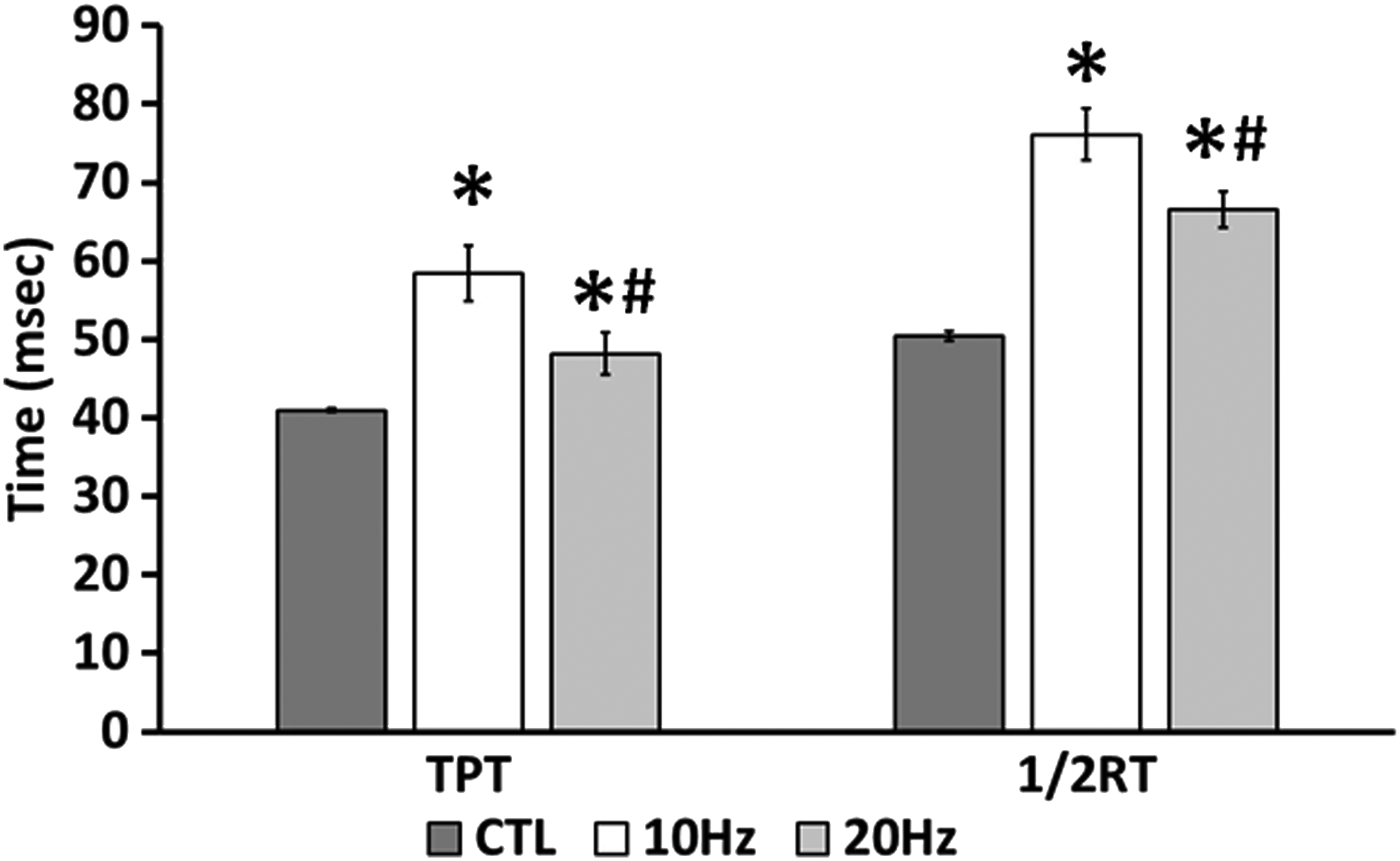
Effect of Frequency on electrical stimulation induced changes in contractile dynamics. Time-to-peak tension (TPT) and half-relaxation time (1/2RT) were measured after 2 weeks of chronic low-frequency electrical stimulation (continuous 0.6 s stimulation, followed by 0.4 s rest). *Significantly different from nonstim and #significantly different than 10 Hz (p<0.05). CTL, control.
