
Abstract
Mechanotransduction plays a role in adipose tissues by transducing the environmental mechanical signals. It is recognized that dynamic or cyclic mechanical strains suppress adipogenesis, but static strains activate the adipogenic signaling pathways. This phenomenon needs to be investigated further, given its potential use in tissue engineering of fat. We used in vitro cultures as model systems for studying differentiation and function of adipocytes. Additionally, using the finite element method, we developed here sets of multiscale models (MSM), which represent single or multiple adipocytes embedded in scaffolds, stimulated mechanically in a static regime. Based on in vitro adipocyte culture work, these models were employed to study the hypothesis that the loading state of the plasma membrane (PM) in adipocytes is influenced by neighboring cells, which could reflect positive feedback loops of en mass adipose cell differentiation. We demonstrate that under static loading, tensile strains at the PM increase with the stage of cell maturation. Furthermore, when the cell density was sufficient (above 19 cells per 100 μm3), progressive differentiation in some of the cells caused higher magnitudes of tensile strains in the PMs of other nearby cells. MSM are currently the only feasible means to correlate continuum (macrolevel) construct deformations to subcellular-level PM stretches in distorted cells. These macro-to-micro mechanobiology relationships, revealed through MSM, point to stimulations that promote the formation of lipid droplet accumulations and the increase of adipogenesis. Such models are a cost-effective useful platform for achieving better understanding of these deformation-driven cell processes toward optimized design of tissue-engineered fat constructs.
Introduction
F
Obesity develops when the body fat mass increases, following proliferation and differentiation of preadipocytes in adipose tissues, as a result of a positive energy balance where calorie intake exceeds expenditure. 5 Understanding the influence of exposure to mechanical loading on adipogenesis is critically important in the research of obesity since adipose cells and tissues in multiple anatomical sites are weight bearing and adjacent to skeletal muscle tissues, which deform them while contracting statically or dynamically. 6 Adipose tissues at the buttocks of seated subjects, for example, were found to be subjected to peak mechanical tensile, compressive, and shear strains of ∼30%, ∼45%, and ∼40%, respectively.7–8 A static lying posture resulted in peak strains in the buttocks that were approximately half these magnitudes. Physical exercise can cause transient large strains in fat tissues, for example, in intramuscular fat when the muscle is contracting or in extramuscular fat interfacing a contracting muscle.
Recent evidence indicates that the excessive fat mass in obesity plays important biomechanical roles through mechanotransduction cascades at the cellular and molecular levels. Studies utilizing cell culture models revealed that dynamic loading modes, such as cyclic stretching or vibration, inhibit differentiation of adipocytes,9–18 whereas static stretching accelerates it.19–21 Multiple signaling pathways, for example, the MEK, 20 the Rho-Rho-kinase, 19 or the TGFβ1/Smad, 11 were found to be activated in adipocytes following exposure to mechanical loading. A common limitation of the aforementioned studies, however, is that they were all conducted in two-dimensional (2D) cultures as opposed to three-dimensional (3D) constructs. One way to attempt to bridge this gap is to employ computational modeling, which considers the biomechanical effects of cell–cell interactions, mediated by an extracellular matrix (ECM).
While modeling mechanobiology interactions in 3D, some research groups focused on developing tissue-engineered fat.22–28 Fischbach et al., 22 for example, developed 3D adipose constructs by differentiating 3T3-L1 preadipocytes on PGA polymeric scaffolds. The same cell line was seeded in 3D fibrous polyethylene terephthalate scaffolds and the cells acquired the morphology and several biological features of mature adipocytes. 23 Young et al. 24 used hydrogel scaffolds produced from decellularized human lipoaspirate. Their hydrogels supported the growth and survival of patient-matched adipose-derived stem cells in vitro. Adipose constructs were also produced by suspending adipose-derived stem cells within injectable carboxymethylcellulose-based 25 alginate 26 or chitosan/glycerophosphate/collagen hydrogels. 27 These studies, however, did not analyze the effects of mechanical stimuli on cell growth or maturation.
We hypothesize that when adipocytes are embedded in a construct, which is being mechanically loaded, the levels of stretch deformations in the plasma membrane (PM) of the cells, which affect adipogenesis, would depend on the stage of growth and maturation of the cells. Furthermore, we surmise that the loading state of the PM in adipocytes is influenced by neighboring cells, which could reflect positive feedback loops of en mass adipose cell differentiation. 6 Finite element (FE) multiscale modeling (MSM) is the only feasible approach to correlate the continuum (macrolevel) construct deformations to subcellular-level PM stretches. In the literature, FE MSM studies of different types of engineered tissues, for example, cartilage, 29 bone,30–33 cardiac, 34 or vascular 35 constructs, were described. MSM of engineered adipose tissues was presented in a previous work of our group as a first modeling step in this field, but the modeling was limited by simplifications in the geometrical description of the cells and construct, both represented in a 2D configuration. 36 In this study, we develop 3D MSM of adipose tissue constructs to better understand mechanotransduction cascades, which can ultimately lead to control of adipogenesis, by either suppressing or enhancing lipid droplet (LD) production. We hence address the following research question: How do growth and maturation levels of adipocytes influence exposure to tensile strains on the PM of neighboring adipocytes in mechanically loaded adipose constructs? Such MSM is useful for developing strategies for fat tissue engineering (TE) in obesity research, wound healing and plastic reconstructive surgery, and cosmetics. Our modeling results were further validated against in vitro experimental data that are also reported here.
