
Abstract
Mesenchymal stem cells (MSCs) usually lose their stemness during in vitro expansion as they are deprived of their niche environment. Cell–extracellular matrix (ECM) interaction is known to play important roles in preserving the stemness of the cells in their stem cell niche environment. Previously, coating with bone marrow MSC (BMSC)-derived ECM was found able to maintain the differentiation potential of in vitro cultured MSCs. This study aimed to determine if this ECM coating could also maintain the stemness of cultured murine adipose-derived stem cells (ASCs) using a regular culture flask as a control. Cells were expanded in ECM-coated and ECM-noncoated flasks for two and four passages and then harvested for various analyses. The results showed that ASCs exhibited fibroblast-like spindle morphology in ECM-coated flasks, whereas ASCs gradually spread and enlarged in the ECM-noncoated flasks. After three and five passages, both groups of cells exhibited similar cytokinetics in the MSC culture medium (MesenPRO RS™ Medium). However, when cultured in Dulbecco's modified Eagles medium (DMEM) plus 10% fetal bovine serum, coating group cells exhibited more potent proliferation than control group cells with a significant difference in both passages 3 and 5 (p<0.01). When seeded at low density (500 cells/10-cm dish), coating group cells formed significantly more and larger sized cell colonies than control group cells with significant difference in cell colony numbers between two groups (p<0.05). In addition, coated colony cells were much smaller and more compactly arranged compared to control colony cells. Furthermore, ASCs expanded in coated flasks exhibited greater potentials for adipogenic, osteogenic, and chondrogenic differentiations than the cells expanded in regular flasks. Quantitatively, the Oil Red O staining area, Alizarin staining area, and Toluidine Blue staining area were all significantly larger than the respective staining areas of control cells (p<0.05). Real-time polymerase chain reaction also revealed significantly higher gene expression levels of peroxisome proliferator-activated receptor gamma (PPARγ), adipocyte protein 2 (aP2), CCAAT/enhancer-binding protein (C/EBP), Rux2, osteocalcin, Sox9, collagen II, and aggrecan in ECM-coated group cells than in control group cells (p<0.05). Collectively, these results suggest that human BMSC decellular ECM coating helps to preserve the stemness of cultured murine ASCs.
Introduction
A
The essential characters of adult MSCs are their multiple differentiation potential and self-renewal function, and these render them an advantageous cell source for tissue regeneration. Multiple differentiation enables MSCs to differentiate into desired cells types, whereas self-renewal function helps MSCs to maintain their stemness during cell division within their in vivo niche environment. 2
One of the prerequisites for MSC application is the need of in vitro expansion to acquire sufficient quantity for therapeutic purpose. However, in vitro culture deprives MSCs of their niche environment, leading to the loss of stemness of cultured MSCs. 3 For example, expanded MSCs at later passages usually lose the differentiation potential and the capability of cell colony formation, 4 and these problems pose a big challenge to MSC application.
It has been a hot research subject regarding how to expand functional MSCs during in vitro culture. Application of growth factors 5 and small molecules 6 is the common method with observable benefits. For example, MSCs are known for their ability to produce growth factors for their biological functions. It was found that the expression of epidermal growth factor, fibroblast growth factor (FGF), and hepatic growth factor (HGF) was down regulated during MSC passage, whereas treatment of FGF increased cell proliferation and HGF could enhance MSC adipogenesis. 5 In a study, Al-Habib et al. demonstrated that the developed small molecules (Pluripotin [SC1], 6-bromoindirubin-3-oxime, and rapamycin) helped human dental pulp stem cells to preserve the cell self-renewal capacity, suppressed differentiation, and decreased cell proliferation. The expressions of STRO-1, NANOG, OCT4, and SOX2 were significantly enhanced, while the differentiation potentials of odonto/osteogenic, adipogenic, and neurogenic lineages were diminished during in vitro culture. 6
Coating of extracellular matrix (ECM) molecules is also one of the ways to maintain the stemness of cultured MSCs. Mauney et al. demonstrated that culture of human BMSCs (hBMSCs) on denatured collagen I was able to preserve the MSC adipogenic potential during ex vivo expansion and the expressions of adipocyte-related markers (fatty acid-binding protein-4, lipoprotein lipase, acyl-CoA synthetase, adipsin, facilitative glucose transporter-4, and accumulation of lipids) were enhanced.
7
Likewise, the topographical structure is also an important part of the niche environment. Jia et al. demonstrated that electrospun poly-
In 2007, Chen et al. reported an interesting culture system to preserve the stemness of expanded BMSCs by culturing the cells on BMSC-derived ECMs. They found that this particular ECM environment not only facilitated the expansion but also prevented osteogenic differentiation of cultured BMSCs. 9 This technique has also been used in inducing human bone marrow-derived MSCs into a smooth muscle cell lineage. 10 Apparently, this technique became an important method to mimic the niche environment to preserve the stemness of cultured BMSCs.
