
Abstract
Chronic back pain is related to intervertebral disc (IVD) degeneration and dogs are employed as animal models to develop growth factor- and cell-based regenerative treatments. In this respect, the differential effects of transforming growth factor beta-1 (TGF-β1) and bone morphogenetic protein-2 (BMP2) on canine and human chondrocyte-like cells (CLCs) derived from the nucleus pulposus of degenerated IVDs were studied. Human and canine CLCs were cultured in 3D microaggregates in basal culture medium supplemented with/without TGF-β1 (10 ng/mL) or BMP2 (100 or 250 ng/mL). Both TGF-β1 and BMP2 increased proliferation and glycosaminoglycan (GAG) deposition of human and canine CLCs. TGF-β1 induced collagen type I deposition and fibrotic (re)differentiation, whereas BMP2 induced more collagen type II deposition. In dogs, TGF-β1 induced Smad1 and Smad2 signaling, whereas in humans, it only tended to induce Smad2 signaling. BMP2 supplementation increased Smad1 signaling in both species. This altogether indicates that Smad1 signaling was associated with collagen type II production, whereas Smad2 signaling was associated with fibrotic CLC (re)differentiation. As a step toward preclinical translation, treatment with BMP2 alone and combined with mesenchymal stromal cells (MSCs) was further investigated. Canine male CLCs were seeded in albumin-based hydrogels with/without female bone marrow-derived MSCs (50:50) in basal or 250 ng/mL BMP2-supplemented culture medium. Although the results indicate that a sufficient amount of MSCs survived the culture period, total GAG production was not increased and GAG production per cell was even decreased by the addition of MSCs, implying that MSCs did not exert additive regenerative effects on the CLCs.
Introduction
L
Extracellular matrix (ECM) synthesis and cell proliferation can be stimulated with growth factors, such as members of the transforming growth factor beta (TGF-β) superfamily, for example, TGF-β1 and bone morphogenetic protein-2 (BMP2). Members of this family transduce their signals by phosphorylation of the type I receptor by the type II receptor. Generally, type I receptors ALK1, ALK2, ALK3, and ALK6 are activated by BMPs and phosphorylate Smad1/5/8, whereas ALK4, ALK5, and ALK7 are activated by TGF-β and phosphorylate Smad2/3. 4 After phosphorylation, Smad2/3 and 1/5/8 form complexes with Smad4, translocate to the nucleus, and assemble target gene transcription. Subsequently, Smad2/3 and 1/5/8 target distinctive genes, resulting into differential effects.4,5 TGF-β and BMP2 have already been tested in vitro6–8 and in vivo in animal models with experimentally induced IVD degeneration.9–11
Growth factor stimulation alone may, however, be insufficient for IVD repair, since cell viability is impaired and cell numbers are decreased in degenerated IVDs. 12 Combined cell- and growth factor-based therapy may solve this problem.13–15 Mesenchymal stromal cells (MSCs) are an emerging cell source for regenerative treatments. They can be isolated from various tissues, for example, bone marrow and adipose tissue, and can differentiate into different cell types, for example, osteoblasts, adipocytes, and chondrocytes.16,17 Tissue regeneration can be supported by MSCs because of their immunosuppressive properties and the trophic factors they secrete. 18 Affirmatively, MSC transplantation increased ECM synthesis in animal models with experimentally induced IVD degeneration.19–25
While the aforementioned growth factors have been studied separately, their differential effects on the IVD have not been addressed. Therefore, in this study, the effect of TGF-β1 and BMP2 was determined on canine and human CLCs derived from degenerated IVDs. Since dogs suffer from spontaneous IVD degeneration with similar characteristics as observed in humans, they are considered a valid in vivo animal model for human IVD degeneration. 26 Based on physical appearance, dog breeds can be divided into chondrodystrophic (CD) and nonchondrodystrophic (NCD). 27 CD dogs have short bowlegs due to disrupted endochondral ossification. This polygenetic trait has strongly been linked with IVD degeneration. In CD dogs, replacement of NCs by CLCs in the NP starts before 1 year of age. IVD disease occurs frequently and develops around 3–7 years of age, usually in the cervical or thoracolumbar spine. 27 In contrast, in NCD dogs, NCs remain the predominant cell type until later in life. If IVD disease develops, it usually occurs around 6–8 years of age due to wear and tear, in the caudal cervical or lumbosacral spine. 27 Since CD and NCD dogs show differences in cause, prevalence, and age of onset of IVD degeneration, the regenerative potential of their CLCs could differ from each other and also from human CLCs. Therefore, we aimed to define whether CD and/or NCD dogs were, besides a valid in vivo, also a suitable in vitro animal model for growth factor-based treatment of human IVD degeneration. Last, as a step toward preclinical translation, the additive regenerative effect of MSCs on growth factor-based treatment of IVD disease was studied in an albumin-based hydrogel that has already been employed in vivo for intradiscal cell delivery. 28