Significantly different from CTL.
Following 7 days of differentiation, engineered muscle constructs were stimulated for 2 weeks with 5 (A–E) electrical stimulation patterns and contractile properties were then measured.
1/2RT, half-relaxation time; CTL, control; TPT, time-to-peak tension.
Effect of streptomycin on active force production following 2 weeks of electrical stimulation
To study the effect of streptomycin on the response of engineered muscle to CLFS in vitro, we performed CLFS (continuous 0.6 s 10 Hz train, followed by 0.4 s rest) on C2C12 engineered muscles for 2 weeks in the presence or absence of streptomycin. Streptomycin decreased force production in both nonstimulated (CTL=0.014±0.001 kN/m2; STREP=0.006±0.0005 kN/m2) and stimulated muscles (CTL=0.036±0.002 kN/m2; STREP=0.017±0.002 kN/m2) (Fig. 2A), but did not affect the fold increase in force seen following CLFS (CTL=165.8%±11.2%; STREP=181.8%±17.6%) (Fig. 2B). CLFS led to a significant increase (CTL=165.8%±11.2%; STREP=181.8%±17.6%) in MHC protein content compared with nonstimulated muscles (Fig. 2C).
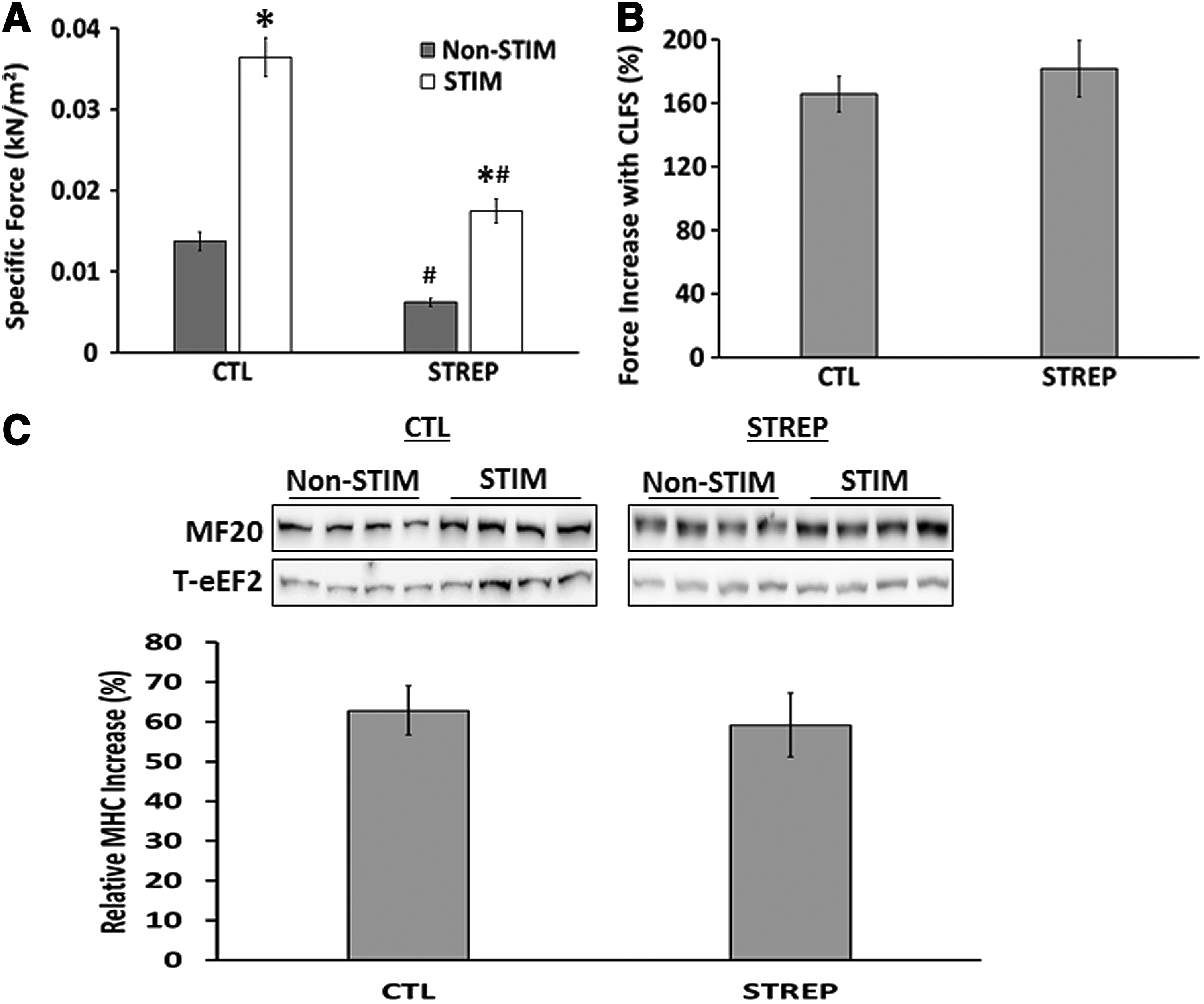
Effect of CLFS and streptomycin on force production and MHC content.