ASCs are an important source of MSCs. The comparison between BMSCs and ASCs provide interesting findings. Semon et al. showed that both cell types had similar efficiency in the treatment of experimental autoimmune encephalomyelitis, 11 whereas BMSCs were more potent than ASCs in the treatment of experimental systemic inflammation. 12 In terms of induced differentiation, BMSCs exhibited stronger osteogenic differentiation potential, 13 whereas ASCs revealed more potent angiogenic potential. 14
Compared to BMSCs, ASCs have the unique advantages as they are much stronger in their proliferation capability, easier to access, and more abundant in their resource with no concern of donor-site morbidity. 15 Therefore, ASCs have become a widely applicable cell source for cell therapy 16 and tissue engineering.17,18
Similar to BMSCs, loss of phenotype and functions is also the major limitation of in vitro expanded ASCs for their application. Much endeavor has been made to deal with this issue by using growth factors 19 and hypoxia condition. 20 Chen's method as well as other derived methods were an interesting try to preserve the stemness of cultured BMSCs and proven to be efficacious in maintaining the multiple differentiation potentials and directing desired cell fate.9,21 It remains unexplored whether this system is also applicable to ASCs. This study aimed to investigate if the coating of culture dish with hBMSC-derived decellular ECM would help to retain the stemness of in vitro cultured mouse ASCs.
Materials and Methods
Animals
Female C57BL/6 mice, 6 to 8 weeks old, were purchased from the Slac Laboratory Animal Center (Shanghai, China). An Institutional Review Committee of the Shanghai Jiao Tong University School of Medicine approved all animal study protocols.
Generation of decellularized BMSC-derived ECM
The culture flasks coated with decellularized hBMSC-derived ECM were generated as previously reported. 21 In this study, all human decellularized ECM-coated flasks were a gift kindly provided by Dr. Xiaodong Chen from the University of Texas Health Science Center at San Antonio and were prepared in Dr. Chen's laboratory. The human ECM-coated flasks were distributed by Shanghai Stemsan Biotechnology Ltd. Briefly, passage 1–2 human bone marrow mononuclear cells were seeded onto tissue culture flasks and expanded with the addition of ascorbic acid until confluence. After extensive wash with phosphate-buffered saline (PBS), the flasks were further treated with 0.5% Triton X-100 containing 20 mM NH4OH in PBS to remove cellular component followed by the treatment of antibiotics and antifungus drugs and stored in 4°C.
Cell culture
In this study, pooled ASCs isolated from the stromal vascular portion (SVF) were used in this study. A total of 18 mice were involved, and fat tissues collected from three mice were pooled to derive one cell sample; in total, six cell samples were derived from the fat tissues of these mice. Briefly, as previously described, 22 subcutaneous adipose tissues were harvested from the inguinal and armpit areas of C57BL/6 mice. The fat tissues were cut into small fragments followed by enzyme digestion with 0.075% collagenase (NB4, Serva, France) in serum-free Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan City, UT) at 37°C for 2 h. The cell suspension was harvested and filtered through a 40-μm nylon filter mesh (BD Falcon, Bedford, MA) to remove tissue residues. The filtrate was centrifuged, and cell pellets were resuspended in the MesenPRO RS™ Medium (Gibco-BRL, Grand Island, NY) or DMEM plus 10% fetal bovine serum (FBS; Hyclone) and seeded onto regular flasks (BD Falcon) or onto human decellularized ECM-coated flasks (kind gift from Dr. Xiaodong Chen) at a density of 3×105 cells/cm2 for primary culture and 2×104 cells/cm2 for subculture. The medium was changed every 3 days. When cultured primary cells of both groups reached about 80% confluence, they were detached with the treatment of PBS containing 400 U/mL type II collagenase (Worthington Bio-chemical, Lakewood, NJ) for 10 min at 37°C followed by subculture. Expanded cells of both groups at the end of passages 2 and 4 were used for the experiments.
Cytokinetics analysis by cell counting kit-8 assay
ASCs of passages 2 and 4 were, respectively, detached from the uncoated or coated culture flasks and plated onto 96-well plates at 2×104 cells per well as passages 3 and 5, respectively, to develop cell growth curves in the MesenPRO RS Medium. Briefly, 10 μL of sterile cell counting kit-8 (CCK-8) solution (Dojindo Laboratories, Kumamoto, Japan) was added to each well and incubated for 2 h at 37°C and then the medium was harvested for measuring the optical density values against a reagent blank control at a wave length of 450 nm using a microplate reader (Varioskan Flash; Thermo Scientific, Waltham, MA). The test was performed on days 0, 2, 4, 6, 8, and 10 in quintuplicate and repeated in three cell samples. To investigate the effect of the culture medium on cytokinetics difference, primary ASCs were also cultured in DMEM plus 10% FBS either on regular flasks or on human ECM-coated flasks for two and four passages, and then the cells were harvested and plated onto 96 wells in the same medium as passages 3 and 5, respectively, for cytokinetics assay in the same way as it was performed in the MesenPRO RS Medium.
