
Abstract
Transplantation of encapsulated islets in a bioartificial pancreas is a promising alternative to free islet cell therapy to avoid immunosuppressive regimens. However, hypoxia, which can induce a rapid loss of islets, is a major limiting factor. The efficiency of oxygen delivery in an in vitro model of bioartificial pancreas involving hypoxia and confined conditions has never been investigated. Oxygen carriers such as perfluorocarbons and hemoglobin might improve oxygenation. To verify this hypothesis, this study aimed to identify the best candidate of perfluorodecalin (PFD) or HEMOXCell® to reduce cellular hypoxia in a bioartificial pancreas in an in vitro model of encapsulation ex vivo. The survival, hypoxia, and inflammation markers and function of rat islets seeded at 600 islet equivalents (IEQ)/cm2 and under 2% pO2 were assessed in the presence of 50 μg/mL of HEMOXCell or 10% PFD with or without adenosine. Both PFD and HEMOXCell increased the cell viability and decreased markers of hypoxia (hypoxia-inducible factor mRNA and protein). In these culture conditions, adenosine had deleterious effects, including an increase in cyclooxygenase-2 and interleukin-6, in correlation with unregulated proinsulin release. Despite the effectiveness of PFD in decreasing hypoxia, no restoration of function was observed and only HEMOXCell had the capacity to restore insulin secretion to a normal level. Thus, it appeared that the decrease in cell hypoxia as well as the intrinsic superoxide dismutase activity of HEMOXCell were both mandatory to maintain islet function under hypoxia and confinement. In the context of islet encapsulation in a bioartificial pancreas, HEMOXCell is the candidate of choice for application in vivo.
Introduction
C
However, the main limitation of this technology is the lack of oxygen inside the devices. 5 The oxygen partial pressure (pO2), reflective of oxygen availability, in a macroencapsulation device is typically around 20 mmHg, 6 which is half of the pO2 in the islets in vascularized pancreas. 7 Islets are highly sensitive to hypoxia. 8 Because of adenosine triphosphate (ATP) depletion in hypoxia, islet function is impaired and islets are exposed to inflammatory factors and reactive oxygen species (ROS), 9 which is detrimental to the function of the bioartificial pancreas. The hypoxic condition is worsened by the increased islet density in the restricted space of the device.8,10 Furthermore, the islets consume oxygen to maintain their metabolism, leading to an even more oxygen-deprived environment. Hypoxia induces the breakdown of ATP, 11 which leads to loss of islet function and finally cell death. In a previous work, we showed that confinement in combination with hypoxia decreased the survival rate of islets in a model mimicking the bioartificial pancreatic environment. 9 In the same study, we showed that the islets suffered from hypoxia-induced inflammation and apoptosis.
Hypoxia and confinement are two major issues for bioartificial pancreas function. Confinement conditions and cell density in the device are dictated by the reversion of diabetes and cannot be modified. Only two options are feasible: to increase the size of the device (MAILPAN®, Defymed 12 ) or to increase the number of devices (ViaCyte®, TheraCyte 13 ). The strategy to overcome hypoxia has been to implant the device before implanting of islets to allow sufficient vascularization of the implantation site to increase oxygenation inside the device; however, the level of oxygenation remains low, resulting in poor cell survival and function. 8 To alleviate these hypoxia-driven issues, researchers have focused on developing artificial oxygen supplies in the device. The βair® device, developed by Beta-O2, was designed to provide exogenous oxygen to the islets. Oxygen is infused in one compartment of the device and can reach the islets in a second compartment through a membrane. In this device, the graft is functional for 10 months 14 ; however, because of the requirement for daily oxygen refills, this system is not capable of completely autonomous function. An example of a molecular strategy to tackle the hypoxia problem involved the use of poly(dimethylsiloxane)-encapsulated, solid calcium peroxide matrices to release oxygen. 15 In this approach, islets survived for 72 h under hypoxia. 15 Other options include the usage of photosynthetic compounds,16,17 hemoglobin, 18 or perfluorocarbons (PFCs)19,20 (Table 1).
PFD, perfluorodecalin; SOD, superoxide dismutase.