Effect of streptomycin on TPT following 2 weeks of electrical stimulation
We next looked at the change in TPT and 1/2RT, classical markers used to help determine muscle phenotype. In nonstimulated muscle, streptomycin led to a significant slowing in TPT (CTL=40.9±1.4 ms; STREP=50.8±1.2 ms). CLFS led to a slowing of TPT (CTL=83.3±2.6 ms; STREP=68.5±2.7 ms), but this was attenuated in the presence of streptomycin (Fig. 3A).
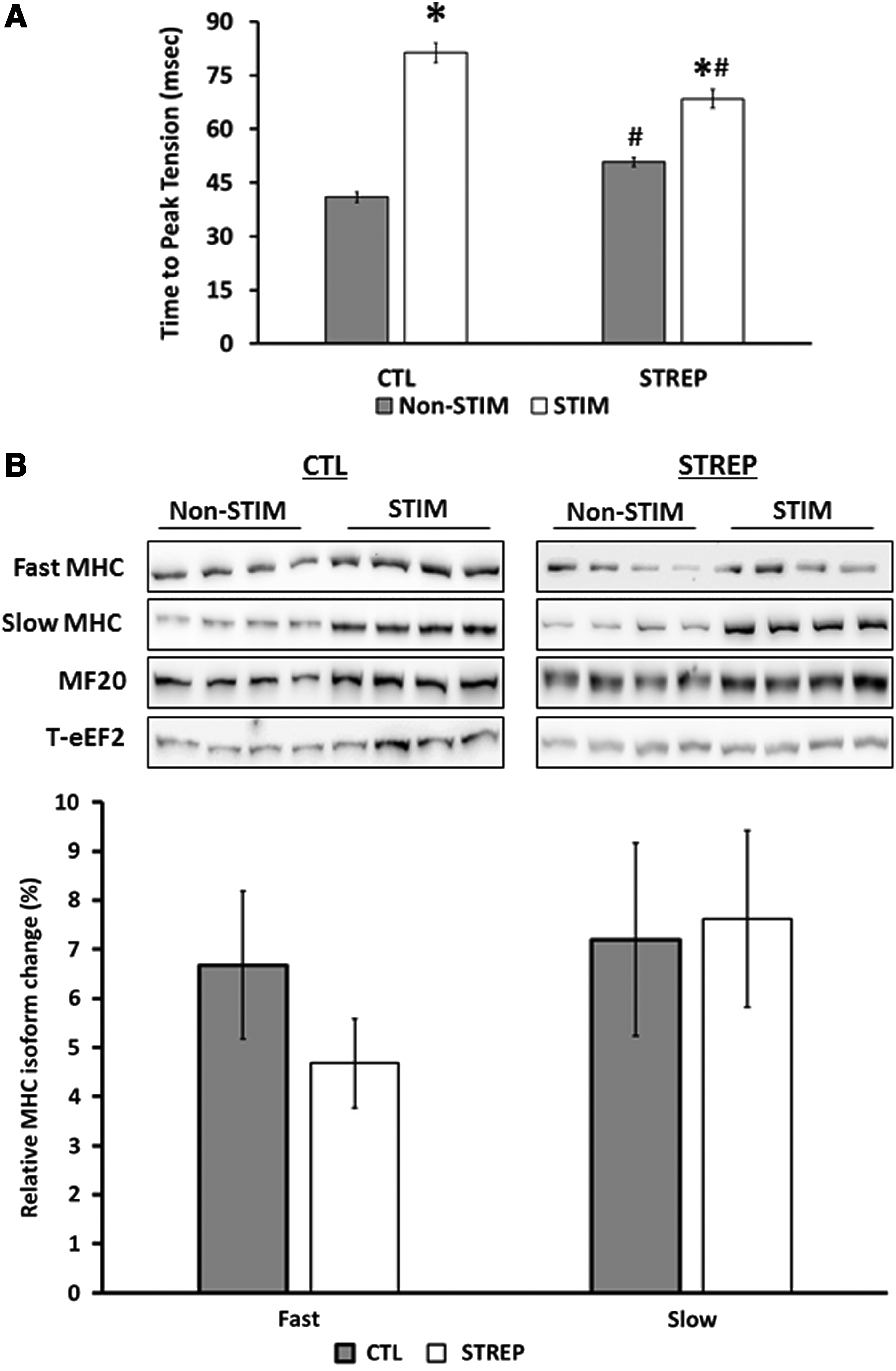
TPT and slow and fast MHC protein content following CLFS.
In adult muscle, myosin isoform is the main determinant of TPT, so we analyzed the relative levels of slow and fast MHC. Relative to total MHC content, we found no change in the relative abundance of the fast MHC isoform following CLFS (Fig. 3B). Relative to total MHC, the relative abundance of slow MHC increased following electrical stimulation (CTL=72.2%±8.0%; STREP=77.6±8.8 ms).
Effect of streptomycin on 1/2RT following 2 weeks of electrical stimulation
Similar to TPT, in nonstimulated muscle, streptomycin led to a significant slowing in 1/2RT (CTL=50.4±1.4 ms; STREP=58.3±1.7 ms). CLFS resulted in slowing of 1/2RT in both the presence and absence of streptomycin (CTL=94.3±4.2 ms; STREP=76.1±3.5 ms), though streptomycin significantly attenuated this slowing (Fig. 4A).
