
Abstract
Introduction:
Current tissue engineering strategies to heal critical-size bone defects through direct bone formation are limited by incomplete integration of grafts with host bone and incomplete graft vascularization. An alternative strategy for bone regeneration is the use of cartilage grafts that form bone through endochondral ossification. Endochondral cartilages stimulate angiogenesis and are remodeled into bone, but are found in very small quantities in growth plates and healing fractures. We sought to develop engineered endochondral cartilage grafts using osteoarthritic (OA) articular chondrocytes as a cell source. Such chondrocytes often undergo hypertrophy, which is a characteristic of endochondral cartilages.
Materials and Methods:
We compared the ability of unmodified human OA (hOA) cartilage and cartilage grafts formed in vitro from hOA chondrocytes to undergo endochondral ossification in mice. Scaffold-free engineered chondrocyte grafts were generated by pelleting chondrocytes, followed by culture with transforming growth factor-β1 (TGF-β1) and bone morphogenetic protein 4. Samples derived from either primary or passaged chondrocytes were implanted subcutaneously into immunocompromised mice. Grafts derived from passaged chondrocytes from three patients were implanted into critical-size tibial defects in mice. Bone formation was assessed with histology after 4 weeks of implantation. The composition of tibial repair tissue was quantified with histomorphometry.
Results:
Engineered cartilage grafts generated from passaged OA chondrocytes underwent endochondral ossification after implantation either subcutaneously or in bone. Cartilage grafts integrated with host bone at 15 out of 16 junctions. Grafts variably remodeled into woven bone, with the proportion of bony repair tissue in tibial defects ranging from 22% to 85% (average 48%). Bony repair tissue bridged the tibial defects in half of the animals. In contrast, unmodified OA cartilage and engineered grafts formed from primary chondrocytes did not undergo endochondral ossification in vivo.
Conclusions:
hOA chondrocytes can adopt an endochondral phenotype after passaging and TGF-β superfamily treatment. Engineered endochondral cartilage grafts can integrate with host bone, undergo ossification, and heal critical-size long-bone defects in a mouse model. However, additional methods to further enhance ossification of these grafts are required before the clinical translation of this approach.
Introduction
B
In the absence of rigid internal fixation, fractures typically heal through a cartilage intermediate that is subsequently remodeled into bone. This process shares many characteristics with endochondral ossification that occurs during development. The use of cartilage to stimulate endochondral bone formation may be a viable alternative to the use of traditional bone grafts.2–5 In a mouse model of segmental bone defects, allogeneic cartilage grafts from fracture callus led to better repair than allogeneic structural bone allografts. 5 One obstacle to translating endochondral grafts to clinical use is the development of an appropriate source of hypertrophic cartilage. 6
We hypothesized that articular chondrocytes could be modified to assume an endochondral phenotype capable of generating bone. Articular cartilage is normally a permanent tissue that resists hypertrophy, vascularization, and ossification. It is characterized by high expression of type II collagen and aggrecan and behaves differently than the transient endochondral cartilages found in growth plates and fracture callus. The latter cells undergo hypertrophy, which involves cellular enlargement and expression of multiple proteins, including type X collagen, matrix metalloproteinase 13 (MMP-13), and vascular endothelial growth factor (VEGF). Differential expression of antiangiogenic molecules such as chondromodulin 1 (Chm-1) and thrombospondin 1 (TSP-1) likely contribute to the phenotypic differences between permanent and transient cartilages. These factors stabilize the phenotype of articular cartilage and inhibit terminal differentiation. Chm-1 promotes chondrogenesis, while inhibiting chondrocyte hypertrophy, vascular endothelial proliferation, and cartilage ossification.7,8 Similarly, TSP-1 inhibits bone morphogenetic protein 7 (BMP7)-induced endochondral ossification of cartilaginous repair tissue formed after microfracture. 9
We were particularly interested in the ability of osteoarthritic (OA) articular cartilage to promote endochondral bone regeneration. OA articular chondrocytes can undergo phenotypic dysregulation, becoming hypertrophic and abnormally expressing genes such as COL10 and MMP-13. 10 Since over 600,000 total knee replacements are performed each year in the United States alone, 11 OA cartilage from total joint replacement resections represents an untapped potential source of cells for tissue regeneration.
The goal of this project was to determine if human OA (hOA) articular chondrocytes could be used to generate endochondral cartilage grafts capable of supporting bone formation. The primary endpoint was histologic evaluation of bone formation in a mouse model of critical-size long-bone defects.
Materials and Methods
Isolation and expansion of articular chondrocytes
The hOA articular cartilage was obtained from the femoral resections of men over the age of 60 undergoing total knee replacement for osteoarthritis. The use of human tissues was reviewed by our Institutional Committee on Human Research and judged to not qualify as human subject research since the specimens would not have identifiers and the research would be done in aggregate. Articular chondrocytes were isolated by digesting cartilage in DMEM/F-12 containing 3% fetal bovine serum (FBS) and 0.2% collagenase P (Roche, Indianapolis, IN) for 8 to 12 h at 37°C. Digested cartilage was passed through 70-μM filters. Cells were collected by centrifugation for 5 min at 500 g and washed thrice with DMEM/F-12 containing 3% FBS. Primary chondrocytes were then pelleted without expansion of cell numbers in tissue culture. Alternatively, chondrocytes were expanded in monolayer culture to passage 5 or 6 (∼240- to 730-fold expansion of cell numbers) using MesenGro® MSC media (StemRD, Burlingame, CA) containing Gibco® GlutaMAX™ supplement (Life Technologies, Grand Island, NY). These media contain a proprietary mix of growth factors. Following monolayer expansion, the passaged chondrocytes were trypsinized and pelleted.