PFCs are inert compounds in which the hydrogen atoms are replaced by fluorine atoms. This unique feature provides PFCs with the particular ability to dissolve and transfer molecular oxygen.21,28 Gases are physically dissolved rather than chemically bound as in hemoglobin. The solubility of oxygen in perfluorodecalin (PFD) is ∼20 times higher than that in aqueous culture media, and the solubility of CO2 is 150 times higher. 7 Furthermore, the oxygen diffusivity in PFD is 2.5-fold higher than that in water or culture media.29,30 As a consequence, the pO2 in PFC emulsions is in equilibrium with the surrounding media.21,27,28,31,32 PFCs have been used in organ preservation and have been tested in islet cultures under normoxic conditions. Perflubron and PFD are both able to prevent hypoxia in the islet core during culture at atmospheric pO2. PFCs can increase the oxygen gradient around islets, which allows oxygen to penetrate more deeply into the organoid.10,33,34 Moreover, the biological inertness and safety of PFCs make them suitable candidate materials for clinical application. Previous studies have shown that the combined use of PFCs and adenosine in pancreas preservation and digestion improved the levels of ATP, resulting in improved islet survival.35–37 Adenosine is an important substrate for ATP production, and ATP is implicated in islet metabolism, including insulin secretion,38,39 and in protection against oxidative stress. 39
Hemoglobin, a natural oxygen carrier, comprises four globins (two alpha and two beta chains), which allow simultaneous binding of up to four molecules of oxygen. 22 HEMOXCell®, a marine extracellular hemoglobin, presents highly interesting characteristics. It comprises 156 globins and thus can bind up to 156 oxygen molecules. p50, defined as the partial pressure corresponding to an oxygen saturation of 50%, is a measure of the oxygen affinity of hemoglobin. Hemoglobin works as a sponge in a simple oxygen gradient between the pO2 and the p50 of the molecule; if the p50 is below the pO2, the molecule will release oxygen, and each free site will be immediately occupied by an oxygen molecule drawn from the environment. This property implies that there is no operating time, just the necessity to respect the gradient. The p50 of HEMOXCell is around 37 mmHg at 37°C. 40 Moreover, this hemoglobin has an intrinsic superoxide dismutase (SOD) activity, which could be highly beneficial for cells in a hypoxic environment. This molecule has been shown to improve cell viability and proliferation 26 ; however, the oxygenation capacity of HEMOXCell in islets under hypoxia has never been tested.
The aim of the present work was to compare the efficiency of two oxygen carriers, PFD and HEMOXCell, supplemented or not with adenosine, in overcoming hypoxia in conditions that mimic the bioartificial pancreas.
Materials and Methods
Oxygen carriers
HEMOXCell
HEMOXCell, a product created using the extracellular hemoglobin M201, was supplied by Hemarina (Morlaix, France).
Perfluorodecalin
PFD (C10F18) (F2 Chemicals Ltd., Lancashire, United Kingdom) is a polycyclic perfluoroalkane. It is a derivative of decalin, with all of the hydrogen atoms replaced by fluorine atoms. Its density (1.92 g/cm3) and viscosity (2.61 cS) 41 are higher than those of water. This molecule is highly stable, biologically inert, and easily eliminated by the body. PFD can contain high concentrations of oxygen, which is released when pO2 decreases in the environment. 42
Islet cell isolation and culture
Animals
Male Wistar rats were supplied by Janvier laboratory (Le Genes St Isle, France). All rats were housed in pathogen-free conditions in standard collective cages in a temperature-controlled room (23°C ± 1°C) with a 12-h light/12-h dark cycle. They had ad libitum access to SAFE-A04 (SAFE, Villemoisson-sur-Orge, France) and water. All experiments were performed according to the National Institutes of Health and local ethics committee (CREMEAS) guidelines (Authorization No. C67-482-28).
Islet isolation
Pancreatic islets were isolated from adult Wistar rat pancreases (200–250 g) using standard collagenase (Sigma-Aldrich, St. Louis, MO) digestion and Ficoll (Eurobio, Les Ulis, France) purification. Islet preparations (from each pancreas separately) were cultured in medium M199 containing 5.5 mM glucose (Gibco, Life Technologies, Paisley, Scotland) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich) and 1% antibiotic/antimycotic (ABAM; 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of Fungizone; Gibco). After culturing for 24 h in 25-cm2 flasks (CELLSTAR®, Greiner Bio-One GmbH, Frickenhausen, Germany), each islet preparation was seeded in 48-well plates (Greiner) at a confined density of 600 islet equivalents (IEQ)/cm2. The culture medium was supplemented with 10% PFD or 50 μg/mL HEMOXCell. These two concentrations were determined based on a cell viability assay on rat β cell lines using concentration gradients of HEMOXCell (25 μg/mL, 50 μg, 75 μg, and 100 μg/mL) and PFD (1%, 5%, 10%, and 20% v/v). As no different effects were observed for the various concentrations, 10% of PFC, the quantity required to cover the surface of the well and known to be effective, 10 and 50 μg/mL, as recommended by the supplier, were used in subsequent experiments. Adenosine (ADE; Sigma-Aldrich) was added at the concentration of 1 mM to PFD and HEMOXCell. Media were prepared and saturated with pure oxygen for 30 min before culture with PFD. To ensure the homogeneity of the preparation, the medium with HEMOXCell was saturated as well despite its complete saturation level at a partial pressure of 150 mmHg.