Materials and Methods
The effect of TGF-β1 and BMP2 on canine and human CLC microaggregates
IVD tissue sources
CLCs of three human (two females, one male, 47–63 years), four CD (Beagles, male, 3–10 years), and four NCD (two mixed-breed dogs, one Jack Russell terrier, one German Shepherd dog, three females, one male, 2–11 years) canine donors were obtained from Thompson grade III IVDs. The NPs from all human and canine donors contained ∼100% CLCs. During standard postmortem diagnostic procedures at the University Medical Centre (UMC) Utrecht, the L2–L5 part of the human spine was collected. Anonymous use of redundant tissue for research purposes is a standard treatment agreement with patients in the UMC Utrecht (Local Medical Ethics Committee number 12-364). The material was used in line with the “Proper Secondary Use of Human Tissue” code installed by the Federation of Biomedical Scientific Societies. Canine spines were collected from dogs euthanized in unrelated research studies (University 3R-policy, approved by the Utrecht University Animal Ethics Committee) and client-owned dogs submitted for necropsy to the Department of Pathobiology (Faculty of Veterinary Medicine, Utrecht University). Briefly, NPs were digested by 0.15% pronase (45 min) and 0.15% collagenase (overnight) treatment. The CLCs were stored in hgDMEM+Glutamax (1966; Invitrogen) with 10% fetal bovine serum and 10% DMSO (−196°C) until use.
Cell culture
First, the CLCs were expanded at 21% O2 and 5% CO2 at 37°C.
29
The CLCs were pooled at passage 2 to have biological representative samples of the respective populations. Human, NCD, and CD canine microaggregates of 35,000 CLCs were formed in low-adherence cell-repellent surface 96-well plates (650970, CELLSTAR®; Greiner Bio-One) in 50 μL basal culture medium/well: hgDMEM+Glutamax, 1% P/S (P11-010; GE Healthcare Life Sciences), 1% ITS+ premix (354352; Corning Life Sciences), 0.04 mg/mL
Gene expression profiling
Microaggregates were collected on day 7 (n = 7). RNA isolation, cDNA synthesis, and reverse transcriptase-quantitative polymerase chain reaction were performed as described previously. 29 Primer specifications for all canine and human reference and target genes are given in (Supplementary Data S1 (Supplementary Data are available online at www.liebertpub.com/tea).
DNA and GAG content and release
Microaggregates were collected on day 28 (n = 7). DNA and GAG sample preparation was performed as described previously. 29 The GAG content of the microaggregates was determined using a dimethyl methylene blue (DMMB) assay. 32 The DNA content was measured using the Qubit® dsDNA High Sensitivity Assay Kit (Q32851; Invitrogen) according to the manufacturer's instructions.
Histology
On day 28, the microaggregates were fixed in 4% neutral buffered formaldehyde (4286; Klinipath B.V.) with 1% eosin (115935; Merck Millipore) and embedded in 2.4% alginate and paraffin. Five micrometer sections were mounted on Microscope KP+ slides (KP-3056; Klinipath B.V.). Alizarin Red S 16 and Safranin O/Fast Green staining and Collagen type I, II, and X immunohistochemistry 29 were performed as described previously using collagen I mouse monoclonal antibody (0.07 μg/mL [canine] and 0.1 μg/mL [human], ab6308; Abcam), collagen II mouse monoclonal antibody (0.02 μg/mL [canine] and 0.4 μg/mL [human], II-II6B3; DSHB), and collagen X mouse monoclonal antibody (2031501005; Quartett). In isotype controls, normal mouse IgG1 (3877; Santa Cruz Biotechnology) showed no specific staining. The positive control for collagen type X (growth plate) showed specific staining.
pSmad1 and pSmad2 enzyme-linked immunosorbent assay
Two hundred thousand human, CD, and NCD canine CLCs (n = 4) were plated per well (12-well plate, 665180, CELLSTAR; Greiner Bio-One) in an expansion medium. After 2 days, the expansion medium was replaced with the basal culture medium with/without 10 ng/mL TGF-β1 or 250 ng/mL BMP2. After 24 h of growth factor treatment, cells were homogenized in the RIPA buffer containing 0.6 mM phenylmethylsulphonyl fluoride, 17 μg/mL aprotinin, and 1 mM sodium orthovanadate (Sigma-Aldrich). Protein concentrations were measured using the Qubit Protein Assay Kit (Q32851; Invitrogen) and enzyme-linked immunosorbent assays (ELISAs) for pSmad1 (SER463/465, PEL-SMAD1-S463; RayBiotech) and pSmad2 (S245/250/255, PEL-SMAD2-S245; RayBiotech) were performed according to the manufacturer's instructions.
