
Abstract
The potential of increasing bone mass and preventing fractures in osteoporosis using stem cell therapy is currently an area of intense focus. However, there are very little data available regarding the postfracture bony defect healing efficacy under osteoporotic conditions. This study aims to investigate whether critical-sized segmental bone defects in a rabbit model of osteoporosis could be repaired using an allogenic stem cell-based tissue engineering (TE) approach and to investigate the potential influence of osteoporosis on the treatment efficacy. Rabbit fetal bone marrow mesenchymal stem cells (BMSCs) were harvested and expanded in vitro. Decalcified bone matrix (DBM) scaffolds were then seeded with allogenic fetal BMSCs and cultivated in osteogenic media to engineer BMSC/DBM constructs. Critical-sized radial defects were created in ovariectomized (OVX) rabbits and the defects were repaired either by insertion of BMSC/DBM constructs or by DBM scaffolds alone. Also, nonovariectomized age-matched (non-OVX) rabbits were served as control. At 3 months post-treatment under the osteoporotic condition (OVX rabbits), the BMSC/DBM constructs inserted within the defect generated significantly more bone tissue when compared to the DBM scaffold as demonstrated by the X-ray, microcomputed tomography, and histological analyses. In addition, when compared to a normal nonosteoporotic condition (age-matched non-OVX rabbits), the defect treatment efficacy was adversely affected by the osteoporotic condition with significantly less bone regeneration. This study demonstrated the potential of allogenic fetal BMSC-based TE strategy for repairing bone defects in an osteoporotic condition. However, the treatment efficacy could be considerably compromised in the OVX animals. Therefore, a more sophisticated strategy that addresses the complicated pathogenic conditions associated with osteoporosis is needed.
Introduction
O
A large amount of research suggested that the impaired osteogenic differentiation potential of bone marrow mesenchymal stem cells (BMSCs) might be one of the major reasons leading to osteoporotic bone loss. Enhancing the proliferation and osteogenic differentiation potential of BMSCs in osteoporotic animals has been investigated as a possible cell-based solution to this disease.3,4 Recently, the application of BMSCs or adipose-derived stromal cells was shown to be able to enhance bone regeneration.5,6 However, the main focus of these studies was the increase of bone mass and prevention of fracture.
A pathologic fracture is one of the major complications of osteoporosis. Typical fragility fractures usually occur in the bones of vertebral columns, ribs, hips, and wrists, leading to further disability and early mortality. Fracture of long bones and hip bones usually requires surgical treatment, and the bone healing is relatively poor in such patients. 7 Tissue engineering (TE) approach has been approved as a new strategy for bone regeneration and repair. However, majority of studies were performed in young and/or healthy recipients that possessed active tissue regenerative capacity.8–10 Whether this approach could repair bone defects under osteoporotic conditions remains unexplored.
Autologous mesenchymal stem cells (MSCs) derived from both bone marrow and adipose tissue have been widely used for bone regeneration. However, the quantity and quality of BMSCs with osteogenic potential would be greatly impaired in osteoporotic patients.11,12 Thus, it is crucial to find alternative cell sources for therapeutic purposes. MSCs could be derived from different tissue sources at various developmental stages, such as fetal bone marrow, umbilical cord, placenta, adult bone marrow, adipose tissue, and so on. Compared with perinatal and adult sources of MSCs, fetal BMSCs exhibited greater cell proliferational capability and osteogenic potentials and lower immunogenicity as reported by previous studies by us and others,13–15 indicating that they might become a good candidate cell source for bone TE application in osteoporotic patients.
Previously, we have conducted a series of research work on fetal BMSC-based bone TE and successfully developed an off-the-shelf-engineered bone graft strategy for repairing large bone defects under normal and healthy conditions.16–19 In the present study, we further explore the potential of fetal BMSC-mediated bone TE strategy for repairing large bone defects in an osteoporotic animal model, and we will meticulously investigate the influence of osteoporotic pathological conditions on the healing efficacy of this strategy.
