
Abstract
Borosilicate bioactive glasses (BBGs) have shown the capacity to promote higher formation of new bone when compared with silicate bioactive glasses. Herein, we assessed the capacity of BBGs to induce osteogenic differentiation of bone marrow mesenchymal stem cells (BM-MSCs) as a function of their substituted divalent cations (Mg2+, Ca2+, Sr2+). To this purpose, we synthesized BBG particles by melt quenching. The cell viability, proliferation, and morphology (i.e., PrestoBlue®, PicoGreen®, and DAPI and Phalloidin stainings, respectively), as well as protein expression (i.e., alkaline phosphatase, ALP; osteopontin, OP; and osteocalcin, OC), of BM-MSCs in contact with BBGs were evaluated for 21 days. We observed an enhanced expression of bone-specific proteins (ALP, OP, and OC) and high mineralization of BM-MSCs under BBG-Mg and BBG-Sr-conditioned osteogenic media for concentrations of 20 and 50 mg/mL with low cytotoxic effects. Moreover, BBG-Sr, at a concentration of 50 mg/mL, was able to increase the mineralization and expression of the same bone-specific proteins even under basal medium conditions. These results indicated that the proposed BBGs improved osteogenic differentiation of BM-MSCs, therefore showing their potential as relevant biomaterials for bone tissue regeneration, not only by bonding to bone tissue but also by stimulating new bone formation.
Introduction
T
Recently, borosilicate bioactive glasses (BBGs) have attracted interest in bone tissue engineering.1,7,8 BBGs have shown the capability to improve new bone formation when compared with silicate-based BGs.9,10 They present controllable degradation rates and have high compositional flexibility that potentially allows BBGs to be tailored with enhanced osteogenic and angiogenic properties, as well as with antibacterial capacity.11,12
Huang et al. showed that a glass network comprising borosilicates have more controllable conversion rates to HA. 13 This has also been demonstrated to occur in vivo. 14 As a matter of fact, the addition of borate to the glass network can also be beneficial for bone healing, as well as formation, and maintenance of new bone while supporting cell osteogenic differentiation. 15 Frequently, it has been associated with the increase in bone resistance to fracture.16,17
The compositional flexibility of BBGs allows the incorporation of inorganic divalent cations, such as, Mg2+, Ca2+, and Sr2+, which play a key role in bone metabolism. For instance, Mg2+ increases bone formation rate as well as stimulates bone cell adhesion, increasing their stability.18,19 Ca2+ is known to be essential during the apatite formation process, being also favorable to osteoblast proliferation, differentiation, and mineralization of the extracellular matrix (ECM). 20 Sr2+ also has bone therapeutic potential. Different studies evidenced its beneficial effects on bone cells and bone formation in vivo,21,22 being even used for the treatment of osteoporosis. 23
In general, BGs are known to be osteoinductive materials capable of stimulating the function and osteogenic differentiation of bone and stem cells without any additional supplementation. Findings from Fu et al. suggested that the borosilicate 13-93B1 scaffolds supported proliferation and function of osteogenic murine osteocyte-like cell line MLO-A5, 15 while Gentleman et al. demonstrated that Sr2+-substituted BGs stimulated osteoblast metabolic activity promoting cell proliferation and ALP activity. 24
More significantly, Santocildes et al. demonstrated that Sr-containing BGs appeared to be capable of promoting osteoblastic differentiation in a proportion of bone marrow mesenchymal stem cells (BM-MSCs) that were in some way precommitted to this lineage. 25 Liang et al. showed that borate glasses support the attachment and differentiation of human BM-derived MSCs and human mesenchymal stem cell-derived osteoblasts. 26
The present study aims to fabricate three substituted BBGs to be used for bone tissue regeneration. We evaluated the impact of incorporating different divalent cations (i.e., Mg2+, Ca2+, and Sr2+) into the BBGs on their ability to induce BM-MSCs (known to differentiate into mesodermal lineage cells, such as osteoblasts, osteoclasts, and osteocytes 27 ) to proliferate, differentiate, and mineralize the ECM while promoting bone formation. 11
Experimental
Preparation of BBGs
The BBGs of general formula 0.05Na2O · xMgO · yCaO · (0.35-x-y)SrO · 0.20B2O3 · 0.40SiO2 (molar ratio, where x, y = 0.35 or 0.00, and x ≠ y) were synthesized by melt quenching. Suitable amounts of silica (SiO2; Macherey-Nagel, Germany), boron oxide (B2O3; Alfa Aesar, Germany), sodium bicarbonate (NaHCO3; Sigma-Aldrich, Australia), and magnesium oxide (MgO; Sigma-Aldrich, Portugal) or calcium carbonate (CaCO3; Sigma-Aldrich, Portugal) or strontium carbonate (SrCO3; Sigma-Aldrich, Portugal) were thoroughly mixed with the addition of ethanol in a porcelain pestle with the help of a mortar, vacuum dried overnight, and transferred to a platinum crucible.
