
Abstract
Currently, there are no established treatments to prevent, stop, or even retard the degeneration of articular cartilage in osteoarthritis (OA). Biological repair of the degenerating articular cartilage would be preferable to surgery. There is no benign site where autologous chondrocytes can be harvested and used as a cell source for cartilage repair, leaving mesenchymal stem cells (MSCs) as an attractive option. However, MSCs from OA patients have been shown to constitutively express collagen type X (COL-X), a marker of late-stage chondrocyte hypertrophy. We recently found that naproxen (Npx), but not other nonsteroidal anti-inflammatory drugs, can induce collagen type X alpha 1 (COL10A1) gene expression in bone marrow-derived MSCs from healthy and OA donors. In this study, we determined the effect of Npx on COL10A1 expression and investigated the intracellular signaling pathways that mediate such effect in normal human MSCs during chondrogenesis. MSCs were cultured in standard chondrogenic differentiation media supplemented with or without Npx. Our results show that Npx can regulate chondrogenic differentiation by affecting the gene expression of both Indian hedgehog and parathyroid hormone/parathyroid hormone-related protein signaling pathways in a time-dependent manner, suggesting a complex interaction of different signaling pathways during the process.
Introduction
A
Since articular cartilage lacks a blood supply, its capacity to self-repair is extremely poor, and if left untreated, localized cartilage defects due to injury or focal degeneration typically lead to osteoarthritis (OA) of the entire joint. OA is characterized by progressive degeneration of articular cartilage, subchondral bone, and other joint tissues, resulting in a major reduction in the quality of life and substantial economic costs. Symptomatic OA is characterized by cartilage deterioration and osteophyte formation, affecting primarily the knees, hips, and hands, and the predominant symptom is pain.2,3 For this reason, patients are usually prescribed nonsteroidal anti-inflammatory drugs (NSAIDs), such as ibuprofen, diclofenac, naproxen (Npx), and celecoxib, as well as acetaminophen. 4 NSAIDs have been used because they can selectively inhibit cyclooxygenase-2 (COX-2), which is upregulated by interleukin-1 beta (IL-1β) and tumor necrosis factor-alpha (TNF-α). 5 One drawback is NSAIDs can alter certain fundamental processes involved in the normal healing of injured tissues. 6 We recently showed that Npx can affect gene expression during osteogenic differentiation of mesenchymal stem cells (MSCs), which are also called mesenchymal stromal cells, and downregulate mineral deposition in the extracellular matrix through Indian hedgehog (IHH) signaling, 7 suggesting that Npx could affect MSC-mediated repair of subchondral bone in OA patients. However, little is known about the effects of Npx on articular cartilage repair in OA patients.
In most cases, NSAIDs are not sufficient to delay the development of OA, and the chronic pain refractory to medication is the number one reason for joint replacement surgery. To delay the requirement for total joint replacement/arthroplasty as a result of OA, a number of conventional therapies such as microfracture and mosaicplasty are utilized. However, the long-term performance of these approaches is very poor. To reverse or retard the degeneration of articular cartilage and repair the subchondral bone, MSCs can be employed in the biological therapy of OA. 8 MSCs can be obtained from multiple tissues, including bone marrow, fat, and synovium. However, regardless of the source, MSCs have a propensity to express markers of hypertrophic chondrocytes, 9 an undesired characteristic in cartilage repair.10,11
Previous studies have shown that collagen type X, a marker of late-stage chondrocyte hypertrophy (associated with endochondral ossification), is constitutively expressed by MSCs isolated from OA patients, 9 and NSAIDs can affect the expression of both hypertrophic and osteogenic marker genes in MSCs,7,12,13 with Npx showing stronger effects than other drugs. The effect of these drugs on the transcriptome of MSCs with inherent effects on their characteristics and differentiation can influence the MSC fate during the repair of OA. Thus, it is important to understand whether Npx affects the chondrogenesis of MSCs.