To establish the ex vivo model, we used a seeding density of 600 IEQ/cm2 for a 24-h culture under hypoxia (pO2, 15 mmHg: 2% O2, 5% CO2, 93% N2; Lindegas, Saint-Priest, France) in a humidified incubator (StemCell Technologies, Vancouver, Canada) at 37°C, as previously reported 9 ; this was considered the control condition. Oxygen measurements were carried out with a four-channel fiber-optic oxygen meter (Oxy-4) and noninvasive oxygen sensors (PreSens GmbH, Regensburg, Germany) glued onto the bottom of each culture well, allowing noninvasive measurements from outside, through the plastic wall. An optic fiber connected to the OXY-4 oxygen meter was guided and positioned from outside the chamber to the sensor. The data are expressed in mmHg and were taken at 0 and 24 h of hypoxic incubation.
Islet viability assay
The viability of 10 islets per treatment condition (islets seeded at 600 IEQ/cm2) under hypoxia (pO2 15 mmHg) in the presence of PFD (10%), ADE (1 mM), PFD (10%)+adenosine (1 mM), HEMOXCell (50 μg/mL), or HEMOXCell (50 μg/mL)+adenosine (1 mM) was analyzed using a LIVE/DEAD® viability/cytotoxicity assay kit (Thermo Fisher Scientific, Illkirch-Graffenstaden, France) following the manufacturer's instructions. Polyanionic dye, calcein AM, is metabolized by living cells and fluoresces in green and EthD-1 penetrates in necrotic cells and labels nuclei in fluorescent red. The empirical ratio between numbers of green cells and EthD-positive nuclei cells indicated the percentage viability. 43 Results are represented as the mean ± SEM. Images were obtained on a Nikon Eclipse 50i microscope with Nis-Element-BR software (Nikon, Amstelveen, The Netherlands).
Proinsulin quantification
Unregulated release of hormones reflects cell death activity. Islets were cultured for 24 h and supernatants were collected and stored at −20°C. Proinsulin was quantified using the Mercodia Proinsulin kit according to the manufacturer's protocol. Results were expressed as proinsulin secreted (pmol/) per 600 IEQ/cm2.
Islet functionality assay
A portion of the islets (n = 10) from each experimental condition was washed extensively and incubated in Krebs Ringer bicarbonate (KRB) solution with 10% FBS and 2.2 mM of glucose (Sigma-Aldrich) for 90 min at 37°C in a humidified 5% CO2 atmosphere. Then, the islets were stimulated with KRB solution containing 10% FBS and 22.6 mM of glucose for 90 min at 37°C in a humidified 5% CO2 atmosphere. 44 Supernatants were collected and stored at −80°C. Insulin measurements were performed using a rat insulin ELISA kit (Mercodia). Insulin content was normalized to the total amount of protein extracted for each condition. Insulin content was expressed as μg/g of protein and as a stimulation index (SI), defined as the ratio of stimulated versus basal insulin secretion. An islet preparation was considered functional when SI >1.
Real-time PCR
Total cellular RNA was extracted from ∼400 islets for each condition by using a Qiagen RNeasy Mini Kit (Qiagen, CA) following the manufacturer's instructions. The RNA was reverse transcribed with an RT 2 First Strand Kit (Qiagen) according to the manufacturer's instructions. The cDNA was diluted 1:100 in RNase-free water and amplified for 35 cycles on the MyiQ real-time PCR system (Qiagen) using QuantiTect SYBR Green PCR kits (Qiagen). Mouse 60S acidic ribosomal protein P1, mouse peptidylprolyl isomerase A, and mouse RNA ribosomal 1 were chosen as the housekeeping genes, and data were analyzed using the ΔΔCt method. The primers for hypoxia-inducible factor 1 alpha (HIF-1α) and housekeeping genes used in this study were purchased from Qiagen and are listed in Table 2.
Protein extraction
Total protein was extracted from 300 islets for each condition using M-PER Mammalian Protein Extraction Reagents, HALT™ Protease, and a phosphatase inhibitor cocktail (Thermo Fisher Scientific). The protein concentration was determined using the Bradford micromethod (Bio-Rad Life Science Group, Marnes-la-Coquette, France) and expressed as mg/mL.
HIF-1α activation (ELISA)
HIf-1α activation was quantified using 20 μg of total protein extract with TransAM® HIF-1 (Active Motif, Belgium) following the manufacturer's instructions. HIF dimers, which bind specifically to the hypoxia response element, were immobilized in the 96-well plate. HIF dimers were detected with HIF-1α antibody (dilution 1:500) and a secondary antibody conjugated to horseadish peroxidase (HRP) (dilution 1:1000). The results are expressed in optical density.