After being entirely dried, each batch was heated to 1450°C in air for 1 h and subsequently the melt was quickly poured into cold water to form the glass frit. Afterward, the as-quenched glasses were ground in an agate mortar (RETSCH, Germany) and sieved to a particle size <63 μm. Before the in vitro tests, BBG-Mg (0.05Na2O · 0.35 MgO · 0.20B2O3 · 0.40SiO2), BBG-Ca (0.05Na2O · 0.35CaO · 0.20B2O3 · 0.40SiO2), or BBG-Sr (0.05Na2O · 0.35SrO · 0.20B2O3 · 0.40SiO2) was weighted, dried, and sterilized at 160°C for at least 2 h.
Morphology and chemical composition of BBGs
The morphology of synthesized BBGs was observed by scanning electron microscopy (SEM, model S360; Leica Cambridge, United Kingdom) equipped with energy dispersive X-ray spectroscopy (SEM/EDS link-eXL-II) for the determination of the surface chemical composition.
Isolation and expansion of mesenchymal stem cells
BM-MSCs were isolated from bone marrow of 4–5-week-old male Wistar rats according to the method established by Maniatopoulos et al. 28 and recently proposed by Santocildes et al. 25 BM-MSCs were expanded in basal medium consisting of Dulbecco's modified Eagle's medium (Sigma-Aldrich, United Kingdom), supplemented with 100 U/mL penicillin (Sigma-Aldrich, United Kingdom) and 1 mg/mL streptomycin (Sigma-Aldrich, United Kingdom). Cells were cultured at 37°C in an atmosphere of 5% CO2.
Before the in vitro studies, BM-MSCs, at passage 2, were harvested and seeded into 24-well plates at a density of 2 × 104 cells per well. Cells were cultured in the presence of the BBGs at concentrations of 20 and 50 mg/mL for 7, 14, and 21 days under static conditions. The BM-MSCs cultured in the absence of BBGs were used as negative control and in the presence of 45S5 bioglass® as positive control.
The BBGs at the desired concentrations and 45S5 bioglass were deposited on top of the cells in cell culture inserts with porous membranes (0.4 μm ThinCerts™ Cell Culture Inserts; Greiner, Germany), as used in previous works. 29 The ThinCerts were used as an inert platform to support the glass particles under cell culture conditions. All BBG compositions and controls were cultured in basal and osteogenic differentiation media (basal medium supplemented with 50 μg/mL ascorbic acid, 10 mM β-glycerophosphate, and 10−8 M dexamethasone). Figure 1 presents a schematic of the experimental design.
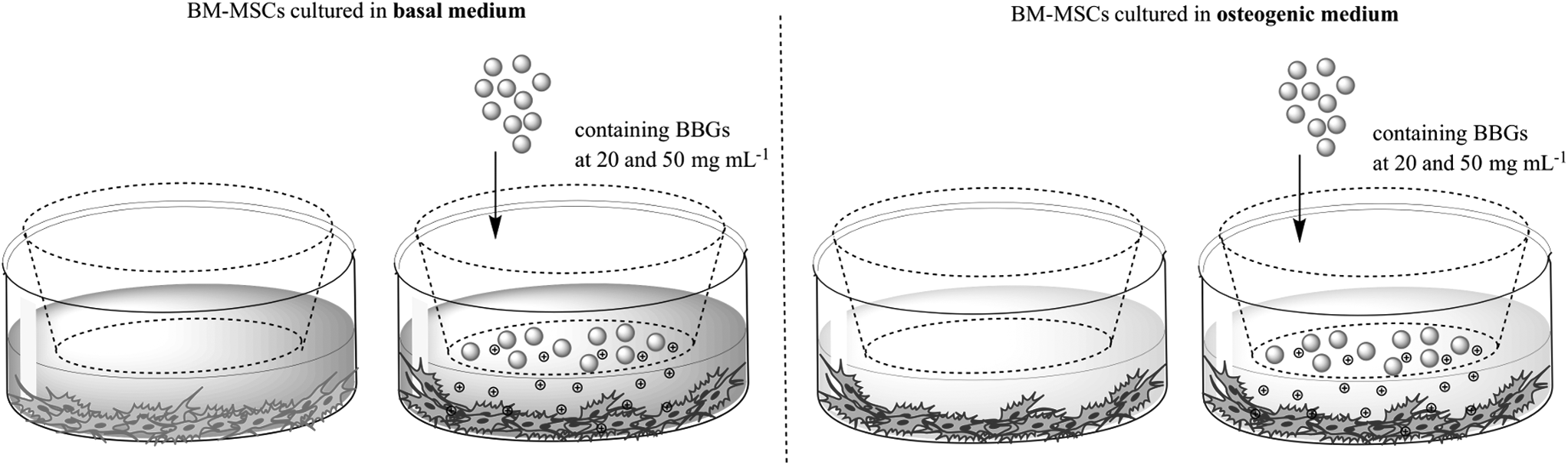
Schematic of the experimental design.