Materials and Methods
Isolation and culture of fetal BMSCs
BMSCs were isolated and expanded in culture as previously described. 5 Briefly, fetal rabbits (28 days old) were sacrificed and their femurs and tibiae were harvested. The ends of the tibiae and femurs were cut and the bone marrow was flushed out with Dulbecco's Modified Eagle's Medium (DMEM; HyClone, Logan City, UT). Cells were then cultured in DMEM in the presence of 10% fetal bovine serum (FBS; Hyclone) and 1% penicillin and streptomycin (Gibco, Grand Island, NY) at 37°C with 5% CO2. Cells were passaged every 3–4 days and used for further experiments at passage 3.
Osteogenic differentiation of fetal BMSCs
Osteogenic differentiation of fetal BMSCs were carried out using a standard protocol.
5
Fetal BMSCs at passage 3 were seeded in a six-well plate at a density of 100,000 cells per well. The culture medium was replaced by an osteogenic medium (DMEM supplemented with 10% FBS, 10 mmol/L β-glycerophosphate, 0.1 mmol/L dexamethasone, and 50 mmol/L
Fabrication of DBM and BMSC/DBM constructs
Decalcified bone matrix (DBM) scaffolds were obtained by demineralization of the porcine caput femoris as previously described. 20 After being degreased, dialyzed, and demineralized, DBM scaffolds were cut into cuboids sized 1.5×0.3×0.5 cm. BMSC/DBM constructs were fabricated as previously described. 21 Rabbit fetal BMSCs at passage 3 were collected and resuspended at a cell concentration of 2×106/mL. Each DBM cuboid was seeded with 0.1 mL of cell suspension. After 20 min of incubation, the BMSC/DBM constructs were replenished with the osteogenic medium and cultured for 10 days before transplantation. To detect the cell growth on DBM scaffolds, a scanning electron microscopy (SEM) examination was performed after 10 days of culture as previously described. 15 Fluorescein diacetate/propidium iodide (FDA/PI) staining was also performed at the same time point to detect live and dead cells in the constructs. FDA stains the cytoplasm of viable cells green and PI stains the necrotic and apoptotic cell nuclei red. Stained samples were viewed under a confocal laser microscope (Leica Microsystems, Wetzlar, Germany).
Rabbit osteoporosis model
The rabbit osteoporosis model was created by bilateral ovariotomy (OVX) as described. 5 A total of 24 female New Zealand white rabbits (5 months old) were randomized into two groups. Group 1 (n=15) animals were subjected to bilateral OVX and group 2 (n=9) animals were subjected to sham surgery. Each animal was individually caged and fed with normal chow. The bone volume was evaluated before and 12 months after OVX. At 12 months postsurgery, three animals from each group were sacrificed and their distal radius and ulna were harvested for microcomputed tomography (micro-CT) measurement (μCT-80; Scanco Medical, Bassersdorf, Switzerland) to confirm the osteoporotic status.
Critical-sized radial defect and transplantation
A diagram of the experimental procedure is shown in Figure 1. Both osteoporotic rabbits at 12 months post-OVX (n=12) and age-matched rabbits at 12 months postsham surgery (non-OVX, n=6) were enrolled in the transplantation study. The rabbits were randomly divided into three groups: (1) BMSC/DBM in the non-OVX group (n=6), in which defects were filled with BMSC/DBM constructs in the rabbits without OVX; (2) BMSC/DBM in the OVX group (n=6), in which defects were filled with BMSC/DBM constructs in the OVX rabbits; and (3) DBM in the OVX group (n=6), in which defects were filled with DBM scaffolds alone in the OVX rabbits. The animal surgery was performed under a sterile condition. All rabbits were anesthetized by intramuscular injection of 3% pentobarbital sodium (30 mg/kg). A longitudinal skin and musculature incision ∼3 cm long was made and a unilateral segment of the periosteum and radius with a critical-sized length of 1.5 cm was cut off in the middle of the radius using a circular saw. Subsequently, the defect site was filled with a DBM scaffold or a BMSC/DBM construct prepared as described above. Finally, the underlying musculature and skin were sutured. After the surgery, the rabbits received 3 days of intramuscular injection of penicillin (1×105 U/kg) and were individually caged and fed normally.
