
Abstract
Boron is an essential metalloid, which plays a key role in plant and animal metabolisms. It has been reported that boron is involved in bone mineralization, has some uses in synthetic chemistry, and its potential has been only recently exploited in medicinal chemistry. However, in the area of tissue engineering, the use of boron is limited to works involving certain bioactive glasses. In this study, we engineer poly(
Introduction
B
The unique characteristics of boron have been exploited by synthetic chemists 6 focusing on important medicinal molecules whose production includes the application of organoborane chemistry. The discovery of a simple synthesis route of sodium borohydride by Schlesinger et al. 7 and the subsequent discovery of the hydroboration reaction by Brown and Rao 8 revolutionized organic synthesis. The preparation of organoboronates through transition metal-catalyzed borylation reactions has also greatly facilitated the development of borane chemistry. 9 Particular mention deserves the Suzuki coupling reaction, 10 which has made the synthesis of several pharmaceuticals significantly easier.
The approval of a boron-containing peptidic proteasome inhibitor, bortezomib (Velcade®), by the U.S. Food and Drug Administration (US FDA) for multiple myeloma and mantle cell lymphoma 11 boosted the area of boron in medicinal chemistry. Currently, several boron-containing molecules are in preclinical and clinical stages of development to treat multiple disease conditions, such as inflammation, diabetes, cancer, and many others.12–18
In terms of tissue engineering and biomedical applications, boron has been used for the optimization of biomaterials especially in the bioactive glass research area,19,20 in which new bioactive glasses based on borate and borosilicate compositions have shown the ability to enhance new bone formation when compared with silicate bioactive glass. Indeed, borate-based bioactive glasses also have controllable degradation rates, providing improved conditions for this type of scaffold with applications in hard and soft tissue regeneration. 21 Boron has also been used in the synthesis of biodegradable polymers. 22
In the context of muscle tissue regeneration, only recently a dose-dependent activity of boron in myogenic differentiation of human adipose-derived stem cells 23 has been described. Notwithstanding, the biochemical mechanism of action of boron is not fully understood yet. Until recently, it was believed that boron and borates are transported by passive diffusion through the cell membrane. 24 However, this was reconsidered after the isolation of the Arabidopsis thaliana borate transporter, AtBor1, 25 and its homolog in mammalians, BTR1 26 or NaBC1. 27 NaBC1 has unique transport features. In the absence of borate, it conducts Na+ and OH− (H+). In the presence of borate, it functions as an electrogenic Na+-coupled borate cotransporter.
Considering the vital role of borate as a micronutrient in plants and animals and the uniqueness of boron as a novel challenging element for tissue engineering, here we set out to investigate whether boron promotes cell differentiation per se when released from a poly(
Materials and Methods
Materials
PLLA (Cargill Dow) was dissolved in 100 mL of chloroform at total polymer concentration of 2% (2 g of PLLA). In all experiments, we used sodium tetraborate decahydrate borax 10 mol (Na2B4O7 · 10H2O) (Borax) to load materials and in the culture medium. For abbreviation, hereafter we will designate it as borax-d. Different solutions of PLLA 2%, containing 2% and 5% of borax-d in relation to total mass of polymer (mT = mPLLA + mborax-d), were performed in continuous stirring using 40 mg of borax-d and 1.96 g of PLLA for the 2% samples and 100 mg of borax-d and 1.9 g of PLLA for 5% samples.
Polymer sheets of PLLA, PLLA-borax-d 2%, and PLLA-borax-d 5% were prepared by solvent casting method and dried in vacuo at room temperature. Die cast materials were prepared with 12 or 25 mm diameter before further characterization.
Protein adsorption and quantification
Collagen I (Stemcell) coating for control glass substrates was adsorbed from a solution of concentration of 1 mg · mL−1 for 1 h at room temperature.
Fibronectin (FN) from human plasma (Sigma-Aldrich) was adsorbed on the different substrates for 1 h using an FN solution of concentration 20 μg · mL−1 in Dulbecco's phosphate-buffered saline (DPBS). After adsorption, the supernatant was collected and quantified by the Micro BCA Protein Assay Kit (Thermo Scientific). The surface density of adsorbed FN was obtained as the difference between the amount of FN in the initial solution and in the supernatant.
