
Abstract
The kidney peritubular microvasculature is highly susceptible to injury from drugs and toxins, often resulting in acute kidney injury and progressive chronic kidney disease. Little is known about the process of injury and regeneration of human kidney microvasculature, resulting from the lack of appropriate kidney microvascular models that can incorporate the proper cells, extracellular matrices (ECMs), and architectures needed to understand the response and contribution of individual vascular components in these processes. In this study, we present methods to recreate the human kidney ECM (kECM) microenvironment by fabricating kECM hydrogels derived from decellularized human kidney cortex. The majority of native matrix proteins, such as collagen-IV, laminin, and heparan sulfate proteoglycan, and their isoforms were preserved in similar proportions as found in normal kidneys. Human kidney peritubular microvascular endothelial cells (HKMECs) became more quiescent when cultured on this kECM gel compared with culture on collagen-I—assessed using phenotypic, genotypic, and functional assays; whereas human umbilical vein endothelial cells became stimulated on kECM gels. We demonstrate for the first time that human kidney cortex can form a hydrogel suitable for use in flow-directed microphysiological systems. Our findings strongly suggest that selecting the proper ECM is a critical consideration in the development of vascularized organs on a chip and carries important implications for tissue engineering of all vascularized organs.
Introduction
T
Recent studies have reported that peritubular vessels are highly susceptible to injury, which contributes significantly to tubular dysfunction, fibrosis, and development of acute kidney injury and chronic kidney disease (CKD).3–7 However, little is known about this microvasculature due to difficulties microvascular system imaging in vivo and the lack of appropriate models in vitro. Engineering a representative human kidney microvasculature with proper cells, extracellular matrices (ECMs) and vessel geometries presents a prospective tool to greatly improve our understanding of their structure and function.
Recently, our laboratory developed methods to isolate Human kidney peritubular microvascular endothelial cells (HKMECs) and engineer a 3D microvascular system within a collagen-I hydrogel. 8 Although collagen-I provides robust mechanical support for vessel fabrication and is an important native ECM component in the kidney, increased collagen-I production has been correlated with fibrosclerosis, a common hallmark in the development of CKD. 9
Furthermore, recent studies have shown that ECM components are more than structural scaffolds that maintain mechanical support for cell and tissue organization—rather, they provide integrin-dependent signal transduction.10–12 Collagen-I has been found to direct vascular, tubular epithelial and mesangial cells toward fibrogenic phenotypes with enhanced collagen synthesis, increased endothelial activation, or epithelial-to-mesenchymal transition.13,14 Thus, there exists a need for substrates that can better simulate the native environment of the human kidney and improve our capacity for studying the complex interactions that occur within this vital organ.
Native human kidney contains a plethora of ECM proteins, whose composition and function vary based on the anatomical area, developmental stage, or pathological state of the kidneys.15,16 Proteins such as collagen-IV (col-IV) and laminins (LAMs) form an intertwining basal lamina for tubules, peritubular microvessels and glomeruli.15,17
In particular, col-IV is present as a triple helical molecule having three α chains coiled around one another, with six distinct chains (COL4A1–COL4A6) available to form unique isoforms. COL4A1 and A2 chains are widely expressed in basement membranes of the kidney, but nearly undetected in mature glomerular basement membrane (GBM), which only expresses the A3, A4, and A5 chains. 15 Laminins are heterotrimeric glycoproteins comprising one α, one β, and one γ chain in a cruciform organization. Five α chains (LAMA1–LAMA5), three β chains (LAMB1–LAMB3), and three γ chains (LAMC1–LAMC3) have been distinguished within the kidney. 16
These basement membrane proteins further bind other glycoproteins and proteoglycans and interact with integrin-type cell receptors and growth factors to regulate cell adhesion, spatial organization, growth, proliferation, and additional complex cellular behaviors.16,18 Moreover, holistically, the basement membranes—together with the vascular and tubular cells—serve as a charge- and size-selective filtration and reabsorption barrier between the circulation and the urinary space.
Substrates comprising individual proteins such as laminin and col-IV have been shown to support kidney epithelial cell phenotypes and prevent their epithelial–mesenchymal transition in vitro. 13 These individual basement membrane proteins, however, lack sufficient structural and mechanical integrity to support complex 3D culture systems—including microvascular and tubular architectures. In addition, native kidney matrices comprise rich compositions of many proteins that have been found to be important for cellular function. Creating native kidney ECM (kECM) substrates from whole kidney tissues presents a promising approach to support tissue engineering and regenerative medicine investigation for human kidney models.
Recently, decellularized whole tissue ECM has become a popular source for tissue engineering substrates in an attempt to maintain endogenous cellular function by utilizing an environment that better recapitulates the complexity of cellular cues found in vivo, with proper matrix composition, mechanical support and biochemical microenvironment. Researchers have decellularized ECM from a number of different organs to create bioengineered matrices for regenerative medicine, including muscle, 19 nerve,20,21 bladder, 22 intestine, 23 heart, 24 lung,25,26 capillary beds,26,27 and liver. 28
Decellularized kidneys have been used as a substrate to create functional kidneys ex vivo by repopulating the whole kidney with different cells. 29 However, outstanding challenges to this approach exist, resulting from the large volume of cells required, as well as competing culture conditions for the many different cell types needed to recapitulate the kidney vascular and tubular structures. Furthermore, using such an approach encounters the same issues as in vivo models including limited imaging capacities and complex multicellular interactions, which prevent the fundamental understanding of the contributions and interactions of individual cellular and matrix components.
In this study, we address these challenges by fabricating and characterizing native human kECM gels from decellularized human kidney cortex tissue and assess their ability to support vascular stasis. We first characterize the decellularization process to demonstrate the removal of cellular components while preserving the biochemical availability of the ECM. We then compare the mechanical properties of kECM, collagen-I, and a mixture of kECM and collagen-I hydrogels, and investigate their capacity to support HKMECs compared with human umbilical vein endothelial cells (HUVECs). Finally, we demonstrate the ability to use a hydrogel comprising kECM to create 3D microvessels in an effort to better recapitulate the microvascular environment of the kidney.