Potential cytotoxic effect of BBG dissolution on BM-MSCs
Cell viability and proliferation
The PrestoBlue reagent (Fisher Scientific, United Kingdom) is a resazurin-based solution that is reduced to resorufin by viable cells, which can be detected fluorometrically. The cell viability assay was executed according to the manufacturer's instructions. In brief, the PrestoBlue reagent was added to a final concentration of 10% to the wells and the cells were incubated for 1 h at 37°C. Afterwards, 200 μL samples of the culture medium were removed and placed in 96-well plates and the resorufin fluorescence was quantified spectrophotometrically using a plate reader (Tecan Infinite M200). Fluorescence was determined at an excitation wavelength of 560 nm and emission wavelength of 590 nm.
The metabolic activity was presented in fluorescence values and compared with the control (cell cultured in the absence of glass particles under basal medium conditions).
The PicoGreen dsDNA reagent (Invitrogen) is an ultrasensitive fluorescent nucleic acid dye for quantification of double-stranded DNA (dsDNA) in solution. This assay enables the measurement of cell proliferation. After each culturing period, the cell monolayers were washed with phosphate-buffered saline (PBS) and then incubated at 37°C for 3 h, followed by a freezing step at −80°C for at least overnight in ultrapure water (1 mL) to ensure cell lysis. The assay was performed according to the manufacturer's protocol, and fluorescence was determined at an excitation wavelength of 485 nm and emission wavelength of 528 nm. The DNA concentration was presented in μg/mL and compared with the control (cell cultured in the absence of glass particles under basal medium conditions).
Cell morphology and distribution
After each culturing period, the cells grown in tissue culture coverslips were washed with PBS and fixed with 4% formalin solution (0.5 mL) for 15 min at room temperature (RT). The cell layers were then washed with PBS, containing 0.2% Triton X, for 2 min. After the fixation and permeation steps, the cell monolayers were washed again with PBS and stained with 4,6-diamidino-2-phenyindole dilactate (1:1000 DAPI; Sigma, United Kingdom) for 2 min at RT and phalloidin-tetramethylrhodamine B isothiocyanate (Sigma, United Kingdom) for 1 h at RT. Finally, the cells were washed and observed using an Axioplan 2 fluorescence imaging microscope with a digital camera QIC AM 12-bit (Zeiss, United Kingdom).
Osteogenic capacity of BBGs on BM-MSCs
Alkaline phosphatase quantification
The concentration of alkaline phosphatase (ALP) was determined for all the culture time periods, using the lysates used for DNA quantification. Briefly, the ALP quantity was assessed using the ALP, Diethanolamine Detection kit (Sigma-Aldrich, United Kingdom) in which p-nitrophenyl phosphatase (pNPP) solution is hydrolyzed by ALP to yellow free p-nitrophenol. In brief, a buffered pNPP solution was prepared and equilibrated at 37°C. Afterward, 2% (v/v) of sample or control was added. Immediately after mixing, the absorbance was read at 405 nm in a plate reader (Tecan Infinite M200) for ≈5 min. An ALP standard solution was used as control and buffer as blank. The units were calculated according to the following equation:
where df = dilution factor; VF = volume of final solution; 18.5 = millimolar extinction coefficient of pNPP at 405 nm, and VE = volume of samples/ALP standard solution. ALP activity was calculated by normalizing ALP concentration per DNA concentration for each condition and time point.
Alizarin red staining
After 21 days of culture, the cells grown in tissue culture coverslips were fixed in 70% ice-cold methanol at −20°C at least for 30 min. The cell layers were then washed with PBS and dried overnight. Afterward, cells were stained with Alizarin red solution [342 mg of Alizarin red (Sigma-Aldrich, United Kingdom) in 25 mL of distilled water and the pH was adjusted to 4.1 with 10% ammonium hydroxide (Sigma-Aldrich, United Kingdom)] for 10 min. Afterward, the coverslips were washed with distilled water, dehydrated in an acetone/xylene (Sigma-Aldrich, United Kingdom) mixture, and mounted using an aqueous mountant.
The stained constructs were observed under an optical microscope (BX51; Olympus Corporation, United Kingdom) and images were captured by a digital camera (DP70; Olympus Corporation, United Kingdom). The BM-MSC morphology and mineral deposition were also observed using SEM (model S360; Leica Cambridge, United Kingdom) equipped with energy dispersive X-ray spectroscopy (SEM/EDS link-eXL-II) for the determination of the surface chemical composition.
