
Abstract
Objective:
When serially passaged in standard monolayer culture to expand cell number, articular chondrocytes lose their phenotype. This results in the formation of fibrocartilage when they are used clinically, thus limiting their use for cartilage repair therapies. Identifying a way to redifferentiate these cells in vitro is critical if they are to be used successfully. Transforming growth factor beta (TGFβ) family members are known to be crucial for regulating differentiation of fetal limb mesenchymal cells and mesenchymal stromal cells to chondrocytes. As passaged chondrocytes acquire a progenitor-like phenotype, the hypothesis of this study was that TGFβ supplementation will stimulate chondrocyte redifferentiation in vitro in serum-free three-dimensional (3D) culture.
Methods:
Human articular chondrocytes were serially passaged twice (P2) in monolayer culture. P2 cells were then placed in high-density (3D) culture on top of membranes (Millipore) and cultured for up to 6 weeks in chemically defined serum-free redifferentiation media (SFRM) in the presence or absence of TGFβ. The tissues were evaluated histologically, biochemically, by immunohistochemical staining, and biomechanically.
Results:
Passaged human chondrocytes cultured in SFRM supplemented with 10 ng/mL TGFβ3 consistently formed a continuous layer of articular-like cartilage tissue rich in collagen type 2 and aggrecan and lacking collagen type 1 and X in the absence of a scaffold. The tissue developed a superficial zone characterized by expression of lubricin and clusterin with horizontally aligned collagen fibers.
Conclusions:
This study suggests that passaged human chondrocytes can be used to bioengineer a continuous layer of articular cartilage-like tissue in vitro scaffold free. Further study is required to evaluate their ability to repair cartilage defects in vivo.
Introduction
A
Chondrocytes, the cell of articular cartilage, may be the optimal cell source for this purpose as, unlike mesenchymal stromal cells (MSCs), they do not progress to hypertrophic differentiation in vitro3,4 and have been shown to maintain a stable phenotype and resist terminal differentiation when implanted in vivo. 4 Furthermore, in a comparative study, human chondrocyte cultures were shown to accumulate significantly more cartilage-specific ECM than chondrocytes derived from adipose-derived stromal cells. 5 Autologous chondrocytes can be easily obtained via enzymatic digestion of a small biopsy of articular cartilage; however, their number is limited and they must be serially passaged to obtain sufficient cells. During this process, they dedifferentiate and lose their phenotype 6 and are subsequently unable to form articular cartilage tissue. A number of investigators have attempted to identify conditions that would prevent dedifferentiation. For example, Martin et al. supplemented the culture media with fibroblastic growth factor 2 during serial passaging of bovine chondrocytes in an attempt to prevent dedifferentiation. 7 This approach was moderately successful at maintaining the hyaline cartilage phenotype of the passaged cells; however, tissues formed by passaged cells did not accumulate as much collagen as the tissues formed by primary cells. Thus, the use of articular chondrocytes for bioengineering has been limited as the conditions necessary to prevent dedifferentiation while favoring cell proliferation have not been identified to date.
An alternative approach to this issue of dedifferentiation would be to redifferentiate the passaged cells. Several methods have been developed to do this, but they have their limitations. Coculture of passaged chondrocytes with primary chondrocytes has been shown to redifferentiate passaged chondrocytes and result in formation of stable articular cartilage in vitro.8,9 This method, however, requires a large number of primary chondrocytes, which are difficult to acquire if the tissue is to be autologous, making this technique problematic to use for cartilage formation on a larger scale. Others have attempted to induce redifferentiation using three-dimensional (3D) culture, such as pellet cultures,10,11 which in a comparative study was shown to promote chondrocyte phenotype more effectively than culture in monolayer. 12 A third approach has been to use porous 3D scaffolds designed to favor chondrogenesis and promote cartilage-specific ECM deposition, but this has had varying success.13–15 In a previous study we showed that passaged bovine chondrocytes could be induced to redifferentiate and form hyaline cartilage scaffold free after serial passage, using a chemically defined serum-free chondrogenic redifferentiation media (SFRM). 16 High glucose concentration, insulin, and dexamethasone were identified as factors essential for this to occur. Unfortunately, preliminary studies suggested that passaged human chondrocytes would not redifferentiate under these conditions, suggesting that additional factor(s) are required for human cells.
