
Abstract
Meniscal injuries are the most frequently encountered soft tissue injuries in the equine stifle joint. Due to the inherent limited repair potential of meniscal tissue, meniscal injuries do not only affect the meniscus itself but also lead to impaired joint homeostasis and secondary osteoarthritis. The presented study compares 3D coculture constructs of primary equine mesenchymal stem cells (MSC) and meniscus cells (MC) seeded on three different scaffolds—a cell-laden collagen type I hydrogel (Col I gel), a tissue-derived small intestinal matrix scaffold (SIS-muc) and a combination thereof—for their qualification to be applied for meniscus tissue engineering. To investigate cell attachment of primary MC and MSC on SIS-muc matrix SEM pictures were performed. For molecular analysis, lyophilized samples of coculture constructs with different cell ratios (100% MC, 100% MSC, and 50% MC and 50% MSC, 20% MC, and 80% MSC) were digested and analyzed for DNA and GAG content. Active matrix remodeling of 3D coculture models was indicated by matrix metalloproteinases detection. For comparison of tissue-engineered constructs with the histologic architecture of natural equine menisci, paired lateral and medial menisci of 15 horses representing different age groups were examined. A meniscus phenotype with promising similarity to native meniscus tissue in its GAG/DNA expression in addition to Col I, Col II, and Aggrecan production was achieved using a scaffold composed of Col I gel on SIS-muc combined with a coculture of MC and MSC. The results encourage further development of this scaffold-cell combination for meniscus tissue engineering.
Introduction
M
However, horses are not only patients suffering from naturally occurring meniscus injuries, but may at the same time serve as models for human disease. Although horses lack the full knee extension seen in humans, their gait (besides sheep and goats) is considered to best resemble the human. 3 The equine standing femorotibial joint angle is approximately 150° as compared to almost 180° in humans. The cranial-caudal translocation of equine menisci during flexion and extension of the knee joint is similar to that reported in humans. 4
Moreover, it was found that hyperextension of the stifle joint may lead to the development of pathological forces and injury in the cranial horn of the equine medial meniscus, analogous to what is observed in the human posterior medial horn upon hyperflexion. 5 The gross anatomic and histologic meniscus structure of most large animals commonly serving as models for human menisci is similar to each other and the human, but a more detailed description of the equine histologic meniscus architecture would be desirable to optimize biomimicry in meniscus tissue engineering. 6
Menisci are paired C-shaped fibrocartilaginous structures within the femorotibial joint providing joint stability, lubrication during movement, protection of the joint margins, and absorption and dissipation of shock. 7 They are composed primarily of a network of collagen fibers, mainly collagen type I and II (Col I and II), interposed with meniscal cells and their extracellular matrix (ECM) consisting mainly of water, collagen, and proteoglycans. 8
In the transverse section menisci are wedge-shaped and three zones may be distinguished according to their different vascularization and neural supply: The abaxial vascular and neural zone, the middle mixed zone, and the axial avascular and aneural zone.9,10 The natural architecture of the meniscus and its highly specialized fiber arrangement allow for optimal redirection of axial forces and durability under compressive, shear, tensile, and hoop stress. 11
Resection of as little as 15–34% of the meniscus has been shown to increase contact pressure in the joint by more than 350%, demonstrating that only an intact system is able to operate at full function and even minor injuries may cause major problems. 12 To date no optimal treatment strategy to heal meniscal tissue and consequently, prevent secondary cartilage degeneration is available. Meniscal tears are currently treated by partial meniscectomy, suturing, or conservative treatment such as rest, controlled exercise, and anti-inflammatory medication. 1 However, the meniscus function cannot be fully restored thereby inevitably leading to secondary osteoarthritis.13–15 Therefore, meniscal injuries are often career-ending incidents for equine and human patients.1,10,16
Regenerative medicine holds the potential for restoring form and function of meniscal fibrocartilage after an injury. Many in vitro and in vivo studies testing potential approaches for meniscus regeneration using biological approaches, collagen scaffolds, or tissue engineering have been performed as reviewed by Moran et al., Myers et al., and Warth and Rodkey.17–19 Ideally, a meniscus implant or transplant should resemble the size, shape, vascularity, and biomechanical properties of a natural meniscus. However, to date the shape, structure, and mechanical requirements of an in vivo implant or transplant, to properly comply with the load-distributing demands in the knee, have not been achieved.16,20