Western blotting
Equal amounts of protein (15 μg) from each sample were mixed with sample buffer, separated by electrophoresis on polyacrylamide gels (4–12% Criterion™ XT Bis–Tris; Bio-Rad), and wet-transferred onto PVDF membranes. The membranes were probed with primary antibodies (4°C, overnight) against cyclooxygenase-2 (COX-2; source rabbit, dilution 1:1000; Abcam, Paris, France), NF-κB p65 (source rabbit, 1:500; Cell Signaling), phospho-NF-κB p65 (source mouse, 1:500; Cell Signaling), and β-actin (source rabbit, 1:2000 [Abcam]; source mouse, 1:2000 [Santa Cruz, Dallas, TX]) as a loading control. Secondary antibodies (1:5000, anti-mouse-HRP, or 1:5000, anti-rabbit-HRP; Sigma-Aldrich) were incubated for 1 h at room temperature. Blots were developed with the Immun-Star™ WesternC™ kit (Bio-Rad) and recorded with Chemidoc® (Bio-Rad). Densitometry was carried out with ImageJ software (National Institutes of Health, Bethesda, MD; http://imagej.nih.gov/ij/). The results are expressed relative to β-actin.
Quantification of IL-6 and VEGF secretion
Vascular endothelial growth factor (VEGF) and interleukin (IL)-6 were quantified in islet supernatants using a rat VEGF ELISA kit (Tebu-bio, Le Perray-en-Yvelines, France) or IL-6 (R&D Systems, Minneapolis, MN). Results are expressed as pg/mL.
Statistical analyses
Statistical analyses were performed using the Statistica software (StatSoft, Maisons-Alfort, France). The results were analyzed with one-way analysis of variance (ANOVA), followed by Tukey's honest significant difference post hoc test for parametric data or by a Kruskal–Wallis test, followed by a Mann–Whitney U test for nonparametric data. The results are expressed as medians and interquartile ranges. A p-value of less than 0.05 was considered statistically significant.