Although many different growth factors could be examined for their ability to induce redifferentiation in human cells, one particular candidate, transforming growth factor beta (TGFβ), appears the most promising. Members of the TGFβ superfamily play vital roles in development and homeostasis of articular cartilage. Three isoforms of TGFβ exist in mammals (TGFβ1, TGFβ2, and TGFβ3) and have been shown to be essential in chondrogenic differentiation of fetal limb mesenchymal cells17,18 and MSCs 19 and in regulating proteoglycan synthesis by primary chondrocytes.20,21 Furthermore, a recent study has demonstrated that the TGFβ signaling pathway is the most highly upregulated gene network during the spontaneous repair of articular cartilage defects in rat. 22 Altogether, these studies indicate that TGFβ signaling plays an important role in both cell differentiation and repair of articular cartilage. Interestingly, in osteoarthritis (OA), a disease of progressive cartilage loss, both TGFβ levels and TGFβ receptors are decreased, 23 further supporting the involvement of this growth factor in regulating cartilage homeostasis. We hypothesize that human passaged chondrocytes will require TGFβ in combination with SFRM to promote redifferentiation and articular cartilage tissue formation. Other groups have shown increased accumulation of Col2 and sulfated proteoglycans by human passaged chondrocytes when grown in the presence of TGFβ in pellet cultures.24,25 However, tissues formed by pellet culture often have nonuniform matrix deposition, with the outer regions of the pellets accumulating more matrix than the center. Furthermore, tissues formed by chondrocyte pellets generally contain collagen type 1 (Col 1), suggesting that the passaged cells have not fully redifferentiated. To the best of our knowledge, this is the first report of the formation of a continuous layer of articular cartilage-like tissue, with a superficial zone formed by passaged human chondrocytes under scaffold-free conditions.
Methods
Chondrocyte isolation and culture
Human articular cartilage was obtained from femoral condyles resected during joint arthroplasty surgery for osteoarthritis following informed patient consent and with Research Ethics Board (Mt. Sinai Hospital) approval. A total of 26 patient samples were used for this study, 20 females and 6 males with an average age of 62 ± 10 years. Tissue was taken from areas that macroscopically appeared undamaged. Chondrocytes were isolated by enzymatic digestion of the tissue in 0.5% protease (Sigma-Aldrich) for 1 h at 37°C followed by an overnight digestion in 0.1% collagenase A (Roche) at 37°C. Cells from each patient were cultured separately for experiments (cells from multiple patients were not pooled). Nonpassaged primary (P0) cells were seeded directly onto filter membranes after enzymatic isolation from the native cartilage tissue. Alternatively, freshly harvested chondrocytes from the same donor were placed in monolayer at 2000 cells/cm2 in Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% fetal bovine serum. Chondrocytes were passaged twice (P2) to allow for ∼400-fold increase in cell number (up to a total of nine population doublings). P2 (2 × 106) cells were seeded onto type II collagen-coated Millicell® membrane culture inserts (12 mm diameter; EMD Millipore) and cultured for up to 6 weeks in SFRM consisting of high glucose DMEM (4.5 g/L), 1% ITS+ (BD Bioscience), L-proline (40 μg/mL), pyruvate (110 μg/mL), dexamethasone (0.1 μM), and L-ascorbic acid (100 μg/mL) in the presence or absence (control) of 10 ng/mL active human recombinant TGFβ3 (R&D Systems). If cultured for longer than 3 weeks the tissues were either “transiently” supplemented with TGFβ3 (first 3 weeks only) or “continuously” supplemented (throughout the entire 6 weeks). In selected cultures, TGFβ was removed after 1 week and β-glycerophosphate (10 mM) was added in combination with triiodothyronine (30 nM) or L-thyroxine (50 nM) or retinoic acid (100 nM) for an additional 2 weeks of culture to evaluate the ability of the tissues to mineralize.
Tissue content analysis
In vitro-formed tissues were digested with papain (40 μg/mL; Sigma-Aldrich) in a digestion buffer containing 20 mmol/L ammonium acetate, 1 mmol/L-EDTA, and 2 mmol/DTT at pH 6.2 for 48 h at 65°C. DNA content was quantified by the Hoechst 33258 dye binding assay and fluorometry (excitation λ = 365 nm, emission λ = 458 nm). The DNA standard curve is generated using calf thymus DNA (Sigma-Aldrich). Hydroxyproline content was quantified following HCl hydrolysis of the papain digest using the chloramine-T/Ehrlich's reagent assay and spectrophotometry (λ = 560 nm). L-hydroxyproline (Sigma-Aldrich) was used to generate the standard curve. Sulfated glycosaminoglycan content of papain digested samples was quantified by dimethylmethylene blue dye binding assay and spectrophotometry (λ = 525 nm). Chondroitin sulfate (Sigma-Aldrich) was used to generate the standard curve.
Histology and immunohistochemistry
Tissues were fixed in 10% phosphate buffered saline (PBS) buffered formalin (pH 7.4) for 4 h, placed in a 30% sucrose/PBS solution overnight at 4°C, and then snap-frozen in Tissue-Tek OCT frozen compound (Sakura Finetek). Blocks were sectioned at 7 μm, and tissue sections were stained with hematoxylin and eosin, toluidine blue, or picrosirius red, mounted with Micromount (Leica Biosystems) and coverslipped.