The two types of scaffolds currently available for clinical use do not fulfill all characteristics of an ideal implant or transplant, hence leading to controversial clinical results.18,21 This may be due to the significantly different biomechanical properties of artificial implants and transplants compared to native menisci, which highlights the need for cellular ingrowth and formation of ECM to gain sufficient viscoelastic properties. 22
The 3D environment is an important feature of engineered meniscus constructs to facilitate a stable fibrochondrogenic phenotype and increased fibrochondrogenesis. It is well known that native meniscus cells (MC), fibrochondrocytes, dedifferentiate during 2D in vitro expansion, losing the typical meniscal and chondrogenic markers and properties.23–25 By transferring them into a 3D environment they redifferentiate and constitute a more stable fibrochondrogenic phenotype. 23 However, obtaining a sufficient number of MC for tissue engineering may be difficult. To avoid extensive culture expansion a complimentary cell source that is easy to obtain and expand in vitro is required.26,27 Coculturing MC with mesenchymal stem cells (MSC) may reduce the required quantity of MC while at the same time facilitating differentiation of MSC toward a fibrochondrogenic phenotype with less hypertrophy and increasing fibrous matrix formation..26–29 While the specific mechanism for MSC differentiation is not yet known, the beneficial effect of MSC on MC proliferation and collagen synthesis is thought to be based on their FGF1 secretion. 30
Another important component of tissue engineering is an appropriate biocompatible and biodegradable scaffold for cell seeding. For meniscus tissue engineering, natural scaffolds such as the ECM component collagen or tissue-derived scaffolds show great promise. After removal of all cellular and nuclear material tissue-derived scaffolds like decellularized porcine jejunum segments (SIS, small intestinal submucosa) and other decellularized matrices consist mainly of collagen and elastin. Despite decellularization, SIŚ mechanical properties and biological activity (SIS contains not only collagen but also GAG and growth factors) are preserved allowing cell migration, infiltration, and formation of a host-derived neomatrix after scaffold biodegradation and native tissue remodeling.28,31–35 In addition, SIS is expected to augment cellular and ingrowth. 36
Therefore, it has gained significant attention for its potential therapeutic application in tissue regeneration and defect repair spanning from bladder, bowl, esophagus, and myocardial and body wall repair over vascular grafting to musculoskeletal indications like the treatment of intervertebral discs, collateral ligaments, Achilles tendon, bone, muscle, or meniscus injuries.37–72 Also, first applications in human patients for treating rotator cuff injuries and bone defects have been reported.36,73–77 No adverse events attributable to the xenograft were observed. 36
However, the SIS matrix alone does not provide the 3D structure or biomechanical properties necessary for meniscus replacement tissue. 78 As a flat and elastic intestinal layer containing collagen in form of oriented fibers, it provides biomechanical stability (mainly longitudinal tensile strength) to the meniscus construct without having significant biomechanical strength on its own. 28
Therefore the aim of this study was to test three different 3D coculture constructs (collagen type I hydrogel, SIS-muc, and a combination of these two) loaded with different ratios of MC and MSC for their potential applicability in meniscus tissue engineering and to compare these tissue-engineered constructs with the histologic architecture of natural menisci. The resulting 3D meniscus construct combines the advantages of MC and MSC coculture at an optimized ratio with the biological activity of SIS-muc and the 3D environment of a Col I gel. Previous studies using MSC and MC cultures in 3D matrices showed promising results with respect to cell phenotype stability and production of ECM components; however, the high amount of ECM and low cell density characteristic for natural meniscus tissue and a relevant cell donor age were not stringently taken into account.26–29
In the current study the cell seeding density and matrix amount were more closely adapted to resemble natural meniscus tissue. To study the suitability of this approach for the age group typically suffering from meniscus injuries, we used only cells from adult, musculoskeletally mature donors.
Materials and Methods
Morphological structure of the equine meniscus