Results
Islet viability
The islet viability did not significantly improve with the addition of PFD (b) or adenosine (ADE) (d) alone, PFD+ADE (e), or HEMOXCell+ADE (f) compared with 600 IEQ/cm2 under hypoxia and confinement without additional molecules (a; control condition) (Fig. 1A, B). Nevertheless, the addition of HEMOXCell (b) improved islet viability significantly (p < 0.05), as indicated by a decrease in dead islets compared with the control (a; Fig. 1A).
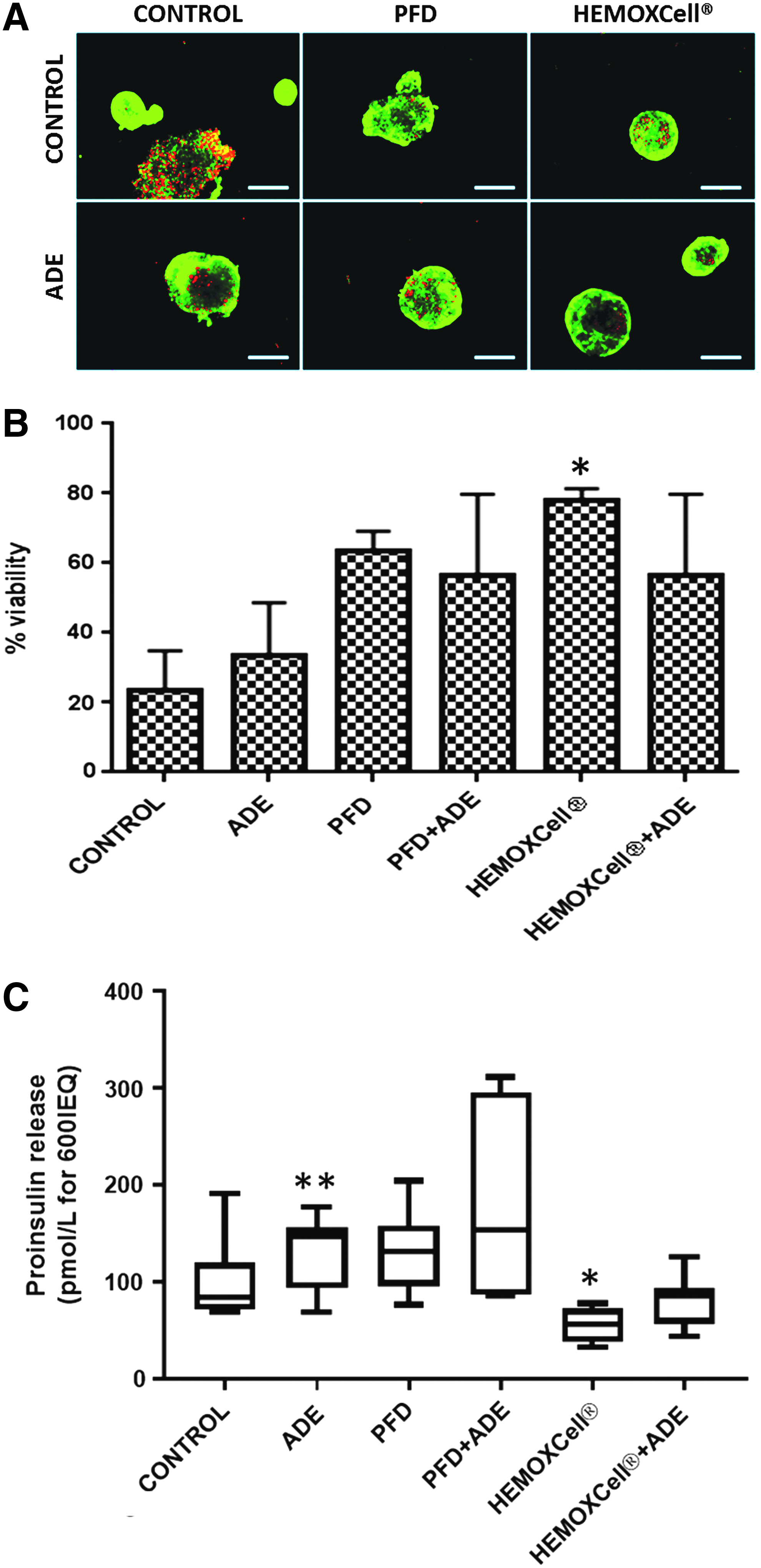
Effect of oxygen carriers on the viability of islets cultured under hypoxia and confinement.
Unregulated proinsulin release into the culture medium was used as a marker of islet cell membrane disruption and cellular destruction. The concentration of proinsulin significantly increased with the addition of ADE in the medium (Fig. 1C, p < 0.01, n = 9). The addition of HEMOXCell during culture decreased proinsulin release compared with the control (p < 0.05, n = 9).
Hypoxia marker expression
HIF-1α mRNA expression (Fig. 2A) and HIF-1α protein activation (Fig. 2B) significantly decreased (p < 0.05) with all treatments. This effect was more prominent in the PFD+ADE and HEMOXCell±ADE treatment groups for both mRNA expression and protein activation. Regarding VEGF secretion in the culture medium, no differences were observed between PFD and/or ADE and the control. Only HEMOXCell±ADE increased the secretion of VEGF significantly (Fig. 2C; p < 0.001).
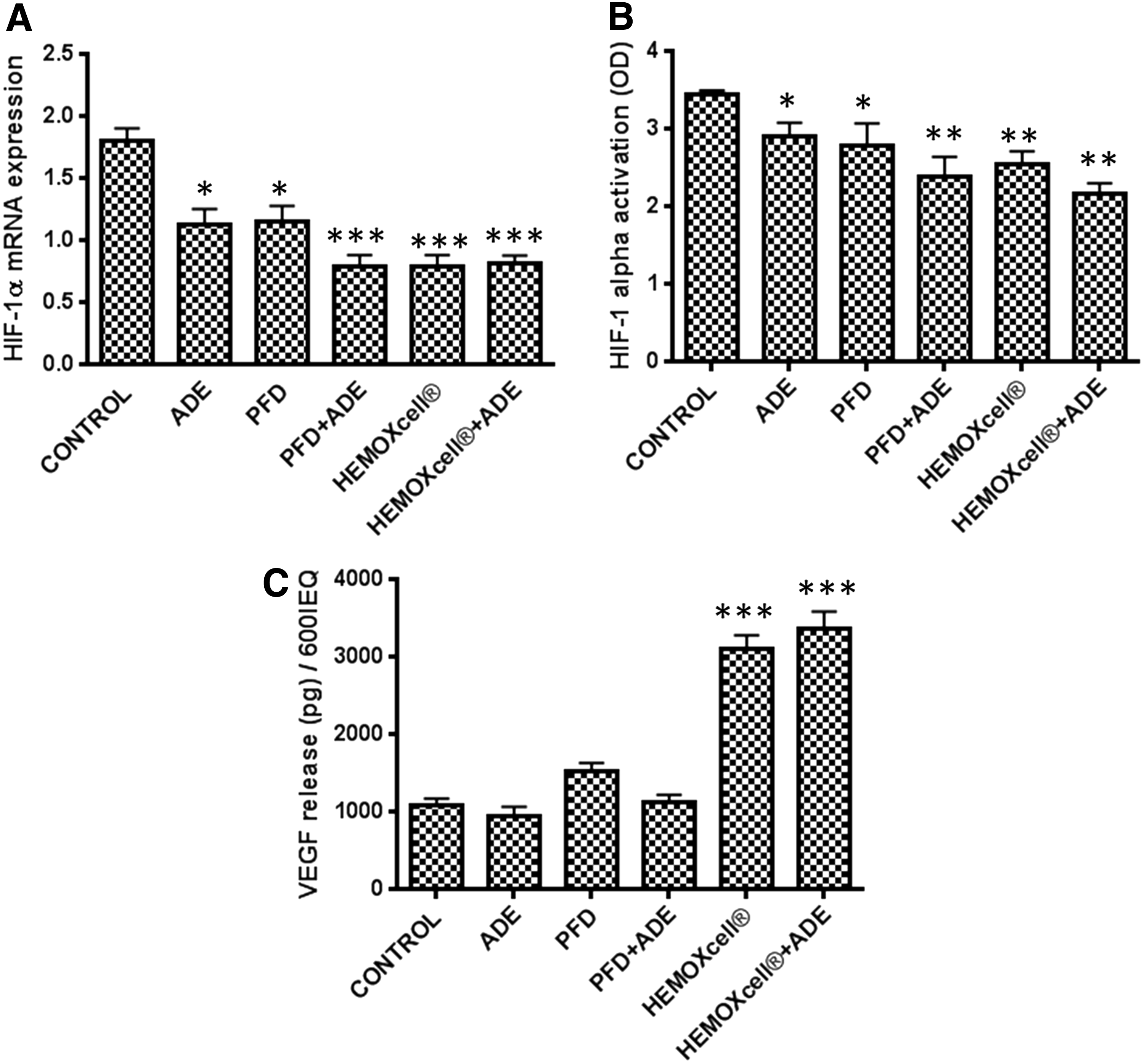
Effect of oxygen carriers on the expression of HIF-1α and VEGF in islets cultured under hypoxia and confinement. Levels of HIF-1α mRNA expression
Inflammation