Methods
Geometry and meshing
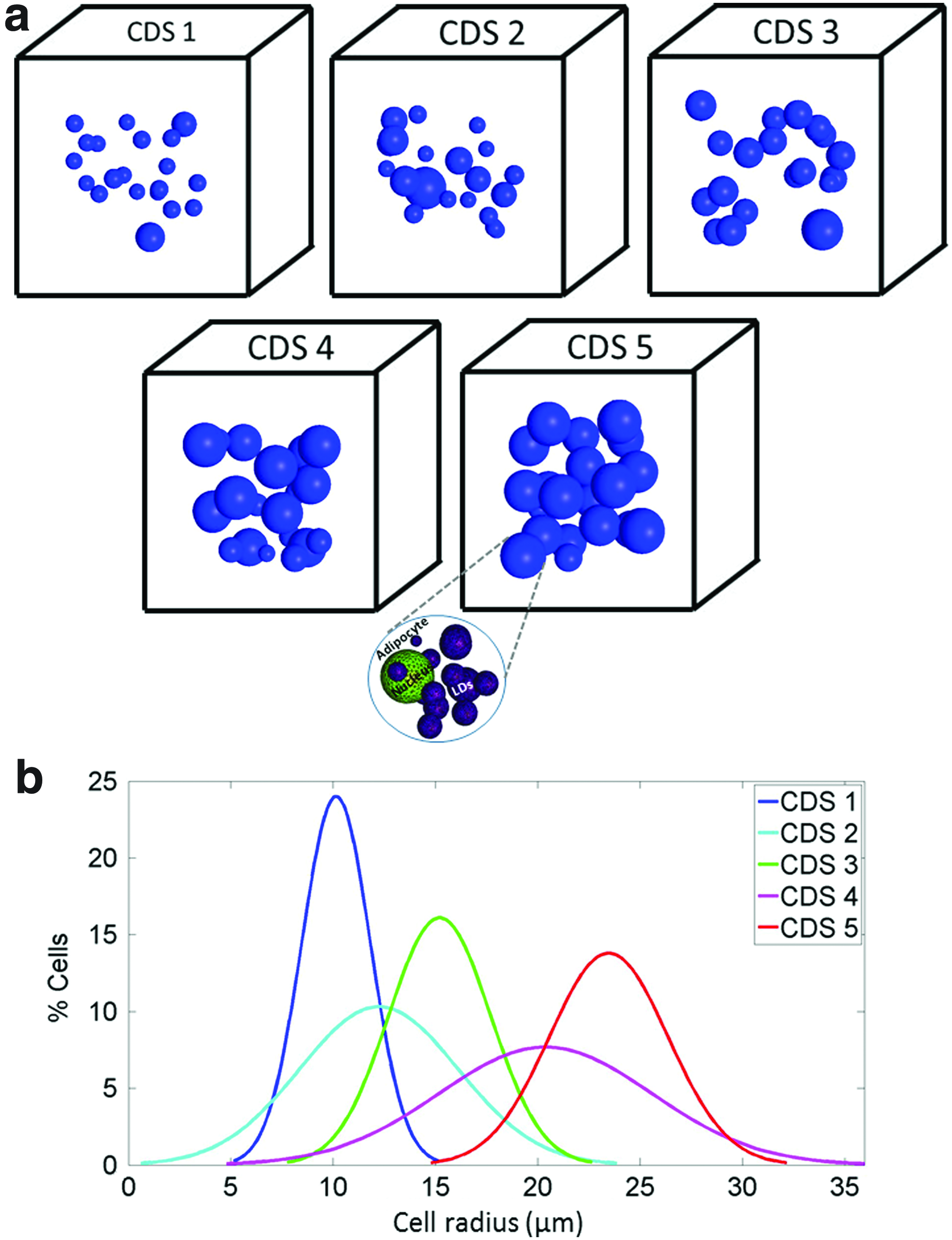
Twenty-eight volumes of interest (VOIs) in 3D adipose constructs were generated using an original custom-made Matlab code (ver. R2013b; The MathWorks, Inc.). The VOIs included either one or multiple adipocytes (Fig. 1). Each simulated adipocyte included the volumes of the cytoplasm, nucleus, and multiple LDs. 36 The shapes of adipocytes, LDs, and nuclei were all assumed to be spherical, which approximates adipocyte behavior in 3D cultures. 37 The nucleus occupied 10% of the total cell volume. 38 The morphological data, including cell sizes and means and standard deviations (SDs) of sizes and numbers of LDs per cell, were obtained previously. 39 During the time course in culture, differentiating adipocytes increase in size to allow accumulation of triglycerides packed as intracytoplasmic LDs. While the sizes of the cells and the LDs grow consistently during differentiation to accommodate the additional triglyceride mass, the numbers of small LDs increase initially, but then decrease as adjacent LDs fuse together to form fewer, but larger LDs, as indicated by our previous studies.20–21,39–40 In this study, we used the data reported in our earlier work 39 to simulate adipocytes at days 5, 15, and 27, which were defined here as the initial, middle, and advanced stages of differentiation, respectively. The maturation level of each simulated cell was raffled randomly, thereby representing our observations of intraculture variability in differentiation levels across different cells in the same culture. The numbers and sizes of LDs in each simulated cell were raffled from within the range of the aforementioned empirically documented cell morphologies 20 assuming normal distributions of these parameters. The LDs and nucleus in each cell were then located randomly in the intracellular space to reflect cell-to-cell variability in intracellular organization. Cell growth and maturation were defined in the modeling as progress in the stage of cell differentiation (per cell in the single-cell simulations, or statistically, for a group of cells) from initial to middle, and then to an advanced stage. Our simulations of adipocytes within a construct were considered related to these three stages of growth/maturation in terms of cell sizes and LD sizes and numbers. Likewise, lipid production or accumulation was not modeled explicitly, but the total volume of LDs in the simulated cells increased with each of the aforementioned stages, to statistically represent lipid accumulation in the cultures. A novel aspect in the present modeling is the stochastic representation of morphologies of cells and intracellular organelles, which is obtained automatically through our custom-made Matlab code. This is currently not possible to implement in standard (commercial) software packages for building the geometry of FE models.