For this part of the study, paired lateral and medial menisci of 15 horses euthanized for reasons unrelated to this study and free of musculoskeletal disease related to the stifle were subjected to histological evaluation. Either the left or right hind limb was randomly assigned to the study, resulting in a total of 15 paired medial and lateral menisci. Age, sex, and breed of all horses were documented. Care was taken that all age groups were represented. Samples were grouped in young (0–5 years, 2 horses), middle-aged (5–20 years, 10 horses), and old (20–30 years, 3 horses).
All samples were fixed with 4% buffered formaldehyde immediately after collection and processed for paraffin embedding. Samples were dehydrated, embedded in paraffin (Shandon Excelsior; Thermo Electron Corporation), and 4–5 μm thick longitudinally oriented sections were cut using a rotating microtome (HM 355S; Histocom). Samples were taken from three anatomic regions, the anterior horn, body ( = pars intermedia), and posterior horn (Fig. 1A).
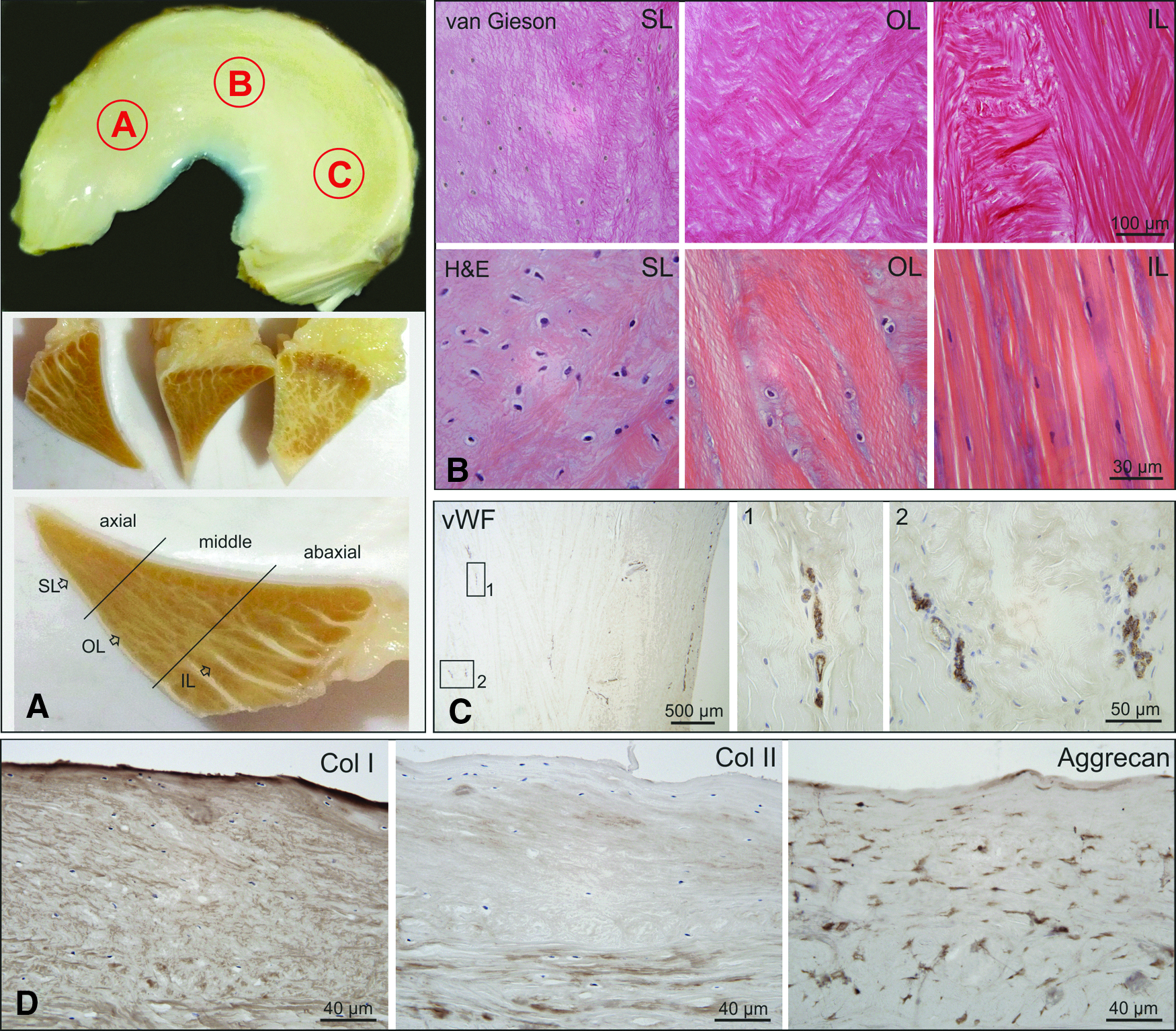
Morphological structure of the equine meniscus.
Stainings for Hematoxilin and Eosin (H&E, to describe cell shapes), Safranin O/Fast Green (counterstain), Alcian blue (to show ECM composition), Van Gieson (to show collagen fibers and fiber orientation), von Willebrand Factor (to visualize vascularization), and immunohistochemical stainings for Col I, Col II, and Aggrecan (Table 1) were performed. Results were compared between the three anatomic regions, the axial, middle, and abaxial zones (Fig. 1A), and between medial and lateral menisci. Furthermore, potential age-related differences were documented (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
Sample collection for cell isolation
For primary cell isolation, paired lateral/medial whole menisci and bone marrow aspirates were harvested from three warm blood horses (age range between 6–19 years) euthanized for reasons unrelated to this study or musculoskeletal disease. All samples were obtained immediately post mortem using aseptic technique and were processed within 14 h after collection.
Isolation and characterization of equine MCs
Whole menisci were washed in PBS (phosphate-buffered saline, without (w/o) divalent ions Ca2+ and Mg2+), diced, and digested overnight (o/n) using 5 mL collagenase solution (500 U/mL) per 1.5 g tissue. After centrifugation the remaining meniscus pieces were washed with MC medium (Dulbeccos's modified Eagle's medium [DMEM] high glucose, 10% fetal calf serum [FCS], 1 mM sodium pyruvate, 1% penicillin/streptomycin) and seeded into a T75 culture flask. During the incubation period in humidified atmosphere at 37°C and 5% CO2, MC migrated out of the meniscus pieces onto the culture dish surface. MC were used between passage 1 and 3. MC characterization was performed by immunohistochemical staining for meniscal markers: Col I, Col II, and Aggrecan.
Isolation and characterization of equine MSCs
Equine bone marrow was harvested from the horses' sterna, into 20 mL syringes preloaded with heparin (Gilvasan, 5000 IU/mL). The bone marrow aspirate was diluted with PBS+ (supplemented with divalent ions Ca2+ and Mg2) and MSC were isolated using the Ficoll-Paque density gradient method as previously described. 79 MSC were used between passage 1 and 5. MSC characterization was performed by trilineage differentiation (adipogenic, chondrogenic, and osteogenic) and via Flow Cytometry (BD FACSCalibur™) using the following markers: CD14-FITC (mouse; eBioscience), CD29-PE (mouse; BioLegend), CD44-PE (mouse; BD Bioscience), and CD45-PE (mouse; Pierce Biotechnology).
Preparation of biomaterials and cell seeding
SIS-muc consisting of decellularized porcine SIS and mucosa (muc) was prepared as described by Reboredo et al. 80 Due to the thickness of the SIS-muc derived from porcine intestinal tissue and the 3D model setup, custom-made cell crowns consisting of two concentric metal rings (1 cm2 growth area) as described in Reboredo et al. 80 were used to stabilize the SIS-muc matrix with the mucosal side facing up for cell seeding. For cell seeding on SIS-muc, 0.5 × 106 cells in 100 μL chondrogenic differentiation medium (DMEM high glucose, 50 μg/mL ascorbic acid 2-phosphate, 100 nM dexamethasone, 100 μg/mL sodium pyruvate, 40 μg/mL L-proline, 1% ITS +1; 10 ng/mL transforming growth factor β-3 (TGF-β3; R&D Systems), and 1% penicillin/streptomycin) were pipetted onto the SIS-muc matrix followed by 3 h incubation at 37°C and 5% CO2 to allow cell attachment.
For Col I gel preparation 1 volume share Col I solution (isolated from rat tail tendon dissolved in 0.1% acetic acid concentration 10 mg/mL) and 4 volume shares of gel neutralizing solution (3.75% 3 M 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and 40% FCS in 4.7 × DMEM high glucose) was used and loaded with 0.5 × 106 cells per mL gel (final concentration of Col I gel 2 mg/mL). Gelation was completed after incubation for 60 min at 37°C. For the biomaterial combination (SIS-muc + Col I gel), cell-laden Col I gel was loaded onto the cell laden mucosal side of the SIS-muc in the cell crowns.