Colony-forming unit-fibroblast assay
Colony-forming unit-fibroblast (CFU-F) assay was performed using a modified technique as described previously. 23 Passages 2 and 4 cells were detached from uncoated and from human ECM-coated culture flasks, respectively, and then cultured in 10-cm Petri dishes in DMEM plus 10% FBS at a density of 500 cells per dish. The medium was changed every 3 days. After 14 days of culture, the cells were fixed with 4% paraformaldehyde, and CFU colonies were visualized with crystal violet staining. The number of colonies (diameter ≥2 mm) was counted. Totally, 10 dishes were included in each group for each cell sample test, and the experiment was repeated in three cell samples.
Multipotent differentiation of ASCs in different culture systems
As previously described,
24
passages 2 or 4 ASCs were cultured to confluence in the MesenPRO RS Medium, and then they were, respectively, detached from uncoated or coated culture flasks and plated onto six-well plates for another passage, respectively, as passage 3 and passage 5. When the cells reached 50% confluence, they were subject to multidifferentiation induction by adding induction media. For adipogenic differentiation, cells were cultured in 10% FBS-containing DMEM and supplemented with 250 nM dexamethasone (Sigma-Aldrich, St. Louis, MO), 66 nM insulin (Sigma-Aldrich), and 0.5 mM isobutylmethylxanthine (IBMX; Sigma-Aldrich). After 7 days, the cells were stained with Oil Red-O. For osteogenic induction, cells were cultured in 10% FBS-containing DMEM supplemented with 10−7 M dexamethasone (Sigma-Aldrich) and 10−4 M
qPCR analysis
Total RNA was extracted using the TRIzol Reagent (Invitrogen, Carlsbad, CA), and complementary DNA was synthesized from 2 μg total RNA per sample using AMV reverse transcriptase (Takara, Kyoto, Japan). The reaction was made in a final volume of 20 μL with 4 μL MgCl2, 4 μL 5× buffer, 2 μL dNTP, 0.5 μL RNase inhibitor, 1 μL oligo-(dT), and 0.5 μL AMV reverse transcriptase, using ddH2O to meet the final volume. The mixture was incubated at 30°C for 10 min, 45°C for 60 min, 98°C for 5 min, and 5°C for 5 min. As previously described, 23 qPCR was performed using a Power SYBRGreen PCR master mix (2×) (Applied Biosystems, Foster City, CA) in a real-time thermal cycler (Stratagene Mx3000PTM QPCR System, La Jolla, CA). It was performed with incubation at 95°C for 3 min followed by 40 cycles (30 s at 95°C, 30 s at 60°C, and 45 s at 72°C), terminated by a 5-min extension at 72°C, and stored at 4°C until analysis. The analysis was conducted in triplicate for each sample and the experiments were repeated in three cell samples. Gene expression was normalized to GAPDH expression using the 2−ΔΔC(t)method. The designed primers for qPCR analysis are listed in Table 1.
Flow cytometry
Single-cell suspensions (1×106) were incubated in 100 μL of diluted PE-conjugated rat anti-mouse CD44, CD90, and CD105 antibodies (1:100 in PBS containing 1% FBS; BD Biosciences, San Jose, CA) for 30 min at 4°C. As a control, cell suspension was incubated in 100 μL of diluted PE-conjugated rat anti-mouse isotype IgG (1:100 in PBS containing 1% FBS; BD Biosciences) for 30 min at 4°C. After two washes with PBS containing 1% FBS, cells were subject to flow cytometry analysis using a Becton Dickinson FACStar.
Statistical analysis
Data are presented as mean and standard deviation. Student's t-test was used for statistical analysis of all paired comparisons and a p-value less than 0.05 was considered statistically significant.
Results
Comparison of cell morphology between ECM-coated and ECM-noncoated groups
Pooled ASCs were isolated from SVF and seeded, respectively, onto ECM-noncoated and human ECM-coated culture flasks in the MesenPRO RS Medium. The medium was changed after 3 days to remove nonadherent cells. When observed under a microscope, there was no apparent difference in cell morphology for both groups of cells, mainly in the spindle shape (Fig. 1A, D). With cell passage, these cells became relatively spread in noncoated flasks with a decreased ratio of cell length to width (Fig. 1B, C). Cells of human ECM-coated flasks became elongated and exhibited a typical fibroblast-like shape (Fig. 1E, F).With further cell passage, cells in noncoated flasks exhibited a bigger cell size than the cells of the ECM group. In addition, coated cells also lost typical fibroblast-like morphology gradually.