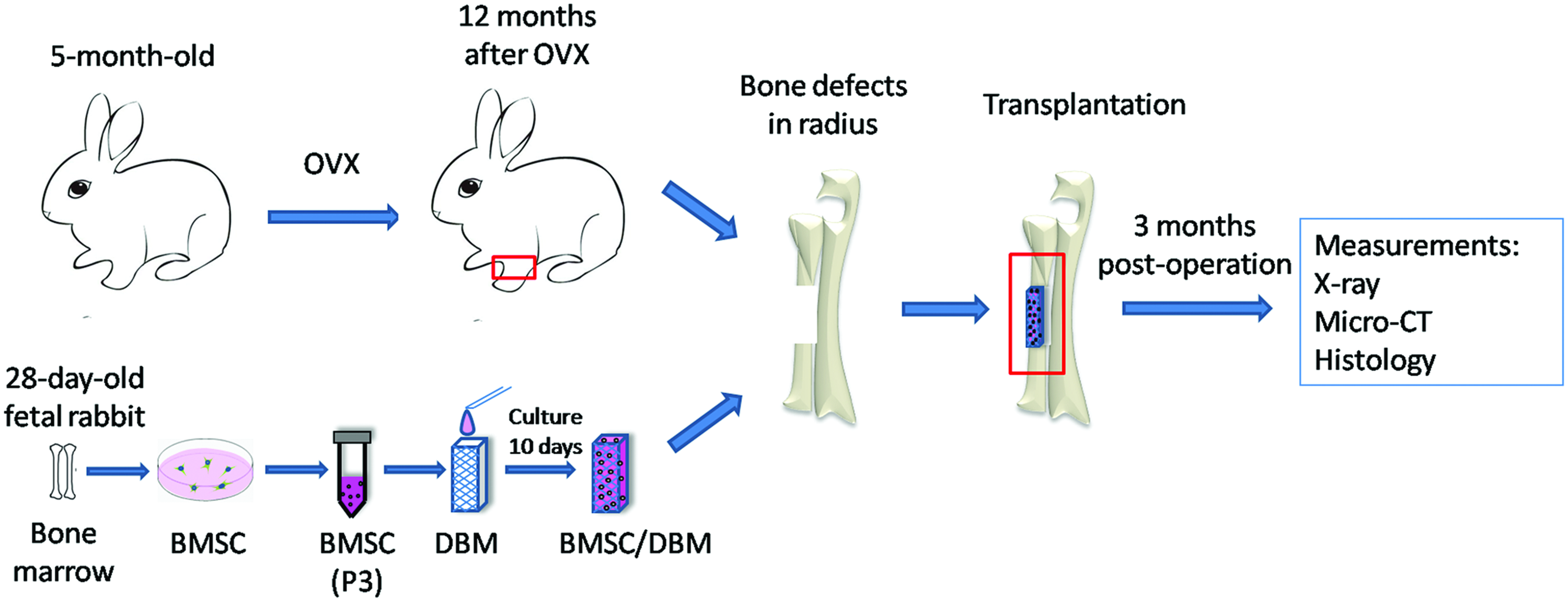
Diagram of experimental procedure. Female rabbits (5 months old) underwent bilateral ovariotomy (OVX) or sham surgery. The osteoporotic status was confirmed at 12 months after OVX. Allogenic fetal BMSCs were obtained from fetal rabbits (28 days old). The third passage cells were seeded on DBM and cultivated in the osteogenic medium for 10 days to make BMSC/DBM constructs. Critical-sized segmental bone defects were created in radii of OVX (n=12) and non-OVX (n=6) rabbits and repaired with DBM scaffolds or BMSC/DBM constructs. Bone regeneration was followed up in all animals up to 3 months postoperation. BMSCs, bone marrow mesenchymal stem cells; DBM, decalcified bone matrix. Color images available online at www.liebertpub.com/tea
Plain X-ray and micro-CT examination
Plain X-ray images were taken at 1 and 3 months post-transplantation, covering the segment of the radius. All animals were sacrificed 3 months later and the radius and ulnas were harvested for micro-CT scanning (μCT-80, 70 kV, 114 mA, 20-mm pixel size). The region of interest (ROI) was set as a cylinder (36 mm in diameter and 10 mm in height) at the central part of the radius, and the three-dimensional image was reconstructed. The quantitative morphometric analysis of the ROI was performed automatically by micro-CT auxiliary software (VG studio Volume Graphics GmbH, Germany), including parameters of bone volume/tissue volume ratio, bone volume (mm3), trabecular number (1/mm), and trabecular separation (mm).