To evaluate the effects of decreased hypoxia by using oxygen carriers under confinement on the inflammatory response, we measured the relative protein expression levels of inflammatory markers, NF-κB, phospho-NF-κB, and COX-2, as well as the secretion of IL-6. While the treatments had no significant effects on NF-κB phosphorylation (Fig. 3A), the effects on COX-2 expression showed a similar pattern, but were significant. PFD alone had no impact on inflammation markers. Adenosine alone or in combination with PFD increased the level of COX-2 significantly compared with the control (Fig. 3B; p < 0.05; p < 0.001). Independent of the presence of adenosine, HEMOXCell significantly increased the level of COX-2 (p < 0.05). The secretion of IL-6 was increased in the presence of adenosine or PFD+ADE (p < 0.05), whereas HEMOXCell decreased IL-6 secretion (p < 0.05) (Fig. 3C).
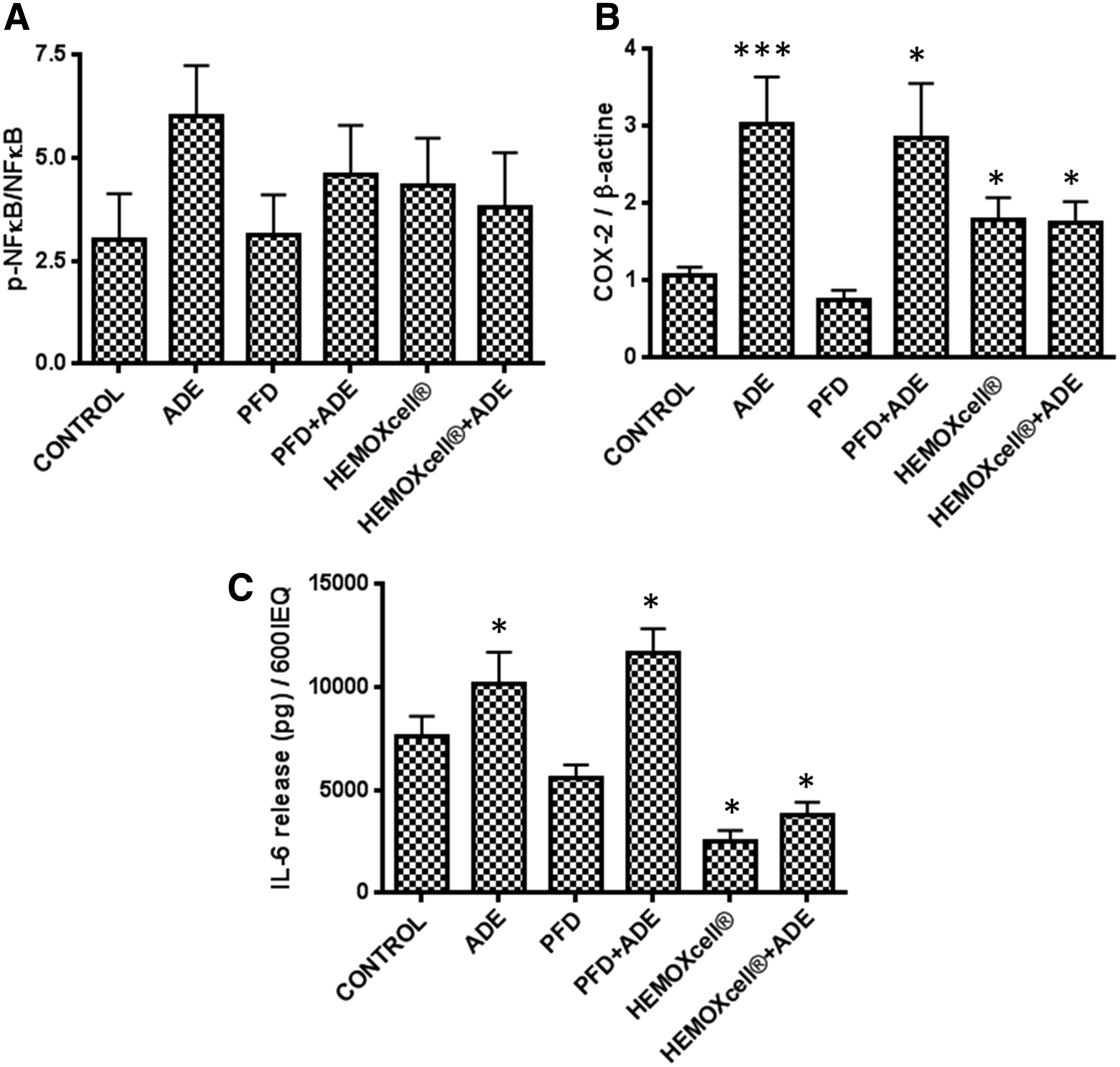
Effect of oxygen carriers on inflammation in hypoxic and confined islets. Protein expression levels of phospho-NF-κB/NF-κB
Islet functionality
In the control condition, islet functionality was lost. High-glucose concentrations did not increase insulin secretion, as reflected by the SIs below 1, indicating an absence of insulin response. The addition of PFD, adenosine, or both did not improve the response to glucose (Fig. 4A). Insulin secretion was independent of the presence of glucose (p < 0.05, p < 0.001), and the SIs were below 1 (Fig. 4B). The addition of HEMOXCell to the medium significantly improved the response to glucose stimulation independently of adenosine presence (p < 0.001), as reflected by SIs between 2 and 4 (p < 0.05). Figure 4B shows that five islet preparations of seven were functional (IS ≤2) in the presence of HEMOXCell and five of eight in the presence of HEMOXCell+ADE, while no functionality was observed in the other conditions.