Characterization of materials
Water contact angle
Static water contact angles (WCAs) were measured using an OCA 20 (DataPhysics Instruments GmbH) and water, reagent ACS (Sigma-Aldrich); the volume of the drop was 5 μL and the measurement was performed after 10 s of substrate water contact. The results are the average of six measurements.
Differential scanning calorimetry
Thermal properties of materials were determined by differential scanning calorimetry (DSC) in N2 atmosphere using a Mettler-Toledo DSC 823e system. Weight of material specimens was 10 mg, and dried samples were sealed in aluminum pans. The range of temperatures for the scans performed was between 20°C and 200°C, and the heating and cooling rate of the scans was 10°C · min−1.
Fourier-transformed infrared spectroscopy
Fourier-transformed infrared spectroscopy (FTIR) was performed using the Thermo NICOLET Nexus FTIR spectrometer using the Smart Multi-Bounce HATR Accessory between 655 and 4000 cm−1 by the smart diffuse reflectance mode. One hundred twenty-eight scans with spectral resolution 7.714 cm−1 were coadded to achieve a good signal-to-noise ratio. All measurements were performed at room temperature.
Atomic force microscopy
Atomic force microscopy (AFM) was performed in a NanoScope III from Veeco operating in the PeakForce QNM mode; the Nanoscope 5.30r2 software version was used. ScanAsyst-Air cantilevers from Bruker were used with drive amplitude of 100 mV and amplitude set point at 250 mV. Cantilever was calibrated against a sapphire crystal to determine the Young modulus. Young's modulus was calculated from nanoindentation experiments using a matrix of 256 × 256 points on different areas of the sample. A total of 65,536 measurements were averaged and represented as mean ± standard deviation (SD).
Borax-d release assay
The in vitro release of borax-d was performed immersing different materials (PLLA-borax-d-loaded 2% and 5% and PLLA. Total mass of the sample was ∼4.5 mg, and total mass of borax-d per sample was 90 μg for 2% and 225 μg for 5% compositions, respectively) of 12 mm diameter in 1 mL of DPBS (Sigma-Aldrich). Short-term release studies (maximum time 180 min) were performed in two different ways: removing a 40 μL aliquot from the medium and removing the total amount of DPBS (1 mL) every 10 min. Long-term release studies were performed simulating a 15-day cell culture in humidified atmosphere at 37°C and 5% CO2. Aliquots consisting of the total amount of DPBS (1 mL) were removed from the plates after 3, 6, 9, 12, and 15 days. Reaction of borax-d present in the 40 μL aliquots with azomethine (Sigma-Aldrich) in acid medium (KAc/HAc buffer pH 5.2) originates a colorimetric reaction measured at 405 nm in a Victor III (PerkinElmer) device. Standards for calibration were prepared at concentrations of 0, 0.2, 0.3, 0.5, 1, 1.5, 2, 5.2, and 52 mg · mL−1 of borax-d, using 40 μL aliquots from the original standard solutions for colorimetric reactions.
Cell culture
Murine C2C12 cells (ATCC CRL-1772) were maintained in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 20% fetal bovine serum (Invitrogen) and 1% penicillin–streptomycin (Lonza) in humidified atmosphere at 37°C and 5% CO2. Cells were subcultured before reaching confluency (approximately every 2 days). For differentiation experiments, 20,000 cells · cm2 were seeded over FN-coated PLLA and PLLA-borax-d 2% and 5% materials in DMEM supplemented with 1% ITS (insulin–transferrin–selenium; Life Technologies) and 1% penicillin–streptomycin to induce myogenic differentiation.
Cytotoxicity assay
MTS quantitative assay (The CellTiter 96 Aqueous One Solution Cell Proliferation Assay; Promega) was performed to assess cytocompatibility of PLLA materials with C2C12 cells and to establish the maximum working concentration of borax-d to use; 20,000 cells · cm2 were seeded onto FN-coated PLLA and glass substrates and metabolic activity was measured after 24, 48, and 72 h of incubation of cells with different concentrations of borax-d in media (52.4, 131, and 262 mM, equivalent to 2%, 5%, and 10% w/v). Cells were incubated for 3 h with MTS (tetrazolium salt) at 37°C and the formation of formazan product was followed by measuring absorbance at 490 nm. All measurements were performed in triplicate.