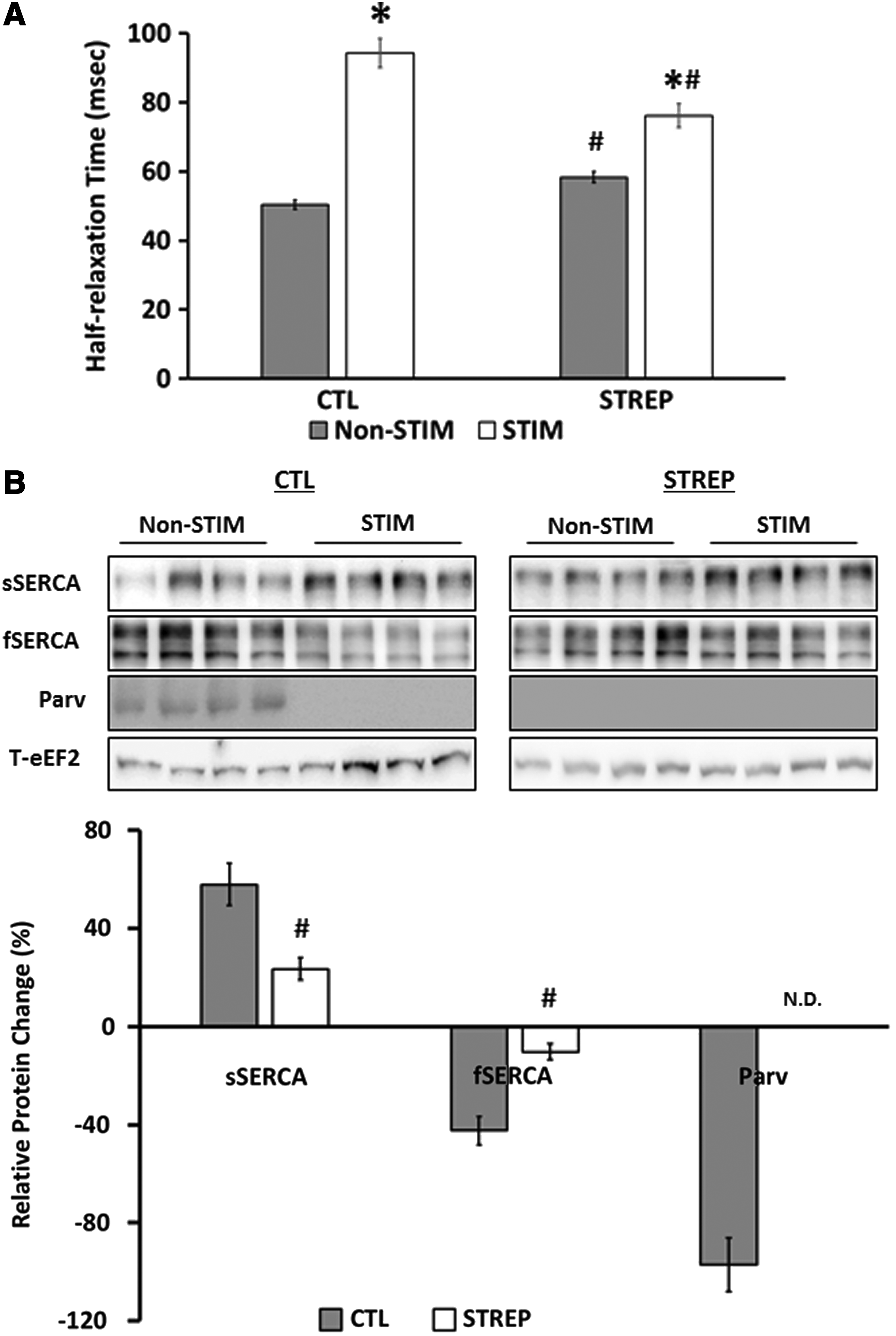
Half-relaxation time and calcium uptake and sequestering proteins levels of engineered muscle following CLFS.
To determine the cause of the shift in 1/2RT, we looked at the levels of the fast-specific calcium-sequestering protein parvalbumin (Parv) and both the slow and fast isoforms of the calcium uptake protein SERCA (Fig. 4B). Following CLFS, the relative increase in the slow SERCA isoform (CTL=58.0%±8.6%; STREP=23.6%±4.6%) was significantly blunted in the presence of streptomycin (p<0.05). Parvalbumin protein could not be detected in streptomycin-treated constructs. In control constructs, Parv was detected in nonstimulated tissues, but not in stimulated constructs.
Effect of streptomycin on fatigue resistance following 2 weeks of electrical stimulation
Following 2 weeks CLFS, fatigue resistance increased significantly in stimulated muscles compared with nonstimulated constructs in the absence of streptomycin (Nonstim=−58.4%±3.6%; Stim=−27.8%±1.7%) (Fig. 5A). CLFS in the presence of streptomycin prevented the improved fatigue resistance (Nonstim=−42.8%±2.5%; Stim=−38.1%±3.6%). However, streptomycin significantly increased fatigue resistance in nonstimulated muscles, though not to the level of stimulated CTL muscles.

Fatigue and metabolic protein levels of engineered muscle following CLFS.
To determine whether the change in fatigue resistance was due to changes in metabolic protein content, we looked at the levels of glycolytic and mitochondrial proteins (Fig. 5B). PFK, the rate-limiting step of glycolysis, 36 was not detected in constructs cultured with streptomycin. In CTL constructs, CLFS resulted in a 53.1%±5.5% decrease in PFK protein. In contrast, GLUT4 protein increased following CLFS (CTL=89.4%±6.7%; STREP=41.0%±4.3%), but this increase was significantly blunted in the presence of streptomycin. The mitochondrial proteins SDH, a protein that participates in both the citric acid cycle and the electron transport chain 37 (CTL=28.5%±3.6%; STREP=−4.6%±2.3%), and ATP synthase, the enzyme that converts ADP and inorganic phosphate to ATP 38 (CTL=22.8%±2.4%; STREP=4.3%±1.8%), were also found to increase in CTL, but not streptomycin fed constructs.