Materials and Methods
Unless otherwise stated, all materials were purchased from Sigma-Aldrich. Expanded methods for cell culture, collagen fabrication can be found in Supplementary Materials and Methods (Supplementary Data are available online at www.liebertpub.com/tea). A list of PCR primers can be found in Supplementary Table S1.
Decellularization of human kidney
Human kidneys were received on ice from LifeCenter NorthWest, after which the renal capsule was removed and the underlying cortex was sectioned and immersed in 1% sodium dodecyl sulfate (SDS) in endotoxin-free cell culture grade water (Thermo Scientific) at pH 7.4 for 5 days with constant stirring (Fig. 1A). Tissue sections were then rinsed in water for 5 days, changing water twice daily, then lyophilized (Labconco), and stored at −20°C.
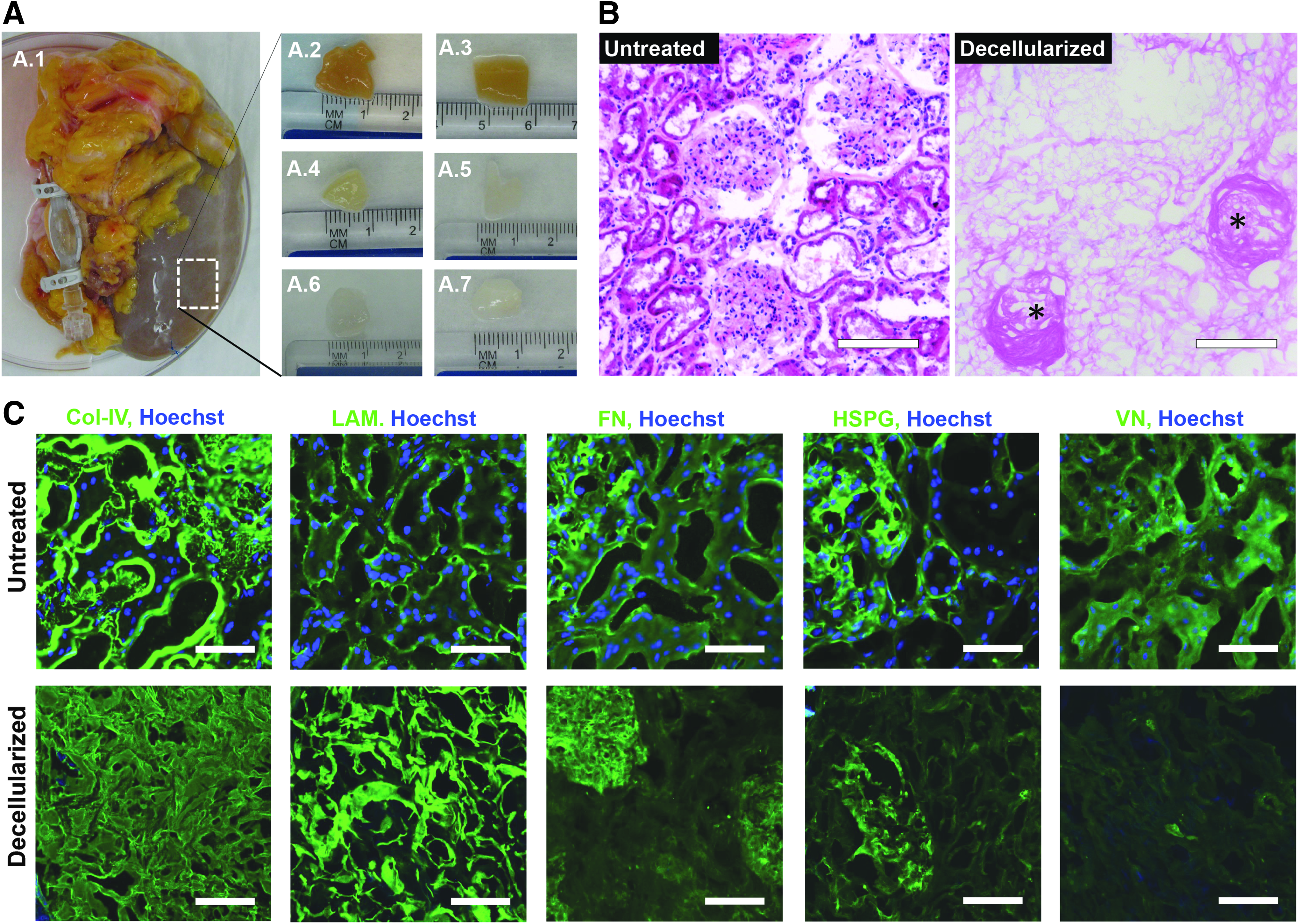
Decellularization and matrix characterization of human kidney cortex.
Characterization and relative quantification of protein composition by nano-liquid chromatography–tandem mass spectrometry
The total protein concentration was determined with a Bio-Rad protein assay. Kidney matrix proteins were solubilized with 1% sodium deoxycholate in 50 mM Tris (pH 8) and sonicated at 4°C for 20 min. The proteins were reduced with dithiothreitol, alkylated with iodoacetamide, and then digested with trypsin (1:10 wt/wt, trypsin/protein; Promega) in 50 mM Tris (pH 8) containing 5% acetonitrile at 37°C overnight. Digestion was halted by acidification with trifluoroacetic acid and the sodium deoxycholate was removed by liquid–liquid extraction into ethyl acetate.