Effect of oxygen carriers on islet function in hypoxia and confinement.
Discussion
In the present study, we showed that the addition of PFD or HEMOXCell with or without adenosine decreased hypoxia. However, inflammation was not reduced by PFD or adenosine, which likely led to the inability to restore islet function. On the other hand, the use of HEMOXCell decreased inflammation and restored islet viability and function under confinement and hypoxia.
PFCs 10 and hemoglobin18,45 have been shown to effectively improve islet isolation, transplantation, and encapsulation.33,46 However, under hypoxia, the effects of the two compounds in hypoxic conditions appear to be different. The two compounds act differently in terms of oxygen release, which can explain the differences in their effects on islets. Both molecules, PFD and HEMOXCell, enhanced decreases in HIF-1α mRNA expression and protein levels, attesting to the positive effect of these compounds on hypoxia of islets cultured at the seeded density of 600IEQ under a PO2 of 15 mmHg. However, only HEMOXCell was able to reduce inflammation and restore function. The difference in action can be explained by the properties of the two compounds. The carrying capacity of PFD is directly proportional to the pO2. 47 Oxygen diffuses to reach equilibrium between PFD and the ambient atmosphere. Because the oxygen is not molecularly tied to the PFD, PFD acts as a reservoir for oxygen. In our conditions of hypoxic culture for 24 h, the pO2 was around 80% of saturation after bubbling and only 2.5% after the culture period (data not shown). The presence of PFD delayed the appearance of hypoxia—7 h after hypoxia induction, there was still 10% oxygen in the wells containing PFD versus 2.5% in the control wells—which explained the decrease in hypoxia markers.