Histological evaluation
At 3 months post-transplantation, radiuses from each group (n=6/group) were harvested and fixed in 10% formalin for 10 days. After being immersed in 10% ethylene diamine tetraacetic acid to demineralize for 30 days, specimens were embedded in paraffin and sectioned into 5-μm slices using a microtome. Sections were then stained with hematoxylin and eosin (H&E) and Masson's Trichrome to visualize the tissue morphology and collagen formation. TRAP (Sigma) staining was performed according to the standard procedure to detect the number of TRAP-positive multinucleated osteoclasts. 22 Five representative fields were recorded and the positive cells were calculated by Image-Pro Plus software (Media Cybernetics, Rockville, MD).
Statistical analysis
All data collected are presented as mean±standard deviation. The statistical analysis was performed with one-way ANOVA. The differences were considered to be statistically significant at p<0.05.
Results
Characterization of fetal BMSCs, DBM, and BMSC/DBM constructs
Rabbit fetal BMSCs showed fibroblastic morphology in the tissue culture plates (Fig. 2A). Cells at passage 3 were induced to differentiate in the osteogenic media for 10 days. Positive alizarin red (Fig. 2B) and von Kossa staining (Fig. 2C) were observed following the osteogenic induction. The multipotent differentiation capacity of the fetal BMSCs was confirmed by chondrogenic and adipogenic induction (data not shown).

Characterization of fetal BMSCs and BMSC/DBM constructs.
To fabricate BMSC/DBM constructs, fetal BMSCs at passage 3 were seeded on DBM scaffolds and cultured in the osteogenic media for 10 days. SEM imaging showed the interconnected porous structure of the DBM before cell loading and that the scaffolds were fully covered by cells and cell-secreted matrices after cell seeding (Fig. 2D, E). The FDA/PI staining showed that in the middle part of the BMSC/DBM construct, most cells remained viable inside the scaffolds with positive staining of FDA and negative staining of PI (Fig. 2F).
Characterization of OVX rabbit model
All the animals survived from OVX or sham surgery. During the following 12 months, neither a sign of infection nor other surgical complications were found. Three animals from each group were sacrificed and the distal radius and ulna were harvested for micro-CT measurement. As shown in Figure 3A and B, the trabecular bone structure in the OVX rabbits became looser and thinner than those in the animals before surgery and sham surgery. Statistical analyses showed that bone volume/tissue volume ratio, bone volume, trabecular number, and trabecular separation in the OVX animals were significantly (p<0.05) lower than those in the animals before surgery. However, no significant changes were observed in the sham surgery group. These data confirmed the osteoporotic status of animals after OVX. The middle region of radius was also analyzed, but no significant changes were observed in the bone volume and BV/TV ratio between the OVX and sham surgery group (data not shown).

Characterization of OVX rabbit model.
Repair of critical-sized bone defects
Critical-sized bone defects were created in the radius of all OVX and non-OVX age-matched rabbits. The defects were repaired with BMSC/DBM constructs or DBM scaffolds alone. Bone regeneration was followed up in all animals until 3 months postoperation. Plain X-ray examination at the first month and third month showed that dense tissues were formed in both OVX and sham-surgery (non-OVX) groups repaired by BMSC/DBM constructs, but not in the group repaired by DBM alone (Fig. 4). Moreover, a better bone union was observed in the non-OVX group compared to the OVX group (Fig. 4). Animals were sacrificed at 3 months post-transplantation and the radius and ulna were harvested. Samples were first analyzed by micro-CT. As shown in Figure 5A, bone defects were repaired by high-density tissues in both OVX and non-OVX groups treated with BMSC/DBM constructs, whereas the nonunion remained in the DBM-treated group. To better demonstrate the bony regeneration, soft tissues in the defect area were removed and samples were photographed. A gross observation confirmed that the defects were repaired by bony tissues in the groups treated with BMSC/DBM constructs, but no bony tissues were observed in the defects treated with DBM alone (Fig. 5B). Quantitative micro-CT analysis showed that in the BMSC/DBM construct-treated groups, the bone density and bone volume of regenerated bone in the OVX group were significantly (p<0.05) lower than those in the non-OVX group, whereas no significant difference was observed in Tb.N between these two groups (Fig. 5C). The OVX group treated with DBM scaffold alone led to the nonunion healing with the lowest bone density, bone volume, and Tb.N compared to the other two groups (Fig. 5C).