IHH regulates both chondrogenesis and endochondral bone formation. 14 HH ligands are secreted from the cells and bind to the receptor Patched (PTC) to relinquish smoothened, frizzled class receptor (SMO) from PTC suppression, resulting in the activation of the zinc finger transcription factor GLI family, which are used as markers for HH signaling activity, and are responsible for HH-induced lineage commitment during MSC differentiation. 15 Parathyroid hormone (PTH) and parathyroid hormone-related protein (PTHrP), which share the same receptor, can affect the chondrogenesis of MSCs.16,17 Chondrocyte differentiation is controlled at multiple steps by PTH/PTHrP and IHH through the mutual regulation of their activities. 18 Ihh acts independently or dependently on PTH-related peptide in a negative feedback loop to regulate early chondrocyte differentiation and entry to hypertrophic differentiation.19,20
The purpose of this study was to investigate if Npx can interfere with the chondrogenesis of MSCs through IHH and PTH/PTHrP signaling pathways. We also determined the mechanisms underlying the effect of Npx on hypertrophic differentiation of human MSCs cultured in chondrogenic differentiation media.
Materials and Methods
Source and expansion of MSCs
Human MSCs from three donors between 20 and 30 years old were obtained from Lonza (Mississauga, ON, Canada). According to the supplier, MSCs were harvested from healthy bone marrow and were positive for CD105, CD166, CD29, and CD44, but negative for CD14, CD34, and CD45. All cells were expanded in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. The culture medium and the supplement were from Wisent, Inc. (St-Bruno, QC, Canada). The cells were used within four passages.
Cell culture
MSCs have been shown to undergo chondrogenesis in vitro using pellet cultures,21–23 which mimics the cellular condensation requirements for embryonic mesenchymal chondrogenesis and provides the physical and biochemical environmental factors conducive to cartilage formation. However, MSCs can also undergo chondrogenic differentiation in monolayer culture with a high density, when supplemented with a defined chondrogenic medium.24,25 In this experiment, the monolayer culture was chosen to study the function of Npx. In every well of the six-well plate (Sarstedt, Saint-Leonard, QC, Canada), 5×105 MSCs were plated and cultured in the expansion medium overnight. The floating cells were removed and the attached cells were cultured until the confluence was more than 90%. Then the cells were cultured in the chondrogenic differentiation medium for 3 days to allow the cells to adapt to the new environment. Afterward, the cells were cultured in 3 mL/well of the chondrogenic differentiation medium with 0.5 μM Npx (Sigma-Aldrich, Oakville, ON, Canada). Since the bone marrow is well vascularized, the concentration of Npx in our experiments was similar to the blood circulating levels in patients taking Npx. 26 The cells that were cultured without Npx were used as control cells. The chondrogenic differentiation medium was prepared with high-glucose DMEM containing 1% ITS+100 nM dexamethasone, 50 μg/mL ascorbic acid, 40 μg/mL proline, 10 ng/mL transforming growth factor beta 3 (TGFβ3), 100 units/mL penicillin, and 100 μg/mL streptomycin. The supplements for the chondrogenic differentiation medium were from Sigma-Aldrich, and penicillin and streptomycin were from Wisent, Inc.
Total RNA isolation
After MSCs were cultured for 3, 6, and 12 days, they were washed with phosphate-buffered saline (PBS) and total RNA was extracted using the TRIzol reagent (Invitrogen, Burlington, ON, Canada) as previously reported. Briefly, after centrifugation for 15 min at 12,000 g at 4°C, the aqueous phase was precipitated in one volume of isopropanol, incubated for 45 min at −20°C, and centrifuged again for 15 min at 12,000 g at 4°C. The resulting RNA pellet was washed with 75% ethanol and then centrifuged and air dried. Then, the pellets were suspended in 50 mL of diethylpyrocarbonate (DEPC)-treated water and assayed for RNA concentration by measuring A260 and purity by calculating A260/A280.