Myogenic differentiation and quantification
C2C12 cells were cultured on FN-coated materials for 4 days under differentiation conditions in the absence of serum and supplemented with 1% ITS. Then, they were immunostained for sarcomeric myosin. Briefly, cultures were fixed in 70% ethanol/37% formaldehyde/glacial acetic acid (20:2:1), and then blocked in 5% goat serum (Invitrogen) in DPBS (Sigma-Aldrich) for 1 h. Samples were sequentially incubated in MF-20 mouse primary antibody (Developmental Studies Hybridoma Bank, University of Iowa) and anti-mouse Cy3-conjugated secondary antibody (Jackson Immunoresearch). Finally, samples were washed before being mounted in Vectashield containing DAPI (Atom) and observed under an epifluorescence microscope (Nikon Eclipse 80i). Images from the fluorescence microscope (DAPI channel—nuclei, and Cy3 channel—sarcomeric myosin) of the C2C12 culture were acquired at 10× magnification (n = 10), transformed to an 8-bit grayscale bitmap (Fiji-ImageJ software), and segmented using the Trainable Weka Segmentation plugin to create a binary mask for both DAPI and Cy3 channels. Total nuclei per image were counted using the particle analysis command. Then, the segmented DAPI channel image was subtracted from the Cy3 channel segmented image, and the remaining nuclei were counted and assigned to nondifferentiated cells. The fraction of differentiated cells was calculated subtracting the nondifferentiated nuclei from the total nuclei counted.
For differentiation experiments, including borax-d as additive in the medium, two different concentrations of the product were tested. The first one was 225 μg · mL−1 (0.59 mM) corresponding to an equivalent amount of borax-d included in the PLLA-borax-d 5% sample. The second one was 22.5 μg · mL−1 (0.059 mM) equivalent to 1:10 dilution. The amount of borax-d is expressed as the final concentration of borax-d in the culture medium. These borax-d solutions were added to media on cells cultured onto FN-PLLA substrates on the days 1 and 3, respectively, as a supplement in differentiation medium with ITS 1% until myogenesis.
Focal adhesion kinase and phosphorylated focal adhesion kinase detection and quantification
For focal adhesion kinase (FAK) assays, cells were cultured under serum-free conditions to reduce the background signal. Cells were then seeded on FN-coated materials in serum-free medium. After 3 h of culture, cells were lysed with RIPA buffer (Tris–HCl 50 mM, 1% Nonidet P-40, 0.25% Na deoxycholate, NaCl 150 mM, EDTA 1 mM) containing protease inhibitor cocktail tablets (Roche). Proteins were concentrated using Microcon YM-30 Centrifugal Filter devices (Millipore) as described by the manufacturer. To determine FAK protein expression and its phosphorylated form (pFAK), concentrated samples were subjected to 7% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Proteins were transferred to a positively charged polyvinylidene fluoride (PVDF) membrane (GE Healthcare) using a semidry transfer cell system (Bio-Rad) and blocked by immersion in 5% semi-skimmed milk in phosphate-buffered saline (PBS) for 30 min at room temperature. The blot was incubated with anti-FAK (400 ng · mL−1; Abcam) and anti-pFAK (1 mg · mL−1; Abcam) in PBS containing 0.1% Tween 20 and 2% semi-skimmed milk. After several washes with PBS/0.1% Tween 20, the blot was incubated in horseradish peroxidase-conjugated antibody (GE Healthcare) diluted 1:25,000 for FAK and 1:10,000 for pFAK in PBS containing 0.1% Tween 20 and 2% semi-skimmed milk for 1 h at room temperature. Supersignal West Femto Maximum Sensitivity Substrate (Pierce) was used before X-ray film exposition.