For visualization of alkaline phosphatase (ALP) activity, tissues were fixed in 10% neutral buffered formalin for 30 min, then embedded and sectioned as described above. Sections were incubated in azo dye (Naphthol AS-MX phosphate and Fast Blue BB salt, both Sigma-Aldrich) for 10 min according to the manufacturer's protocol, counterstained with eosin, mounted with Micromount (Leica Biosystems), and coverslipped.
For immunohistochemistry, sections were digested with 0.4% pepsin, blocked with 20% goat serum, and then incubated overnight at 4°C with antibody reactive with type I collagen (1:250 CL-50111AP-1 Cedarlane), type II collagen (1:100, MAB8887, EMD Millipore), type X collagen (1:1500 C7974, Sigma-Aldrich), Lubricin (1:250, MAB401, EMD Millipore), or Clusterin (1:250 NBP1-19637, Novus Biologicals). For aggrecan, no antigen retrieval was used (1:500 AHP0022, Thermo Fisher Scientific). To visualize reactivity, the tissues were incubated with Alexa-594 goat anti-rabbit IgG (1:1000 Life Technologies), Alexa-488 goat anti-mouse IgG (1:1000, Life Technologies), Alexa-594 goat anti-mouse IgM (1:1500, Life Technologies), or Alexa-488 goat anti-rabbit IgG secondary antibody depending on the species and isotype of the primary antibody. Tissues were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) (1:2000). Negative controls consisted of replacing the primary antibody with isoform and species-matched antibody at the same concentration.
Mechanical testing
Mechanical testing was performed with a Mach-1 micromechanical tester (Biomomentum, Laval, QC) equipped with a 1 kg load cell (0.05 g resolution, 2.5 kHz sample rate). Thicknesses of the constructs were determined through the needle probe method 26 and were measured twice at different locations near the center of the construct and averaged. Construct mechanical properties (bulk modulus, Young's modulus, and Poisson's ratio) were determined through Hayes's double indentation method27,28 using two plane-ended indenters (2 and 4 mm), indenting the samples to 10% strain at a rate of 10%/s. Constructs were allowed to equilibrate ∼20 min between indentations.
Statistics
Three independent experiments were performed, and all conditions were done in triplicate. The data are pooled, expressed as mean ± 95% confidence interval, and analyzed using analysis of variance with Tukey's post hoc. Significance was assigned at p < 0.05.
Results
TGFβ is required for in vitro articular cartilage tissue formation by passaged human chondrocytes
To determine if human passaged chondrocytes could be redifferentiated and form articular cartilage, P2 cells were grown in SFRM in the presence or absence of TGFβ. Preliminary studies indicated that both TGFβ1 and TGFβ3 had similar efficacy in stimulating tissue formation at the same concentration; therefore, we chose to use TGFβ3 for the remainder of the study. When a concentration of 1 ng/mL TGFβ3 was utilized, patient-specific differences in the amount of ECM accumulated was observed; however, when 10 ng/mL TGFβ3 was used, consistent ECM was accumulated despite potential patient variability (data not shown). Therefore, 10 ng/mL was used for the remainder of the study for consistent tissue formation. When P2 chondrocyte cultures were supplemented with 10 ng/mL TGFβ3, a continuous layer of robust cartilage tissue formed (Fig. 1A, B). In contrast, in the absence of TGFβ, the tissue was fragile and could not be easily handled. After 3 weeks of in vitro culture, treated tissues had accumulated significantly more sulfated glycosaminoglycan (Fig. 1C) and collagen (Fig. 1D) than untreated controls. Histological evaluation shows that the tissues cultured with TGFβ3 form a continuous layer of hyaline cartilage-like tissue with ovoid-shaped cells and rich in proteoglycans when stained with toluidine blue (Fig. 1F, H). In contrast, the cells cultured in the absence of TGFβ3 were elongated and there was little accumulation of proteoglycans (Fig. 1 E, G).

TGFβ3 is required for cartilaginous tissue formation by passaged human chondrocytes. P2 chondrocytes were cultured on membrane inserts for 3 weeks in SFRM with 10 ng/mL TGFβ3.