Scaffold-free cartilage grafts
Scaffold-free tissues were formed by centrifuging 250,000 to 500,000 cells for 5 min at 500 g. Either primary chondrocytes or chondrocytes that had been expanded in monolayer culture with MesenGro MSC media were used. The resultant cell pellets were cultured for 1 week in insulin-transferrin-selenium (ITS) media (DMEM/F-12, ITS premix [#354352; Becton Dickenson, Franklin Lakes, NJ], 50 μg/mL ascorbic acid, 0.1% bovine serum albumin [BSA], 100 μg/mL sodium pyruvate, and 25 mM HEPES) containing (1) no additional supplement, (2) transforming growth factor β1 (TGF-β1; 10 ng/mL), or (3) TGF-β1 (10 ng/mL) and BMP4 (200 ng/mL). Pellets were then cultured for at least an additional 3 weeks in ITS media without growth factors. Pellets were then harvested for Alcian green staining for glycosaminoglycans 12 and quantitative reverse transcription polymerase chain reaction (qRT-PCR) for genes associated with chondrogenesis and endochondral ossification. Pellets grown with TGF-β1 and BMP4 were implanted subcutaneously or into tibial defects in mice. Implanted grafts were harvested after 4 weeks in vivo.
mRNA isolation and qRT-PCR
mRNA isolation qRT-PCR were performed using the TaqMan® Gene Expression Cells-to-CT™ kit (Life Technologies, Grand Island, NY) and TaqMan Gene Expression assays according to the manufacturer's instructions. Assays included aggrecan (Hs00153936_m1), β2-microglobulin (Hs99999907_m1), COL1A2 (Hs01028956_m1), COL2A1 (Hs00156568_m1), COL10A1 (Hs00166657_m1), MMP-13 (Hs00233992_m1), VEGF (Hs0090005_m1), Chm-1 (Hs00993254_m1), and TSP-1 (Hs00962908_m1). Relative gene expression was calculated by normalizing to the reference gene β2-microglobulin as 2−ΔCT. Mean ± standard deviation for three to five replicates from each condition was calculated. Statistical difference was evaluated using the Mann–Whitney test when evaluating gene expression of cells grown in monolayer. For evaluation of pellet-culture gene expression, analysis of variance, followed by Tukey's honestly significant difference test comparisons of all pairs was performed.
Murine subcutaneous model
All murine studies were approved by our Institutional Animal Care and Use Committee. Grafts were implanted subcutaneously on the dorsal aspect of adult (6–9 week old), male, immunocompromised hairless mice (Homozygous Nu/J, Strain #2019; Jackson River Laboratories, Sacramento, CA). Implanted grafts included primary and passaged hOA chondrocyte pellets that were first cultured in vitro with TGF-β1 and BMP4. Pellets were derived from 300,000 cells and were generated under carrier-free conditions. Skin incisions were made 1 cm lateral to the spine. Subcutaneous pockets were created by blunt dissection and grafts implanted away from the incision site. Skin was closed with 6.0 silk suture, antibiotic ointment was applied, and postoperative analgesics were administered per protocol. Animals were allowed to heal for 4 weeks.
Murine segmental defect model
Segmental 3 mm defects were created unilaterally at the tibial middiaphysis of immunocompromised mice under microscope visualization. Each tibia was externally stabilized with a customized circular fixator consisting of two 2 cm rings held concentrically by three threaded rods. 13 The fixator stabilizes and prevents overlap of the fracture ends. Defects were filled with unmodified hOA cartilage grafts (3 mm length × 1 mm width). 8.0 suture was used to close soft tissues and secure the grafts in place. Animals were monitored for postoperative pain, analgesics were provided, and free ambulation was allowed. Alternatively, defects were filled with engineered cartilage grafts formed from hOA articular chondrocytes that were passaged in monolayer with proprietary serum-free media and then were cultured in cell pellets in ITS media with exposure to TGF-β1 and BMP4. Control animals received no grafts in the tibial defects. Animals were allowed to heal for 4 weeks.