Effect of calcium, calcium chelator, and calcium channel blocker on C2C12 differentiation
C2C12 cells were cultured for 4 days in the differentiation medium (in the absence of serum and substituted for ITS 1%) supplemented with 0.35 mM calcium chloride (Sigma-Aldrich) as exogenous calcium ions or 0.35 mM calcium chelator ethylene glycol tetraacetic acid (EGTA; Sigma-Aldrich) or 10 μM of verapamil (Sigma-Aldrich) as an L-type calcium channel blocker. After induction of differentiation under the different conditions assayed, culture cells were fixed and immunostained for sarcomeric α-actinin (Abcam) with the same procedure explained above. Tile images were obtained with confocal microscopy (Zeiss LSM 780, Axio Observer). The tile images acquired were the representation of total of samples analyzed (25 images per sample). Myotubes were defined and counted as cells with three or more nuclei and the total amount of nuclei was defined as explained above, creating a binary mask. All experiments were performed in triplicate.
Gene expression analysis by quantitative real-time polymerase chain reaction
Total RNA was extracted from C2C12 cells cultured for 4 days under different experimental conditions using an RNeasy micro kit (Qiagen) according to the manufacturer's protocol. RNA quantity and integrity were measured with a NanoDrop 1000 (Thermo Scientific). Then, 150 ng of RNA was reverse transcribed using the Superscript III reverse transcriptase (Invitrogen) and oligo dT primer (Invitrogen). Real-time quantitative polymerase chain reaction (qPCR) was carried out using the SYBR select master mix and 7500 Real-Time PCR system from Applied Biosystems. The reactions were run in triplicate for both technical and biological replicas. The primers used for amplification were designed based on sequences found in the GenBank database and included MyoD1 (GenBank M18779.1, Forward: 5′-CGC TCG TGA GGA TGA GCA T-3′, Reverse: 5′-AGC GTC TCG AAG GCC TCA T-3′) and Myogenin (NM_031189.2, Forward: 5′-TGC CGT GGG CAT GTA AGG T-3′, Reverse: 5′-TGC GCA GGA TCT CCA CTT TAG-3′).
The fractional cycle number at which fluorescence passed the threshold (Ct values) was used for quantification using the comparative Ct method. Sample values were normalized to the threshold value of the housekeeping gene GAPDH:
Statistical analysis
Results are shown as mean ± standard deviation for experiments in which the sample size was low and mean ± standard error of the mean (SEM) for those experiments in which the sample size was higher (n > 3) (SEM quantifies precisely the true mean of the population as it takes into account both the value of the SD and the sample size).
Protein quantification for western blot bands and differentiated cell quantification were analyzed by the nonparametric ANOVA test using a Dunn post hoc multiple column comparison test. p < 0.05 was considered significant and is indicated with an asterisk on each figure. All experiments were performed at least per triplicate.
Results and Discussion
Characterization of material substrates
PLLA is a well-known biodegradable material approved by the FDA for several biomedical applications with remarkable properties for tissue engineering such as excellent biocompatibility and processability. 30 We have confirmed the presence of borax-d within the samples using DSC and FTIR. DSC heating curves show cold crystallization and melting induced by the presence of borax-d as a nucleating agent (pure PLLA is unable to crystallize on heating due to slow kinetics) (Fig. 1A). 31 Likewise, the characteristic borax-d peak is observed by FTIR (Fig. 1B). The peak at 1350 cm−1 corresponds to the B-O bond of the borax-d and its intensity increases as the content of borax-d on the sample does. To assess the role of borax-d on surface properties, WCA was measured on the base material and after loading with different borax-d ratios. Figure 1C shows similar WCA values regardless of the amount of borax-d loaded in PLLA, suggesting that the presence of borax-d does not alter surface wettability of the base polymer and disregards any effect of this important surface property on cell behavior.