In the single-cell-in-a-construct (SCinC) model geometries, cells with radii of 9.6–24.4 μm were located at the center of a cubic scaffold with edge sizes of 40–100.8 μm (Fig. 1), with the specific edge length being at least four times the cell diameter. A total of 18 SCinC geometries were generated, of which 6 adipocytes were at an initial stage, 6 other adipocytes were at a middle stage, and the last 6 adipocytes were at an advanced stage of differentiation (Fig. 1 and Table 1).
All elements were 4-node linear tetrahedrons.
LD, lipid droplet; PM, plasma membrane; SD, standard deviation.
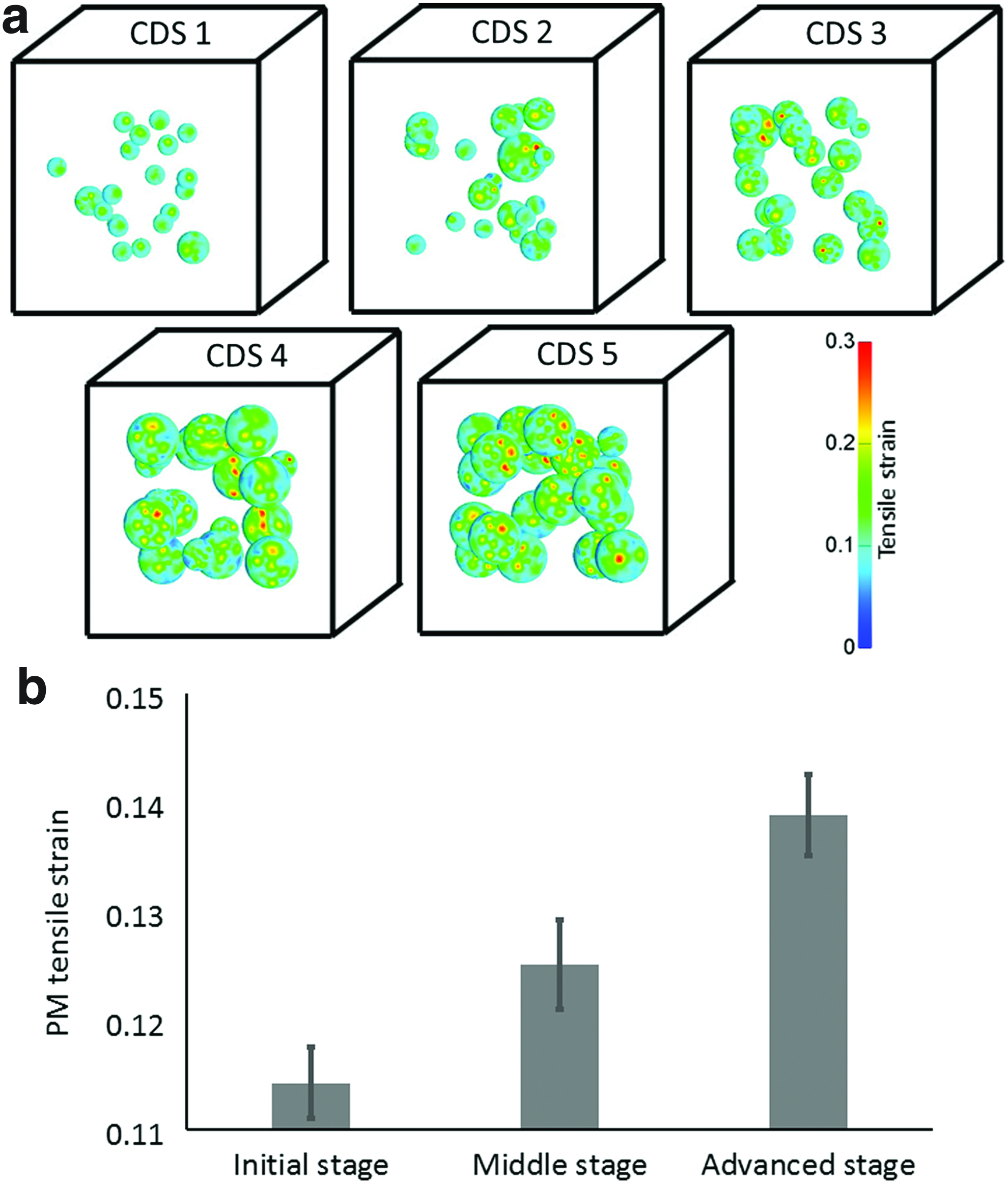
Ten multiple-cells-in-a-construct (MCinC) model geometries were further generated where each model included 20 cells spatially distributed in a 3D cubic scaffold with edge sizes of 118–250 μm (Table 2). The sizes of the cubes were adjusted to the maximal cell size so that edges were always five times longer than the maximal cell diameter. The positions of the cells in the constructs and the levels of maturation of the individual cells in the MCinC geometries were determined stochastically to reflect variability in maturation levels and intracellular contents (Figs. 2 and 3). Each simulated culture was characterized by a culture differentiation stage (CDS) index (1<CDS<5), which was determined from the mean cell size in the specific model variant. A greater CDS index represented