Reverse transcription and real-time PCR
First, 1 μg total RNA isolated from the cells was digested with DNase I (Invitrogen) according to the protocol of the supporter. Then, the purified RNA was reverse transcribed as described previously. Briefly, 1 μg RNA was mixed with random primers (final concentration 0.15 μg/μL), dNTP mixture (final concentration 0.5 mM), and DEPC-treated distilled water with a total volume of 12 μL. After the solution was incubated at 65°C for 5 min, it was mixed with a first-strand buffer, Dithiothreitol, RNaseOUT, and SuperScript II Reverse Transcriptase with a final volume of 20 μL. Then, the solution was incubated at 45°C for 50 min to perform the reverse transcription and then in 70°C for 15 min to inactivate the reverse transcriptase. For LightCycler real-time polymerase chain reaction (PCR), a master mix of the following reaction components was prepared with the final concentrations as 10 μL SYBER PCR master mix (Qiagen, Mississauga, ON, Canada), 8 μL distilled water, 0.5 μL forward primer (final concentration 0.25 μM), and 0.5 μL reverse primer (final concentration 0.25 μM). To each 19 μL master mix, 1 μL of cDNA was mixed as a PCR template. The sequences of primers are in Table 1. The reaction conditions included one cycle of PCR initial activation step (95°C for 15 min, 20°C/s ramp rate), 45 cycles of amplification and quantification (94°C for 15 s, 57°C for 30 s, 72°C for 30 s), one cycle of melting curve (65–95°C with heating rate of 0.1°C/s with a continuous fluorescence measurement), and a final cooling step to 4°C. The crossing points (CPs) were determined by the LightCycler software 3.3 (Roche Diagnostics, Indianapolis, IN) and were measured at constant fluorescence level. The ratio of gene expression relative to glyceraldehyde 3 phosphate dehydrogenase (GAPDH) as the reference gene was determined by the following equation:
ACAN, aggrecan; ADAMTS, a disintegrin and metalloproteinase with thrombospondin motifs; ALP, alkaline phosphatase; COL2A1, collagen type II alpha 1; COL10A1, collagen type X alpha 1; COMP, cartilage oligomeric matrix protein; GAPDH, glyceraldehyde 3 phosphate dehydrogenase; IHH, Indian hedgehog; MMP, matrix metalloproteinase; PTC1, Patched 1; PTHR1, PTH/PTHrP receptor; SOX9, Sry-related high-mobility-group box 9; TIMP, tissue inhibitors of metalloproteinase.
Alkaline phosphatase activity analysis
After MSCs were cultured in the chondrogenic differentiation medium with or without Npx for 6, 9, and 12 days, the cells were lysed and alkaline phosphatase (ALP) activity was assayed with the StemTAG™ Alkaline Phosphatase Activity Assay Kit (Colorimetric; Cell Biolabs, Inc., San Diego, CA) according to the protocol from the supplier. Briefly, the cells were washed with cold PBS and then lysed with the lysis buffer. After incubating at 4°C for 10 min, the lysis solution was centrifuged at 4°C at 12,000 g for 10 min. The supernatant was collected as cell lysate and the protein concentration was measured with the BCA assay (Thermo Scientific, Burlington, ON, Canada). Then 50 μL of cell lysate was added to a 96-well plate, while the well containing 50 μL cell lysis buffer was applied as a blank control. The reaction was initiated by adding 50 μL of the StemTAG AP Activity Assay Substrate, and the plate was incubated at 37°C for 15 min. Then the reaction was stopped by adding 50 μL of 1× stop solution and mixed by placing the plate on an orbital plate shaker for 30 s. Finally the absorbance value of each well was read at 405 nm. To analyze pNP concentration produced by the reaction, a serial twofold dilution of pNP standard was prepared in 1× stop solution, and the absorbance of each dilution was measured at 405 nm. Then pNP concentration in the reaction buffer was calculated by comparing with the standard curve. Alp activity was calculated with the formula below:
where 30 min is the reaction time.
GAG and DNA analysis
During the chondrogenesis of MSCs, the culture media were changed every 3 days. The media were collected and stored in −20°C for sulfated GAGs analysis. GAGs were quantified in culture media by a modified dimethylmethylene blue dye-binding assay.27,28 Samples were diluted to fall within the middle of the linear range of the standard curve. To compare the number of cells at different time, DNA was measured on day 3 and 21 by using a Quant-iT dsDNA High-Sensitivity Assay Kit (Invitrogen), following the manufacturer's instructions. 25
Statistical analysis
Statistical analysis was performed using analysis of variance, followed by Fisher's protected least significant difference post hoc test using StatView (SAS Institute, Inc., Cary, NC). The results of three experiments with MSCs from three different donors were assessed on 80% power to detect a difference in gene expression. Results are presented as the mean±standard deviation of three experiments. Differences were considered statistically significant with p<0.05.