Plain X-ray images of the radial defects at 1 and 3 months after transplantation with BMSC/DBM constructs or DBM alone in OVX and non-OVX animals.

Repair of critical-sized radial bone defects 3 months after transplantation.
Histological analyses
To further confirm the gross views and micro-CT findings, the histology of repaired defects at 3 months post-transplantation was examined using H&E staining and Masson's Trichrome staining of decalcified tissue sections. Representative photographs from each group are present in Figure 6A and B. In the defects treated with BMSC/DBM constructs, an irregular osteon formation was observed together with some undegraded DBM particles. In addition, a bony union was observed in the groups treated with BMSC/DBM constructs in either OVX animals or non-OVX animals. However, in the defects treated with DBM alone, only fibrous tissue with undegraded DBM particles was observed (Fig. 6A). Interestingly, many adipocytes were observed in the BMSC/DBM of OVX rabbit group, but not in the other two groups (Fig. 6A). Masson's Trichrome stains collagen fibers blue. The DBM scaffold, which contained collagen, was also stained blue. However, bone lacunas, which represented a newly formed bone, were only observed in the cell-seeded groups, while absent in the DBM-alone group (Fig. 6B).
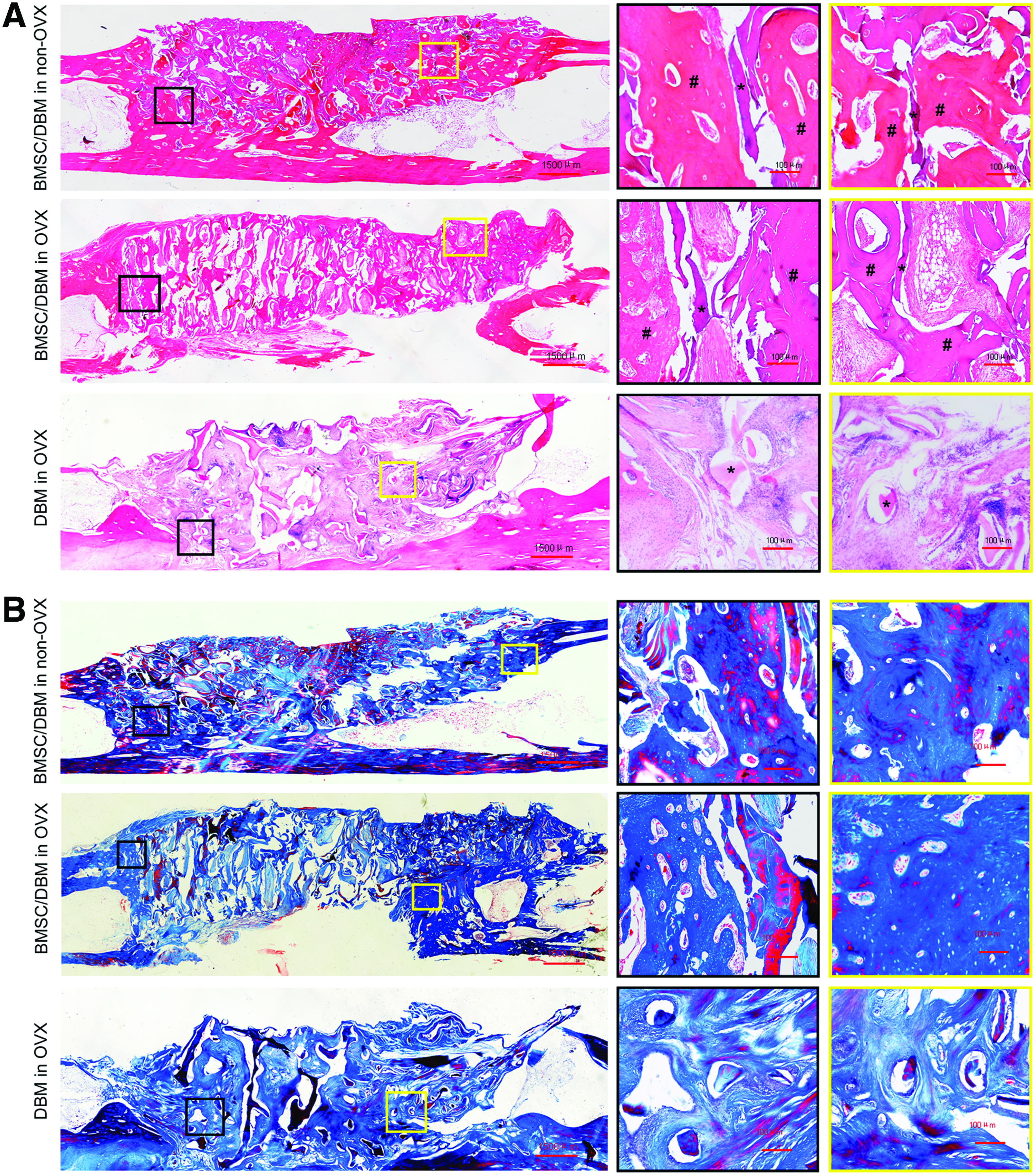
Histological analyses of regenerated tissues.
TRAP staining
By X-ray examination, micro-CT analyses, and histological observation, we demonstrated that the critical-sized bone defect could be repaired by BMSC/DBM constructs in both OVX and non-OVX animals. However, the therapeutic efficacy was compromised in the OVX animals (Fig. 5). To find out the possible mechanism, TRAP