Three-dimensional in vitro coculture constructs
To optimize the cell ratio for meniscus tissue engineering, in vitro constructs of cocultures of MC with MSC at different ratios (20% MC and 80% MSC, 50% MC and 50% MSC) or with 100% MC or 100% MSC were studied. The constructs were established using each of the three scaffolds: (1) SIS-muc, (2) Col I gel, or (3) SIS-muc + Col I gel. SIS-muc and Col I gel w/o cells served as negative controls. All 3D constructs were submerged in chondrogenic differentiation medium.
After 21 days the 3D tissue constructs were harvested and cut into four pieces with a surface of 0.25 cm2 each. These tissue construct pieces were subjected to cell metabolic activity analysis by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT; Serva) and evaluation of cell morphology and attachment by scanning electron microscopy (SEM). Immunohistochemical stainings of cryosections with primary antibodies were applied as listed in Table 1. On day 7, 14, and 21 conditioned medium aliquots were collected for matrix metalloproteinases (MMP) evaluation by zymography.
Scanning electron microscopy
SEM images were acquired using the field emission scanning electron microscope (FE-SEM) AURIGA 60 (Carl Zeiss Microscopy). For SEM, samples were fixed in 3% glutardaldehyde, pH 7.4 (Sigma-Aldrich) and in 1.5% osmiumtetroxid (Science Services), dehydrated with an ascending acetone row, critical point dried (Baltec CPD 030), and coated first with a carbon and then a platin layer.
Molecular analysis of DNA and GAG amount
For molecular analysis, lyophilized samples were digested with papain to release residual DNA and GAG. A Quant-iT™ PicoGreen® dsDNA Assay was performed as described in the instruction manual. A Blyscan™ Sulfated Glycosaminoglycan Assay was conducted as described in the manufacturer's protocol. Finally, the determined GAG amount was normalized by the measured DNA amount.
Data are presented as mean ± standard deviation. Statistical analysis was performed by nonparametric Kruskal–Wallis test using SPSS software (IBM, version 2.3) to detect differences between the used scaffolds and cell ratios of the 3D constructs. Differences were considered as statistically significant when *p < 0.05 and biologically relevant when Δp < 0.1. Follow-up analysis by homogenous subsets using SPSS software (IBM, version 2.3) was performed to detect significant differences.
Matrix remodeling
To detect MMP-2 and MMP-9 involved in the matrix remodeling process, zymography of medium supernatants on day 7, 14, and 21 was performed with the Bio-Rad system according to their instruction manual. 10% Ready Gel® Zymogram Gels containing gelatin as substrate for MMP-2 and MMP-9 were used. Coomassie Brilliant Blue staining was performed according to the manufacturer's instructions (Bio-Rad).
Results
Morphological structure of the equine meniscus
According to the location, orientation, and morphology of fibrochondrocytes and collagen fibers three different layers were defined: (1) a superficial layer (SL), (2) an outer deep layer (OL), and (3) an inner deep layer (IL) (Fig. 1A).
The SL was shown to mantle the meniscus on its femoral and tibial surface. It was characterized by a meshwork of very thin collagen fibrils, which were masked by GAG and had no particular fiber orientation (Fig. 1B). Equally, the fusiform fibrochondrocytes in the SL had no characteristic cell shape, arrangement, or orientation (Fig. 1B). Below this superficial cover was the OL (second layer) characterized by coarse Col I fibers arranged in bundles but with no distinct fiber orientation allowing them to cross one another at different angles (Fig. 1B). The inner most (third) layer IL accounted for the main portion of the equine meniscus. It was composed mainly of thick fibers with circumferential orientation (Fig. 1B). In the abaxial zone these fibers showed a strictly parallel alignment, with long fibroblast-like fibrochondrocytes arranged in lines between the collagen bundles. These bundles were interrupted only by a few radially oriented branches of connective tissue, which originated from the adjacent connective tissue and enclosed the blood vessels responsible for the blood supply of the meniscus (Fig. 1A). The axial and middle zone were additionally interwoven with collagen fibers oriented in a proximo-distal direction originating from the SL and contained round fibrochondrocytes with no particular cellular arrangement embedded in a lacunae type cavity (Fig. 1A, B).
On cross sections stained for von Willebrand Factor (vWF) to show vascularization, the presence of three zones (abaxial, middle, and axial zone corresponding approximately to the red-red, red-white, and white-white zone in human menisci) with age-dependent extent of vascular supply was confirmed. Single blood vessels marginally reached the middle zone in young animals (Fig. 1C), whereas in older (>5 years) horses only the abaxial zone was vascularized.
Col I and Aggrecan could be detected throughout the whole equine meniscus whereas Col II positive staining was typically rare within the meniscus fibrocartilaginous tissue (Fig. 1D). Only the axial and middle zone contained Col II and stained positive for GAG throughout all age groups particularly adjacent to fibrochondrocytes but also between Col I fibers (Supplementary Fig. S1A). While no age-related changes in the collagen fiber alignment were detected, the overall Col II content increased and fibrochondrocytes elongated with age.
In young horses (0–5 years) a weak Col II staining was detected only immediately around oval or round fibrochondrocytes. In older (>5 years) horses the halo of Col II containing ECM surrounding the cells increased and was also found between collagen fiber bundles (Supplementary Fig. S1A). Similarly, the proteoglycan content and distribution were found to be subjected to age dependent differences with an increase in GAG content in the abaxial zone with age and the even GAG distribution of young menisci giving way to a patchier distribution in older horses (>5 years) (Supplementary Fig. S1B).