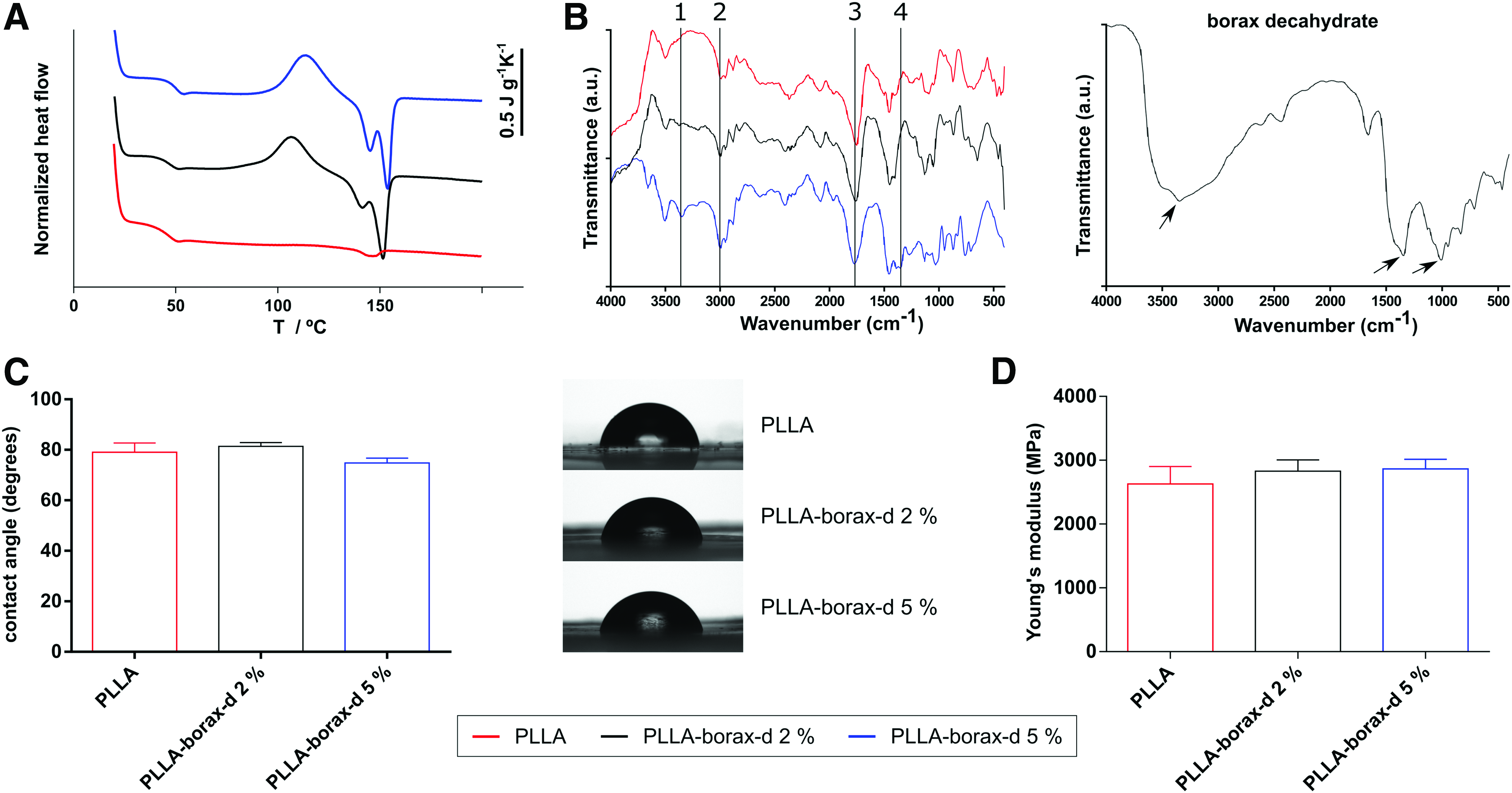
Physicochemical characterization of the material substrates (poly(
In addition, we disregarded any effect of borax-d on the mechanical properties of the different systems as this is known to be a key property to trigger cell fate.32,33 We have measured local elasticity on the material surfaces by AFM measurements operating in the PeakForce QNM (Fig. 1D), which reveals that borax-d content does not affect the stiffness of the base PLLA.