TGFβ supplementation must be maintained for 2 weeks to allow for significant collagen accumulation by passaged human chondrocytes in the in vitro-formed tissues
The time in culture with TGFβ required for significant ECM accumulation was determined. Although sulfated glycosaminoglycan content was significantly greater than untreated controls by week 1 (Fig. 2A), hydroxyproline content was not significantly greater until the second week of culture (Fig. 2B), indicating that continuous treatment with TGFβ for at least 2 weeks is necessary. Histological examination of tissues harvested at weeks 1, 2, and 3 (Fig. 2C) appeared to increase in thickness with time in culture compared to control tissues cultured for 3 weeks without TGFβ. Immunohistochemical studies showed that tissues cultured with TGFβ for longer periods of time appeared to stain more strongly for Acan and for Col 2. Of note, no Col 1 was detected in any of the in vitro-formed tissues. The cells showed no evidence of hypertrophy, as the tissues were negative for ALP activity (Fig. 3B) and collagen type X (Col X) (Fig. 3C). Furthermore, tissues did not mineralize when TGFβ was removed and cultures were supplemented with β-glycerophosphate alone or in combination with triiodothyronine or thyroxine or retinoic acid (data not shown), known inducers of mineralization by growth plate chondrocytes and MSC-derived chondrocytes.19,29–31

A minimum of 2 weeks of culture with TGFβ3 is required for significant accumulation of collagen and proteoglycan. In vitro-formed tissues were treated continuously with 10 ng/mL TGFβ3 unless otherwise stated and harvested at 1, 2, or 3 weeks of culture.
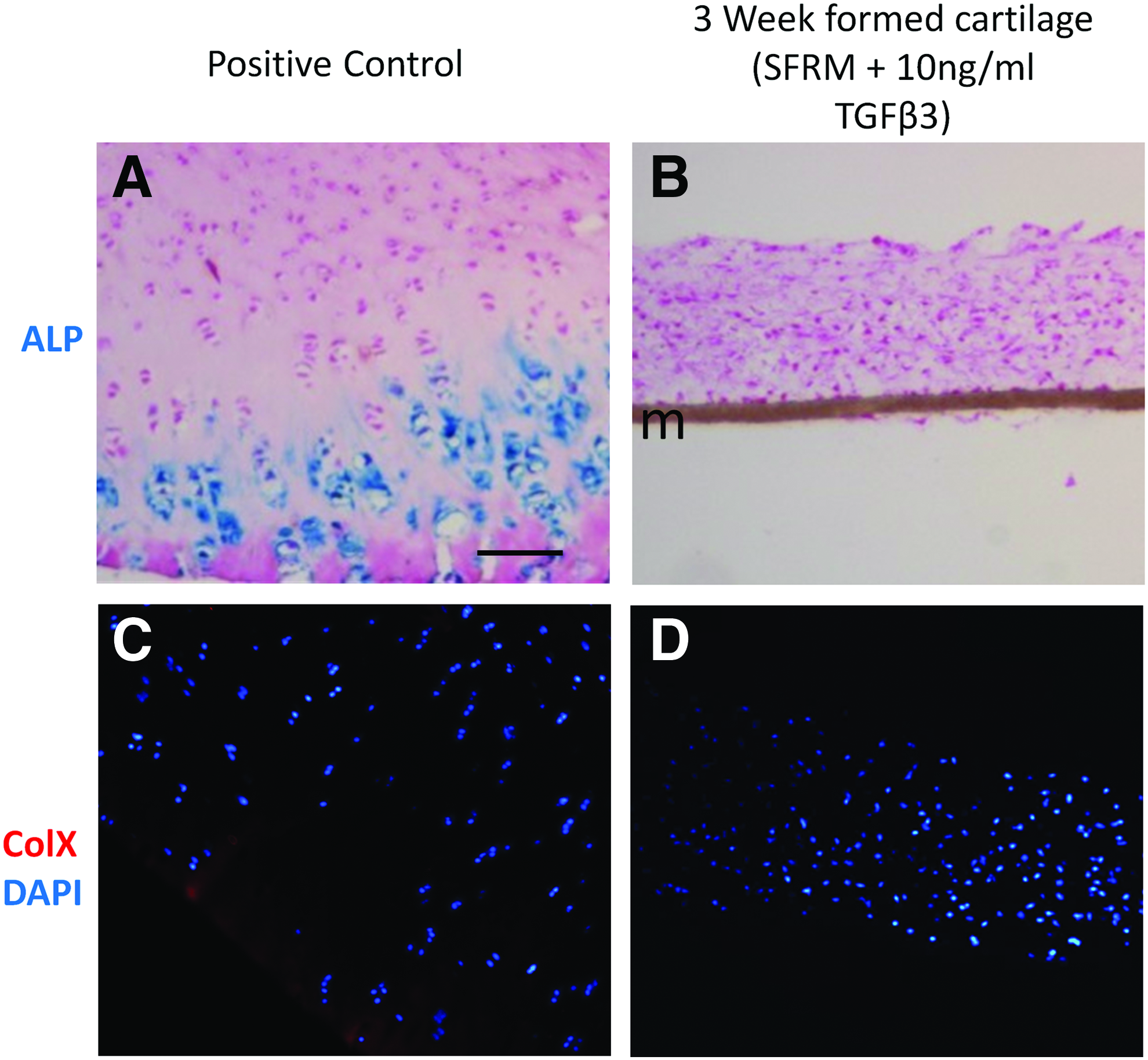
Hypertrophic markers, ALP and Col X, are not expressed in 3 weeks cultured in vitro-formed cartilage tissue.