The anatomic regions anterior horn, meniscal body ( = pars intermedia), and posterior horn had no influence on either the collagen content or fiber arrangement, GAG content or distribution, or cell shape, arrangement, or orientation of fibrochondrocytes. Differences between the lateral and medial meniscus were limited to the GAG and Col II content, both of which were more prominent in the lateral meniscus (Supplementary Fig. S1A).
Cell characterization of primary equine MCs and MSCs
Equine primary cells were successfully isolated. Equine MC showed a stretched morphology typical for elongated fibroblast-like chondrocytes (data not shown). Positive expression of Col I (Supplementary Fig. S2Aa) and Aggrecan (Supplementary Fig. S2Ac) was detected for MC whereas Col II expression (Supplementary Fig. S2Ab) was negative comparable with the isotype controls (upper right corner).
Equine MSC showed characteristic MSC spindle-shaped morphology (elongated and thin cells with a small cell body comprising a large and round nucleus; data not shown), MSC surface marker expression (CD44+, CD29+, CD14−, and CD45−; Supplementary Fig. S2Ba–d) and trilineage differentiation capacity (Supplementary Fig. S2C). Adipogenic, osteogenic, and chondrogenic differentiation potential was confirmed by Oil Red O (lipid droplets; Supplementary Fig. S2Ca), Alizarin Red (calcium hydrogen phosphate; Supplementary Fig. S2Cb), Alcian blue (GAG) and Col II expression (Supplementary Fig. S2Cc,d) compared to the nondifferentiated controls (data not shown). Isotype controls for Col II showed no positive expression. MSC showed a high potential to differentiate to osteocytes and chondrocytes, whereas adipogenic differentiation was weak.
Cell attachment and growth on SIS-muc
To investigate the cell attachment and cell growth, cells were seeded directly onto the mucosal site of the SIS-muc. After 21 days of culture in chondrogenic differentiation medium, SEM demonstrated cell attachment with a well-developed cell layer with round cell morphology on the mucosal side of the SIS-muc (Supplementary Fig. S3).
Three-dimensional in vitro coculture constructs
Different cell ratios were investigated to study coculture of MC and MSC to develop a 3D meniscus construct in comparison to mono-culture.
The generated matrix of the 3D in vitro coculture constructs was stained immunohistochemically for Col I, Col II, and Aggrecan. Isotype controls showed no positive expression. Structured matrix formation after cell seeding on SIS-muc was achieved for all cell ratios (Fig. 2 and Supplementary Fig. S4). Moreover, using the SIS-muc scaffold alone (Fig. 2 and Supplementary Fig. S4a), we observed intensive Col II deposition in the 100% MSC and (although to a lesser degree) also in the 20%MC and 80% MSC cocultures (Supplementary Fig. S4). 3D in vitro coculture constructs merely based on Col I gel showed positive expression of Col I, Col II, and Aggrecan illustrated in Figure 2 and Supplementary Figure S4b. Similar to native equine meniscal tissue (Fig. 2d, right column) Col I as predominant meniscal marker was highly expressed in the 3D constructs in contrast to a lower Col II expression observed.

Immunohistochemical analysis of 3D in vitro coculture constructs (n = 3). Immunohistochemical staining for Col I, Col II, and Aggrecan on day 21 with isotype controls in the bottom row. Schematic illustrations of the 3D constructs for coculturing MC and MSC as cell suspension directly on SIS-muc, resuspended in a Col I gel or embedded in a Col I gel on SIS-muc (SIS-muc + Col I gel) are illustrated in the upper left corner of the Col I stainings. The three left columns show the 3D in vitro coculture constructs of the 50% MC and 50% MSC ratio based on
However, shrinkage of the cell-laden Col I gel was observed for the constructs merely based on Col I. Less shrinkage of 3D constructs combining the two biological scaffolds was noticed. Moreover, the SIS-muc was firmly attached to the cell-laden Col I gel (Fig. 2c, arrows show junction of SIS-muc with Col I gel).
Cell metabolic activity of the constructs was confirmed by MTT assay (Fig. 3A). Regarding DNA quantity, no significant differences between either the biomaterials or the cell ratios were detectable (Fig. 3B). However, DNA quantification of all constructs confirmed biocompatibility of the scaffolds and cell growth.