We also quantified adsorbed FN on PLLA and PLLA-borax-d-loaded substrates by the Micro BCA Protein Assay Kit. Results of quantification showed similar values for all substrates without statistical significance, which disregard any influence of borax-d on the amount of FN adsorbed (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
Borax-d release
We next examined the kinetics of borax-d release from the material systems. Figure 2 shows borax-d released into the medium using the azomethine colorimetric reaction and measuring the absorbance at 405 nm. 34 Substrates were immersed in DPBS, and aliquots of 40 μL and 1 mL of buffer were collected every 10 min for short-term release studies (Fig. 2A, B). For long-term release assays, 1 mL aliquots were removed after 3, 6, 9, 12, and 15 days, simulating a standard long cell culture (Fig. 2C). Borax-d release followed a one-phase exponential decay, indicating that the release of borax-d increases as the concentration of borax-d in the sample does. It is important to emphasize that major release was produced during the first 3 h of culture. Note that myotube formation during C2C12 differentiation reaches a plateau after 4 days of culture, ensuring that borax-d is participating in the myogenic process during the timing of experiments. We also wondered if PLLA degradation may have a role in the release of borax-d from samples. The initial release of borax-d will occur through dissolution of superficial borax-d after immediate contact with the culture media. The sustained release should then be related to the hydration of PLLA that will lead to PLLA degradation in the long term. For the samples used in this work, the percentage of PLLA mass lost is only ca. 9% after 1 month of degradation experiment (data not shown). Considering that the degradation kinetics of PLLA is much slower than the time needed for cells to differentiate on our surfaces and that borax-d release occurs at the very initial time disregards the effect of degradation of PLLA as a consequence of release mechanism of borax-d from the materials.

Borax-d release studies.
Cell differentiation on borax-d-loaded materials
We used borax decahydrate in this work, so the uptake of boron by cells must be actually in the form of borate anions. To evaluate the biological activity of borax-d-loaded materials, we quantified the expression of sarcomeric myosin and the presence of fused myotubes in C2C12 myoblasts as markers of myogenesis. 35 Myotube formation was significantly higher on the PLLA-borax-d 5% substrates (Fig. 3A, B) compared with the PLLA-borax-d 2% and PLLA ones. Surprisingly, myogenic differentiation was considerably more robust on the PLLA-borax-d 5% than on collagen type I, which represents a standard substrate for myogenic differentiation. 36

Myogenic differentiation on the different substrates.
Despite that the initial cell density was the same for all substrates, final cell density at the end of the culture was not constant throughout substrates due to the effect of intrinsic material properties in cell behavior (in this particular case determined by borax-d content). It is also important to note that the final cell number does not correlate with the level of myogenic differentiation as shown in Figure 3C, which disregards differentiation to be a consequence of cell density as previously reported. 37 Furthermore, the number of cells on the PLLA-borax-d 5% and PLLA-borax-d 2% samples was slightly lower than on the collagen I control (Fig. 1C), which reveals the potential of borax-d to induce fusion into multinucleated myotubes. To further evaluate the biological activity of borax-d in cell differentiation and to ensure that the effect on cell differentiation was due solely to the borax-d released into the culture medium, we used, as a control, culture media with two different concentrations of borax-d, whereas cells were seeded on FN-PLLA surfaces. Previous to these experiments, we tested several concentrations of borax-d diluted in the culture medium to assess cytotoxic levels. We found that a solution of concentration of borax-d 52.4 mM in the culture medium was the maximum amount previous to cell death (Supplementary Fig. S2). We then estimated the equivalent amount of borax-d included in the PLLA-borax-d 5% sample, weighting a die cut disc and assuming that 5% of the sample weight corresponded to borax-d. Subsequently, an equivalent amount of borax-d was added to the culture medium, which effectively corresponded to the solution of concentration 225 μg · mL−1. Both this solution (equivalent to PLLA-borax-d 5% sample) and another one diluted 10 times (22.5 μg · mL−1) showed enhanced myotube formation (Fig. 4A) and similar level of differentiation than cells on PLLA-borax-d 5% (Fig. 4A, B). These results support the idea that the effect of borax-d loaded in PLLA occurs through a sustained release of borax-d in the medium—as characterized in Figure 2—and it might be related to dissolution of superficial borax decahydrate salt after immediate contact with the culture media. Once again, differentiation levels attained due to the presence of borax-d in the culture medium were not due to differences in the final number of cells on the substrates (Fig. 4C). Our results demonstrate that sustained release of borax-d from PLLA enhances myoblast differentiation at a very early stage, probably accelerating the entry of cells to the differentiation pathway earlier than control myoblasts.

Myogenic differentiation on the different substrates cultured with addition of borax-d in the medium.