Passaged chondrocytes accumulate more cartilage ECM than primary chondrocytes
To determine if the effect of TGFβ3 was specific to passaged chondrocytes, ECM accumulation by human passaged (P2) chondrocytes and primary (P0) chondrocytes (never passaged) was quantified. The passaged chondrocytes formed more robust tissue after 3 weeks of culture with TGFβ, accumulating significantly more collagen (Fig. 4A) than primary chondrocytes. Both passaged and primary cells accumulated similar amounts of sulfated proteoglycan when cultures were supplemented with TGFβ3 (Fig. 4B). Similar to passaged chondrocytes, primary chondrocytes did not form cartilage tissue in the absence of TGFβ3 (SFRM alone).
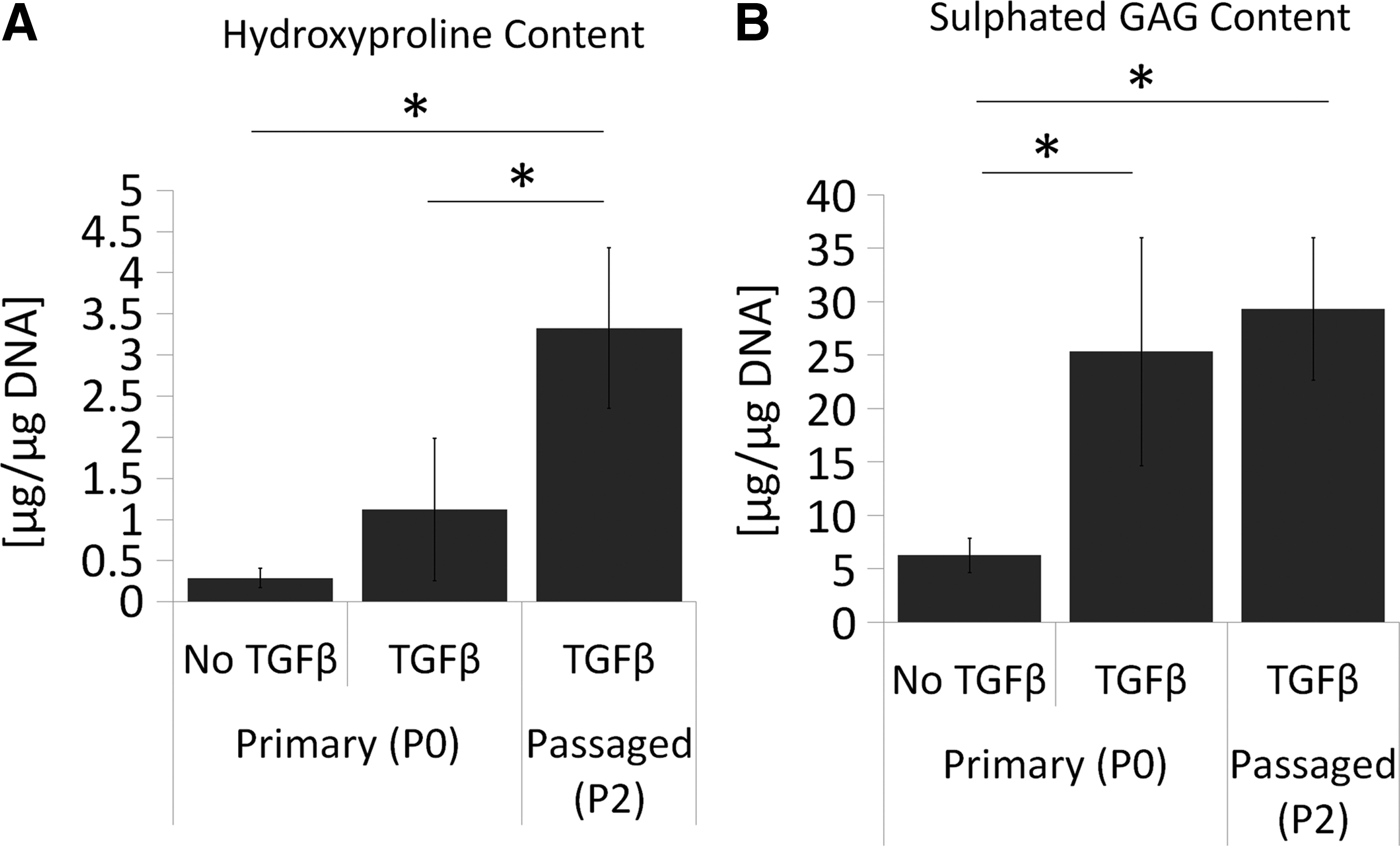
Passaged chondrocytes accumulate significantly more collagen in response to TGFβ3 than primary chondrocytes. Chondrocytes were seeded directly onto membranes (P0) or after serially passaging twice (P2) and grown for 3 weeks in SFRM with or without 10 ng/mL TGFβ3 as stated.