a more differentiated 3D culture with larger cells (Fig. 2). Specifically, the mean±SD of the cell radius was 10.14±1.66, 12.23±3.87, 15.2±2.48, 20.37±5.19, and 23.46±2.89 μm in CDS models 1,2,3,4, and 5, respectively. For determining the influence of the volumetric cell density on the levels of tensile strains developing in the PMs of the cells, each MCinC model variant was assigned two sizes of a bounding cube (i.e., total scaffold dimensions): (1) the minimum cube size able to include all the cells in the model, and (2) a constant size of 250 μm3 (width×depth×height), which was the minimum cube able to contain 20 cells at an advanced stage of differentiation, and additional geometrical parameters, which are provided in Table 2. For numerically solving the mechanical loading conditions in the 3D constructs and the stretching levels of the PM in the embedded cells, in particular, all models were meshed with tetrahedral elements using the ISO2MESH toolbox 41 in Matlab (Tables 1 and 2). Special care was taken to refine meshes in LDs and the PM of cells. Convergence tests yielded that greater mesh densities resulted in less than a 1.5% change in our outcome measures.
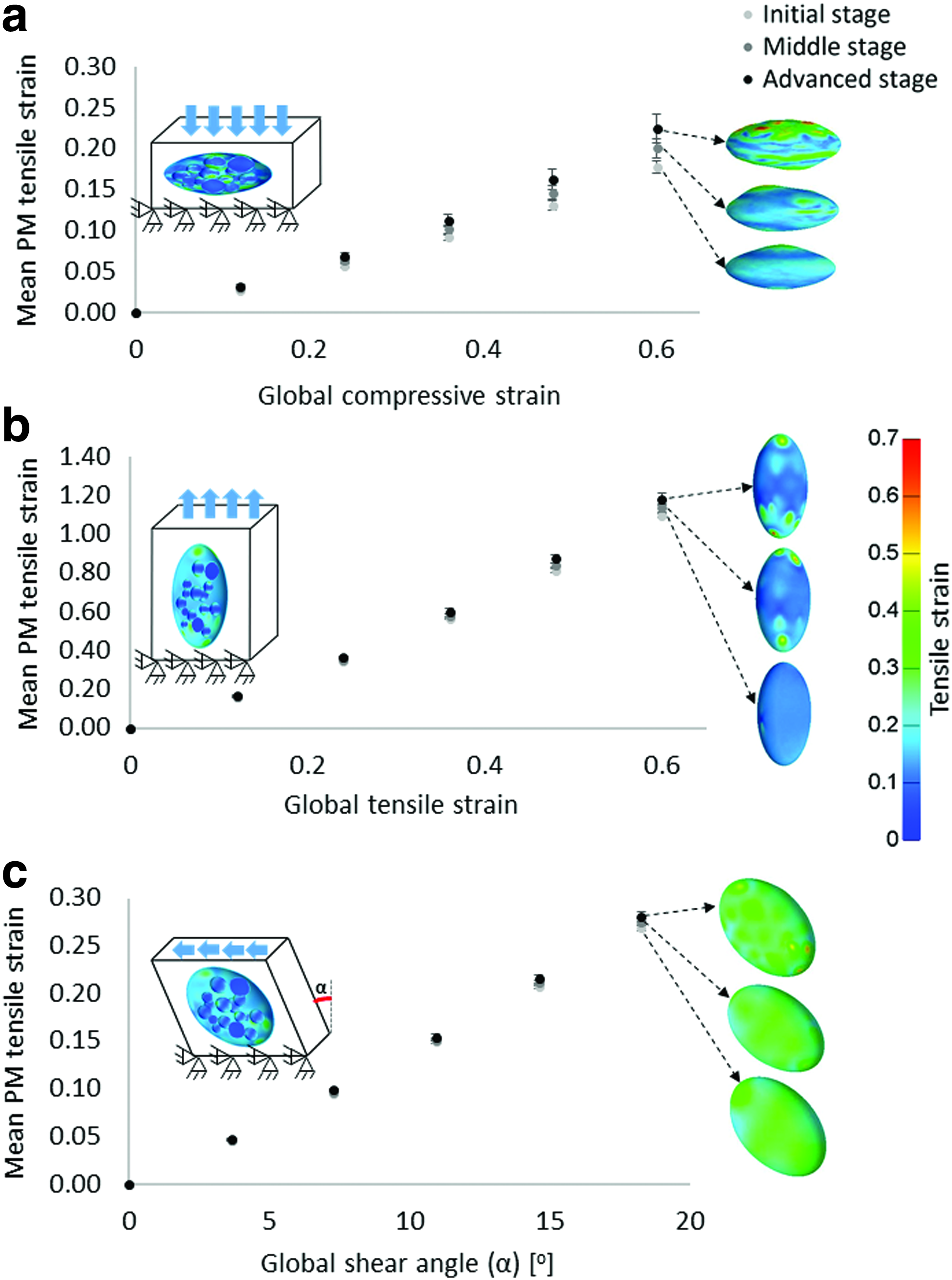
The levels of tensile strains developed in the plasma membranes (PMs) of the cells in the single-cell-in-a-construct simulations when global compression
All elements were 4-node linear tetrahedrons.