Tryptic peptides were analyzed by liquid chromatography–tandem mass spectrometry (LC-MS/MS) with a Thermo Scientific LTQ OrbitrapVelos mass spectrometer coupled to a Waters nanoAcquity Ultra Performance LC system with a nanoUPLC BEH130 C18 column (100 × 0.075 mm, 1.7 μm; Waters). Peptides were eluted using a linear gradient of 5–35% acetonitrile with 0.1% formic acid over 90 min. MS/MS spectra were searched against the human protein database using Proteome Discoverer 1.3 software (Thermo Scientific). All protein identifications required detection of two unique peptides per protein. Relative protein abundance was determined by the average area of the top three most abundant unique peptides of the protein identified. Percent composition was evaluated by dividing the peak area of the protein of interest by the total area of all the proteins identified.
Gelation of decellularized human kECM and collagen-I
Lyophilized kECM was minced, then homogenized (Polytron; Brinkmann Instruments) in 0.01 N HCl on ice. Additional 0.01 N HCl was added to reach 15 mg/mL. Next, 30 mg of pepsin per 30 mL kECM solution was added to digest the sample for 48 h at room temperature with constant stirring. After digestion, the sample was stored at −20°C for long-term storage. Collagen-I was prepared from rat tails at 15 mg/mL in 0.1% acetic acid, as described previously. 8 Liquid collagen-I, kECM, and mixture gel solutions were prepared on ice by mixing the stock gel solutions with neutralizing diluents to reach the targeted concentrations.
Rheology test
Parallel plate rheology was performed to compare the complex modulus of the kECM, 1:1 mixture, and collagen-I hydrogels. All samples were measured at 37°C in oscillatory mode at a fixed frequency of 1 Hz with varying strain from 0.01% to 1000%. Complex moduli were calculated from storage (G′) and loss (G") moduli.
Histology and immunostaining
Whole and decellularized tissues, gelled kECM, and engineered microvessels were all fixed with 4% paraformaldehyde, then embedded in Tissue-Tek® O.C.T. compound, and sectioned using a Leica CM1850-3-1 cryostat. Following embedding and sectioning, samples were processed following standard immunohistochemical procedures.
Sections were blocked using 2% bovine serum albumin (BSA) for 1 h before the addition of primary antibodies against selected ECM components. Primary antibodies against col-IV (ab6586), fibronectin (FN; ab2413), heparan sulfate proteoglycan (HSPG; ab23418), LAM1 (ab11575), and versican (VN; ab19345) were added overnight at 4°C, then rinsed repeatedly before incubation in 2% BSA containing the appropriate fluorescently conjugated secondary antibodies (Life Technologies) for 1 h at room temperature. In all immunostaining experiments, secondary antibodies in 2% BSA were added without a primary antibody as a negative control.
For experiments using cultured cells, samples were permeabilized for 15 min in a 2% BSA/0.1% Triton X-100 solution before blocking with 2% BSA and subsequent addition of primary antibodies against CD31 (ab28364) and plasmalemma vesicle-associated protein (ab81719) and fluorescent conjugates, FITC-von Willebrand factor (VWF; ab8822), phalloidin 647 (A22287), Hoechst counterstain (H1399), and secondary antibodies.
Imaging and quantifications
Sections stained with hematoxylin and eosin were imaged with a Nikon TS-100 microscope and a Canon Vixia HFS20 camera. Immunofluorescence images were taken using a Nikon A1R confocal microscope. Image processing was performed using ImageJ software. Sprouts from endothelial monolayers were counted from confocal z-stacks. For cell density and sprouting frequency, three images were counted to reach an averaged value for each replicate. For both assays, three to five biological replicates were processed for analysis.
Cell culture
Primary HKMECs were cultured between passages 1 and 3 as described previously. 8 The culture media consisted of EBM-2 basal media (Lonza) with fetal bovine serum (10%), heparin (50 μg/mL), endothelial cell growth supplement (100 μg/mL), and vascular endothelial growth factor (VEGF, 20 ng/mL; R&D) supplements. HUVECs were acquired from Lonza and cultured between passages 3 and 7 according to the manufacturer's suggestions.
Engineered microvessels
Microvessels were fabricated from a 7.5 mg/mL kECM/collagen mixture as described previously. 30 Cells were delivered to the inlet of the channel using ∼8 × 104 cells/device. Gravity-driven flow was introduced by replenishing media in the inlet reservoir twice per day.
Real-time quantitative PCR analysis
Total RNA was isolated from cells cultured in a well of a six-well plate using an RNeasy mini kit (Qiagen) with on-column DNase digestion (Qiagen) to remove genomic DNA, then read on a NanoDrop 1000 Spectrophotometer. cDNA was used in a 10 μL reaction with Power SYBR® Green PCR Master Mix (Applied Biosystems) to determine the mRNA level of specific genes using β-actin as an internal control using a model 7900HT Real-Time PCR System (Applied Biosystems). Comparative CT was used to analyze the relative quantification of gene expression between the samples. Each set of oligonucleotides was designed to span two different exons to avoid PCR products from genomic DNA contamination.
Statistical analysis
For all quantitative measurements, the entire population was used to calculate statistical significance, whereas mean values with n = 3 were used to calculate standard error and graphical confidence intervals. Data were then analyzed using a one-way ANOVA test, followed by post hoc analysis using Tukey-HSD to determine significance between groups. On each graph, error bars represent ±2 SEM (standard error of the mean), a 95% confidence interval. A single asterisk was used for p-values <0.05, two asterisks for p < 0.01, and three asterisks for p < 0.001.
Results
Decellularization of human kidneys
Discarded human kidneys originally intended for transplantation were received for decellularization and kECM extraction (Fig. 1A.1). Human kidney cortex was diced into 1 cm × 1 cm × 1 mm pieces (Fig. 1A.2) and submerged in 1% SDS solution for 5 days. The tissues became translucent around the edges after 1 h in SDS and on the fifth day were semitranslucent and blanched—indicating the removal of cellular components (Fig. 1A.3–A.5). The decellularized tissues were then rinsed with water for another 5 days to remove residual detergent (Fig. 1A.6–A.7).