Results
Aggrecan (ACAN), collagen type II alpha 1 (COL2A1), and cartilage oligomeric matrix protein (COMP) are important genes that define the chondrocyte phenotype.29,30 SOX9 is a transcription factor belonging to the Sry-related high-mobility-group box (Sox) proteins family, essential for chondrogenesis of MSCs.31,32 The effect of Npx on the expression of these genes was therefore assessed after culturing MSCs in chondrogenic differentiation media for 3, 6, and 12 days. At day 3, the expression of ACAN (p=0.009), COL2A1 (p=0.012), and SOX9 (p=0.005) was increased significantly, while no significant effect was found for COMP, when compared to controls (Fig. 1). In contrast, when MSCs were cultured with Npx for 6 and 12 days, the expressions of ACAN (p=0.005 on day 6, p<0.001 on day 12), COL2A1 (p=0.003 on day 6, p=0.012 on day 12), COMP (p<0.001 on day 6, p=0.007 on day 12), and SOX9 (p=0.007 on day 6, p=0.032 on day 12) decreased significantly when compared to controls. This indicates that at early time points Npx has the potential to stimulate chondrogenic differentiation, but with a longer period, Npx can suppress the chondrogenesis of MSCs.

The effect of naproxen (Npx) on the expression of aggrecan (ACAN)
Collagen type X alpha 1 (COL10A1) is a marker gene for hypertrophic chondrocyte differentiation, a process that is undesirable for effective cartilage repair.33,34 When MSCs were cultured in a chondrogenic differentiation medium with Npx, the expression of COL10A1 was significantly upregulated on day 3 (p=0.009), but was significantly downregulated on day 6 (p=0.006), while no significant difference was observed on day 12 (Fig. 1E). Thus, Npx appears to decrease hypertrophic differentiation when MSCs were cultured for 6 days, while stimulating hypertrophy in the short term. Alkaline phosphatase (ALP) plays an active role in initiating the calcification process after chondrocytes have become hypertrophic. 35 When MSCs were cultured in a chondrogenic differentiation medium with Npx, the expression of ALP was significantly upregulated on day 3 (p=0.006), but was significantly decreased on day 12 (p=0.011), while no significant difference was observed on day 6 (Fig. 1F).