The effect of BMP2 on canine CLCs and MSCs in a hydrogel
The same CD and NCD canine CLC donor pools were used as described for the microaggregate experiments. Bone marrow-derived MSCs were obtained from a Beagle (CD) and a mixed-breed dog (NCD) (both female, 2 years old), which were euthanized in unrelated research studies. The MSCs were isolated, expanded, and characterized. 16 Thereafter, CLCs and MSCs were incorporated in 40 μL hydrogels composed of albumin crosslinked by polyethylene glycol spacers to hyaluronic acid 33 (3 × 106 CLCs/mL hydrogel [CLC], 1.5 × 106 CLCs/mL hydrogel + 1.5 × 106 MSCs/mL hydrogel [CLC:MSC], and 1.5 × 106 CLCs/mL hydrogel [½ CLC]) and cultured in the basal culture medium with/without 250 ng/mL BMP2 at 21% O2 and 5% CO2 at 37°C for 28 days. Since the differentiation potential of canine MSCs differs between breeds, 34 it was decided to only combine CLCs and MSCs of one breed: CD (Beagle) CLCs+CD (Beagle) MSCs and NCD (mixed breed) CLCs+NCD (mixed breed) MSCs.
DNA and GAG content measurements (n = 8), Safranin O/Fast Green staining, and collagen type I and II immunohistochemistry (n = 3) were performed as described above, with the adjustment of pH of the papain and DMMB solution to 6.8 and addition of 2.16 M guanidinium chloride to mask the hyaluronic acid. To determine the fate of the MSCs after 28 days, the CLC (male): MSC (female) ratio was determined by SRY:GAPDH PCR on genomic DNA isolated from 20 μL papain-digested sample using the DNEasy blood and tissue kit (69581; Qiagen). The DNA was diluted 10× and used for SRY or GAPDH qPCR (Supplementary Data S1). To determine the samples' male DNA percentage, a standard series with known female:male genomic DNA amounts was used. Cq SRY:Cq GAPDH was used to interpolate the amount of male DNA in the samples from that of the known standard series.
Statistical analysis
Statistical analyses were performed using IBM SPSS statistics 22. Data were examined for normal distribution using a Shapiro–Wilks test. General linear regression models based on ANOVAs were used for normally distributed data and Kruskal–Wallis and Mann–Whitney U tests for nonnormally distributed data. To correct for multiple comparisons, a Benjamini and Hochberg False Discovery Rate post hoc test was performed. p-Values <0.05 were considered significant.
Results
The effect of TGF-β1 and BMP2 on CLC proliferation and apoptosis
The effects of 100 and 250 ng/mL BMP2 (BMP2100 and BMP2250, respectively) and 10 ng/mL TGF-β1 were determined on microaggregate cultures of CD and NCD canine and human CLCs from degenerated IVDs. The DNA content of the human, CD, and NCD canine microaggregates was significantly increased by TGF-β1, BMP2100, and BMP2250 treatment compared with controls, with the lowest increase in BMP2100-treated microaggregates (Fig. 1A, p < 0.01). TGF-β1 induced the highest DNA content in human (p < 0.01) and CD canine (p < 0.001) microaggregates, whereas in NCD canine microaggregates, TGF-β1 and BMP2250 were equally potent in this respect. On day 7, expression of proliferation marker CCND1 was not increased by TGF-β1 in human nor canine CLCs, while both BMP2 concentrations induced CCND1 expression compared with controls in CD canine and human CLCs (p < 0.05, Supplementary Data S2).

The effect of TGF-β1 and BMP2 on human and canine CLC proliferation, matrix production, and Smad signaling. Microaggregates of CLCs derived from degenerated human/canine intervertebral discs were cultured for 28 days (DNA and GAG content) or 24 h (phosphorylated [p]Smad1, pSmad2 signaling).