In contrast, in hemoglobin, oxygen is bound to iron. The binding depends on the affinity of oxygen for the binding site, the pO2, and the p50 of hemoglobin. Unfortunately, because of this molecular binding, the oxygen available cannot be measured using an oximeter. The concentration of oxygen in the medium was around 2.5% after 24 h of hypoxic culture; however, the decrease in hypoxia markers proved that there was oxygen delivery to the cells. The p50 of hemoglobin is the critical parameter determining its effect in a hypoxic environment. For instance, when a hemoglobin molecule with a p50 of 26 mmHg is placed in an environment of 7 mmHg oxygen, only 10% of its oxygen binding sites are saturated. In myoglobins with a p50 of 2 mmHg, 78% of the binding sites will be saturated with oxygen at the same partial pressure.18,48 We chose hemoglobin with a p50 of around the partial pressure in bioartificial pancreas to compensate for the hypoxic environment. HEMOXCell has a p50 of 37 mmHg at 37°C, which means that at a pO2 below 37 mmHg, HEMOXCell releases oxygen against the gradient. The release follows a sigmoid pattern, which implies that hemoglobin buffers the pO2 between two limits. Thus, oxygen release depends on the partial pressure gradient, but within the limits of the specific hemoglobin. 49
Despite this difference, both PFD and HEMOXCell appeared to be able to decrease hypoxia markers. The advantage of HEMOXCell over PFD could be the intrinsic SOD activity, which eliminates ROS, thus decreasing inflammation. 50 This is consistent with a previous report on SOD activity in grafted hemoglobin. 48 ROS are known as major activators of inflammation and can trigger a detrimental pathway, leading to membrane breakdown and proinsulin release. We have shown in previous studies that hypoxic conditions combined with confinement triggered inflammatory pathways by inducing COX-2 mRNA expression. 9 In this study, we observed that the addition of PFD alone to the culture medium had no impact on inflammatory markers; however, the addition of adenosine enhanced inflammation by activating COX-2, NF-κB, and subsequently IL-6 secretion. Surprisingly, despite the activation of COX-2 in HEMOXCell treatment conditions, IL-6 secretion was reduced. Cytokine secretion is a major issue in islet transplantation and more so in solid organ transplantation as it can trigger graft rejection.51,52 The decrease in IL-6 secretion, most likely related to the SOD activity of HEMOXCell,26,50,53 can be expected to improve the tolerance of bioartificial pancreas in vivo. A similar discrepancy was observed regarding VEGF secretion; despite a decrease in HIF expression, VEGF was significantly increased in the presence of HEMOXCell. Contrary to PFC, hemoglobins interact with cells and are able to induce signaling pathways. For instance, it has been shown in macrophages that hemoglobin can activate the receptor CD163, which exerts anti-inflammatory effects by activating heme oxygenase 1.54–56 Thus, it is possible that the increase in VEGF and the decrease of IL-6 observed in the presence of HEMOXCell are independent from oxygenation, but are instead activated through other pathways.
In our results, the decrease in HIF-1α was not correlated with VEGF release. However, it is possible that the 24-h time point used in this study was too early to see any effects of hypoxia on VEGF secretion. In contrast, the increase in VEGF release by the addition of HEMOXCell without adenosine could be attributed to the addition of this oxygen carrier. In support of this hypothesis, some studies have suggested that hemoglobin enhances tissue factor expression, resulting in upregulation of VEGF expression and secretion.57,58
The decrease in hypoxia seems to be important for islets within the first 24 h to maintain islet mass. Indeed, when islets were assessed after 5 days under hypoxia, the ones that survived the first 24 h were functional, independent of the culture condition (data not shown). This observation seems to imply that when an islet survives the first hours of culture in suboptimal conditions, it develops defenses that maintain its survival and function in the long term. Therefore, the main objective should be to maintain islet mass. As shown by the increased proinsulin release, the islet number decreased drastically in some conditions with PFD or adenosine. The effects of adenosine are dual; it has been shown that adenosine can trigger cell death59,60 or cell survival, 61 depending on the type of receptor activated. Moreover, it has been shown that adenosine is unstable and is rapidly degraded to inosine and hypoxanthine, precursors of ROS. 62 In our conditions of hypoxia and confinement, adenosine at 5 mM was detrimental for islet survival. Moreover, the presence of antioxidant activity in HEMOXCell enhanced islet survival and function,63,64 as shown by the lack of such effects of PFD.
In conclusion, we showed that the supply of HEMOXCell without adenosine to confined islets restored islet glucose-sensing function only after 24 h of incubation under hypoxic conditions. Future studies should aim at validating the efficiency of HEMOXCell to preserve the viability and function of encapsulated islets in vivo and of long-term surrogate cells.
Footnotes
Acknowledgment
This work received financial support from the Cooperation Program of the European Community's FP7 (Grant No. HEALTH-F2-2012-305746).
Disclosure Statement
The authors declare that F.Z. is the founder of and holds stock in HEMARINA SA, which produced the substance under investigation. V.P. is an employee of HEMARINA SA and does not hold stock.