Effect of streptomycin on excitability following 2 weeks of electrical stimulation
We next looked at the markers of excitability, rheobase, and chronaxie (Fig. 6). Rheobase was unaltered following CLFS in control tissues (Nonstim=0.35±0.01 V/mm; Stim=0.37±0.02 V/mm). Compared with CTL tissues, streptomycin resulted in an increase in rheobase (Nonstim=0.48±0.02 V/mm; Stim=0.43±0.02 V/mm), but CLFS did lead to a decrease in rheobase (Fig. 6A).
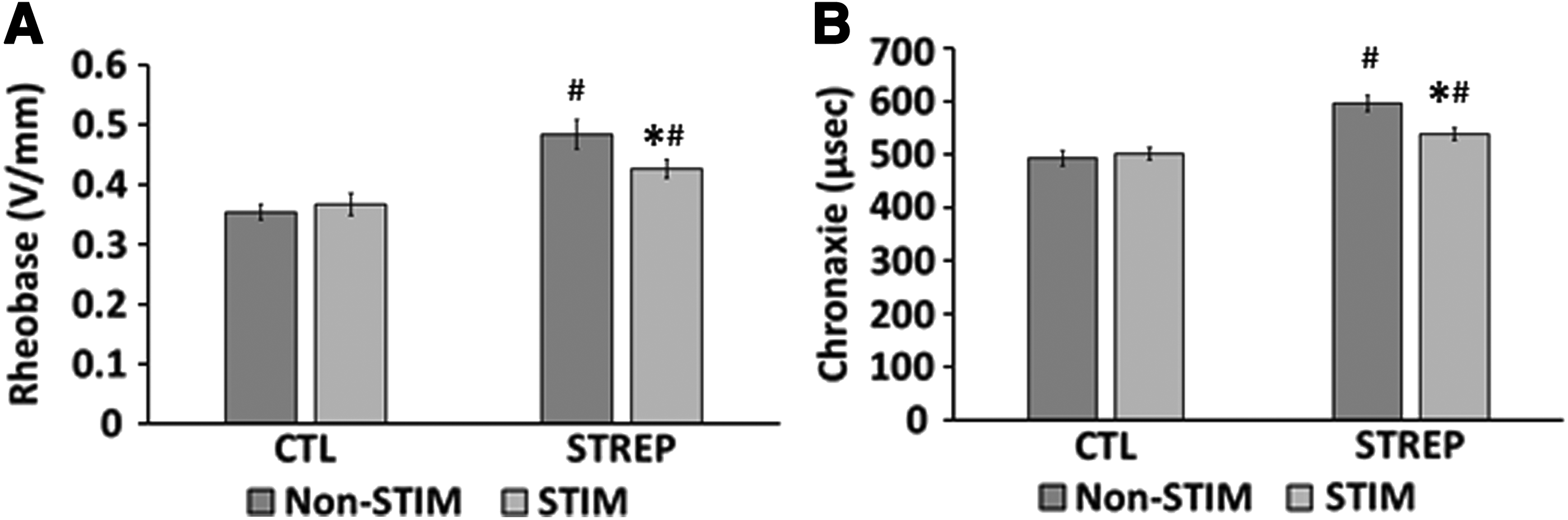
Excitability of engineered muscle constructs following CLFS.
Chronaxie was unaltered following CLFS in control tissues (Nonstim=492.6±14.6 μs; Stim=501.7±11.5 μs). Compared with CTL tissues, streptomycin resulted in an increase in chronaxie (Nonstim=595.8±14.6 μs; Stim=538.6±11.5 μs), but CLFS did lead to a decrease in chronaxie (Fig. 6B).