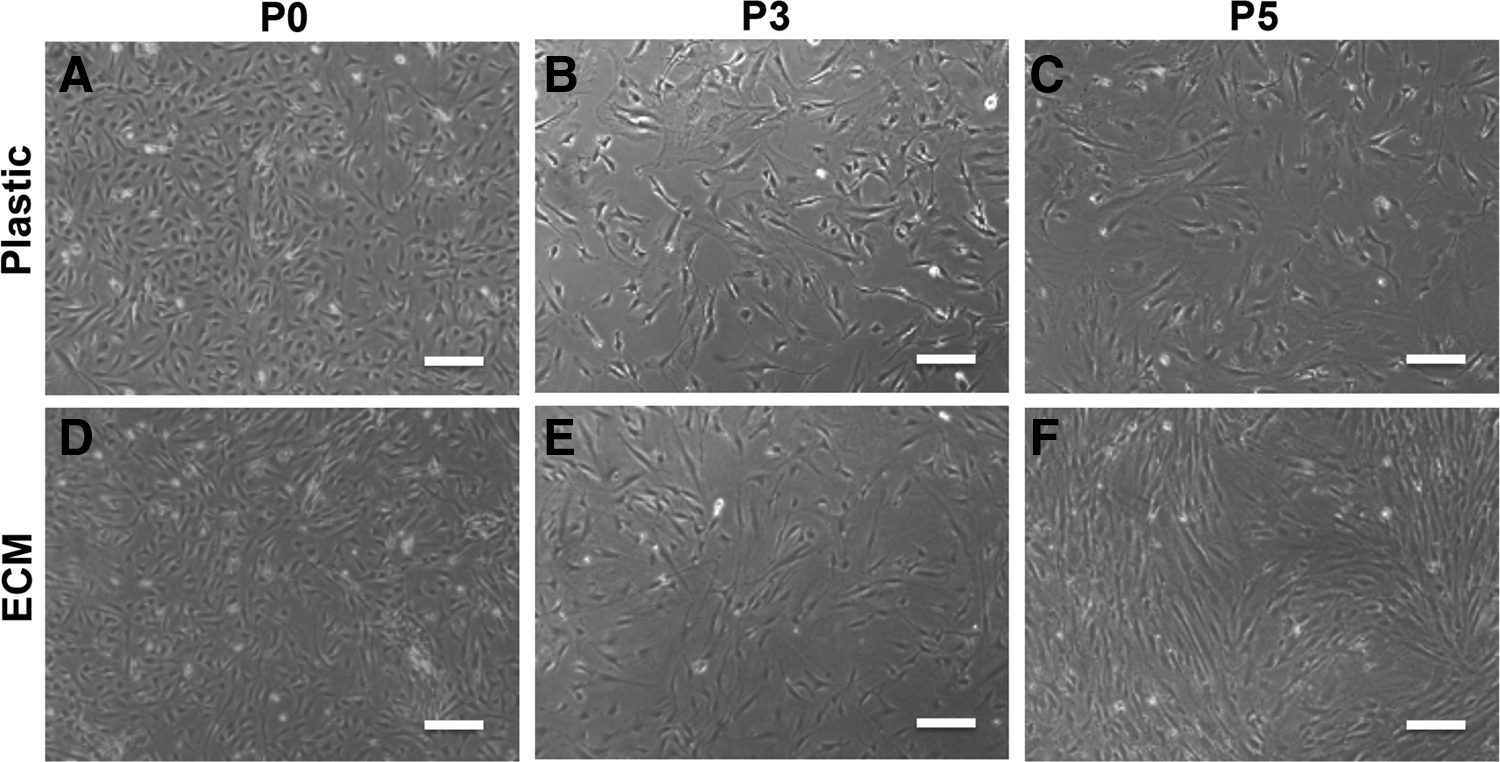
ASC morphology comparison between ECM-coating group and Plastic (ECM-noncoating) group. Both groups of cells exhibited spindle cell shape in primary cells
Comparison of cell proliferation between ECM-coated and ECM-noncoated groups
Cell proliferation capacity was examined by the CCK-8 assay. When cells were cultured in the MesenPRO RS Medium, CCK-8 assay revealed similar cell growth patterns between two groups at both passage 3 (Fig. 2A) and passage 5 (Fig. 2B) with no significant difference (p>0.05) in most time points, although a slight but significant difference was found in some time points (day 4 of passage 3 and day 10 of passage 5 with faster proliferation in the Plastic group; day 4 of passage 5 with faster proliferation in the ECM group, p<0.05). However, when ASCs were cultured in DMEM plus 10% FBS, CCK-8 assay revealed much faster cell proliferation of the human ECM-coated group than the Plastic group at both passage 3 (Fig. 2C) and passage 5 (Fig. 2D) with significant difference (p<0.01) in most time points except for day 6 of passage 3 (p>0.05).