Cell signaling
The requirement of β1 integrins for myoblast fusion is essential both in vivo and in vitro. 38 However, the downstream effectors of integrins that regulate myoblast fusion are unclear. To gain insights into the mechanisms controlling the enhanced cellular responses to borax-d-loaded PLLA, we examined FAK as a downstream effector of integrins. FAK has been identified as the key cytoplasmic tyrosine kinase that transmits integrin-mediated signals at focal adhesions in several cell types.39,40 FAK phosphorylation at Tyr397 plays a central role during myoblast differentiation in 2D cultures. 41 To test whether FAK is activated due to the release of borax-d from PLLA, we examined the phosphorylation of Tyr397 by western blot. No differences in FAK phosphorylation were observed among borax-d-loaded substrates and collagen I-coated control of differentiation (Fig. 5), suggesting that enhanced cell differentiation triggered by borax-d comprehends other metabolic pathways rather than phosphorylation of FAK. We have previously found the same level of FAK phosphorylation on materials that promote a significant level of C2C12 differentiation due to the activation of contractility phospho myosin light chain (pMLC) downstream FAK. 42

Effect of calcium in cell differentiation and gene expression on borax-d-loaded materials
Extracellular Ca2+ plays an important role in muscle development. Various reports have previously demonstrated the importance of Ca2+ signaling in the activation of myogenic transcription factors and myogenesis.43–47 Indeed, it has been reported that migration or fusion, which precedes myotube formation, requires Ca2+ influxes.48,49
To further assess the role of borax-d in skeletal myogenesis and to elucidate its possible mechanism of action in relation to Ca2+, we performed different studies combining borax-d and Ca2+ ions. We used (i) addition of CaCl2 in the culture medium, (ii) a chelating agent (EGTA) to sequester metal complexes (e.g., based on Ca2+), and (iii) verapamil as a calcium channel blocker directed against L-type Ca2+ channels in muscle cells. Myotube formation under different conditions was evaluated by immunostaining of sarcomeric α-actinin and further visualized by laser scanning confocal microscopy. After induction of differentiation, PLLA-borax-d 2% and 5% presented clear accumulation of sarcomeric α-actinin in a punctate manner along the longitudinal axis of the myotube (Fig. 6A inset, open arrowhead) associated with actin filaments. These aggregations of sarcomeric α-actinin that are not yet laterally aligned correspond to Z bodies, a key marker of early stage sarcomere development.50,51 This result further suggests that borax-d enhances skeletal myogenesis, regulating the developmental program that directs myoblasts to differentiate into muscle fibers.

Results showed that the addition of CaCl2 has a positive effect on myotube formation in the absence of borax-d (FN-PLLA, Fig. 6B) as we expected based on previous reports.45,52 The observed effect of CaCl2 seems to be null on PLLA-borax-d 2% substrates and, furthermore, it leads to lower levels of differentiation on PLLA-borax-d 5% substrates (Fig. 6A, B), which strongly suggest that the enhanced differentiation associated with the presence of borax-d involves a mechanism that has to do with the presence of Ca2+ in the medium as well. To test this further, we assessed that either the chelation of Ca2+ in the culture medium (addition of EGTA) or the blocking of the Ca2+ channel (verapamil) led to strong effects on myotube formation for all samples analyzed—in agreement with other studies—which further reveals that the effect of borax-d depends strongly on the presence of Ca 2 . For example, Zanou et al. 52 reported that inhibiting Ca2+ fluxes decreases PI3K/Akt activation and subsequently slows down myoblast migration. Other studies have reported that Ca2+ channel blockers reduce—and Ca2+ stimulation increases—transcription and impair muscle regeneration. 53 Nevertheless, despite the absence of myotube formation, we detected a great number of cells positive for sarcomeric α-actinin in samples treated with verapamil (Fig. 6A). It is important to remark this observation as we considered differentiated cells to be both positively stained and with myotubes containing three or more nuclei (to avoid counting replicating binucleated cells).