staining was performed to detect the activity of osteoclasts in the defect areas. More TRAP-positive osteoclasts were observed in the OVX group treated either with DBM or BMSC/DBM construct than in the non-OVX group, which had a very few number of osteoclasts (Fig. 7A, B).

TRAP staining of regenerated tissues.
Discussion
The repair of critical-sized bone defects using TE approaches in normal and disease-free animals has been previously shown to be effective.23–25 However, in clinical practice, the delayed union or nonunion of fractures occurs more commonly in elderly patients or those suffering from pathological conditions such as diabetes mellitus, osteoporosis, and so on. It is thus critical to address the need of TE bone repair in such pathological conditions.26–28 This study demonstrated the feasibility of repairing critical-sized radius bone defects with allogenic fetal BMSCs in osteoporotic animals of OVX rabbit model.
Primary osteoporosis is generally categorized into two types, namely senile and postmenopausal osteoporosis. A significantly decreased estrogen level is the major cause of postmenopausal osteoporosis. Consequently, in line with recommendations from the American Food and Drug Administration, bilateral OVX is an established method to reduce the estrogen level and establish an osteoporotic model. 29 As rabbits possess Harversian systems within their cortical bone similar to that of human beings, they become an ideal experimental model for the study of osteoporosis.30,31 We confirmed the successful establishment of a rabbit osteoporosis model by bilateral OVX with a significant decrease in both bone mass and BV/TV ratio and Tb.N at the distal radius at 12 months postprocedure (Fig. 3).