Histology
Mice were euthanized and tibiae were collected and fixed in 4% paraformaldehyde (pH 7.2–7.4) for 24 h at 4°C. Specimens were decalcified in 19% EDTA (pH 7.4) for 14 days at 4°C and then processed for paraffin histology. Engineered cartilage grafts after in vitro culture or subcutaneous implantation were similarly fixed with or without decalcification for 3 days in 19% EDTA (pH 7.4) at 4°C before embedding and sectioning. Standard histology protocols were used to visualize the bone and cartilage: Masson's Trichrome (bone = blue or red; cartilage = light blue; American Mastertech, Lodi, CA), safranin-O/fast green (glycosaminoglycans = red), and Von Kossa (mineral = black, #KTVKO; American Mastertech). Immunofluorescence was used to detect collagen II (1:200, II-II6B3; Developmental Studies Hybridoma Bank, Iowa City, IA) and collagen X (1:200; Gary Gibson, Henry Ford Hospital, Detroit, MI) as previously described. 5 The primary antibody or IgG control diluted in 1% BSA was incubated on slides overnight at 4°C. Alexa Fluor 594 goat anti-mouse (#150116; Abcam, Cambridge, MA) was used to detect the primary antibodies. Immunohistochemistry for human osteocalcin was performed with a human-specific antibody (ab13420; Abcam) as previously described. 14 Histomorphometric analysis to quantify the amount of bone in tibial repair tissue was performed as previously described. 5
Results
Intact articular cartilage retains native phenotype in segmental bone defects
While healthy articular cartilage is resistant to endochondral ossification, osteoarthritis is associated with aberrant gene and protein expression that alter phenotype and promote hypertrophy. 10 We examined whether hOA articular cartilage grafts could promote endochondral bone formation in a critical-size murine tibial defect. Tibiae were stabilized with external fixators and defects filled with size-matched native hOA articular cartilage grafts. Masson's trichrome staining of unimplanted tissue showed initial tissue histology (Fig. 1). After 4 weeks of implantation into a tibial defect, hOA grafts maintained an articular phenotype with no evidence of remodeling, bone formation, integration with host bone, or vascularization.
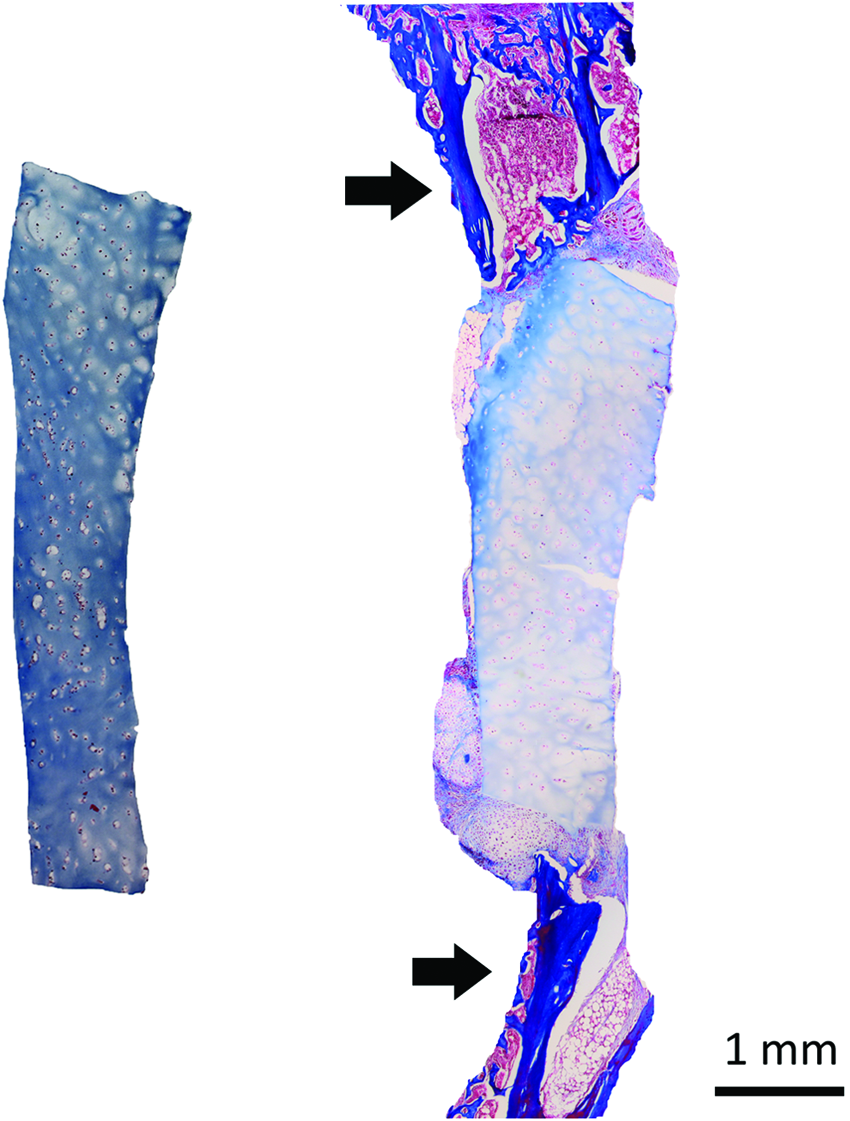
Unmodified human osteoarthritic (hOA) articular cartilage does not ossify in a mouse segmental tibial defect model. Left panel: Masson's trichrome staining of unmodified hOA articular cartilage. Right panel: Masson's trichrome staining of an unmodified hOA articular cartilage graft after 4 weeks of implantation in a segmental tibial defect. There has been no remodeling of the graft. No ossification of the graft has occurred and there is minimal new periosteal bone formation from the tibia. Instead, cartilaginous tissues have formed close to the implanted hOA cartilage. Arrows indicate the proximal (top) and distal (bottom) ends of the tibia. Color images available online at www.liebertpub.com/tea
Engineered endochondral cartilage grafts
Since unmodified hOA cartilage did not undergo endochondral ossification, we next evaluated whether hOA articular chondrocytes could be engineered into bone-forming tissues. To increase cell numbers, we first expanded articular chondrocytes in monolayer culture to passage 6 using commercially obtained serum-free media (MesenGro MSC media), which contain a proprietary mix of growth factors. Gene expression studies indicated that passaging led to loss of the chondrocyte phenotype (Fig. 2). In addition, passaging did not lead to upregulation of genes associated with endochondral ossification. Compared to primary cells (P0), articular cartilage-associated genes such as COL2 and aggrecan (ACAN) were downregulated by passaging, while COL1 was upregulated. Markers of hypertrophy such as VEGF and MMP-13 were also significantly downregulated during passaging, while COL10 was not detectable in these samples.