Following decellularization, the physiological architecture of the kECM remained intact, as visualized using standard histological methods (Fig. 1B, C). Both glomerular and peritubular basement membrane architectures were well preserved following decellularization (GBM: asterisks), despite the clear removal of nucleic material as evidenced by the absence of hematoxylin nucleic counterstaining (Fig. 1B). Immunofluorescence microscopy revealed the absence of nuclei (blue color) following decellularization, whereas the structure of col-IV, LAM1 FN, and HSPG remained intact (Fig. 1C)—verified through antibody affinity for each matrix component. VN, however, showed much lower expression following decellularization, indicating loss during processing.
Some nonspecific blue color appeared in small portions of decellularized tissue, likely the result of trace residual anionic detergent, which has been known to strongly bind to proteins during decellularization26,31–33 but could be trace residual nucleic material. Very rarely, nuclei were observed following decellularization; however, they did not resemble the diffuse staining that was more commonly observed in our sections, suggesting that the diffuse staining depicts anionic residues and not nucleic material.
Quantitative characterization of the decellularized kidneys
Following decellularization, acellular tissues were lyophilized and assessed with mass spectrometry (MS) to determine protein composition (Fig. 2A). Collagens and LAMs dominated the ECM composition, with col-IV found to be the most abundant protein, comprising 37.0% ± 7.2% mass area of all proteins. All six α chains of col-IV were detected, of which α1 and α2 were the most abundant at ∼28% and 43% area, respectively. The other four subunits α3, α4, α5, and α6 were detected at 6%, 5%, 13%, and 6%, respectively (Fig. 2B.1). The decellularization process preserved not only COL4A1 and A2 chains, which are ubiquitous in all basement membranes, but also COL4A3–A5 chains, which are only located in the specialized basement membranes within the GBM of kidneys.
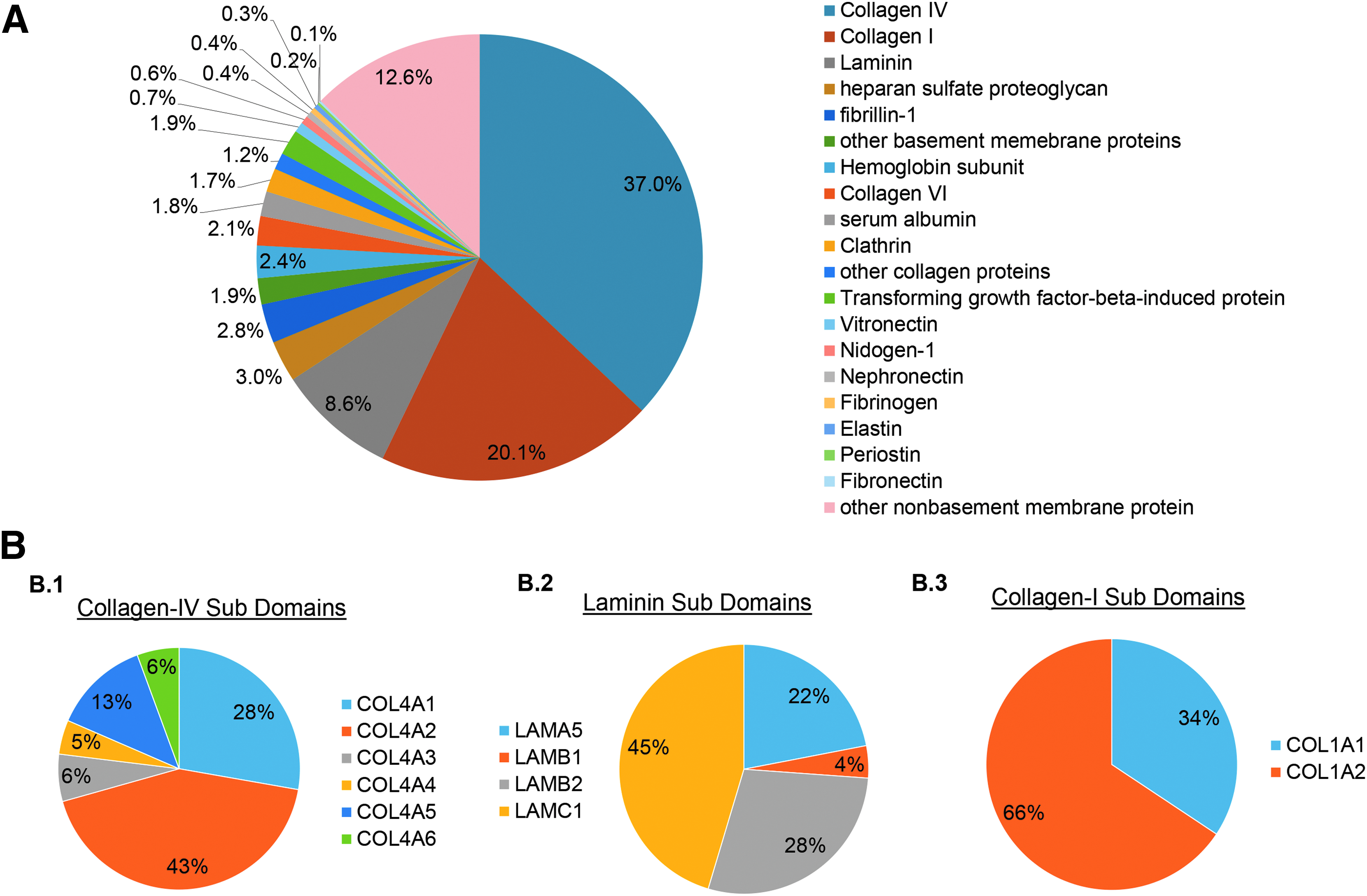
Quantitative assessment of decellularization by LC-MS/MS.