Stem cells play an essential role in tissue regeneration. Research shows that BMSCs decline in their number with age 32 and that the osteogenic differentiation potential of these cells is impaired in osteoporotic patients. 12 We found that it was difficult to obtain a large amount of BMSCs from OVX rabbits due to their poor proliferation capacity in culture (data not shown), thus finding that an appropriate cell source is essential for the treatment of such kind of bone fractures. Our previous study as well as others proved that allogenic MSCs were suitable for repairing bone defects in large animals.33–35 Fetal BMSCs have the advantages of higher proliferative and osteogenic capacities and exhibit lower immunogenicity, making them a suitable candidate cell source for allogenic bone repair.15–19 In this study, we used rabbit fetal BMSC-engineered bone to repair the radial critical-sized defect in OVX and non-OVX rabbits. In this study, the repair of created bone defects with allogenic fetal BMSCs plus DBM was proved effective in both OVX and non-OVX rabbit. By contrast, the implantation of DMB scaffold alone was unable to repair the defects to the same extent (Figs. 4 and 5), confirming the crucial role of stem cells in bone tissue regeneration. Many studies, including ours, have unveiled the successful repair of bone defects using allogenic MSCs.33–35 However, there remains a concern about the immune rejection of implanted cells, which may affect the healing efficiency in osteoporotic patients. Further comparison of autologous BMSCs (freeze preserved before OVX) versus allogenic fetal BMSCs in this model could address this concern.
A discrepancy exists among the research literature regarding the responses of osteoporotic animal models to biomaterial-mediated bone formation. Some studies demonstrated an impaired bone regeneration in osteoporosis,36,37 while others showed no difference between osteoporosis models and disease-free animals.38,39 This is likely due to the differences in animal models and experimental designs in these reported studies. In the current study, we found decreased bone mass and bone density of regenerated bone in OVX animals, as opposed to those in non-OVX controls (Fig. 5). TRAP staining proved that active osteoclasts contributed to the compromised bone regeneration in the OVX animals (Fig. 7). Inhibition of osteogenic differentiation of MSCs may also be involved in reduced bone regeneration in osteoporotic animals.11,12 The exact role of allergenic cells in engineered bone repair remains unclear. Previous studies indicated that implanted cells might not only directly differentiate to form a new bone but also recruit native stem cells to the defect site and thus promote bony regeneration through paracrine secretion. 40 Since the amount of native MSCs is decreased in osteoporotic conditions, it is possible that fewer host stems cells are recruited to the defect site, explaining the difference observed in the radial repair between the OVX and non-OVX rabbit models in this study. Unfortunately, cells were not labeled before transplantation; it is difficult to determine the percentage of new bones generated by transplanted cells or by host stem cells.
In addition, the long-term fate of transplanted allogenic cells is also unidentified. It is certainly worth performing cell-tracing studies in the future. Another interesting finding is that many adipocytes were observed in the BMSC/DBM-mediated bone formation in the OVX rabbits group, but not the other two groups (Fig. 6A), suggesting that transplanted BMSCs may differentiate into adipocytes instead of osteoblasts, which may also need to be confirmed with cell fate-tracing study. Collectively, the current results indicate that the tissue environment plays a significant role in engineered bone regeneration, and correcting an osteoporotic condition is essential to achieve the therapeutic effect in these patients.
In clinical practice, the prevention of fracture by enhancing a patient's bone density is as important as the promotion of postfracture bone regeneration in osteoporotic patients. By injecting autologous BMSCs or adipose-derived stem cells into local sites of osteoporotic models, bone mass and bone density at the injected site have shown to be enhanced.5,6,41,42 Our study demonstrated the potential of allogenic fetal BMSCs in bone regeneration. Future studies should investigate the potential of local injection of these cells in the treatment of osteoporosis.
Conclusion
This study demonstrated that the allogenic fetal BMSC-based TE strategy possesses a promising capacity in healing critical-sized bone defects in osteoporotic animals, although its therapeutic efficacy could be considerably compromised by the osteoporotic pathological condition. To develop more effective TE therapy for bone defect treatment under an osteoporotic condition, the adverse influence of pathological factor should be carefully addressed.
Footnotes
Acknowledgments
This work was supported by the Major State Basic Research Development Program of China (2011CB964704), the National Basic Research Program of China (30800231, 31170944) to W.J.Z., the National Natural Science Foundation of China (81101353, 81371964), the Program for Professor of Special Appointment (Eastern Scholar) at Shanghai Institutions of Higher Learning (No.1220000187), National Young Thousand-Talent Scheme, Shanghai Rising-Star Program (13QA1402400), and the Shanghai Jiao Tong University Medicine-Engineering Integrated Research Grant (YG2012MS44) to Z.-Y.Z.
Disclosure Statement
No competing financial interests exist.