Proteoglycan production during chondrogenic differentiation can also be monitored with GAG analysis, and this was used to assess the effect of Npx on MSC differentiation. After MSCs were cultured in a chondrogenic differentiation medium with Npx for 21 days, the quantity of GAG secreted into the culture medium was significantly lower than that in the control medium (Fig. 2). Since the DNA content of the cultures were not significantly different at day 3 and 21, the decreased GAG synthesis is likely the result of decreased production by each cell rather than a consequence of less cells due to cell death.
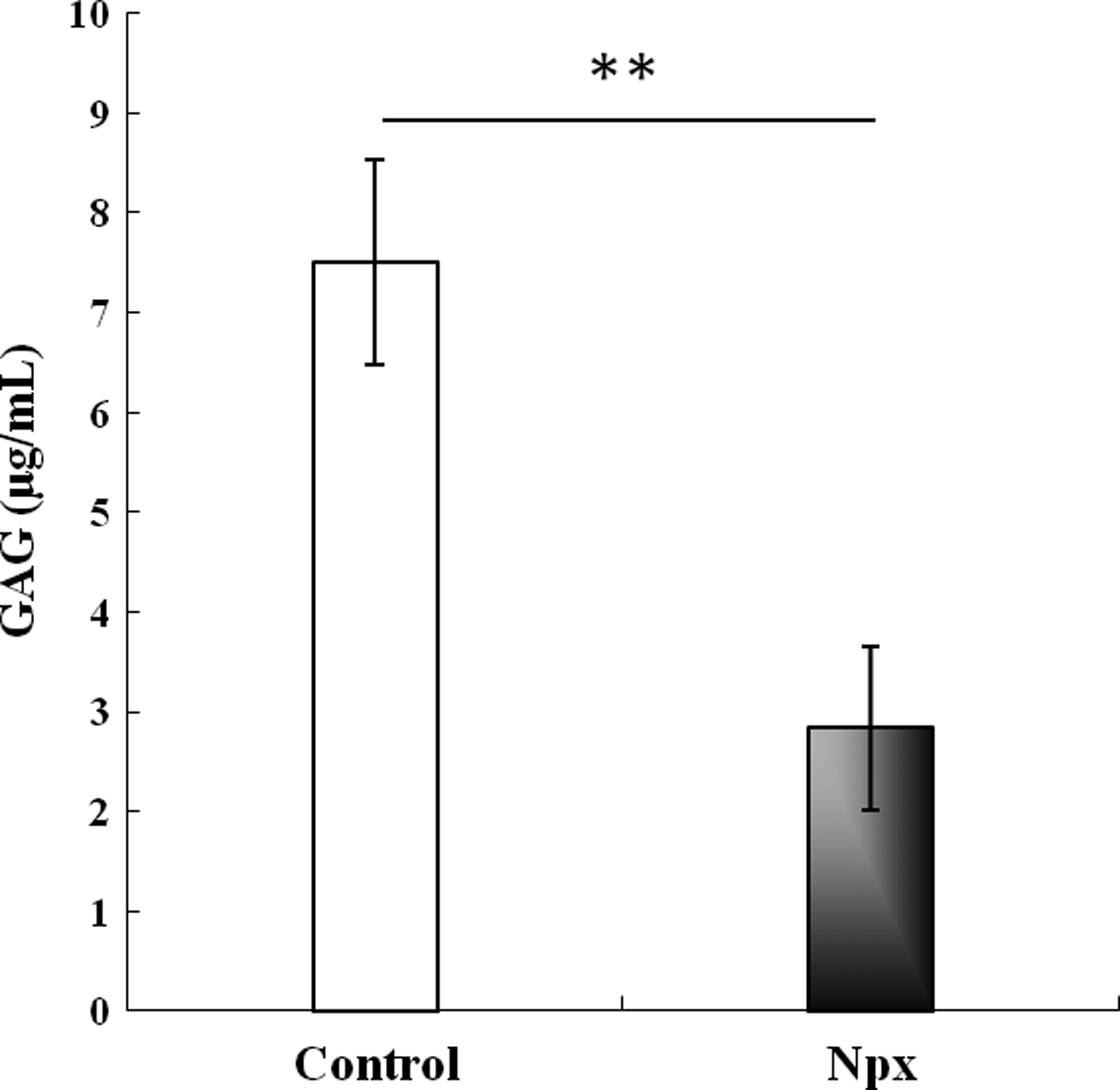
Glycosaminoglycan (GAG) concentration in pooled culture media of MSCs cultured for 21 days in chondrogenic differentiation medium without (control) or with 0.5 μM Npx. The results are shown as mean±SD of three independent experiments with MSCs from three different donors. **p<0.01 versus control.
During the chondrogenic differentiation, ALP can also be monitored by its activity and this was used to assess the effect of Npx on MSC differentiation. Even though the mRNA level could be detected on day 3 through real-time PCR, the enzyme activity was too weak to be detected. So we determined the activities of ALP on days 6, 9, and 12. After MSCs were cultured in a chondrogenic differentiation medium with Npx for 9 and 12 days, ALP activity was significantly lower than that in the control medium, although no significant effect was observed on day 6 (Fig. 3).

ALP activity in MSCs cultured in chondrogenic differentiation medium without (control) or with 0.5 μM Npx for different periods. The results are shown as mean±SD of three independent experiments with MSCs from three different donors. *p<0.05 versus control.
Matrix metalloproteinases (MMPs) are essential for chondrogenic differentiation of adult human MSCs. 36 However, while the function of ADAMTS (a disintegrin and metalloproteinase with thrombospondin type I motifs) family does not appear to be required for cartilage development in mice, 37 its role in chondrocyte homeostasis is vital. 38 The effect of Npx on the expression of these genes was therefore assessed.