Passaging of hOA articular chondrocytes in monolayer under serum-free conditions decreases chondrogenic gene expression and does not promote hypertrophy. Quantitative RT-PCR
Since chondrogenesis occurs in three-dimensional conditions, we next transferred sixth-passage chondrocytes to pellet culture in ITS media. The following three conditions were evaluated in pellet culture: (1) no additional supplement (control), (2) TGF-β1, and (3) TGF-β1 and BMP4. As assessed by qRT-PCR, COLI expression was similar for all three conditions (Fig. 3). In contrast, only the combination of TGF-β1 and BMP4 led to upregulation of molecules associated with chondrogenesis and endochondral ossification, including COL2, COL10, ACAN, MMP-13, and VEGF. The combination of factors also led to an increased expression of TSP-1 and Chm-1. Similarly, only samples treated with TGF-β1 and BMP4 showed accumulation of glycosaminoglycans as assessed by Alcian green staining (data not shown).

Effects of growth factor treatment during pellet culture on gene expression of engineered cartilage grafts derived from passaged hOA articular chondrocytes. Quantitative RT-PCR of sixth-passage hOA articular chondrocyte pellets that were exposed to (1) no additional differentiation factors (controls), (2) 10 ng/mL TGF-β1, or (3) 10 ng/mL TGF-β1 and 200 ng/mL BMP4 for 1 week, followed by three additional weeks of culture in ITS media. Gene expression relative to the reference gene β2-microglobulin is shown. Means and standard deviations are shown. *Significantly more expression (p < 0.05) than the two other groups. BMP, bone morphogenetic protein; ITS, insulin-transferrin-selenium; TGF-β1, transforming growth factor-β1.
We then compared pellet cultures generated from either primary articular chondrocytes or chondrocytes that had been first passaged in monolayer with serum-free media. For both cell types, pellet cultures were grown in ITS media with TGF-β1 and BMP4. After 4 weeks of in vitro culture, pellets generated from primary cells were friable and showed minimal safranin-O, type II collagen, or type X collagen staining (Fig. 4A–C). The cells did not exhibit hypertrophic morphology. In contrast, pellets formed from passaged cells demonstrated intense safranin-O, type II collagen, and X collagen staining (Fig. 4D–F).