In BMP2100-treated CD and NCD canine and BMP2250-treated CD canine microaggregates, a central area containing cells with karyorrhexis and pyknosis was observed, while there were no signs of apoptosis in the human microaggregates (Fig. 2). BMP2250, however, significantly reduced apoptosis marker BAX expression in NCD canine CLCs (p < 0.05, Supplementary Data S2) and tended to decrease BAX expression compared with controls in human and CD canine CLCs (p < 0.1). Expression of BAX was significantly decreased by TGF-β1 in CD and NCD canine CLCs compared with controls (p < 0.05).
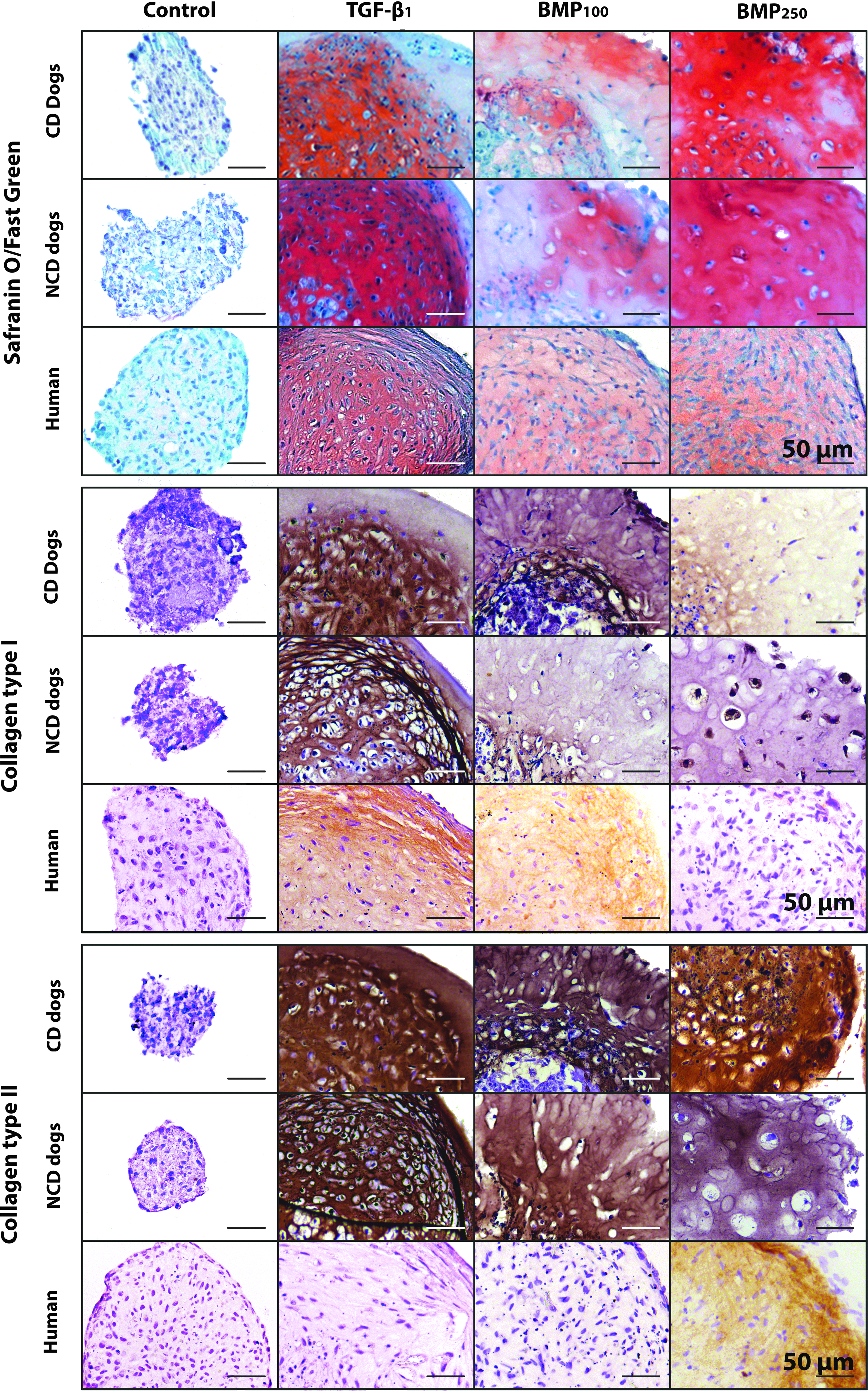
The effect of TGF-β1 and BMP2 on GAG, collagen type I and II deposition of human and canine CLC microaggregates. Microaggregates of CLCs derived from degenerated human/canine intervertebral discs were cultured for 28 days. Red staining in Safranin O/Fast Green indicates GAG deposition. n = 7.