LAMs were detected to account for 8.6% ± 2.0% of all proteins, with subunits for α5, β1, β2, and γ1 (Fig. 2B.2), suggesting the preservation of the isoforms, LAM-511, the most abundant LAM in kECM, and LAM-521, which is highly restricted to glomerular and arteriolar basement membranes. 15 Collagen-I was also detected at 20.1% ± 2.3% of all proteins (Fig. 2B.3), a relatively higher value than expected—possibly a result of the average age of kidney donors (age >50) available for decellularization.
A number of other ECM components were also detected (Fig. 2A). HSPG was detected at 3.0% ± 0.2%, despite the use of a strong anionic detergent. Collagen XVIII, an abundant HSPG component of basement membranes in kidney cortex, was also present at nearly 1%. These HSPGs have the ability to sequester soluble growth factors important to cell viability and transduce signals to maintain endothelial cell stasis.34–36 Nidogen-1, a linker between LAM and col-IV in the formation of an intertwining basal laminar network, 37 was detected at ∼0.6%. Other matrix components such as vitronectin, FN and elastin—all of which are important for dictating cellular fate and signaling 38 —were detected at 2.4%, 0.4%, and 0.3%, respectively.
Characterization of a decellularized kECM gel
The lyophilized kECM matrix proteins were further digested and homogenized in the presence of an acid on ice (Fig. 3A). The kECM solid gel was made at 7.5 mg/mL after neutralizing with NaOH and gelling at 37°C for 30 min (Fig. 3B). Collagen-I at 7.5 mg/mL and a mixture gel of kECM and collagen—at a ratio of 1:1 to obtain the same 7.5 mg/mL final mass fraction—were made for comparison. The kECM formed a weak gel that was relatively delicate to handle, whereas the mixture gel at 1:1 ratio had better structural integrity (Fig. 3B.1, B.2). Immunohistochemical assessment revealed structural preservation and abundance of ECM proteins, such as col-IV, HSPG, and LAM in kECM gel (Fig. 3B).
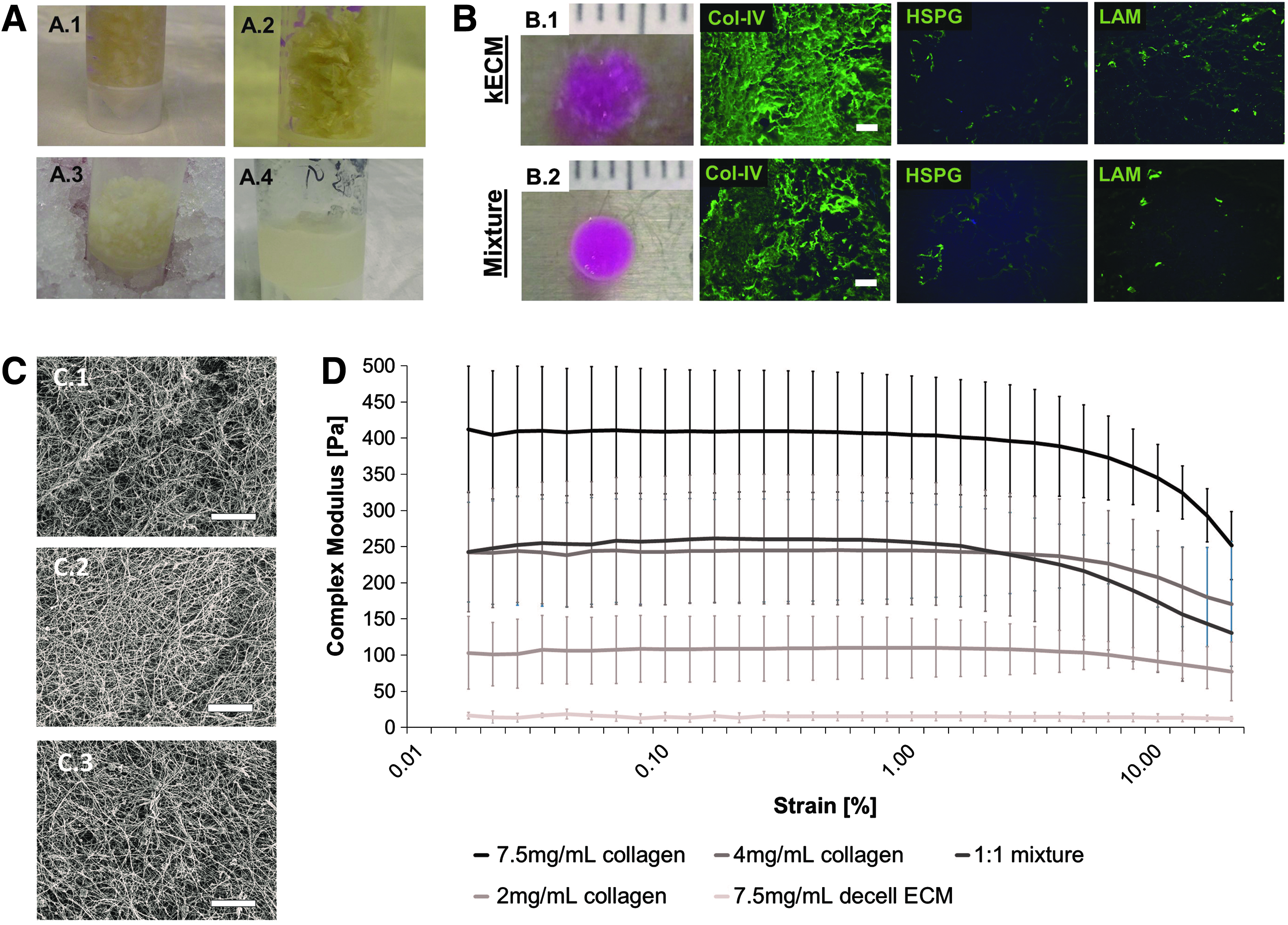
Fabrication and characterization of kECM gels.