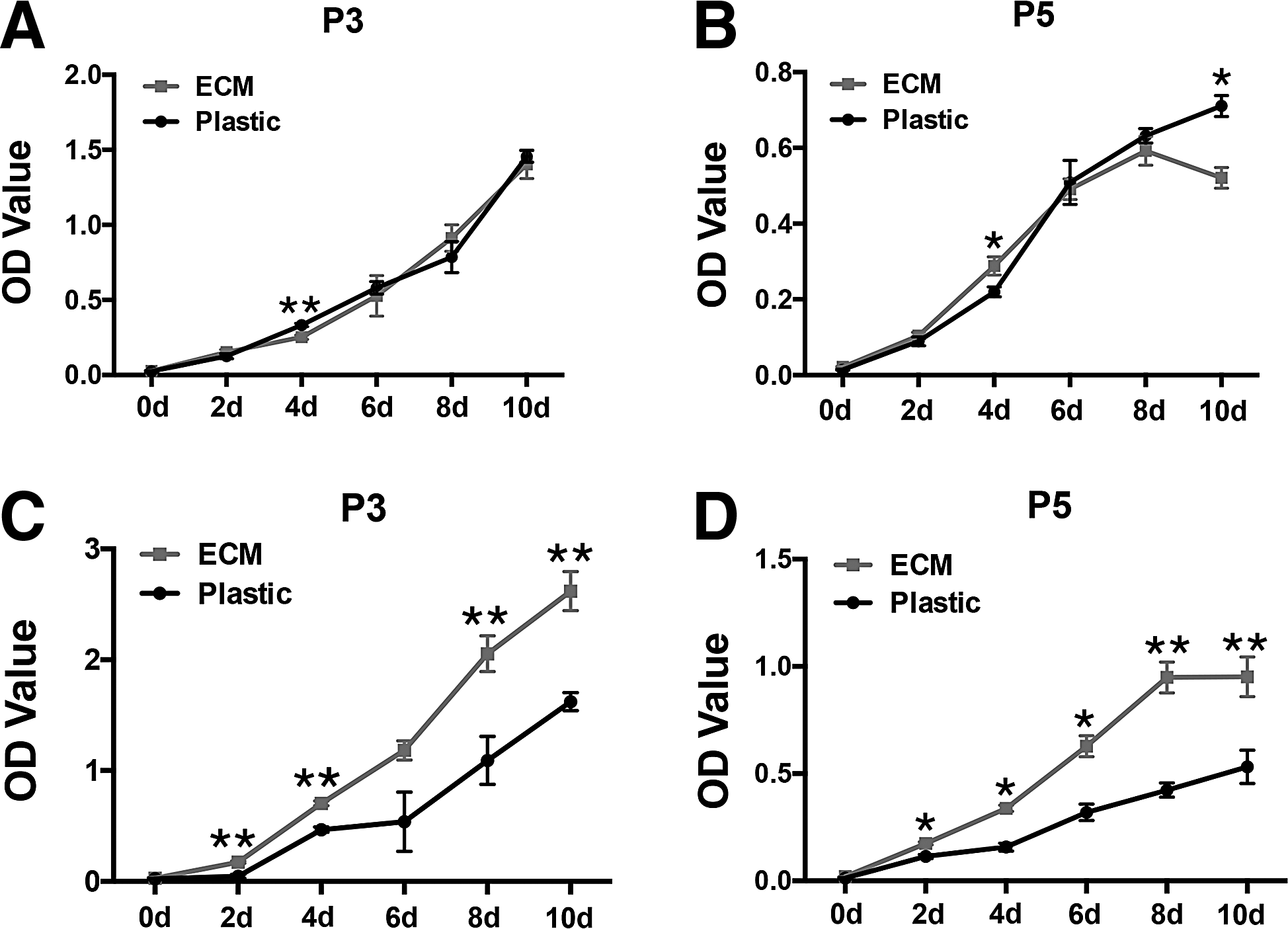
CCK-8 assay for cytokinetic comparison. ASCs expanded for three
Comparison of colony-forming ability between ECM-coated and ECM-noncoated groups
Cells were expanded on both ECM-noncoated and human ECM-coated flasks until passages 2 and 4, respectively, and then they were harvested and seeded onto regular Petri dishes at a low density (500 cells/10-cm dish). After one more passage culture, cells of the human ECM-coated group showed a stronger colony-forming ability than the cells of the ECM-noncoated group (Fig. 3). It was observed that colonies formed by the human ECM-coated group were bigger with higher cell density when compared to the colonies of the ECM-noncoated group. Statistical analysis also revealed that averaged colony number per dish reached 20.20±4.78 in the ECM group at passage 3, which was significantly higher than the averaged colony number of the Plastic group (5.68±0.29, p<0.05). At passage 5, the colony-forming ability generally decreased relatively to passage 3 cells, and the averaged colony number of the ECM group (5.23±0.54) remained significantly higher compared with the Plastic group (2.57±0.67, p<0.01). It was also noted that cells of the control colony became larger in their size and lost their fibroblast morphology when compared to the morphology of ECM colony cells.