The effect of TGF-β1 and BMP2 on CLC matrix production and remodeling
ACAN expression was significantly increased by BMP2250 treatment in CD canine and human CLCs compared with control- and TGF-β1-treated CLCs (p < 0.05), but not in NCD CLCs (Supplementary Data S2). Nevertheless, GAG deposition was significantly increased by BMP2100, BMP2250, and TGF-β1 treatment in human and canine CLCs compared with controls (p < 0.05, Figs. 1B and 2). BMP2 concentration dependently increased GAG deposition in human and canine CLCs (p < 0.05). In CD canine CLCs, TGF-β1 was significantly more potent than BMP2 in inducing GAG deposition (p < 0.05). In contrast, NCD canine microaggregates treated with BMP2250 had a significantly higher GAG content than those treated with TGF-β1 (p < 0.001), whereas TGF-β1 and BMP2250 were equally potent in increasing GAG deposition by human CLCs. The GAG/DNA content of the BMP2250-treated human and canine microaggregates was significantly increased compared with all other conditions (p < 0.05, Fig. 1C). In human and CD canine CLC microaggregates, the GAG/DNA content was significantly more increased by TGF-β1 than by BMP2100 treatment, whereas in NCD dogs, the opposite occurred.
TGF-β1 and BMP2250 were equally potent in inducing COL2A1 expression in canine and human CLCs (Supplementary Data S2), while collagen type II protein deposition was induced by both TGF-β1 and BMP2 in canine CLCs, but only by BMP2250 in human CLCs (Fig. 2). COL10A1 expression was only detected in TGF-β1-treated human CLCs (data not shown), but collagen type X protein was neither detected in canine nor human microaggregates, regardless the treatment (Supplementary Data S3). In addition, mRNA of the osteogenic marker OSX was not detected in any culture condition, and RUNX2 and BGLAP gene expression was very low (Cq-values >37) and not significantly different between conditions (data not shown). Moreover, Alizarin Red S staining showed no calcium deposition regardless the treatment (Supplementary Data S3).
In all species, TGF-β1-treated microaggregates showed signs of fibrotic (re)differentiation. Expression of collagen type I gene (Supplementary Data S2) and protein (Fig. 2) was increased by TGF-β1 compared with control and BMP2 treatment in both species, except for COL1A1 expression in CD canine CLCs. Morphologically, a GAG-depleted, collagen type I-rich outer rim was formed after TGF-β1 treatment, which contained almost no cells in canine, and fibroblast-like cells in human microaggregates (Fig. 2).
Expression of the matrix remodeling gene MMP13 was significantly induced by BMP2 in canine CLCs, but only induced by TGF-β1 treatment in human CLCs (p < 0.05, Supplementary Data S2). ADAMTS5 expression was not significantly affected by the different growth factors in human CLCs (data not shown). In contrast, TGF-β1 decreased CD canine CLC ADAMTS5 expression 10-fold, whereas BMP2250 decreased NCD canine CLC ADAMTS5 expression 7-fold compared with controls (p < 0.05, data not shown). TIMP1 (inhibitor of matrix metalloproteinase) expression was decreased in TGF-β1- and BMP2250-treated human CLCs (2- and 1.5-fold, respectively) compared with controls (p < 0.05), whereas it was 30-fold decreased by BMP2250 in NCD canine CLCs (p < 0.05), and not affected by growth factor treatment in CD canine CLCs (data not shown).
The effect of TGF-β1 and BMP2 on Smad signaling
To determine the effect of TGF-β1 and BMP2250 on Smad signaling, ELISAs for phosphorylated (p)Smad1 and Smad2 were performed. In CD canine CLCs, TGF-β1 significantly induced Smad1 and Smad2 signaling, whereas BMP2250 only induced pSmad1 levels (p < 0.05, Fig. 1D, E). In NCD canine CLCs, TGF-β1 significantly increased pSmad2 (p < 0.05) and tended to increase pSmad1 levels (p = 0.1), whereas BMP2250 significantly increased pSmad1 levels (p < 0.05). TGF-β1-treated human CLCs showed a tendency toward increased pSmad2 levels (p = 0.1), whereas BMP2250-treated human CLCs showed significantly increased Smad1 signaling (p < 0.05).