Scanning electron microscopy revealed a greater amount and organization of fibrils in collagen-I gels compared with both mixture and kECM gels (Fig. 3C). This structural disparity is likely correlated with the amount of collagen-I present in each gel, as col-IV, the most abundant kECM protein, is not fibrillar.
Rheology testing of the 7.5 mg/mL kECM gel resulted in a complex modulus of ∼15 Pascals (Pa), almost a 10-fold decrease compared with 2 mg/mL collagen-I. The mixture gel of 7.5 mg/mL at 1:1 ratio had a complex modulus of ∼250 Pa, which is equivalent to the modulus of a 4 mg/mL collagen-I gel (Fig. 3D), and about half the value of the 7.5 mg/ml collagen-I gel.
Taken together, these data suggested that the most important component of the ECM gel to mechanical structure was collagen-I. MS data indicated that ∼20% of the kECM comprised collagen-I; however, interference by the other proteins within the kECM may have prevented the acidified collagen monomers from forming a more mechanically robust gel. Overcompensation of this effect by increasing the total collagen-I content resulted in gels with favorable mechanical integrity.
Differential endothelial cell response to ECM gels
When cultured on 2D gels of 7.5 mg/mL collagen-I, kECM, and a 1:1 mixture of the two, HKMECs responded differently. Immunofluorescence microscopy revealed differential phenotypes for cells cultured on the three different gels, highlighted by the varied distribution of CD31 around the cell surfaces and junctions (Fig. 4A). HKMECs cultured on collagen-I displayed a uniform distribution of CD31 on the surface of the cells (Fig. 4A.1); whereas HKMECs cultured on gels containing kECM displayed less expression with uneven distribution around the cell borders (Fig. 4A.2, A.3). The matrices appeared not to affect the low VWF and robustly high PV1 expression on the HKMECs for all three conditions (Fig. 4A.4–A.6).
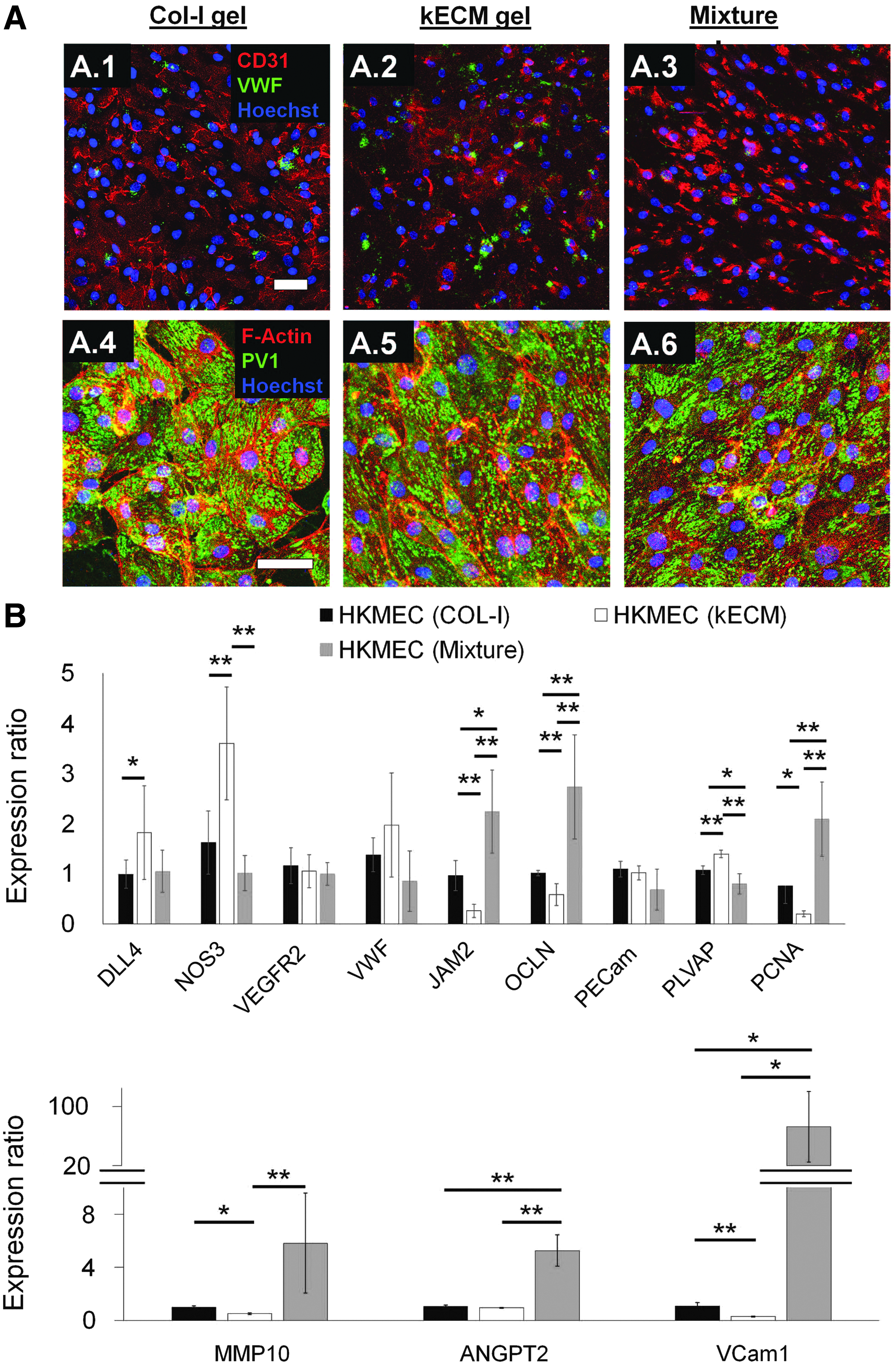
Morphological, functional, and molecular differences of HKMECs on different ECM gels.