Comparison of colony-forming ability.
Comparison of adipogenic differentiation potential between ECM-coated and ECM-noncoated groups
Both groups of cells at passages 2 and 4 were detached and seeded onto six-well plates and were subject to adipogenic induction as passages 3 and 5 cells, respectively. As shown in Figure 4A, a much stronger Oil Red O staining appeared in the ECM group than in the control group after 7 days of induction when tested at both passages 3 and 5. In addition, statistical analysis also revealed a significantly larger stained area in the ECM group than in the control group with significant difference at both passages 3 (p<0.05) and 5 (p<0.01) (Fig. 4B).

Comparison of adipogenic differentiation potential. ECM cells revealed stronger adipogenic potential than Plastic cells in both passage 3 and passage 5 cells as demonstrated by Oil Red O staining
Stronger potential for adipogenic differentiation in the ECM group was also confirmed at the gene level. qPCR analysis revealed significantly higher levels of gene expression of peroxisome proliferator-activated receptor gamma (PPARγ) (Fig. 4C), adipocyte-specific gene adipocyte protein 2 (aP2) (Fig. 4D), and CCAAT/enhancer-binding protein alpha (C/EBPα) (Fig. 4E) in the ECM group than those in the Plastic group with a significant difference at both passages 3 (p<0.01) and 5 (p<0.01).
Comparison of osteogenic differentiation potential between ECM-coated and ECM-noncoated groups
Similarly, ASCs grown in both ECM-noncoated and human ECM-coated flasks were detached and seeded onto six-well plates for osteogenic induction. As shown in Figure 5A, a much stronger Alizarin Red staining appeared in the ECM group than in the control group after 3 weeks of induction when tested at both passages 3 and 5. In addition, statistical analysis also revealed a significantly larger stained area of Alizarin Red in the ECM group than in the control group with significant difference at both passages 3 (p<0.01) and 5 (p<0.01) (Fig. 5B).

Comparison of osteogenic differentiation potential. ECM cells revealed stronger osteogenic potential than Plastic cells in both passage 3 and passage 5 cells as demonstrated by Alizarin Red staining
To further confirm this phenomenon, qPCR analysis was also performed to verify the gene expression. As shown, significantly higher levels of Runt-related transcription factor 2 (RUNX2) (Fig. 5C) and osteocalcin (OCN) (Fig. 5D) in the human ECM-coated group than those in the ECM-noncoated group with significant difference in both passage 3 (p<0.05) and passage 5 (p<0.01).
Comparison of chondrogenic differentiation potential between ECM-coated and ECM-noncoated groups
As shown, after 4 weeks of chondrogenic induction, Toluidine Blue staining revealed a stronger staining (Fig. 6A) and bigger staining area (Fig. 6B) in the human ECM-coated group than in the ECM-noncoated group with significant difference at passage 3 (p<0.01) and passage 5 (p<0.01). At gene levels, qPCR analysis revealed higher expression levels of SRY (sex determining region Y)-box 9 (SOX9) (Fig. 6C, p<0.05 in p3, p<0.01 in p5), collagen type II (COLII) (Fig. 6D, p<0.05 in p3, p<0.01 in p5), and aggrecan (AGG) (Fig. 6E, p<0.01 in p3 and p5) in the ECM group than in the control group with significant difference between two groups at both passages 3 and 5.