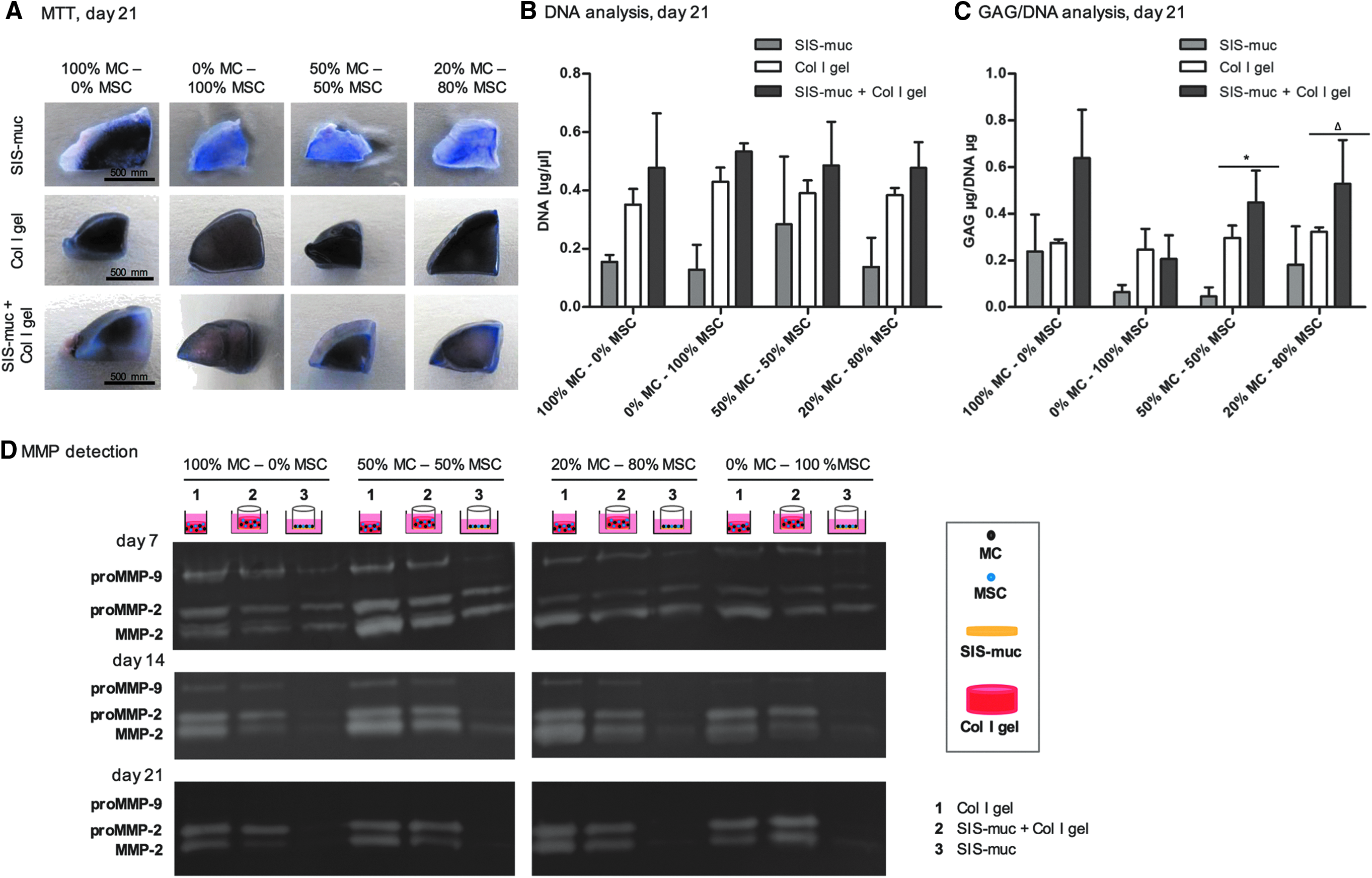
MTT staining, DNA analysis, GAG/DNA analysis, and MMP detection of 3D in vitro coculture constructs (n = 3).
All cell ratios are shown in Figure 3C for GAG/DNA quantification. Constructs based on mono-culture of MC showed GAG/DNA expression, whereas constructs based on mono-cultures of MSC showed low GAG/DNA expression. In contrast to MSC mono-cultures, increased GAG/DNA expression of the coculture of 50% MC and 50% MSC, in addition to 20% MC and 80% MSC was observed.
Moreover, the coculture constructs based on the biomaterial combination SIS-muc and Col I gel showed a significantly higher expression of GAG/DNA in contrast to constructs merely based on SIS-muc or Col I gel. This was identified and confirmed by statistical analysis (50% MC and 50% MSC-based constructs: X2 (df = 2) = 6.49, p = 0.039; 20% MC and 80% MSC-based constructs: X2 (df = 2) = 5.60, p = 0.061). Thus, significantly higher GAG/DNA values were achieved by combining the two biomaterials for 3D in vitro constructs using the two coculture ratios (50% MC and 50% MSC, 20% MC and 80% MSC). Furthermore, Aggrecan expression confirmed by immunohistochemical staining (Fig. 2 and Supplementary Fig. S4) indicated GAG presence.
Matrix remodeling of 3D in vitro coculture constructs
As an essential process of tissue engineering, matrix remodeling should be mediated by the embedded cells causing timely degradation of the 3D biomaterial and new matrix formation. Although MMPs are responsible for matrix degradation they also indicate dynamic processes like matrix remodeling. For 3D in vitro coculture constructs merely based on SIS-muc, less expression of proMMP-2, MMP-2, and proMMP-9 in medium supernatants was detected for all cell ratios compared to the 3D constructs based on the cell-laden Col I gel or the combination of SIS-muc and Col I gel (Fig. 3).
The different MC and MSC coculture ratios of the 3D constructs showed no significant influence on the detected level of proMMP-2, MMP-2, and proMMP-9. On day 7 the 3D coculture constructs based on the biomaterial combination (SIS-muc + Col I gel) showed a band for proMMP-9, proMMP-2, and MMP-2 (Fig. 3D, top). MMP detection was repeated on day 14 (Fig. 3D, middle) and day 21 (Fig. 3D, bottom). ProMMP-2 and MMP-2 expression increased over time whereas the proMMP-9 band became weaker and finally disappeared for the 3D coculture models based on the biomaterial combination). This detected change of MMP bands indicated comparable dynamic matrix remodeling for all cell ratios as shown in Figure 3D.
Discussion
When it comes to developing and testing new implants and transplants, one crucial point is the choice of appropriate animal models.