CDS, culture differentiation stage.
Mechanical properties of the model components
All components were assumed to be isotropic compressible materials that behave according to a Neo-Hookean strain energy density function (Table 3). A Poisson's ratio of 0.45 and elastic moduli of 5, 4.17, and 0.77 kPa were set for the nuclei, LDs, and cytoplasm (including the PM), respectively. 42 An elastic modulus of 2.28 kPa and Poisson's ratio of 0.3 were assigned to the scaffold material. 28
Loading and boundary conditions and protocol of simulations
Large compression, tension, and shear strains were applied quasi-statically to the SCinC model variants by lowering, elevating, or horizontally translating the superior face of each cubic VOI while keeping the inferior face fixed for all motions. The magnitudes of the loadings, which included global compression or tension strains of up to 0.6 or a global shear angle of up to 20o, were consistent with physiological strains developing in weight-bearing adipose tissues in vivo.7–8 Considering that in the SCinC simulations the level of maturation of the cells had the most substantial influence on the results in the compression mode and that compression is a primary loading mode in vivo,7–8 all the MCinC simulations were loaded quasi-statically in compression up to a global compressive strain of 0.4. Tie constraints were imposed at all material interfaces in all simulations. Using FEbio (http://mrl.sci.utah.edu/software/febio) in its structural mechanics mode, we ran all computations on a 64-bit Windows 7-based workstation with an Intel Core i7 920 2.57 GHz CPU and 64GB of RAM. Runtimes ranged between 1–7 h, depending on the model size. The input and output parameters of the computational modeling are specified in Table 4.
Outcome measures and data analysis
We analyzed tensile strains in the PM of the cells as follows: For the SCinC simulations, we compared tensile strains developed in the PM across the cell differentiation stages (initial, middle, advanced; Fig. 1); For the MCinC simulations, we not only made the same comparisons but also separately calculated the tensile strain levels in the less mature cells in each model, that is, in the cells that are smaller than the mean cell size in each model variant. Eighteen, 12, 19, 8, and 2 such less mature cells with mean cell radii of 9.6, 9.6, 14.72, 14.33, and 15 μm were considered in CDS models 1,2,3,4, and 5, respectively.
Cell culturing
Evaluating intraculture variability in cell differentiation levels
Mouse embryonic 3T3-L1 preadipocytes (American Type Culture Collection) at passage 8 were seeded at a density of 5×104 cells/well in six wells. The growth medium (GM) consisted of high-glucose Dulbecco's modified Eagle's medium (4.5 mg/mL; Biological Industries), 10% fetal bovine serum (Biological Industries), 1% L-glutamine (Biological Industries), 0.1% penicillin–streptomycin (Sigma), and 0.5% 4-(2-hydroxyethyl)-1-piperazineethane -sulfonic acid (Sigma). The GM was changed twice a week. When the cultures reached a confluence of ∼90%, the following differentiation factors were added to the GM: 3.15 μg/mL insulin, 1 μM dexamethasone, and 2 μM 3-isobutyl-1-methylxanthine; this medium was termed a differentiation medium (DM). One to four cultures were photographed under a phase contrast microscope. At each time point, three micrographs were taken in four randomly chosen regions within each well for the purpose of evaluating the intraculture variability in differentiation levels of the cells. The micrographs were analyzed for adipogenesis levels by calculating the mean number of LDs per cell, the mean radius of LDs, and the ratio of areas occupied by lipids over the field of view, and the SDs around these means. Briefly, each micrograph was converted into a binary image, in which only LDs were colored white and everything else was blackened. Then, we manually marked cells in each micrograph and a Matlab code automatically calculated the aforementioned adipocyte differentiation parameters as described in our previous studies.20–21,40 The input and output parameters were differentiation factors and lipid production levels, respectively.
Evaluating cell differentiation in cultures depleted of differentiation factors
We used staining to evaluate cell differentiation in 3T3-L1 preadipocytes that were maintained only in the GM (without differentiation factors). Specifically, the cells were cultured in the GM as described in the Evaluating intraculture variability in cell differentiation levels section. The GM was replaced every 2–3 days. At day 24, the cells were stained with Nile red (for visualizing LDs) and 4′,6-diamidino-2-phenylindole (DAPI; for marking nuclei). Depletion of differentiation factors and lipid production levels were the input and output parameters, respectively, for these experiments.