In vitro-formed cartilage maintains the ability to accumulate significant amounts of collagen after TGFβ3 supplementation is stopped
If used clinically for cartilage repair, following implantation, the in vitro-formed tissue would no longer be exposed to exogenous TGFβ. Therefore, we investigated whether the chondrocytes would continue to accumulate ECM when supplementation of TGFβ was discontinued. Cultures in which TGFβ3 was discontinued after weeks 3, but grown for an additional 3 weeks (termed “transient TGFβ3”), the sulfated glycosaminoglycan content decreased (Fig. 5A). This loss was not significant compared to 3-week-old cultures but was significantly reduced when compared to tissues cultured continuously for 6 weeks in the presence of TGFβ3. In contrast, significant amounts of collagen continued to accumulate in the absence of TGFβ3 (Fig. 5B). Immunohistochemical studies revealed that it was Col 2 and not Col 1 that accumulated after media supplementation with TGFβ3 ceased (Fig. 5C).
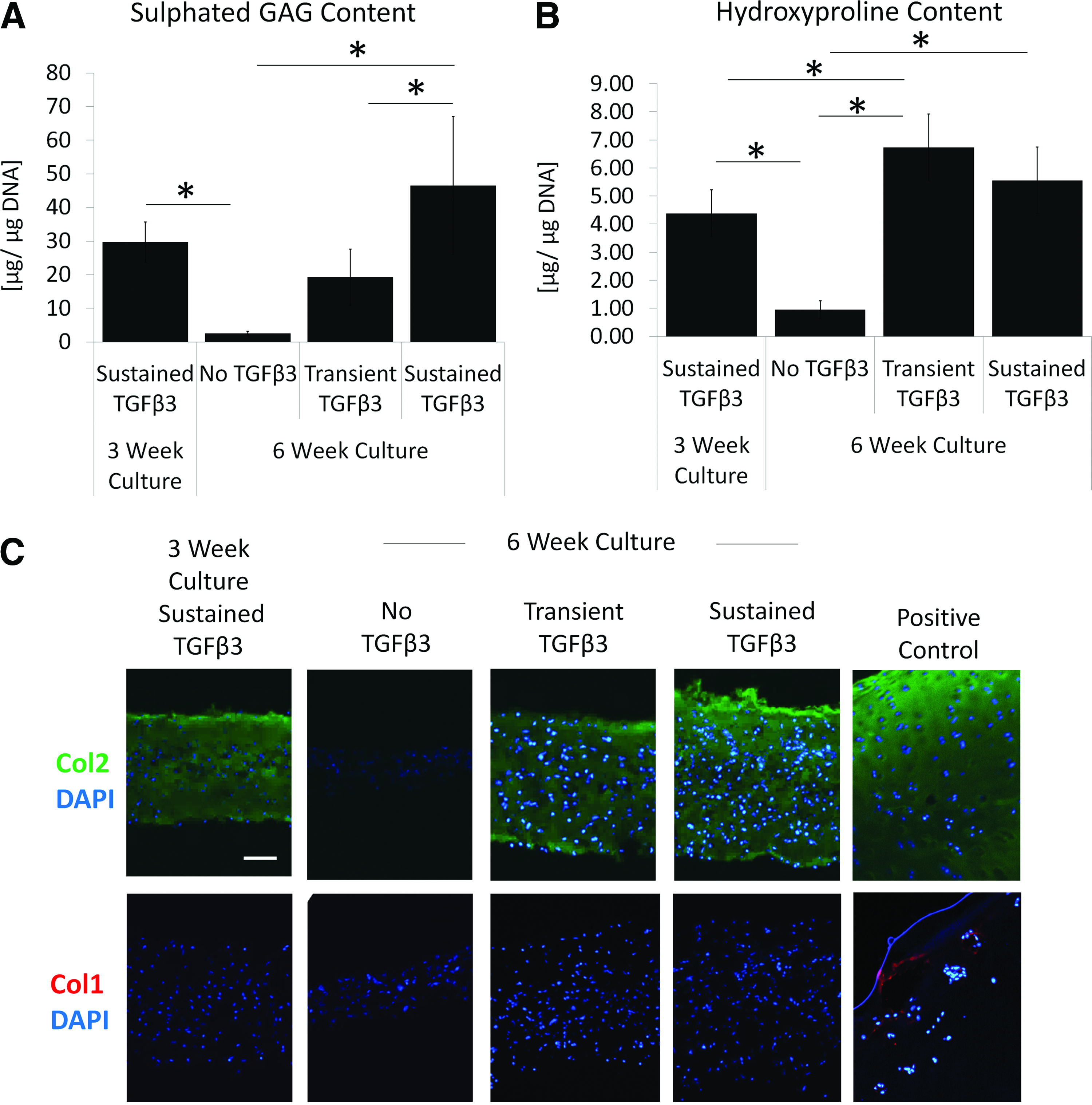
In vitro-formed tissues continue to accumulate significant amounts of Col 2 if treatment with exogenous TGFβ3 is discontinued. Passaged (P2) chondrocytes were cultured for 3 weeks in SFRM with 10 ng/mL TGFβ3 to allow for tissue formation and then cultured for an additional 3 weeks in the absence (transient TGFβ3) or presence (sustained TGFβ3) of TGFβ3 treatment. Controls were grown in the absence of TGFβ3 for 6 weeks.