Real-time quantitative PCR (RT-qPCR) showed upregulation of DLL4, NOS3, and PV1 and downregulation of PCNA, MMP10, and VCAM1 on HKMECs when cultured on kECM compared with collagen-I (Fig. 4B). These expression profiles indicate that HKMECs were in a more quiescent and mature state when cultured on kECM, with higher endothelial nitric oxide synthase (eNOS) synthesis capacity, lower proliferation rate, lower proteolytic activity, and less expression of adhesion molecules. JAM2 and OCLN were also significantly downregulated in HKMECs cultured on kECM, indicating decreased tight junction formation and increased vessel permeability potential.
Interestingly, HKMECs cultured on mixture gel decreased NOS and PLVAP, but increased JAM2, OCLN, PCNA, MMP10, ANGPT2, and VCAM1 expression. Thus, the incorporation of collagen-I into kECM disturbed HKMEC stasis and led to a more activated phenotype characterized by decreased eNOS synthesis, increased proliferation, increased proteolytic activity, and an increase in surface adhesion molecules.
In contrast to HKMECs, HUVECs showed little differences in morphology for all three conditions, with robust CD31 staining present along the surface of cells and at cell–cell contacts (Fig. 5A). Immunofluorescence microscopy revealed an apparent increase of cell density when cultured on kECM compared with collagen-I at the same mass fraction. HUVECs experienced a significant increase in cell density when kECM was incorporated in the gel composition (Fig. 5B).

Morphological, functional, and molecular differences of HUVECs on different gels.
Real time qPCR showed significant downregulation of DLL4, JAM2, PECAM1, and PV1, but upregulation of PCNA, MMP10, ANGPT2, and VCAM1 for HUVECs cultured on kECM compared with collagen-I (Fig. 5C). This indicates that HUVECs were in a more stimulated state when cultured on kECM, denoted by higher proliferation, more proteolytic activities, less junctional integrity, and higher expression of adhesion molecules. Expression of JAM2 and PECAM1 was less upregulated when collagen-I was added into kECM to form a mixture gel, whereas VCAM1 stayed highly upregulated. No significant changes were observed in the remaining genes that were investigated.
We further cultured HUVECs on 4 mg/mL collagen and 7.5 mg/mL kECM mixture gels, which have equivalent complex moduli, supplemented with 40 ng/mL VEGF (Fig. 5D). HUVECs readily sprouted into both matrices; however, the number of sprouts into the mixture gel was significantly higher compared with collagen-I alone (Fig. 5D.3) (p < 0.05).
kECM supports endothelial cell growth in a 3D kidney microphysiological system
Ultimately, our aim is to use the kECM gel to better recapitulate the kidney-specific extracellular matrix microenvironment to support and mimic the cellular structure and functions of the kidney for in vitro investigation. As a proof-of-principle demonstration, we used the kECM and collagen mixture gel at a 1:1 ratio to support a 3D microphysiological system (MPS) seeded with endothelial cells under gravity-driven flow (Fig. 6A), similar to previous work from our laboratory. 8
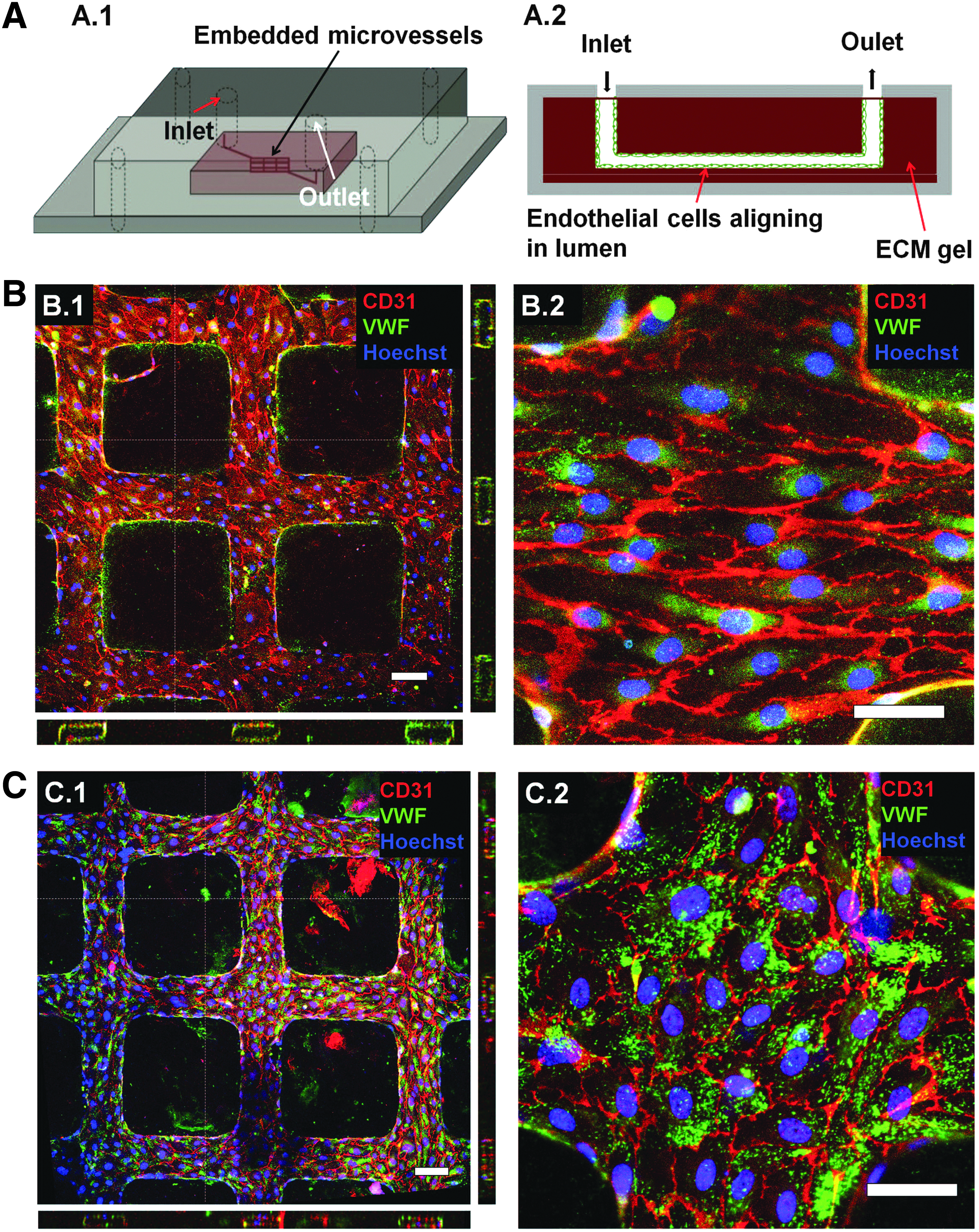
kECM mixture gels support the formation of engineered microvessels.