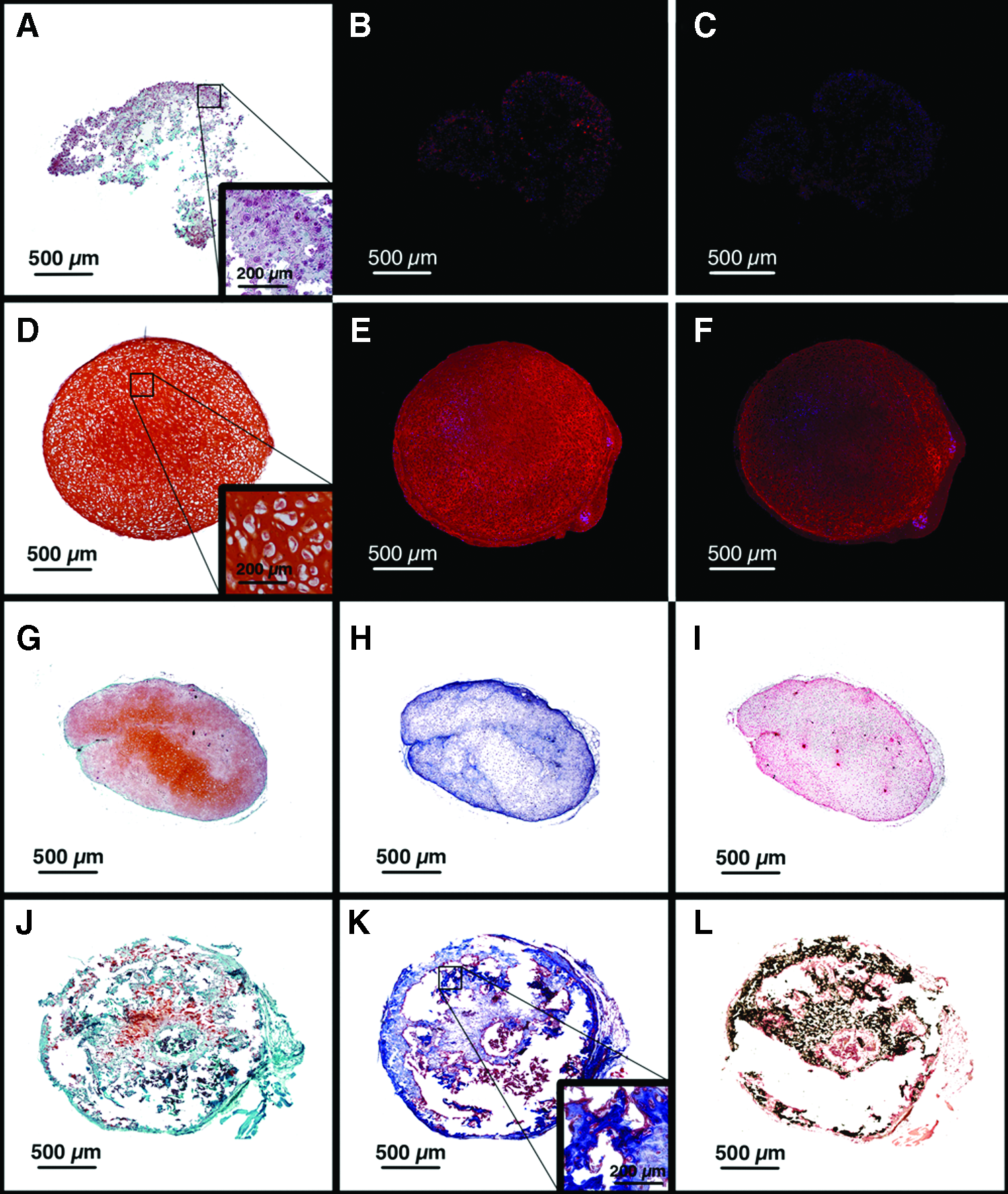
Endochondral ossification of passaged, but not primary hOA articular chondrocyte grafts. Primary
Pellets that had been grown in vitro for 4 weeks were then implanted for an additional 4 weeks in subcutaneous pockets in mice. The primary pellets generated a cartilaginous matrix, but did not undergo mineralization, remodeling, or endochondral ossification (Fig. 4G–I; three pellets derived from cells from a single patient were evaluated). In contrast, pellets from passaged chondrocytes lost glycosaminoglycan staining, underwent vascular remodeling, mineralized, and showed areas of woven bone formation (Fig. 4J–L; three pellets derived from cells from a single patient were evaluated).
We next tested the ability of engineered cartilage grafts derived from passaged hOA articular chondrocytes to stimulate endochondral bone repair in the murine tibial-defect model (Fig. 5). Chondrocytes were passaged in serum-free media and then pelleted with TGF-β1 and BMP4 stimulation. Grafts derived from cells from three patients were implanted unilaterally into eight animals (Table 1). After 4 weeks of implantation, integration between the grafts and the host bone occurred at 15 out of 16 junctions. As evaluated with histology, there was continuity between the matrix of the grafts and the adjacent bone, with evidence of partial replacement of the cartilaginous graft matrix with vascularized woven bone. Based on histomorphometric analysis, the amount of bony repair tissue in the tibial defects varied from 22% to 85%, with an average of 48%. In half of the animals, the woven bone bridged the defects. In contrast, none of four control defects exhibited bridging with bone or cartilage. Periosteal bone formation occurred at the osteotomized bone ends in the control animals, however, fibrous tissue separated the ends of the bones.