After the cells were cultured for 6 days in the chondrogenic differentiation medium, MMP3 expression with Npx increased significantly compared to controls (p=0.006) (Fig. 4A). However, no significant effect was observed compared with control cells at both day 3 and 12 (Fig. 4A). In contrast, the expression of MMP13 was increased significantly on day 3 (p=0.007), but was decreased significantly on day 6 (p<0.001) and day 12 (p<0.001) (Fig. 4B). Compared with control cells, there was no significant effect of Npx on the expression of ADAMTS4 (Fig. 4C), while the expression of ADAMTS5 was increased significantly on day 3 (p=0.012) and day 12 (p<0.001) (Fig. 4D). Thus Npx appeared to affect MMP3, MMP13, and ADAMTS5 expressions during the chondrogenic differentiation of MSCs. Striking changes in the expression of matrix remodeling enzymes in control cultures over time was observed, which have previously been observed in MSCs during chondrogenic differentiation. 25

The effects of Npx on the expression of matrix metalloproteinase (MMP)3
We next examined the expression of MMPs endogenous inhibitors, the tissue inhibitors of metalloproteinases (TIMPs). MSCs have been shown to secrete high levels of TIMPs. 39 After culturing the cells for 12 days in the absence of Npx, MSCs show TIMP1, TIMP2, and TIMP3 gene expression, with the highest expression at day 6 (Fig. 4E–G). With Npx supplementation, the expression of TIMP1 was increased significantly on day 3 compared to controls (p=0.012). This effect was not observed at day 6 and 12 (Fig. 4E). Npx supplementation increased the expression of TIMP2 significantly on day 3 (p=0.009) and day 12 (p=0.011) (Fig. 4F), but not on day 6. After the cells were cultured for 3 days in the chondrogenic differentiation medium with Npx, TIMP3 expression significantly increased compared to controls (p=0.003), and the increase was also observed after 6 days (p=0.001) and 12 days (p<0.001) (Fig. 4G). Thus Npx appeared to stimulate the expression of TIMPs during chondrogenic differentiation of MSCs.
We have previously shown that Npx affected the expression of IHH, PTC1, GLI1, and GLI2 genes belonging to the Indian hedgehog signaling pathway when MSCs were cultured in the osteogenic differentiation medium. 7 The effect of Npx on the expression of PTC1, IHH, GLI1, GLI2, and GLI3 when MSCs were cultured in chondrogenic differentiation medium was therefore assessed (Fig. 5). When the cells were cultured in this medium with Npx, the expression of PTC1 significantly decreased on day 6, compared with that in control cells (Fig. 5A). This difference was not observed on day 3 and 12. The expression of IHH significantly increased on days 3 and 6, compared with that in control cells, but its expression significantly decreased on day 12 compared with that in control cells (Fig. 5B). Npx supplementation