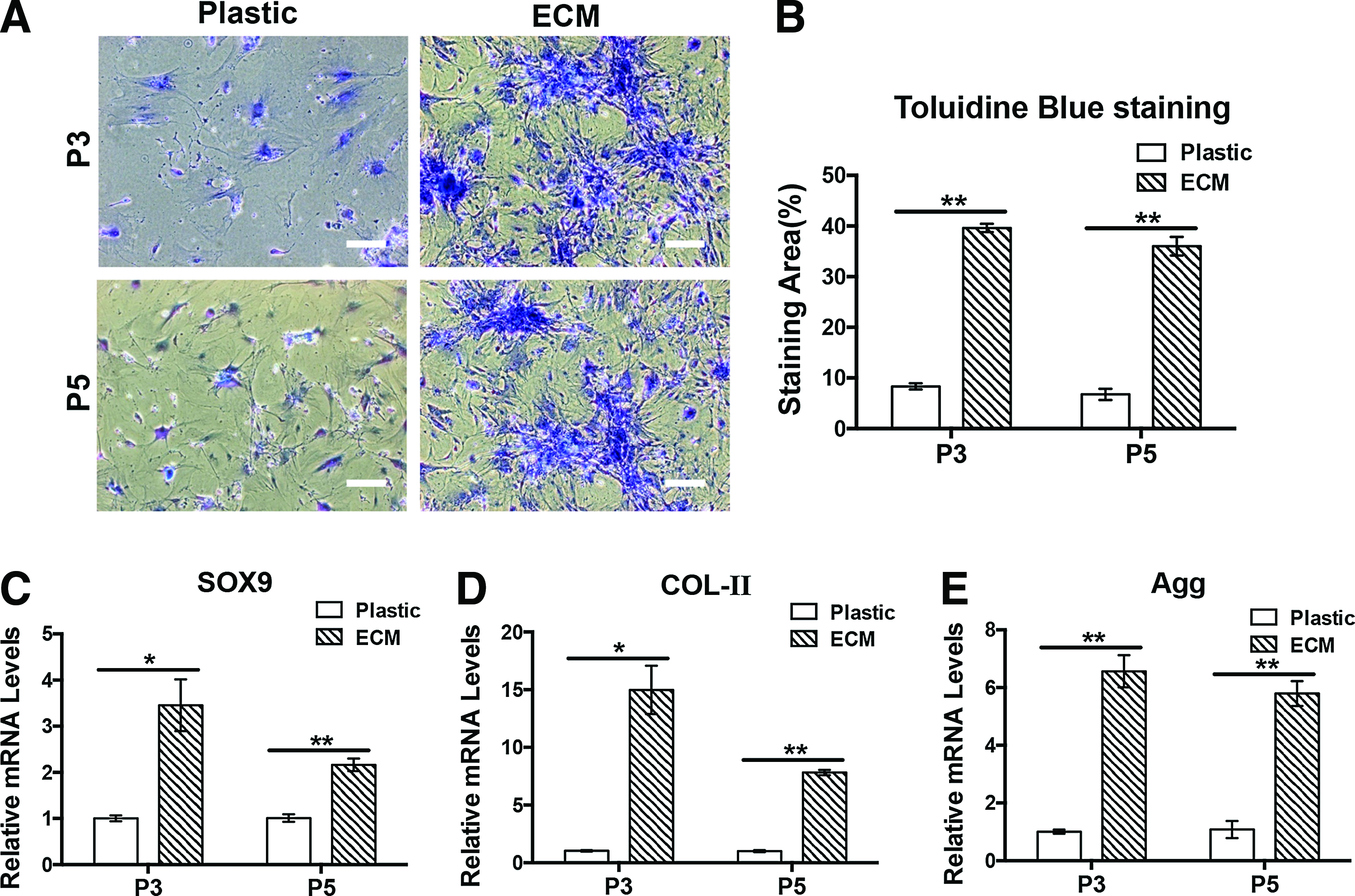
Comparison of chondrogenic differentiation potential. ECM cells revealed stronger chondrogenic potential than Plastic cells in both passage 3 and passage 5 cells as demonstrated by Toluidine Blue staining
Surface marker expression analysis between ECM-coated and ECM-noncoated groups
The MSC surface marker expression was examined with flow cytometry. As shown in Figure 7 and Table 2, there was generally no significant difference in the expression rate between human ECM-coated and ECM-noncoated groups for the three marker expressions, although CD44 exhibited slightly but significantly increased expression at passage 5 only (99.37±0.19 vs. 95.26±1.10 p<0.05). In addition, fluorescence staining intensity of CD 44 was apparently stronger in the ECM group than in the noncoated group at passage 5.

Flow cytometry analysis of marker expression in both groups of cells. Similar expression rate was observed in both groups for CD44
ECM, extracellular matrix.
Discussion
Phenotype drift and loss of function remain a major concern for MSC application after their in vitro expansion. 25 This is likely due to the deprivation of niche environment of MSCs during their in vitro culture. 3 Mimicking the stem cell niche environment has become an important strategy for in vitro expansion of functional MSCs. Such a biomimetic approach may include growth factors, ECM, nanostructure, and matrix elasticity.
Interestingly, Chen et al. reported that the use of ECM derived from mouse or human bone marrow cells as a coating substance could well preserve the stemness of in vitro expanded BMSCs.9,21 This biomimetic approach not only provided the cells with essential matrix components of their native niche environments, such as collagen types I and III, fibronectin, biglycan, and decorin, but also the signals of matrix containing growth factors and the signals of topographical structure and elasticity of the native ECM, representing an interesting and important step toward comprehensive mimicking of niche factors.