Calcium signaling is essential for the differentiation of many cell types, including skeletal muscle cells, but its mechanisms remain elusive due to broad effects of calcium participating in different metabolic pathways. The lack of effect of extracellular Ca2+ on cell differentiation observed on borax-d-loaded substrates could be controversial if we consider only the transport of Ca2+ through the channel on the plasma membrane: one could argue that the higher the amount of Ca2+, the higher the level of cell differentiation. 54 However, it has to be considered that an increase in the intracellular concentration of Ca2+ can also be caused by the release from intracellular stores controlled by second messengers or Ca2+ itself.45,55
An alternative explanation concerning Ca2+ effects on borax-d-loaded materials lies in the fact that borax-d actually chelates Ca2+ as borax decahydrate is widely used for water softening (calcium ion sequestration) in detergents. In this case, addition of extracellular Ca2+ might sequester borax-d released from PLLA and prevent its association and active transport by Na+-coupled borate cotransporter, NaBC1, 27 hindering Ca2+ entering through the cell membrane. The formation of the complex borate anion-Ca2+ would involve less Ca2+ available and as a result similar effects to EGTA or verapamil chemicals were obtained. This chelating effect is increased as the content of extracellular borax-d does (Fig. 6).
We also analyzed the effects of borax-d in the gene expression of two myogenic regulatory factors, MyoD and Myogenin, as they are essential transcriptional regulators of skeletal muscle differentiation. These myogenic transcription factors are controlled by cellular signaling, resulting in repression of their activity in proliferating myoblasts and enhanced activity during myogenic differentiation.35,56 Real-time qPCR reactions were performed for all samples using collagen I-coated substrates as standard control for myoblast differentiation. The results showed diminished values of relative expression for MyoD on borax-d-loaded materials after CaCl2, EGTA, or verapamil addition compared with control substrates (Fig. 7A) in agreement with results obtained in Figure 6. Nevertheless, Myogenin levels were weakly reduced under the different conditions assayed (Fig. 7B) and no relevant differences were observed among samples.

Total RNA from C2C12 cells cultured under different conditions was isolated for real time quantitative polymerase chain reaction (real time qPCR). Ct values obtained were normalized with Ct values of GAPDH as the housekeeping gene and referenced to collagen I control of differentiation.
Differentiation of skeletal muscle cells is a multistep complex process, in which muscle precursor cells initially express early differentiation markers, exit the cell cycle, then express muscle-specific structural genes, and finally fuse to form multinucleated myotubes; so, all these events are temporarily separable and could be potential steps on which borax-d can play a role, acting over different unknown targets. Our results can be understood if we consider that MyoD and Myogenin are proteins participating in different steps of the differentiation process. MyoD is required for commitment to the myogenic lineage, whereas Myogenin plays a critical role in the expression of the terminal muscle phenotype previously established by MyoD and other myogenic factors such Myf5 and MRF4. 57 Indeed, some transcription factors belonging to the MyoD family have redundant functions, thus we cannot argue similar expression levels for both transcription factors analyzed.
Overall, our results suggest a potential effect of borax-d enhancing myogenesis at early initial steps, although further research is needed to shed light on the molecular mechanism of borax-d in cells.
Conclusions
We have engineered and characterized a degradable PLLA-based system to release borax-d and determined the possible effects of borax-d on myogenesis. We have carried out a comparative analysis of genes that act as early markers of myogenesis and evaluated their expression in the presence of borax-d. Our results suggest a great potential of borax-d to enhance myogenic differentiation at early differentiation steps, and we also elucidate the possible mechanism of action of borax-d in relation to Ca2+. This research highlights borax-d-loaded materials as a relevant and robust strategy for striated muscle tissue engineering.
Footnotes
Acknowledgments
The financial support from ERC (306990), the Spanish Ministry of Economy and Competitiveness (MINECO), and FEDER funds through the MAT2012-38359-C03-01 project are acknowledged. CIBER-BBN is an initiative funded by the VI National R&D&I Plan 2008–2011, Iniciativa Ingenio 2010, Consolider Program, CIBER Actions and financed by the Instituto de Salud Carlos III with assistance from the European Regional Development Fund. The Electron Microscopy Service of the UPV and Laser Scan Confocal Microscopy Service of IBMCP are acknowledged as services for obtaining the images realized for this work. The authors also acknowledge Solaz MA from Borax España for the borax decahydrate supply used in the experimental research.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.