significantly increased the expression of GLI1 on day 3 (p=0.001), but its expression was significantly decreased on day 6 (p=0.024) (Fig. 5C). There was a tendency toward a decrease of GLI1 expression on day 12 (p=0.08). The expression of GLI2 significantly increased on day 12 (p=0.031), while no significant difference was observed on days 3 and 6 (Fig. 5D) when cells were supplemented with Npx. Finally, the expression of GLI3 significantly increased on day 3 (p=0.017) and day 12 (p=0.044), while no significant effect of Npx on the expression of GLI3 was observed on day 6 (Fig. 5E).
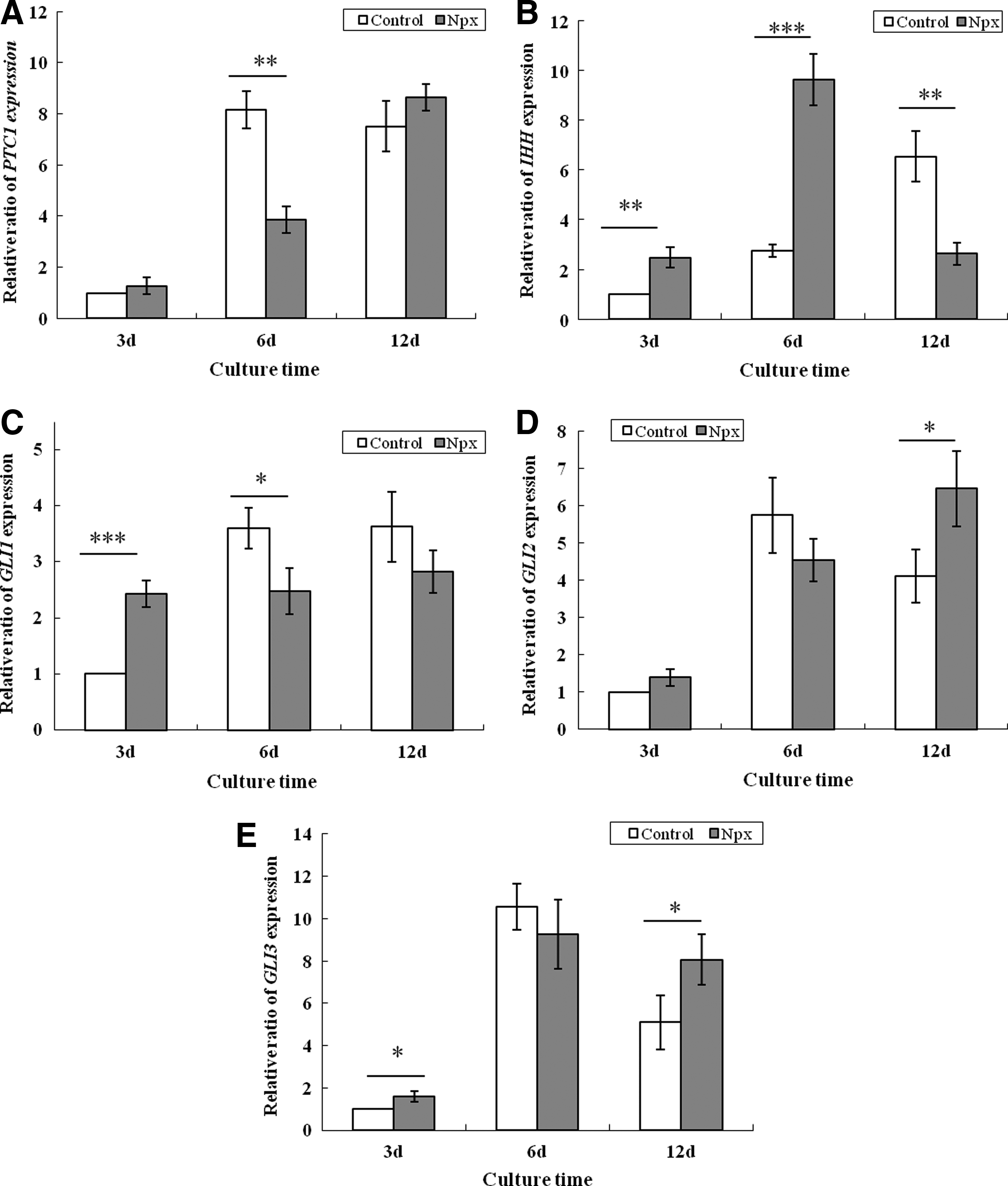
The effects of Npx on the expression of genes Patched 1 (PTC1)
To study whether Npx can affect PTH/PTHrP signaling, the effects of Npx on the expression of PTH and PTH/PTHrP receptor (PTHR1) were studied. Npx increased the expression of PTH significantly on day 3 (p=0.04) and day 12 (p<0.001), but its expression was significantly decreased on day 6 compared with that in control cells (p=0.035) (Fig. 6A). The expression of PTHR1 was significantly increased on day 3 (p=0.045), but significantly decreased on day 6 (p=0.003) (Fig. 6B). No significant effect of Npx on the expression of PTHR1 was observed on day 12. Thus Npx not only suppresses the expression of PTH, but also the expression of PTH/PTHrP receptor PTHR1.

The effects of Npx on the expression of parathyroid hormone (PTH)
Discussion
In the present study, we used Npx as an NSAID supplement and determined that it can interfere with chondrogenesis of MSCs. In a clinical context this indicates that NSAIDs present in the synovial fluid may have an influence on the ability of endogenous and exogenous MSCs to differentiate and stimulate cartilage repair.
Previous work has demonstrated a potential of Npx to interfere with MSC differentiation.7,11,12 Studies utilizing MSCs isolated from the bone marrow of patients with OA or normal donors cultured with NSAIDs showed that Npx could induce the expression of COL10A1, ACAN, COL1A1, as well as ALP, bone sialoprotein (BSP), osteocalcin (OC), and Runt-related transcription factor 2 (RUNX2).11,12 In addition, these results indicated that Npx can affect gene expression during osteogenic differentiation of MSCs through IHH signaling. 7 Elevated basal COL10A1 expression in OA MSCs also involved the activation of MAPK pathway and stimulation of the 5-lipoxygenase signaling pathway by Npx. 13 The present study is the first to demonstrate that Npx can regulate chondrogenic differentiation by affecting both the IHH and PTH/PTHrP signaling pathways. Further studies are underway to determine if the gene expression observed with IHH is translated to protein. This is important to determine the role of Npx in IHH signaling pathways related to the process of chondrogenesis and terminal chondrocyte differentiation.