Discussion
In this article, we studied several CLFS patterns and found that frequency was the main determinant of contractile dynamics following CLFS. Even though streptomycin had no effect on the increase in force production seen with electrical stimulation, it greatly blunted the slowing of TPT and 1/2RT and completely blocked the increase in fatigue resistance seen with CLFS. Concomitant with this blunting of function, streptomycin blocked the increase in mitochondrial proteins (SDH and ATP synthase) and reduced the changes seen in both fast and slow SERCA content and the glycolytic protein GLUT4.
One of the main differences found between CLFS between different species and muscle is the primary determinant of contractile phenotype. While some laboratories maintain that frequency7,15,39 is the main determinant, others suggest that impulse number, 40 active time, 40 or time allowed between contractions 41 determine the phenotypic shift. We found that frequency was the primary determinant of the response to CLFS in vitro (Fig. 1) with impulse number and active time having no effect (Table 1). It should be noted that those experiments were performed in the presence of streptomycin and this may have blocked or blunted some of the response. However, we have found that performing CLFS in the presence or absence of streptomycin with six times fewer impulses induces a similar change in TPT and 1/2RT (data not shown) supporting the fact that frequency, in the ranges studied, is the primary determinant of contractile dynamics in engineered muscle.
The optimal electrical stimulation protocol found in this study was a continuous 0.6 s 10 Hz train, followed by 0.4 s rest. The decision to shift to a more intensive stimulation protocol than used previously 14 was instigated by the fact that single EMG recordings recorded from the adult rat soleus muscle show that slow motor units are active approximately 60% of the time. 13 A previous electrical stimulation protocol of a continuous 0.4 s 20 Hz train, followed by 3.6 s rest, was shown to induce a slow shift only in tissue engineered from satellite cells derived from the fast phenotype tibialis anterior muscle. 14 The slowing of contractile properties were similar in the current study when using a frequency of 20 Hz. Coupled with the increased force generation and fatigue resistance, this demonstrates that the engineered C2C12 constructs can adapt to a more adult-like neural pattern and are a useful model to study CLFS-induced changes.
Specific force production increased approximately 2.5-fold in this study with a concomitant increase in MHC content. The increase in MHC is in keeping with previous observations that electrical stimulation increases MHC content of in vitro cultures. 42 Despite the increase in MHC, the increase in force production occurs entirely in the first 24 h before an increase in MHC is seen. 24 It is not entirely clear why the increase in MHC does not increase force production further as electrical stimulation continues, but it suggests either that impaired force transmission prevents further increases in force or more than isometric contractions are required to improve force production. Force transmission is the link between the contractile machinery and the extracellular matrix. In this study, the muscles were engineered from a pure myoblast population. In vivo, fibroblasts produce the functional connective tissue that is vital to force transmission. 43 We have found that adding fibroblasts to the C2C12 cultures decreases force production under static conditions (Khodabukus and Baar unpublished observation), but these cells may be required to improve force transmission when the constructs are stimulated. Alternatively, isometric contractions might not be sufficient to increase force in response to stimulation. In support of this hypothesis, the maintenance of force in denervated muscles in vivo requires all three contraction types (isometric, concentric, and eccentric). 3 Therefore, it is likely that further increases in force production with engineered muscle may require contraction types other than the isometric contractions employed in this study.
Following 2 weeks of CLFS, we saw no shift in the relative levels of MHC isoform regardless of the presence of streptomycin. Smaller mammals such as mice and rats 21 are less adaptive in fast-to-slow shifts than larger animals such as rabbits,7,15 and it can take up to 60 days of chronic 10 Hz stimulation (i.e., 24 h a day) to see a shift in rat muscle.21,22 Therefore, it is not surprising that we didn't see a shift in MHC isoform with our 2-week stimulation protocol in vitro.
Even in the absence of a shift in MHC isoform, we did find that CLFS induced a slowing of 1/2RT, but the slowing was reduced by streptomycin. The changes in calcium sequestering and uptake proteins were consistent with the functional changes recorded and, importantly, the absence of streptomycin mirrored the shifts seen following CLFS in vivo.16,44,45 These shifts included a switch from fast-to-slow SERCA and the elimination of parvalbumin, a calcium-sequestering protein found only in fast phenotype muscles, 46 and likely underlie the slowing of relaxation.