The effect of BMP2 on canine CLCs and MSCs in 3D hydrogel cultures
The best growth factor was considered the one that induced GAG and collagen type II production and cell proliferation, and inhibited apoptosis and fibrotic (collagen type I rich) and osteogenic (collagen type X rich and calcium rich) ECM deposition. In this respect, human and canine CLC microaggregates responded best to 250 ng/mL BMP2. Therefore, the effect of 250 ng/mL BMP2 alone or in combination with MSCs was determined on canine CLCs using hydrogels employed for intradiscal cell transplantation. 28 Canine CLCs and MSCs from CD and NCD breeds were incorporated at different concentrations: 3 × 106 CLCs/mL (CLC), 1.5 × 106 CLCs/mL + 1.5 × 106 MSCs/mL (CLC:MSC), and 1.5 × 106 CLCs/mL (½ CLC).
In hydrogels containing CD (Fig. 3) and NCD (Supplementary Data S4) canine ½ CLC, CLC, and CLC:MSC, BMP2 treatment significantly increased the DNA content, GAG release, GAG deposition, and collagen type I and II deposition compared with controls (p < 0.05). Generally, BMP2 treatment also significantly increased the GAG/DNA content of NCD (Supplementary Data S4) and CD (Fig. 3D) canine cell containing hydrogels (p < 0.01), but not the GAG/DNA content of CD CLC:MSC containing hydrogels.
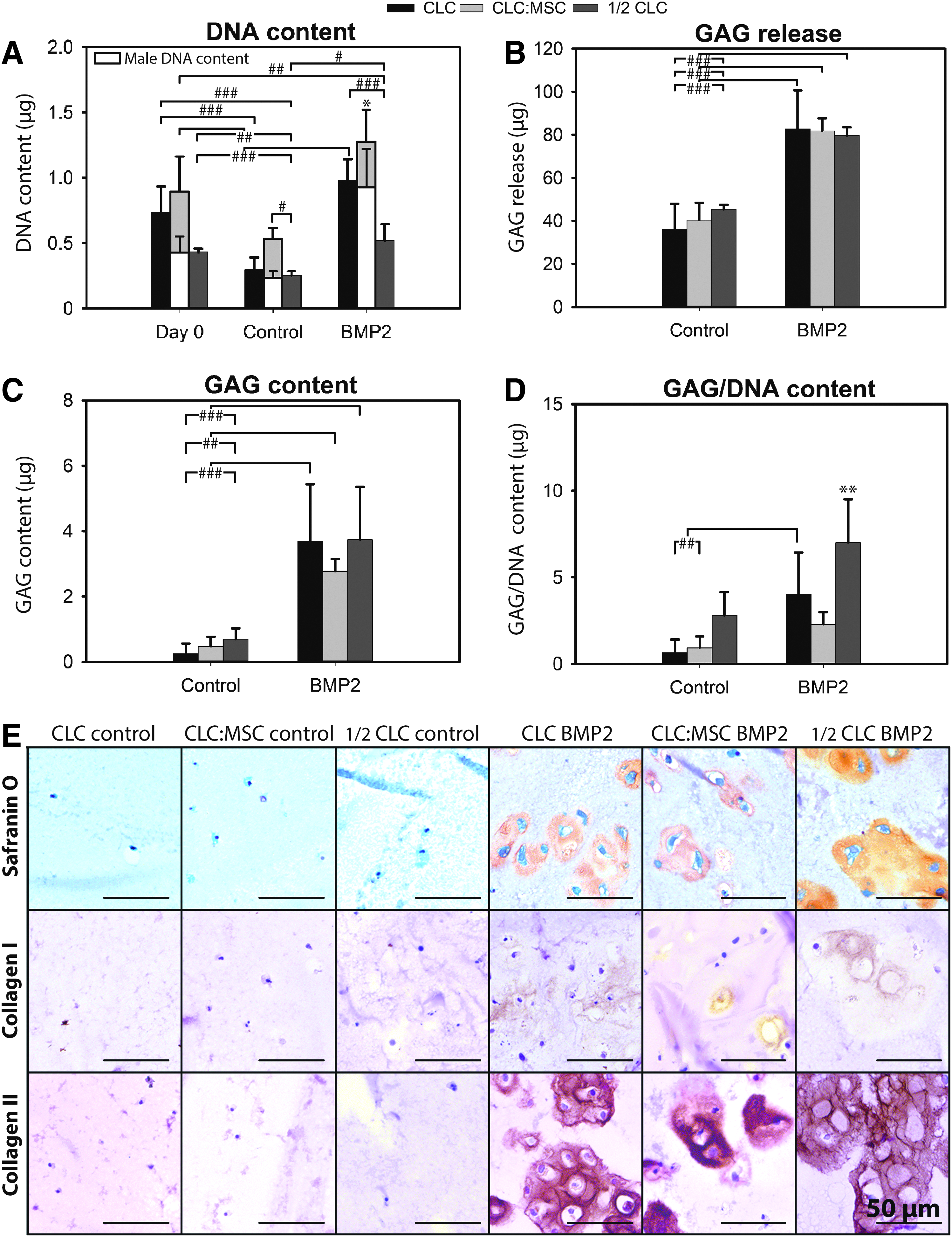
The effect of BMP2 on matrix production and cell proliferation of CD canine CLCs and MSCs. CLC (3 × 106/mL hydrogel), ½ CLC (1.5 × 106/mL), or CLC:MSC (1:1, both 1.5 × 106/mL) hydrogels were cultured for 28 days with or without 250 ng/mL BMP2.