Results
The levels of the mean tensile strains, which developed in the PMs of the cells in the SCinC simulations, increased with the level of maturation of the cells in all loading modes when global compression, tension, or shear were applied (Fig. 3). The most substantial effect of the level of cell maturation, though, was observed in the SCinC model variants that were loaded in compression (Fig. 3a). Moreover, differences in tensile strains of the PM across levels of cell maturation grew in the SCinC model variants as the magnitudes of the global loading on the constructs increased (Fig. 3). Specifically, the means±SDs of the PM tensile strains in the initial, middle, and advanced differentiation stages were 0.18±0.01, 0.20±0.01, and 0.22±0.02, respectively, when a global compression strain of 0.6 was applied (Fig. 3a). When a global tensile strain of the same magnitude was simulated, the levels of the PM tensile strains were 1.18±0.03, 1.13±0.02, and 1.09±0.01 in the initial, middle, and advanced differentiation stages, respectively (Fig. 3b). In the shear simulations, the mean tensile strains in the PM were 0.2687±0.0031, 0.2692±0.0039, and 0.2699±0.0024 in the initial, middle, and advanced stages, respectively, for a global shear angle of 20o. Although differences in absolute values between maturation conditions are slight, these data do show a consistent trend of rise in PM stretches when the constructs were subjected to increasing levels of external shear (Fig. 3c). The MCinC modeling data revealed similar trends of effects, as substantially greater PM tensile strains occurred in the more mature cells embedded in these simulated constructs (Fig. 4). When averaging the PM tensile strains across all model variants with the same external construct dimensions (model variants #1,3,5,7,9) for a simulated compression loading mode, the means±SDs of the data were 0.11±0.003, 0.12±0.004, and 0.13±0.03 for the initial, middle, and advanced stage cells, respectively (Fig. 4b). When further analyzing the analogue results in the models with the varying construct dimensions (model variants # 2,4,6,8,10), the mean PM tensile strains in the less mature cells (cells that were smaller than the mean cell size in each model variant) increased with the CDS index of the model, that is, with the mean cell radius (Fig. 5a). For example, in the CDS 1 (mean cell radius 10.1 μm) and CDS 5 (mean radius 23.5 μm) models, the tensile strains in the PM of the less mature cells were 0.106±0.004 and 0.126±0.008 (mean±SD), respectively. Furthermore, the distributions of tensile strains in the PMs of the less mature cells expanded with the CDS level of the model (Fig. 5b). Tensile strains of up to 0.18, 0.195, and 0.215 occurred in the less mature cells in the model variants with CDS 1–2, 3–4, and 5, respectively. To summarize, the PM stretch levels in these less mature cells became more inhomogeneous, within and across cells, and greater in magnitude as the culture in the construct became more differentiated as a whole (Fig. 5b). To support these computational results, we documented the differentiation patterns of 3T3-L1 cells in vitro. In these cultures, we observed substantial heterogeneity across the individual cells in the cultures exposed to the DM (Fig. 6a, b); while some areas were covered by large adipocytes (Fig. 6b), cells in other areas were yet in their preadipocyte stage (Fig. 6a). Variability in differentiation levels among the adipocytes that were already differentiating was further quantified (Fig. 6d–f). The mean radius of LDs and the lipid area ratio increased up to 1.8±0.11 μm and 0.16±0.01, respectively, at day 15, which nicely demonstrated the impact of variability across cells in our simulations (Figs. 1 and 2). Moreover, the present empirical observation that adipocytes differentiate and produce large LDs in cultures even without any differentiation factors (Fig. 6c) indicates that there is spontaneous differentiation as cells become denser, which is consistent with the modeling results and our theory.

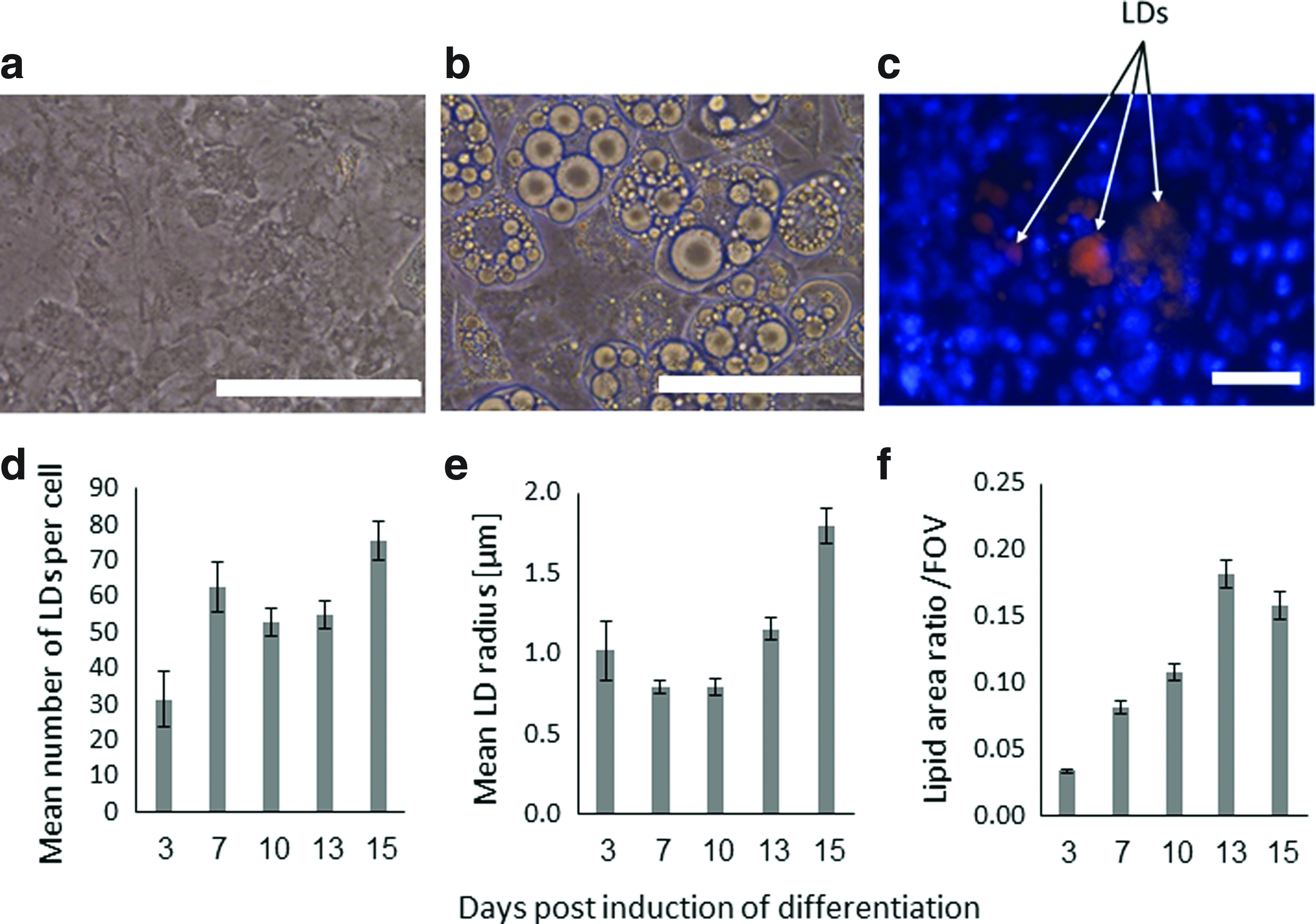