Mechanical properties of in vitro-formed cartilage
No significant differences in the mechanical properties of the in vitro-formed tissues were observed with longer culture (3 vs. 6 weeks) or continued compared to transient TGFβ3 supplementation (Table 1). Although Poisson's ratio increased with sustained culture with TGFβ3 for 6 weeks, this was not a significant change. It was not possible to assess the mechanical properties of tissues formed in the absence of TGFβ as there was insufficient tissue for testing.
Passaged (P2) chondrocytes were cultured for 3 weeks in serum-free redifferentiation media with 10 ng/mL TGFβ3 to allow for tissue formation and then cultured for an additional 3 weeks in the absence (transient TGFβ3) or presence (continuous TGFβ3) of further TGFβ3 treatment. The mechanical properties of the tissues were measured for each condition. Data are represented as mean ± standard error of mean.
TGFβ3, transforming growth factor beta-3.
Tissues cultured for 6 weeks show collagen fiber orientation and protein expression consistent with superficial zone cartilage
To determine if the in vitro-formed tissues showed evidence of zonal organization with the development of a superficial zone, polarized light microscopy was used to assess collagen fiber orientation in the formed tissues. Cells cultured for 3 or 6 weeks with TGFβ3 showed collagen fiber orientation parallel to the surface (Fig. 6). Immunohistochemical studies demonstrated that at 3 weeks the expression of lubricin, also known as proteoglycan 4 (PRG4), was variable, with some tissues showing no expression and others showing discontinuous expression along the surface (Fig. 6 white arrow). No clusterin expression was detected at 3 weeks. Tissues grown for 6 weeks either continuously in the presence of TGFβ3 or transiently (for the first 3 weeks only) showed both clusterin and PRG4 at the surface of the cartilage tissue, indicative of the presence of a superficial zone. Some PRG4 expression was present at the base of 6 week formed tissues cultured continuously with TGFβ for 6 weeks (Fig. 6 white arrowhead).

In vitro-formed tissues show collagen fiber alignment and protein expression consistent with a superficial zone of articular cartilage. (Upper Row) Polarized light microscopy images of histological sections stained with picrosirius red. (Middle Row) Immunohistochemical staining for PRG4. DAPI counterstain. (Lower Row) Immunohistochemical staining for clusterin. DAPI counterstain. White arrow shows area of discontinuous PRG4 expression in 3 weeks formed tissues. White arrowhead points to area of PRG4 expression at the bottom of the construct. Scale bar = 100um. m, membrane. Color images available online at www.liebertpub.com/tea
Discussion
In this study, we showed that human passaged chondrocytes could form a continuous layer of articular cartilage tissue in vitro when cultured in high density (3D) on top of a membrane in SFRM supplemented with 10 ng/mL TGFβ3 for a minimum of 2 weeks. The tissue developed a superficial zone and was rich in Col 2 and Acan. Importantly, the tissue did not contain Col 1 and the cells did not acquire a hypertrophic phenotype as they did not express ALP activity or accumulate Col X, and it did not show any potential to mineralize. To the best of our knowledge this is the first study demonstrating this ability by passaged human chondrocytes to form cartilage tissue in the absence of scaffold and with only one growth factor, TGFβ3. The mechanism by which TGFβ3 promotes redifferentiation has not been elucidated; however, TGFβ3 is crucial for promoting chondrogenesis and cartilage tissue formation by MSC in vitro.19,32 Interestingly, it has been shown that when chondrocytes are serially passaged, they acquire plasticity resembling MSC33,34 characterized by spindled morphology and the ability to differentiate into all three of the mesenchymal lineages (adipose, chondrogenic, and osteogenic), a phenotype not found in primary chondrocytes. Therefore, TGFβ3 may be acting in a similar manner as it does in MSC chondrogenesis to promote articular cartilage tissue formation by the passaged human cells in our system. In support of this, we observed that primary chondrocytes (P0) were unable to accumulate significant amounts of collagen, when cultured under the same conditions as P2 chondrocytes.