Although on day 0, the DNA content of CLC and CLC:MSC-containing hydrogels was comparable, after 28 days, the DNA content of NCD CLC:MSC-containing control hydrogels was higher compared with NCD CLC-containing control hydrogels (p < 0.01, Supplementary Data S4). The DNA content of the BMP2-treated CLC:MSC-containing hydrogels was higher compared with the BMP2-treated CLC-containing hydrogels in both CD and NCD dogs (p < 0.05, Fig. 3A and Supplementary Data S4). On day 0, the CLC (male)-derived DNA content of CD CLC:MSC-containing hydrogels was 50% (0.45 μg) of the total DNA content. After 28 days of culture, 57% (0.30 μg) of cells in CD CLC:MSC-containing control hydrogels appeared female (MSC) derived, whereas this was 27% (0.34 μg) in CD BMP2-treated CLC:MSC-containing hydrogels (Fig.3A), indicating that in absolute terms, the MSC-derived DNA content remained relatively unchanged throughout culture. On day 28, the CLC (male)-derived DNA content of CD CLC:MSC-containing hydrogels was comparable with the total DNA content of CD CLC-containing hydrogels, both in the presence and absence of BMP2 (Fig. 3A).
GAG release and deposition were comparable in all BMP2-treated cell–hydrogel combinations. The GAG/DNA content was highest in the hydrogels containing BMP2-treated ½ CLC, followed by BMP2-treated CLC hydrogels (Fig. 3D and Supplementary Data S4). The GAG/DNA content of NCD and CD BMP2-treated CLC:MSC-containing hydrogels was significantly lower compared with BMP-treated ½ CLC and CLC-containing hydrogels (p < 0.01).
A schematic overview of the observed effects of TGF-β1 and BMP2 on CD and NCD canine and human CLCs is given in Supplementary Data S5.
Discussion
Both humans and dogs experience chronic back pain related to IVD degeneration. As current treatments do not achieve IVD repair, there is need for cell- and growth factor-based regenerative treatments leading to functional IVD restoration. This study demonstrates that both growth factors BMP2 and TGF-β1 increased proliferation and matrix production of human and canine CLCs, but TGF-β1 induced a fibrotic phenotype, whereas BMP2 did not. To determine whether MSCs exerted additional regenerative effects, canine CLCs and MSCs were cultured in a hydrogel that has already been shown to facilitate IVD repair. 28 Although the results indicate that a considerable amount of MSCs survived the culture period, they did not exert additive regenerative effects on the CLCs.
The effect of TGF-β1 and BMP2 on human and canine CLCs
It has already been shown that BMP26,7 and TGF-β8,29 are able to induce GAG deposition in CLCs. In this study, the response of CD and NCD canine CLCs to these growth factors was in a similar direction as observed in human CLCs, but sometimes with a different intensity. Both BMP2 and TGF-β increased cell proliferation and ECM production, while they decreased the expression of apoptosis-related genes in human and canine CLCs. These effects were reflected in a high DNA and GAG content of the CLC microaggregates at the end of the study. TGF-β1 stimulated Smad1 and Smad2 signaling in canine CLCs (as also shown in chondrocytes 35 ) and induced GAG, collagen type I and II deposition, and a cell/GAG-depleted fibrotic rim. In contrast, BMP2 only stimulated Smad1 signaling in canine CLCs, and induced GAG and collagen type II deposition, but no collagen type I deposition or fibrotic phenotype. In human CLCs, TGF-β1 tended to stimulate only Smad2 signaling and induced a fibroblast-rich construct, GAG and collagen type I, but no collagen type II deposition. Moreover, BMP2 only stimulated Smad1 signaling in human CLCs and induced GAG and collagen type II, but no collagen type I deposition. Altogether, these results suggest that Smad1 signaling was associated with collagen type II production, whereas Smad2 signaling accompanied fibrotic (re)differentiation in human and canine CLCs. In line with our results, TGF-β is a well known, collagen type I-inducing, fibrotic agent, 36 whereas BMP2 did not increase COL1A1 expression in human CLCs. 37 In chondrocytes, however, TGF-β1-mediated Smad2/3 signaling results in protective, and Smad1/5/8 signaling in deleterious responses, 35 indicating that CLCs respond distinctively different to (TGF-β1-mediated) Smad2/3 signaling than chondrocytes.