Culturing MSCs in chondrogenic differentiation media for 3, 6, and 12 days in the presence of Npx used in the present study mimic short-term, intermediate, and long-term intake of Npx by patients with OA, where it can be present in the synovial fluid and influence the ability of MSCs to differentiate and stimulate cartilage repair. For long-term use, Npx is preferred to other NSAIDs in patients because it is associated with less vascular risk than other NSAIDs. 40 Previous studies, in line with the findings of this study, showed that NSAIDs have the potential to suppress chondrogenic differentiation by MSCs. 41 We showed that Npx was also capable of suppressing osteogenic differentiation. 7 However, despite its extensive use as a prescription and over-the-counter medication, it still needs to be established at what dosage and duration it affects the repair of skeletal tissues.
The effect of Npx on the expression of genes in MSCs was time dependent, suggesting a complex interaction of different proteins involved in IHH signaling. Additionally, the independent functions of IHH and PTH/PTHrP signaling,19,20,42,43 and the interaction between IHH and PTH/PTHrP signaling on the chondrogenesis of MSCs, can also lead to the complex results of gene expression.17,18 In skeletogenesis, IHH regulates chondrogenesis during endochondral ossification, while Gli zinc finger proteins regulate IHH signaling. 44 Previously, we showed that Npx did not affect the expression of GLI3 when MSCs were cultured in osteogenic differentiation media. 7 In the present study, the expression of GLI3 increased significantly on days 3 and 12. The difference may be caused by the different culture media.
In this study, for most of the genes, only mRNA but not protein expression levels were studied. Although the fluctuation of mRNA expression level usually is parallel with protein level, the effects of Npx on protein level are on the way for further confirmation. Furthermore, IHH gene expression was not consistent, increasing on day 6 and decreasing at day 12 in the presence of NPx. Although decreasing levels of IHH may be explained by the antihypertrophic effects of NPx during long-term treatment, it is possible that the variability in the cells' response to NPx could be due to differences in the cell sources. According to the supplier, these cells were harvested from healthy donors aged between 20 and 30 years and were positive for CD105, CD166, CD29, and CD44, but negative for CD14, CD34, and CD45. Altogether, the present data indicate that Npx can directly interfere with MSC chondrogenesis potentially through the IHH and PTH/PTHrP signaling pathways. Other studies showed that Npx could alter MAPK signaling, in particular p38-MAPK, in the induced expression of COL10A1 in human MSCs. 12 Furthermore, Npx appears to be capable of affecting chondrogenesis and hypertrophic differentiation of MSCs depending on different culture time.
Our studies and those of others raise the question whether bone marrow, adipose, and synovium are the best sources of MSCs for the repair of permanent cartilages such as articular cartilage, since MSCs derived from these tissues consistently undergo hypertrophic differentiation during in vitro chondrogenic differentiation. 8 An alternative source would be articular cartilage stem cells (ACSCs), which do not differentiate down the endochondral pathway through hypertrophy in vitro in chondrogenic assays.45,46 Although some methods, such as polymer substrates surface modified by glow discharge plasma,11,47–49 can result in a decrease of hypertrophic markers in differentiated cells, they do not reach the levels found in stable (permanent) chondrocytes. 50 Conversely, ACSCs differentiate into chondrocytes that do not have the hypertrophic phenotype. However, it remains to be determined if Npx supplementation can influence ACSC hypertrophic differentiation.
Conclusions
Npx interferes with chondrogenesis of MSCs. At 12 days of culture, Npx negatively affects matrix production by the MSCs in the chondrogenic lineage and it seems to suppress termination of hypertrophic differentiation. These effects may be mediated by the interplay of PTH and IHH signaling pathways. Therefore, in principle, Npx supplementation could interfere with the expected outcome of medical treatment of degenerative lesions in articular cartilage.
Footnotes
Acknowledgments
This research was supported by grants from the Canadian Institute of Health Research (CIHR). M.T. and B.M. were supported by the Dutch Arthritis Foundation (LLP22).
Disclosure statement
No competing financial interests exist.