Micrographs demonstrating undifferentiated
Discussion
Using FE MSM we found that under a given mechanical loading state, the levels of tensile strains that are developing in the PM of adipocytes increase with the levels of cell growth and maturation. In other words, cells become structurally stiffer as they grow and accumulate lipid mass during differentiation.28,42 Our results were consistent in both the SCinC and MCinC simulations. Furthermore, we found that differentiation of adipocytes can lead to higher magnitudes of tensile strains in the PMs of the other (less mature) nearby cells, again when there is continuum-level loading. Our experimental observations further support this hypothesis by showing intracellular LDs in superconfluent preadipocyte cultures that were depleted of differentiation factors. In other words, preadipocytes in the dense cultures were deformed by adjacent cells, which triggered spontaneous differentiation and considerable adipogenesis.
Deformations of the PM lead to activation of mechanosensitive signaling that triggers the downstream signal cascades involved in adipogenesis.6,9–21 Interestingly, while dynamic loading modes such as cyclic stretching or vibration were generally found to suppress adipogenesis, for example, in preadipocytes from cell lines, mesenchymal stem cells, and adipose tissue stromal cells,9–18 static mechanical stretching was found to substantially accelerate adipogenic differentiation.6,19 Hence, we suggest that when differentiating adipocytes embedded in a construct are being mechanically loaded, deformations imposed by more mature structurally stiffer adipocytes on neighboring less differentiated cells will accelerate the overall adipogenesis.
This mechanism needs to be studied further, given its potential use in adipose TE in vitro and in applications where fat tissue mass needs to be enhanced or repaired in vivo (e.g., in breast enlargement or reconstruction and wound repair). Indeed, the FE MSM approach was taken in TE research of cartilage, bone, or cardiac. Kelly and Prendergast, 29 for example, used FE MSM to optimize the production of engineered cartilage for repairing osteochondral defects. In their work, a mechanoregulation algorithm for describing the process of cell differentiation in cartilage constructs was used to determine the influence of scaffold material properties on chondrogenesis. They found that increasing the stiffness of their scaffold in the simulations (within a certain range) resulted in more cartilage formation and reduced the amount of fibrous tissues in the defect. Likewise, FE MSM of bone TE was considered by Milan et al. 30 and Byrne et al. 31 In the Byrne article, optimal properties for a scaffold that maximizes differentiation of osteoblasts were again sought by simulating the influence of parameters like the porosity and dissolution rate of the scaffold. In the Milan article, fluid dynamics was further taken into account to simulate flow in polylactic acid–calcium phosphate glass scaffolds, which allowed determining the perfusion inlet velocity and compression strain rate magnitudes that can either promote or hinder cell proliferation, osteoblast differentiation, and osteogenesis. Another example is the study of Jean and Engelmayr, 34 in which FE models of accordion-like honeycomb scaffolds designed for cardiac TE were used to determine their elastic effective stiffness and their anisotropy properties. The modeling allowed these authors to design variants of the scaffold that simultaneously provided proper cardiac anisotropy and reduced the directional stiffness to enhance heart cell-mediated contractility. Zahedmanesh and Lally 35 also used FE modeling to investigate the influence of scaffold compliance and loading regimes for regulating the growth of vascular smooth muscle cells in tissue-engineered intimal hyperplasia. The above literature and our results indicate the potential for achieving a cost-effective optimized design of tissue-engineered fat constructs by employing FE MSM as opposed to the trial and error approach, which was common in the earlier days of TE.