Hypertrophic differentiation is known to precede BMP (Smad1/5/8)-induced bone formation in articular chondrocytes.35,38 In this study, however, no signs for hypertrophic differentiation or osteogenic matrix production were detected in BMP2-treated canine or human CLCs. In line with our results, others showed only chondrogenic, but no osteogenic effects of BMP2 on rabbit NCs, 30 human CLCs,7,37 and degenerated goat IVDs, 11 indicating that CLCs may also respond rather different to BMP2 (Smad1/5/8 signaling) than chondrocytes.
Altogether, the results of this study suggest that BMP2 can be a valuable candidate for IVD regeneration, since it induced chondrogenic, but no osteogenic or fibrotic ECM production in canine and human CLCs. Future studies should, however, investigate long-term exposure to BMP2. Last, CD and NCD canine CLCs responded rather comparably to growth factor treatment as human CLCs, indicating that CLCs from both CD and NCD dogs can be used as in vitro animal models for human IVD degeneration.
The effect of MSCs on canine CLCs
Hydrogels that are already applied in clinical studies for intradiscal cell transplantation 28 were used to determine the possible additive regenerative effect of MSCs on BMP2-treated CLCs. The results of this study indicate that a considerable amount of MSCs survived the 28-day CLC:MSC coculture period. Also in vivo, injected MSCs maintain their viability for at least 28 days within rat IVDs. 39 In cocultures, MSCs have been shown to stimulate chondrocyte proliferation and ECM production. 40 Although previous work on human CLC:MSC cocultures also showed (modest) trophic effects of MSCs,41,42 this study on canine CLC:MSC cocultures did not. Comparable results as observed in this canine coculture study have been demonstrated for human chondrocyte:MSC cocultures, 43 stressing the importance of using proper controls. Cells should be seeded at comparable densities, but also at lower densities to account for possible differences in available nutrition or cells producing factors inhibiting their own proliferation.
Taken together, MSCs did not exert regenerative effects on CD or NCD canine CLCs in vitro. The discrepancy between these results and results of previous work41,42 could be due to species differences. In addition, our experiments were performed in normoxia, whereas the NP is a hypoxic tissue. 44 Hypoxia produces a favorable microenvironment that improves MSC cell viability, as well as differentiation into NP-like cells and ECM synthesis. 45 Furthermore, previous work indicated that the optimal CLC:MSC ratio for MSC differentiation appeared 75:25.42,46 Therefore, future studies should test different CLC:MSC ratios under hypoxic culture conditions. The results of this study do not directly imply that MSCs will not have regenerative effects on degenerated IVDs in vivo. Given that regenerative effects of MSCs were encountered in different animal models with experimentally induced IVD degeneration,19–25 future studies should investigate whether MSCs also have beneficial effects in dogs with spontaneously degenerated IVDs, a valid in vivo animal model for human IVD degeneration. 26
Conclusions
This study demonstrates that, while both TGF-β1 and BMP2 exerted regenerative effects on human and canine CLCs, BMP2 induced a more healthy NP matrix than TGF-β1. The (BMP2-mediated) Smad1 signaling increased collagen type II production, whereas (TGF-β1-mediated) Smad2 signaling induced fibrotic CLC (re)differentiation. MSCs did not exert additive regenerative effects on control- or BMP2-treated canine CLCs. Since CD and NCD canine CLCs responded rather comparable to TGF-β1 and BMP2 as human CLCs, support for the dog as a valid in vitro model for human IVD degeneration was provided.
Footnotes
Acknowledgments
This work was funded by AOSpine International (SRN2011_11) and the Dutch Arthritis Foundation (LLP22). The authors would like to thank Anita Krouwels for supplying the human CLCs and Willem de Jong and Sanne Loos for assistance during the execution of experiments.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.