Horses are a well-accepted, well-established, and clinically relevant animal model particularly for studying joint disease.81,82 Therefore, this study was carried out using equine meniscal tissue and cells. Horses fulfill the requirements of the European Medicines Agency (EMA), the USA Federal Food and Drug Administration (FDA), and the International Society for Stem Cell Research (ISSCR) who recommend the use of large animal models to evaluate efficacy, durability, dose response, degradation, and safety before obtaining market approval of advanced therapeutic medicinal products. 83 Above that, horses are also patients who suffer from naturally occurring meniscus lesions and osteoarthritis and as such might benefit from new treatment technologies in the future.1,2,13–15,84
At first, the histologic architecture of equine menisci was comprehensively evaluated to define target structure and properties of an equine meniscus model. The results were related to the histologic architecture of human menisci and the engineered meniscus pilot construct. From a histological point of view the equine meniscus is similar to the human, with a superficial thin meshwork of fibers (SL) followed by a lamellar layer (OL) and a main central layer of circular fibers with interwoven radial fibers (IL) (Fig. 1A, B). 11
As in human menisci Col I was present throughout the entire meniscus whereas Col II positive staining was typically rare with higher concentrations in the axial zone (Fig. 1D). The fibrochondrocytes were found to be round and embedded in lacunae-like cavities in the axial and middle zone of the meniscus while in the abaxial zone they showed a more elongated fibroblastic shape (Fig. 1B) as described similarly for human menisci. 6 Also, vascularization could be shown to be similar to the human counterpart.9,10 It reached the middle zone (red-white zone) only in young horses (Fig. 1C), whereas in older (>5 years) horses only the abaxial zone (red-red zone) was vascularized. Col II and GAG content fibrochondrocyte morphology was shown to be subjected to age-related changes (Supplementary Fig. S1).
Meniscus injuries are a significant clinical challenge due to the poor healing potential of the avascular axial and middle zone.85,86 As current treatment regimes do not yet offer ideal long-term solutions, tissue engineering is considered a promising prospect for meniscus regeneration. A variety of cells and scaffolds for meniscus regeneration extending from autologous tissues such as perichondrium or tendon and natural scaffold materials such as collagen, fibrin, hydrogels, polymer-hyaluronan, silk to synthetic materials such as nanomaterials, polycaprolacton-urethane, Teflon, or polyurethane are being investigated.87–91 However, although research efforts are strong no sufficiently convincing meniscus transplant has yet been established.
In this study, to generate a bioartificially engineered tissue comparable to native equine meniscal tissue, 3D coculture constructs based on a SIS-muc-derived scaffold, a cell-laden Col I gel, and a combination thereof were compared.
SIS-muc has gained approval for clinical implantation in the United States, Europe, and Australia and has been utilized as a scaffold in multiple attempts to regenerate different organs and tissues.28,37–72,92,93 As meniscus scaffold SIS-muc does not only provide longitudinal tensile strength and collagen but also growth factors such as basic fibroblast growth factor, vascular endothelial growth factor, and TGF-β.28,33–35 It is biodegradable and promotes cell proliferation and differentiation. 48 Several researchers have already tested SIS-muc to induce regeneration of meniscal defects or to provide an implant or transplant with contradictory results.37–40,94 Reasons for the diverging outcome may be the lack of appropriate shape and mechanical properties due to the unstable nature of the biomaterial. Fibrocartilage is a tissue with a high percentage of ECM,10,95 which the SIS-muc alone cannot offer. Above that, on the SIS-muc scaffold alone, we detected intensive Col II deposition similar to the pattern seen in old horses with altered meniscus morphology (metaplasia toward more hyaline-like cartilage), with 100% MSC and (although to a lesser degree) also 20% MC/80% MSC (Supplementary Fig. S4). The low amount of ECM offered by SIS-muc may have caused cell cluster formation (similar to pellet culture for chondrogenic differentiation) within the tissue, which may have resulted in a more cartilage than meniscus like differentiation in these areas, making SIS-muc alone a less than ideal scaffold choice.
On the other hand, in accordance with other studies, we observed contraction of the low-density collagen hydrogel scaffold when using only Col I gel.96,97 However, the contraction was altered by combining Col I gel with SIS-muc. The overall width and length of the constructs was maintained and only the height of the construct was reduced. This is most likely attributed to the longitudinal tensile strength provided by SIS-muc thereby encouraging a combination of these two biomaterials to prevent shrinkage. 28 In the future this effect may be even strengthened by using high-density collagen gels, which were shown to be suitable as injectable, moldable scaffolds seeded with meniscal fibrochondrocytes, but with reduced contraction. 97 However, the one-dimensional shrinkage, causing the reduction in height only might lead to better collagen fiber alignment.
Furthermore, higher the MSC portion in the 3D coculture constructs studied here the less shrinkage of the compared scaffolds was observed. This finding is interesting because it is in contrast to McCorry et al. who reported more contraction of high density Col I gels seeded with MSC than with MC, although using culture medium without TGF-β. 26 As the gel shrinkage is likely caused by cells with fibrocytic (i.e., undifferentiated MSC) or fibrochondrocytic (i.e., MC) properties due to their high actin activity this disparity may be explained by the differences in TGF-β content of the culture medium. In our study, the influence of TGF-β may have driven the MSC toward a more chondrocyte-like phenotype with less actin activity.
Nonetheless, during 21 days of culture matrix remodeling of the combined scaffold to a fibrocartilage phenotype was observed and further corroborated by a chronological change of proMMP-2, MMP-2, and proMMP-9 expression. Although MMPs are involved in matrix degradation they also indicate dynamic processes like matrix remodeling and were interpreted as such in this study. 98 3D constructs comprising Col I gel resulted in higher proMMP-2, MMP-2, and proMMP-9 levels compared to 3D constructs merely based on SIS-muc. This suggested a higher matrix remodeling degree of the cell-laden Col I gel-based 3D constructs as also confirmed by GAG/DNA analysis.