The present modeling is supported by our in vitro results, particularly those, which demonstrate spontaneous adipocyte differentiation in dense confluent cultures. Our results obtained using the 3T3-L1 line were consistent with those reported in other cell lines, including in Ob1754, Ob1771, and 3T3-F442A cells, where all these cell cultures were deprived of differentiation factors.43–48 We hence assume that the cell-to-cell mechanical forces that were produced en mass by the cells when pushing against each other activated signaling pathways, which are responsible for the spontaneous differentiation process, thereby providing a new description for a mechanism of fat tissue growth.
Our results are consistent with previous research, in which mechanical strains were calculated in and around loaded adipocytes using FE analyses.36,49 In previous work of our group, Or-Tzadikario and Gefen 49 used a cell-specific modeling approach and created geometrically realistic FE cell models of a single undifferentiated preadipocyte and a single differentiated adipocyte from z-stack confocal microscopy scans. Similarly to our data, greater tensile strains occurred in the PM of the differentiated adipocyte with respect to the preadipocyte model when an equivalent global compressive deformation was applied. In a later study, Ben-Or Frank et al. 36 used a set of 2D models of multiple adipocytes embedded in the ECM. They found that when the constructs were loaded in compression and/or shear deformations, the levels of localized deformations in the PM increased with the levels of maturation of the neighboring cells, but their work was limited to a more simplified 2D geometrical representation. In this study, we extended the approach considerably to 3D geometries and determined the positions of the cells and intracellular nuclei and LDs stochastically to represent the inherent variability in cell sizes and intracellular contents. We appreciate that the modeling parameters may further depend on culture/construct-specific conditions, for example, the biochemical composition of the medium and ECM, presence of growth factors, and the mechanical properties of the ECM. 40 Nevertheless, the trends of effects that were demonstrated here should not be affected given that such conditions are kept consistent across trials. The automatic process of generation of the present models further allows accounting for the influence of such specific culture conditions as well, pending that the relevant empirical data are available.
We surmise that a cell biomechanics deformation-driven positive feedback loop of adipogenesis is very likely to be involved in differentiation of adipocytes in mechanically loaded constructs and tissues (Fig. 7). At the cell scale, adipocytes that differentiate and accumulate LDs become effectively stiffer. 42 These structural changes at the individual cell level lead to greater tensile strain magnitudes in the PM of the differentiating cells. The elevated PM strains are very likely to then lead to cytoskeletal rearrangement and activation of different signaling pathways that translate the mechanical stimuli to a differentiation process. In addition, and importantly, when cells are differentiating, tensile strains in the PMs of the adjacent cells increase locally, which again appear to be involved in activation of mechanotransduction signaling pathways that result in acceleration of differentiation in such neighboring less mature cells. The outcome at the cell population level is a progressive culture differentiation where the more mature cells push the adjacent less mature cells to differentiate as well, by distorting and deforming them.

The deformation-driven positive feedback loop of adipogenesis. At the cell scale, adipocytes that differentiate and accumulate LDs become effectively stiffer. These structural changes at the individual cell level lead to greater inhomogeneity of tensile strains and also to greater tensile strain magnitudes in the PM of the differentiating cells. The elevated PM strains, in turn, are very likely to then lead to cytoskeletal rearrangement and activation of different mechanotransduction pathways. The differentiation process is amplified by this signaling and so on and so forth. This feedback loop plays an important role at the mesoscale as well, which is an intermediate scale between the cell (micro) and organ (macro) scales, which refers here, specifically, to a group of cells embedded within a volume of interest in an adipose construct. When cells are differentiating, tensile strains in the PMs of the adjacent cells increase locally (i.e., there are cell-to-cell mechanical interactions), which again appear to be involved in activation of mechanotransduction signaling pathways that result in acceleration of differentiation of such neighboring less mature cells.
Future work should be focused on applying different regimes of global tension/compressive and/or shear strains to 3D-engineered adipose constructs in vitro while evaluating the differentiation responses in the deformed adipocytes. MSM is currently the only feasible means to correlate continuum (macrolevel) construct deformations to subcellular-level PM stretches in the distorted cells. Developing FE MSM in this regard connects loading states at the construct level, which can be controlled in a laboratory setting, with deformations of the PM (at the subcellular scale) that are difficult to estimate directly in constructs. Revealing these macro-to-micro-to-nano relationships characterizing the influence of mechanical loads on adipocyte differentiation, that is, by adopting a multiscale approach to adipose TE, is extremely important for basic understanding of mechanotransduction phenomena in bioartificial and native fat tissues, and also, from a more practical point of view, for optimizing TE protocols in this field and the respective experimental costs.
Footnotes
Acknowledgments
This research work is being supported by a grant from the Israel Science Foundation (no. 611/12, A.G. and D.B.) and by the Israel Ministry of Science and Technology (Women in Science Scholarship awarded to N.S.).
Disclosure Statement
No competing financial interests exist.