Considering the similar characters between BMSCs and ASCs, we also investigated the effect of human marrow-derived ECM on the functions of cultured ASCs using the same ECM-coated culture flasks kindly provided Dr. Xiaodong Chen, and the results demonstrated the beneficial effect on cultured ASCs as well.
Multiple differentiation potentials and self-renewal represented by colony-forming efficiency are considered as the important characteristics of MSC stemness. We investigated passages 3 and 5 cells to observe the effect of ECM-coating system on the functions of passaged cells during in vitro expansion. As shown in Figure 3, the colony-forming efficiency was significantly enhanced in the ECM-coated group when compared to that of ECM-noncoated (Plastic) group, indicating that the ECM could better retain the self-renewal functions of cultured ASCs. As the supporting evidence, cell morphology was also better retained in the ECM group as apposed to the cell morphology of Plastic group, in which cells became larger and spread. In addition, the potentials of multiple differentiations were also better retained in the ASCs cultured on ECM-coated flasks. As shown, enhanced potentials for adipogenic (Fig. 4), osteogenic (Fig. 5), and chondrogenic differentiations (Fig. 6) were observed in the ECM-coated group than in the ECM-noncoated group.
Lai et al. reported that SSEA-4 expression could be maintained in SSEA-4 sorted BMSCs when they were cultured on ECM-coated flasks. By contrast, SSEA-4 expression dropped sharply after expansion on plastic flasks. 21 In this study, SSEA-4 expression was not observed in mouse ASCs (data not shown). Instead, we examined the expression profile of CD44, CD105, and CD90, which are considered as the common markers for MSCs or ASCs. 24 The results revealed no significant difference in general expression profiles between the two groups of cells, although the CD44 expression rate was slightly but significantly increased in passage 5 cells expanded in ECM-coated flasks (Fig. 7).
According to the literature report,26,27 all these molecules are membrane-anchored adhesion molecules and their expression rates usually increased with cell passage. Similarly, our results demonstrated that both CD44 and CD90 reached the expression rate above 95%, and CD 105 expression rate was above 60%.
Of note, this study employed adherent cells derived from pooled SVF cells to analyze the effect of ECM coating on ASC biological behaviors. Similarly, Chen et al. used marrow-derived adherent cells for the assays in their first report. 9 Chen's group later reported using SSEA-4 sorted BMSCs as a cell model to test the effect of ECM coating on stemness preservation of sorted cells, which also demonstrated the beneficial effect on maintaining SSEA-4 expression. 21 Recently, Rodeheffer et al. 28 reported murine ASCs purified from pooled SVF cells by cell sorting based on the markers of Lineage−, CD34+, CD29+, and Sca-1+ and showed that purified ASCs had more potent differentiation potential than unsorted SVF cells. It will be an interesting step of our future study to investigate the role of ECM coating on biological behaviors of purified ASCs and to examine the expression of more specific markers that are truly associated with the stemness of cultured cells and usually decrease in their expression with cell passage such as CD34 and Oct 4.
This study also showed that the culture medium is likely to play a role in retaining the stemness of cultured ASCs. Although low serum stem cell culture medium (MesenPRO RS Medium) could much better preserve the stemness of coated ASCs than the regular DMEM culture medium (data not shown), it failed to enhance cell proliferation of the coated cells. By contrast, DMEM plus 10% FBS could significantly enhance cell proliferation of coated ASCs than the control ASCs, indicating that an optimized culture system should include both ECM mimicking and an optimized culture medium.
One of the shortcomings of this study was the use of human ECM coating to test the stemness preservation on mouse ASCs. The mismatch of species may have an effect on the biological behavior of mouse ASCs, as structural difference in the ECM protein does exist between two different species, such as collagens. 29 Although positive evidence was demonstrated, we were not able to maintain cell subculture for more than 10 passages, as the mouse cells were not as robust as human ASCs that grew well in DMEM and exhibited apparently more potent cell passage capacity (data not shown). When allogeneic human cell-derived decellular ECM is applied, it will certainly function better on human cells and should be applicable to different individuals as the immunogenic cells are already removed from the produced ECM coating.
Collectively, the results of this study demonstrated the beneficial effect of human marrow cell-derived ECM on the retention of mouse ASC stemness, including self-renewal function and multiple differentiation potentials, as well as enhanced cell proliferation. In the future, tissue-matched ECM coating and species-matched studies will be the important directions.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (30872694, 31170937) and the Specialized Research Fund for the Doctoral Program of Higher Education (20130072120028). We also appreciate Dr. Xiaodong Chen from the University of Texas Health Science Center at San Antonio for providing the ECM-coated culture flask.
Disclosure Statement
No competing financial interests exist.