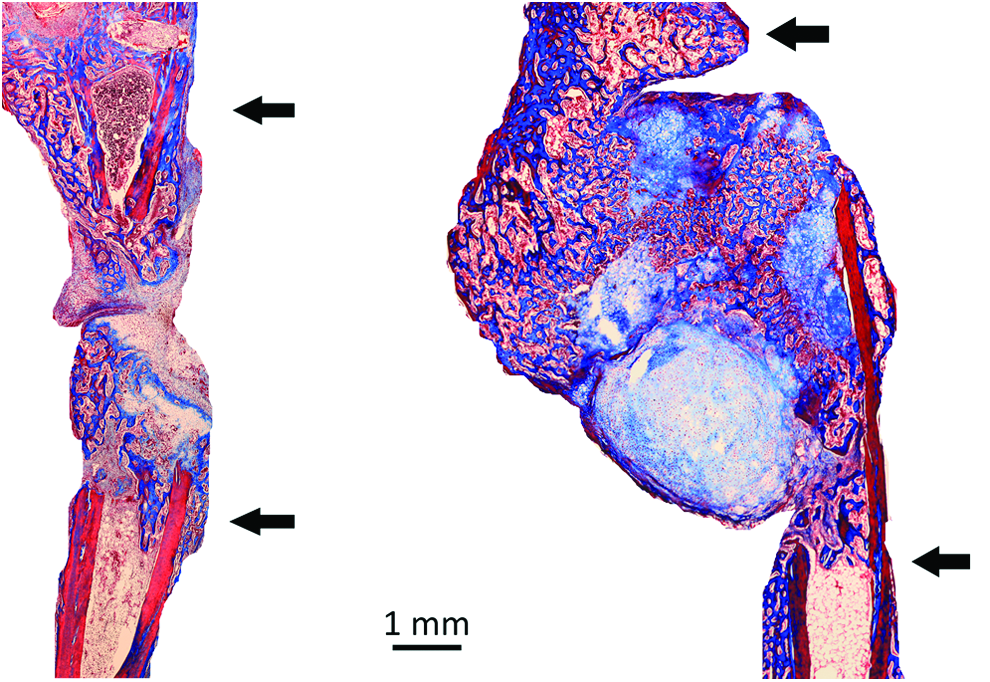
Cartilage grafts from passaged hOA articular chondrocytes can heal critical-size segmental tibial defects in mice. hOA articular chondrocytes were passaged in monolayer with serum-free media and then pelleted with TGF-β1 and BMP4 stimulation in vitro. The resulting grafts were transplanted into externally stabilized murine tibial defects and healing evaluated histologically after 4 weeks (right panel). An empty control defect is on the left. Arrows indicate the proximal (top) and distal (bottom) ends of the tibiae. In the control, fibrous tissue separated the ends of the tibia. There has been remodeling of the engineered cartilage graft into woven bone, with bony bridging of the defect. Masson's trichrome stain. Color images available online at www.liebertpub.com/tea
The host response to the engineered endochondral cartilage grafts was very different from the response to the unmodified hOA articular cartilage. In the latter case, cartilage formed adjacent to the human cartilage, with no evidence of ossification or remodeling of the graft (Figs. 1 and 6, top panel). There was a clear demarcation between host and graft matrices. Compared to empty controls, there was minimal periosteal bone formation. In contrast, the matrix of the endochondral cartilage grafts was continuous with the host woven bone, and channels containing red blood cells penetrated into the grafts (Fig. 6, two center panels). There was only one interface between the host bone and an implanted engineered cartilage graft that showed no evidence of integration, with a clear demarcation between the host bone and repair tissue (Fig. 6, bottom panel).
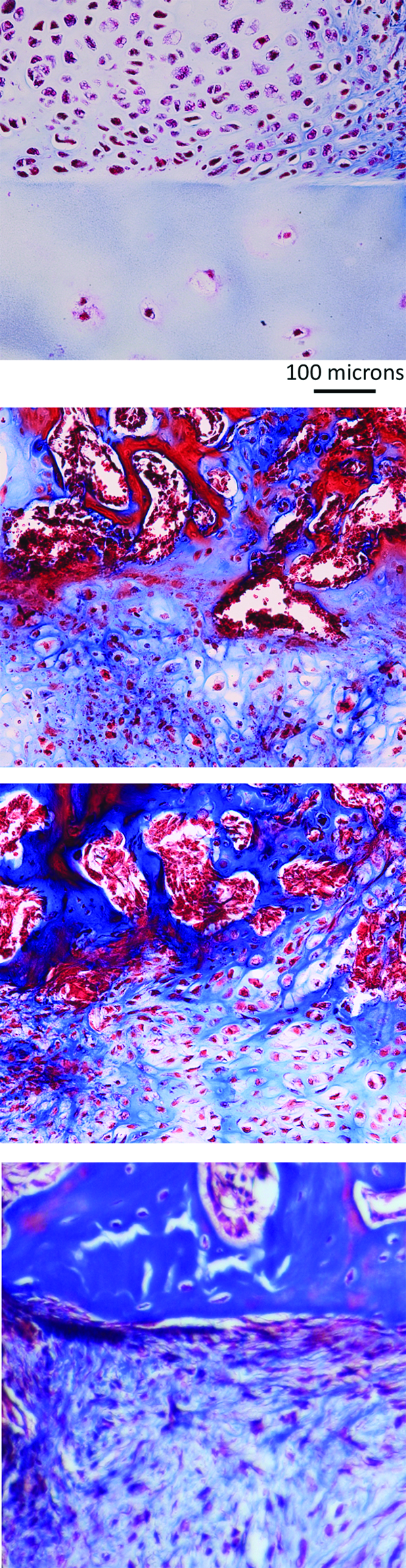
Junctions of host tissue with unmodified hOA articular cartilage and engineered endochondral chondrocyte grafts (tibial defect model). The top panel shows the junction between host tissue and unmodified hOA cartilage. There has been no remodeling of the grafted cartilage, and the host and graft matrices are distinct. The host tissues consist of cartilage that shows no evidence of endochondral bone formation. The two middle panels show junctions between host tissue and engineered cartilage grafts derived from passaged hOA chondrocytes. Implanted grafts included primary and passaged hOA chondrocyte pellets that were first cultured in vitro with TGF-β1 and BMP4. Pellets were derived from 300,000 cells and generated under carrier-free conditions. The host tissue consists of woven bone. There is remodeling of the graft by vascular tissue. The boundaries between bone and cartilage are indistinct. The bottom panel shows the only interface between host bone and an engineered graft that failed to integrate. There is a clear demarcation between the host bone at top and the repair tissue below. In each panel, host tissue is at the top and graft tissue is at the bottom. Masson's trichrome stain. Color images available online at www.liebertpub.com/tea
Finally, we used immunohistochemistry with a human-specific anti-osteocalcin antibody to determine whether the new bone in the tibial defects arose from the host mice or the implanted human cells. This antibody has previously been used to distinguish human-derived bone from mouse-derived bone in xenograft experiments. 14 There was minimal staining of the newly formed bone, suggesting that most, if not all, of the repair bone was derived from the host (Fig. 7).