Interestingly, streptomycin blocked the improvement in fatigue resistance and changes in metabolic expression seen with CLFS (Fig. 5). CLFS led to a decrease in PFK protein in the absence of streptomycin, whereas the other glycolytic protein we studied, GLUT4, increased with electrical stimulation, though to a lesser extent with streptomycin. GLUT4 is known to increase following chronic electrical stimulation, both in engineered muscle 47 and in vivo, 48 and increase glucose transport into the cell. PFK is the first enzyme that commits glucose to anaerobic glycolysis, 36 so the loss of PFK and increase in GLUT4 would result in a shift to higher rates of aerobic glycolysis, that is, a more oxidative metabolism.36,49
We found increases in the mitochondrial proteins SDH and ATP synthase following electrical stimulation, but not in the presence of streptomycin (Fig. 5B). SDH participates in both the tricarboxylic acid (TCA) cycle and the electron transport chain, 37 and thus in conjunction with a decrease in PFK, an enzyme which metabolizes glucose in a nonoxidative manner, 36 supports a shift to the oxidative metabolism of glucose. ATP synthase catalyzes the final step of the electron transport chain, 38 the conversion of ADP and inorganic phosphate to ATP, and has been shown to increase following CLFS in vivo. 50 The increase in SDH and ATP synthase would be expected to increase the maximal oxidative capacity of the engineered tissues, ultimately leading to the improved fatigue resistance recorded (Fig. 5A).
One of the key findings of the current work was that streptomycin blocked or blunted changes in calcium handling, leading to less pronounced changes in contractile dynamics, and metabolic proteins preventing changes in fatigue resistance. Although the exact mechanisms underlying the slow shift induced by CLFS are unclear, the sustained increase in intracellular calcium caused by CLFS 51 is thought to be a key signaling mediator. 52 Streptomycin may partially block calcium flux by blocking SACs 31 and/or other calcium channels not limited to, but, including L-type 53 and Ca2+-activated K+ channels. 54 In support of this, streptomycin reduces calcium influx following electrical stimulation, 55 and thus may prevent or decrease some of the changes induced by CLFS through perturbation of calcium flux. Streptomycin also blocks mTOR activation, a key regulator of protein synthesis, following mechanical stress through blockade of SACs. 33 Therefore, it is possible that the smaller protein increases found were also due, in part, to a decrease in protein translation and elongation that are found with decreased mTORC1-S6K signaling. 56
We found that streptomycin in both stimulated and nonstimulated muscle resulted in a decrease in excitability (Fig. 6) and a decrease in the frequency and amplitude of spontaneous contractions (data not shown). Following denervation, excitability decreases in both animal 15 and human 57 tissue and electrical stimulation can reduce the decrease in excitability, albeit very modestly.15,58 Engineered muscle has much lower excitability compared with native muscle,27,34 and electrical stimulation has been postulated to increase engineered muscle excitability. 59 The fact that CLFS does not increase excitability in the absence of streptomycin suggests that the spontaneous contractility in the nonstimulated tissue may be sufficient to maintain excitability. In support of this, in the presence of streptomycin, active time was the key variable that promoted increases in excitability (Table 1). Active times of 30% and 60% induced incremental improvements in excitability, but an active time of 15% was insufficient to induce a change in excitability. In the presence of streptomycin, and hence decreased spontaneous contractility, CLFS may be able to modestly improve CLFS through higher contractile activity.
Using the C2C12 cell line we have been able to induce a functional contractile shift (TPT, 1/2RT, and fatigue) to a slow phenotype using electrical stimulation, which is blunted in the presence of streptomycin. Furthermore, this shift has resulted in changes in calcium handling isoforms and glycolytic and mitochondrial enzymes, suggesting that this may be another model to study the control of skeletal muscle isoform plasticity.
Footnotes
Acknowledgments
This work was supported by the Hellman Family Foundation and the E. Baar Research Trust. A.K. was supported by a fellowship from the CIRM Stem Cell Training Program.
Disclosure Statement
No competing financial interests exist.