HKMECs cultured in this 3D system expressed more robust junctional proteins (CD31) as observed on the cell surface and cell–cell contact with uniform distribution (Fig. 6B) compared with 2D culture conditions on the same matrix composition (Fig. 4A). Likely resulting from the low mechanical integrity of these kECM incorporated gels, these microvessels showed a more than 50% reduction in vessel area, although we were able to successfully perfuse these kECM-supported microvessels over time. Overall, the incorporation of kECM provided a matrix microenvironment, which allowed HKMECs to adopt a phenotype closer to the human kidney.
Alternatively, HUVECs cultured in the 3D micropatterned network were supported and appeared similar in cellular morphology and expression of junctional proteins (CD31) and granules (VWF) compared with previous experiments utilizing collagen-I (Fig. 6C).8,30
Discussion
There has been increasing interest to use tissue engineering approaches to recreate organ-specific functional models or so-called organs on a chip. This engineering process utilizes three major components: cells, matrices, and spatial organization, with each component specifically selected to recreate organ-specific function. Kidneys function as the primary filter for harmful substances circulating in blood. As such, kidney vessels carry a high fraction of blood flow, ∼20–25% of the cardiac output. This complex system contains many specialized vascular and tubular cells and various matrix components that form a unique architecture.
Recently, decellularization strategies have been developed to generate whole organ kidney scaffolds from animals and human discarded kidneys.29,39–42 Small animal kidneys have shown promise toward this goal 29 ; however, bioengineering whole organ human kidneys remains challenging due to the large number of cells required to repopulate the large and complex architecture of the kidneys. Furthermore, it remains difficult to investigate many fundamental questions regarding the cell–cell and cell–matrix interactions and their response in injury. Alternatively, our effort to generate a human kidney MPS offers a better tool for understanding human kidney microvasculature, their interaction with the surrounding matrix and their response to injury.
Our recent work showed that kidney-specific endothelial cells, or HKMECs, have a distinct genotype, phenotype, and function compared with HUVECs, a commonly used cell source for studying endothelial biology in health and diseases. 8 In this study, we hypothesized that creating a more relevant kidney-specific matrix for use in MPSs could further the development of a human kidney on a chip that will respond appropriately to environmental cues. To this aim, we decellularized adult human kidney cortex, preserved the complex kECM, and formed kECM hydrogels. We demonstrated that HKMECs cultured on kECM, in contrast to collagen-I, had upregulated gene expression of molecular signatures suggestive of cellular quiescence and maturation and concomitantly decreased expression of genes associated with proteolytic activity and cell surface activation.
This was in marked contrast to HUVECs, which appeared more quiescent on collagen-I and more stimulated on kECM. These data follow evidence in vivo, in which increased collagen-I ECM concentration is associated with pathology in kidneys, but stasis in the umbilical vein.9,43
Taken together, our findings strongly indicate that selecting the proper ECM is a critical consideration in the development of vascularized organs on a chip and carries important implications for tissue engineering of other vascularized organs as well. As a final embodiment of the kidney MPS, incorporation of the kECM gel mixture into an engineered 3D microvascular network demonstrated its supportive function for HKMEC growth. To our knowledge, this is the first time that human kidney cortex has been formulated into a hydrogel suitable for use in a flow-directed MPS.
There remain several technical limitations elucidated in our studies that require addressing in future work. First, we found that the induction of HKMEC quiescence and maturation with kECM gels was lost when collagen-I was incorporated. Additionally, the kECM gel was mechanically insufficient to support our engineered microvessels. Engineering designs are needed that will allow for the sole incorporation of kECM gel, without additional collagen-I, which we expect to promote vascular quiescence and maturation in physiological conditions.
The differential responses of the two types of endothelial cells cultured on the different matrices are in agreement with previous work demonstrating their different intrinsic properties. 8 HUVECs are derived from the umbilical vein and are subjected to much lower hydrostatic and oncotic pressure than peritubular HKMECs. Further studies could be made to understand the role of pressure and flow on endothelial cell heterogeneity. Overall, we believe this study is an important step forward toward the goal of bioengineering a human kidney MPS with full biomimetic potential.
Footnotes
Acknowledgments
The authors acknowledge the Microfabrication facility, Electron Microscope facility, and Lynn and Mike Garvey imaging core in the University of Washington, and the Mass Spectroscopy facility in BloodWorks NW research institute. The authors thank LifeCenter NorthWest for providing discarded human kidneys and Dr. Edward Kelly for helpful discussions. This project was supported by National Institutes of Health grants, UH2/UH3 TR000504 (to J.H.) and DP2DK102258 (to Y.Z.), an NIH T32 training grant DK0007467 (to R.J.N.), and an unrestricted gift from the Northwest Kidney Centers to the Kidney Research Institute.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.