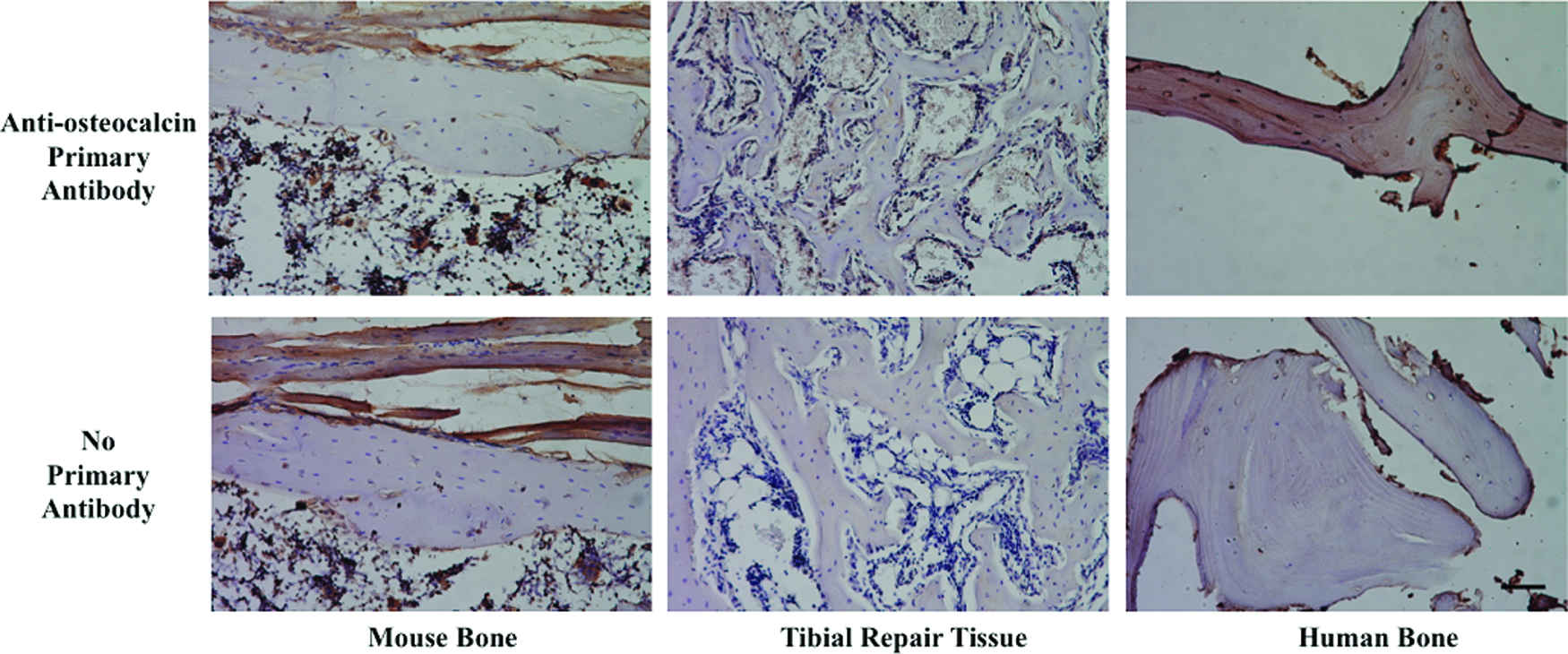
Expression of human osteocalcin in tibial repair tissue. Immunohistochemistry was performed on mouse bone (left column), woven bone from the tibial repair tissue generated by engineered cartilage grafts implanted into mouse tibial defects (center column), or human bone (right column). The top row shows staining with an anti-osteocalcin antibody that recognizes human, but not mouse osteocalcin. The bottom row shows control samples that were not exposed to the primary antibody. Unambiguous staining for human osteocalcin is only seen in the control human bone. Scale bar in bottom right panel is 100 μm. Color images available online at www.liebertpub.com/tea
Discussion
Endochondral grafts represent an emerging paradigm for stimulating bone regeneration, with both cell-based and cell-free matrix-based approaches under development. 15 A number of challenges remain in translating endochondral grafts toward a clinical therapy, including identification of an appropriate cell source to generate hypertrophic cartilage. In this study, we explored the use of hOA articular chondrocytes to form endochondral cartilage grafts. OA cartilage is readily available from discarded tissues from total joint replacements and our study specifically developed a protocol for proliferating and modulating the phenotype of these cells.