The GAG amount was increased by addition of the cell-laden Col I gel suggesting production of ECM by fibrochondrocytes (Fig. 3C). The GAG/DNA production of the coculture constructs based on SIS-muc plus Col I gel and just Col I gel reached levels in closer approximation to MC mono-culture than 100% MSC (Fig. 3C). Although 100% MC would of course closest mimic the natural conditions in a native meniscus and showed the overall highest GAG/DNA production in this study, adding MSC can reduce the required quantity of MC. Thus, by adding MSC fewer passages of fibrochondrocytes will be needed to reach a clinically or experimentally relevant number of cells, which in turn reduces dedifferentiation. As reviewed by Taylor et al., 99 MC are not abundantly available due to the low cell density in the meniscal tissue. In addition, MC have to be either harvested from dead horses immediately post mortem or for autologous use from healthy meniscus areas of horses undergoing knee surgery, which would potentially inflict donor site morbidity and further damage to the already harmed knee. In contrast, MSC can easily be harvested from different tissues for autologous or allogeneic transplantation with relatively little risk and donor site morbidity. Furthermore, MSC are known to provide paracrine trophic factors as essential conductors for immune modulation, differentiation, cell proliferation, and ECM synthesis.100–103 However, they tend to undergo hypertrophy when chondrogenically differentiated. Coculturing primary MC with MSC has been shown to increase fibrochondrogenesis of MSC with a stabilized fibrochondrogenic phenotype and has been repeatedly suggested to support healing and remodeling.26–29,104,105
In this study, coculture of MC obtained from whole menisci and MSC at different cell ratios were tested. While the location of MC harvest (axial or abaxial) may influence their performance in meniscus tissue engineering, studies have shown equivalent differentiation potential toward fibrochondrogenic phenotype and ECM production of axial and abaxial MC.106–109 Additionally, in a study particularly evaluating the influence of outer (axial) and inner (abaxial) MC on MSC it was demonstrated that both interact similarly and synergistically with MSC. However, as abaxial MC were shown to decrease hypertrophic differentiation of MSC and axial MC maintain a more chondrogenic phenotype, we decided to use whole menisci in our study to utilize both properties.29,108
In our study we chose a low cell seeding density (0.5 × 106 cells per mL Col I gel and 0.5 × 106 cells on the SIS-muc with 1 cm2 growth area) reflective of the hypocellular native meniscus. In contrast to other studies with higher seeding density (25 × 106 cells per mL Col I gel or 0.9 × 106 0.5 cm2 growth area), we achieved the best results with MC monoculture.26,28 This may be explained by the lower cell density and therefore probably lower cell interaction in the constructs used in our study as compared to previous studies performed with higher seeding densities. Another factor contributing to this difference may be the pellet culture used in other studies, which does not provide ECM and thus does not reflect the environment of a native meniscus.27,29
Alternatively, the TGF-β contained in chondrogenic differentiation medium, which was shown to induce hypertrophy of MSC and MC, may have influenced the results. 28 The TGF-β supplemented medium may have had a stronger hypertrophic influence on MSC than on differentiated MC, leading to superior results of the 100% MC group. Cui et al. therefore suggested that by coculturing 50% MC with 50% MSC or 75% MC with 25% MSC fibrochondrogenesis may be achieved even without the addition of TGF-β thereby bypassing the risk of inducing the hypertrophic pathway. 27 On the other hand, TGF-β may be beneficial for the regeneration of the most axial meniscus zone, which is naturally containing more Col II. 110
Also, cell donor age may lead to diverse results between studies. The equine MC and MSC used in this study were all derived from adult and skeletally mature donors (11–19 years old) to represent an age group that would typically suffer from meniscus injuries and require a transplant. In other studies performing coculture experiments of MC and MSC on different kinds of scaffolds cells from very young donors (3 days old bovids or 3 months old pigs) were used.26,28 Young cells may have a different inherent differentiation potential and spectrum of factors they release compared to cells from skeletally mature animals therefore leading to different results upon cell interaction.
Taken together, the equine meniscus is anatomically and histologically similar to the human, therefore suggesting the horse as a potentially good model animal in particular when it comes to researching naturally occurring disease. The results suggested that primary equine MC seeded on a scaffold combining the cell-laden Col I gel and the decellularized SIS-muc as potentially suitable construct may lead to the most native like meniscus phenotype. However, due to the limited access to sufficient numbers of MC for tissue engineering MSC represent a valuable complimentary cell source to compensate a potential lack of MC. By coculturing MC with MSC on SIS-muc and Col I gel results in closer approximation to MC mono-cultures could be achieved than using 100% MSC while in addition offering stable and sustainable in vitro culture properties.
Conclusion
In this study, primary equine MC and MSC were successfully cultured in a combined scaffold composed of Col I gel on a decellularized porcine jejunum segment SIS-muc as confirmed by MTT test, GAG/DNA production analysis and histologic evaluation for meniscus-like ECM production. Coculture of MC and MSC led to a meniscus phenotype with promising similarity to native meniscus tissue in its GAG/DNA expression and Col I, Col II, and Aggrecan production while in addition offering stable and sustainable in vitro culture properties. These results encourage continuation of this promising approach on the way to develop a scaffold-based personalized treatment for meniscal lesions with modulation capability, however, without addition of TGF-β to the culture medium.
In the current consistency the construct is soft and plastic (gel-like consistency), which makes it predestined to be applied as patch for fixing meniscal tears of different size and shapes. However, by implementing fibers and/or high-density collagen together with mechanical stimulation a construct more closely mimicking the mechanical properties of a native meniscus can be fabricated, which may be used for (partial) meniscus replacement. The horse proved to be a good model for the continuation of the study.
Footnotes
Acknowledgments
This work was supported by grants from Interdisciplinary Clinical Research Center (IZKF) Würzburg, Projects No D-101.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.