Cartilage formation by P2 human chondrocytes required exogenous TGFβ, in contrast to bovine P2 chondrocytes that we showed in previous studies did not require the addition of any growth factors. 16 Interestingly, it has been shown recently that passaged bovine chondrocytes do require exogenous TGFβ for cartilage tissue formation in vitro, but only after passage number 3 (P3), 35 at which point these cells showed significant decreases in messenger RNA (mRNA) levels of TGFβ2 and both TGFβ receptor types 1 and 2. It is known that chondrocytes from human OA cartilage synthesize less TGFβ and possess lower levels of TGFβ receptors than chondrocytes from nondiseased cartilage.23,36,37 Therefore, TGFβ may be required for human cells as they already have impaired endogenous TGFβ signaling, which may be further affected with serial passaging.
Use of these tissues for biological repair requires that tissue will not degrade in the absence of exogenous growth factor supplementation, as intra-articular injections of TGFβ can induce adverse effects in joint and synovial tissue.38,39 Although not entirely similar to the in vivo situation, the in vitro-formed cartilage continued to accumulate significant amounts of collagen, which was predominately Col 2, when TGFβ was discontinued after 3 weeks of treatment. This suggests that the redifferentiated phenotype may be stable, and, if implanted in vivo, the cartilage tissue may continue to mature in the absence of growth factor. Of note, proteoglycan content did not continue to increase in our culture system when TGFβ was discontinued. The reason for this is not known, but importantly this did not affect the mechanical properties of the formed tissues. It is possible that the formed tissues may simply be behaving like native articular cartilage explant cultures, which lose proteoglycan when placed in vitro culture. 40 Alternatively, the absence of mechanical loading in the in vitro environment may contribute to the inability of the tissue to further accumulate sulfated proteoglycan. Previous studies have demonstrated the role of biomechanics on promoting proteoglycan accumulation. 41 Furthermore, shear forces have been shown to activate latent TGFβ produced by cells in synovial joints. 42 It may be that once the cartilage tissue is implanted in vivo, mechanical stimulation from joint movements and weight bearing may act to maintain the TGFβ and proteoglycan content and allow for further maturation and ECM accumulation. Further study is required to confirm this.
Interestingly, more than 3 weeks of culture was required for the tissues to develop a continuous superficial zone characterized by parallel collagen alignment and expression of PRG4 and clusterin. Continuous 6 weeks exposure to TGFβ3 was not required for this to occur. TGFβ has been shown to enhance PRG4 expression and accumulation by superficial zone chondrocytes in vitro.43,44 There are several potential reasons this occurred. Recently, Albro et al. showed that in bovine chondrocyte tissue constructs, media supplementation with exogenous active TGFβ1 resulted in high affinity binding and internalization by cells in the construct periphery (first 1 to 2 mm) only. 45 This has also shown to occur in the superficial zone of native bovine cartilage explant cultures. 46 This gradient of exposure to exogenous active TGFβ may be the reason the tissues in our system developed a superficial zone and may explain why PRG4 is most highly expressed and localized to the surfaces of our tissues. Although the effect of TGFβ3 on clusterin expression by chondrocytes is not known, it has been shown to upregulate clusterin mRNA and protein expression in other cell types such as lung epithelial cells47,48 and may act similarly through a TGFβ-mediated pathway in chondrocytes. In our system, 6 weeks of culture was needed for the development of a superficial zone. It may be that 6 weeks is required for sufficient amounts of PRG4 and clusterin to be accumulated so they can be detected immunohistochemically and for collagen to be detected by polarized light microscopy. Alternatively, it may be a maturation process that, similar to development, requires time. Gannon et al. demonstrated that it was not until 4 weeks of age that porcine cartilage had developed collagen fibers aligned parallel to the articulating surface. 49 Furthermore, it was not until skeletal maturity was reached that the fibers in the deep zone thickened and became aligned perpendicular to the articulating surface.
As chondrocytes passaged in the presence of serum containing media have been Food and Drug Administration (FDA) approved for use in other cartilage repair procedures and are readily obtainable using an arthroscopic approach, they would seem to be the optimal source of cells for cartilage repair. In this study, we describe a method to reverse the dedifferentiation that occurs during cell number expansion in monolayer culture and to re-establish the ability of these cells to form a continuous layer of articular cartilage-like tissue in the absence of a scaffold when grown in 3D culture in serum-free media supplemented with TGFβ3. Further study is required to determine whether the in vitro-formed cartilage tissue will be suitable to repair focal joint surface defects. Developing a method to expand cell number in the absence of serum supplementation may further enhance clinical translation.
Footnotes
Acknowledgment
This research was supported by a grant from The Arthritis Society (SOG-13-023).
Disclosure Statement
The authors have no conflicts of interest.