Since unmodified hOA cartilage grafts did not support endochondral ossification, we established an in vitro culture protocol that included initial passaging of articular chondrocytes in monolayer followed by pelleting and treatment with growth factors from the TGF-β superfamily. Consistent with previous studies, passaging of articular chondrocytes altered gene expression and led to downregulation of COL2 and aggrecan, and upregulation of COL1.16–20 This process has been referred to as dedifferentiation, but it is unclear if cells regress to a progenitor state or assume an alternative nonphysiologic phenotype. Although previous studies of dedifferentiation involved monolayer expansion in the presence of 10% bovine serum, we used a commercial, proprietary serum-free medium for expansion. This medium is likely to contain fibroblast growth factor 2, platelet-derived growth factor, and TGF-β1 at a minimum and may have different effects than serum-containing media. However, gene expression changes during monolayer expansion of chondrocytes with this medium were grossly similar to those described with serum-containing media.
Downregulation of cartilage-specific genes during monolayer expansion, including those associated with hypertrophy, suggests that passaged hOA chondrocytes lose cartilage specification. Consistent with this, primary articular chondrocytes form stable, ossification-resistant cartilaginous tissues when injected into nude mice, but passaged chondrocytes, lose this ability. 21 Reprogramming to a multipotent state could explain the acquired ability of the passaged cells to differentiate into the alternative endochondral phenotype. Expansion of cell numbers in tissue culture may also select for metabolically active cells that are capable of cell division and differentiation.
The optimal cell source to generate endochondral grafts is unclear. Mesenchymal stem cells (MSCs) and embryonic stem cells (ESCs) can also form tissues that undergo endochondral ossification.2–5,22 However, in contrast to articular chondrocytes, hMSCs lose the ability to undergo chondrogenesis with prolonged passaging. hESCs hold promise for cell-based therapies due to their potential to differentiate toward any tissue phenotype. The challenge with these cells is developing protocols to efficiently control differentiation: heterogeneous cell differentiation and teratoma formation remain problematic. One therapeutic advantage of hMSCs is that they appear to be immunoprivileged due to the absence of major histocompatibility complex class II cell surface markers. 23 In addition, autologous hMSCs may be harvested from patients and used to generate endochondral grafts. Clinical translation of endochondral grafts from hOA cartilage will require the use of allogeneic cells, so the immunogenicity of these grafts must be determined. Encouragingly, articular chondrocytes have also been shown to be immunoprivileged 24 and allogeneic articular cartilages are routinely transplanted into patients.25,26 Furthermore, cells from bioengineered neocartilage derived from juvenile articular chondrocytes do not stimulate an allogeneic T-cell response and instead suppress T-cell proliferation. 27 Since cells from engineered chondrocyte grafts may persist in the host even if the majority of new bone is host derived, the fate and immunological effects of these grafted cells must be determined before clinical evaluation of this technology.
Scalability remains a key challenge to the use of engineered endochondral grafts. We utilized scaffold free, pellet culture to form grafts. Incorporation of cells into an appropriate scaffold may be required to repair larger, clinically relevant bone voids in human subjects. Possibilities for these scaffolds include clinically available osteoconductive materials such as tricalcium phosphate or one of the many novel scaffolds that are in development. 28 Previously, we have shown that engineered scaffolds can be tuned to support specific biological processes 29 and retain phenotype of encapsulated cells given the appropriate growth factor treatment.5,30 Alternatively, a biomaterial consisting of poly(lactide-co-glycolide) and integrated hyaluronic acid may serve as an appropriate scaffold since the combination of this matrix with bone autograft promotes endochondral ossification in a rabbit model of spinal fusion. 31
In conclusion, we have shown that hOA articular chondrocytes from total joint arthroplasty resections can be transformed into endochondral tissues that promote bone formation in vivo. These grafts are able to integrate with host bone, undergo vascularization, and heal critical-size long-bone defects in mice. However, while these cartilaginous tissues efficiently integrated with the host bone, they did not completely remodel into bone. Cartilage constituted the majority of the remaining tissue. Limbs with residual cartilage are likely to resemble fractures in the soft-callus stage of healing and are unlikely to support full weight-bearing. The upregulation of Chm-1 and TSP-1 in the engineered endochondral grafts may contribute to this resistance to complete ossification. In addition, although animals were allowed ambulation with the external fixators, the lack of complete loading may also have inhibited ossification. Techniques to more completely drive the differentiation of articular chondrocytes into a transient bone-forming phenotype must be developed before translation of this proof-of-concept study into clinical practice.
Footnotes
Acknowledgments
Research reported in this publication was supported by the National Institute of Arthritis and Musculoskeletal and Skin Disease (NIAMS) of the National Institutes of Health (NIH) under the following award numbers: CSB (#5F32AR062469). Additional research support was provided by the Musculoskeletal Transplant Foundation (CSB: MTF Junior Investigator Award), the UCSF Graduate Education in Medical Sciences (GEMS), the UCSF Department of Orthopaedic Surgery, and the San Francisco Veterans Affairs Medical Center.
Disclosure Statement
Dr. Kuo has filed a patent on the described method of forming endochondral cartilage grafts. No other competing financial interests exist.