Immunodetection of bone-specific proteins
Osteopontin (OP) and osteocalcin (OC) protein expression of BM-MSCs was assessed by immunoassay technique to evaluate their osteoblastic differentiation. The procedures were executed according to the manufacturer's instructions. The concentrations of OP and OC were determined for all the culture time periods, using the lysates used for DNA quantification.
The OP quantitative determination was performed using Mouse/Rat Osteopontin Quantikine ELISA Kit (R&D Systems, United Kingdom). In brief, 50 μL of assay diluent RD1W and 50 μL of standard (2500–39 pg/mL), control, and samples were added and the plate incubated for 2 h at RT. After four washing steps and drying, 100 μL of Mouse/Rat OP Conjugate was added and incubated for 2 h at RT. The sandwich complex was washed four times and allowed to react with 100 μL of substrate solution before adding 100 μl of stop solution. Finally, the optical density was determined at 450 nm and the concentration of OP obtained from a standard curve plot.
OC quantitative determination was performed by the use of Rat Gla-Osteocalcin High Sensitive EIA kit (Takara Clontech, Japan). In brief, 100 μL of samples and standard solution (16–0.25 ng/mL) were incubated for 1 h at 37°C with the capture antibody, rat osteocalcin C-terminus-specific antibody. After OC capture and three washing steps, 100 μL of the enzyme-labeled antibody (GlaOC4-30) specific to Gla-OC was incubated for 1 h at RT. The sandwich complex was washed four times and allowed to react with 100 μL of substrate solution for 10–15 min. Finally, after adding the stop solution, the optical density was determined at 450 nm and the concentration of OC obtained from a standard curve plot. OP and OC content was calculated by normalizing OP or OC concentration per DNA concentration for each condition and time point.
Statistical analysis
Results are expressed as mean ± standard deviation with n = 3 for each sample. Error bars represent standard deviations. The data were analyzed by nonparametric statistics: Kruskal–Wallis test (p < 0.0001), followed by a Dunn's Multiple Comparison test. ***p < 0.001; **p < 0.01; *p < 0.05 in respect to the control.
Results and Discussion
Morphology of BBGs and their chemical composition
BBG frits were successfully obtained by melt quenching and ground in a controlled manner. Figure 2a–c shows the SEM/EDS analysis of BBGs, which exhibited an angular shape with low sphericity and confirmed the successful incorporation of different modifier divalent cations (i.e., Mg2+, Ca2+, and Sr2+). The composition of BBGs was confirmed in a prior work by the use of X-ray fluorescence and follows the general formula 0.05Na2O · xMgO·yCaO·(0.35-x-y)SrO ·0.20B2O3·0.40SiO2 (molar ratio, where x, y = 0.35 or 0.00, and x ≠ y). 30 Moreover, studies after immersion in SBF, showed that the studied BBGs are bioactive due to the formation of bone-like apatite structures onto their surface and the constant release of ions to the reaction media over time. 29

SEM/EDS micrographs of BBGs,
In vitro biological evaluation
Osteoblast differentiation can be divided into three stages: cell proliferation, ECM synthesis and maturation, and ECM mineralization, each with a cellular characteristic behavior. 31
Potential cytotoxic effect of BBG leachables on BM-MSCs
The effect of BBG concentration on cell viability and proliferation was studied in a previous work.29,30 It showed that a concentration between 20 and 50 mg/mL did not significantly affect cell viability and proliferation. On the other hand, Romero 32 studied the osteogenic response of BM-MSCs to strontium-substituted bioactive glasses (SrBG) and observed that 20 mg of Sr50BG promoted the osteoblastic differentiation of BM-MSCs. Based on these results, we decided to use BBG concentrations of 20 and 50 mg/mL.
Herein, we cultured BM-MSCs in basal and osteogenic differentiation media for 7, 14, and 21 days under static conditions, in the presence and absence of BBGs, to evaluate their biological activity. The cellular metabolism and proliferation were evaluated by quantifying the conversion of resazurin to resorufin by viable cells and the amount of double-stranded DNA (live cells) in the culture wells, as well as their morphology.
Fluorescence microscopy images showed the morphology of BM-MSCs in culture containing BBGs or 45S5 bioglass (Fig. 3). The adhered BM-MSCs exhibited a well-spread morphology, exhibiting cell-to-cell contacts in a comparable manner on the BBG-conditioned cultures as in the control experiment. While BM-MSCs cultured with 45S5 bioglass presented a round shape for the last time point (21 days), suggesting cell death, at the same time point, especially for cultures under osteogenic differentiation media (e.g., BBG-Sr), there were distinguishable well-spread polygonal-shaped cells, suggesting osteoblast-like morphology.
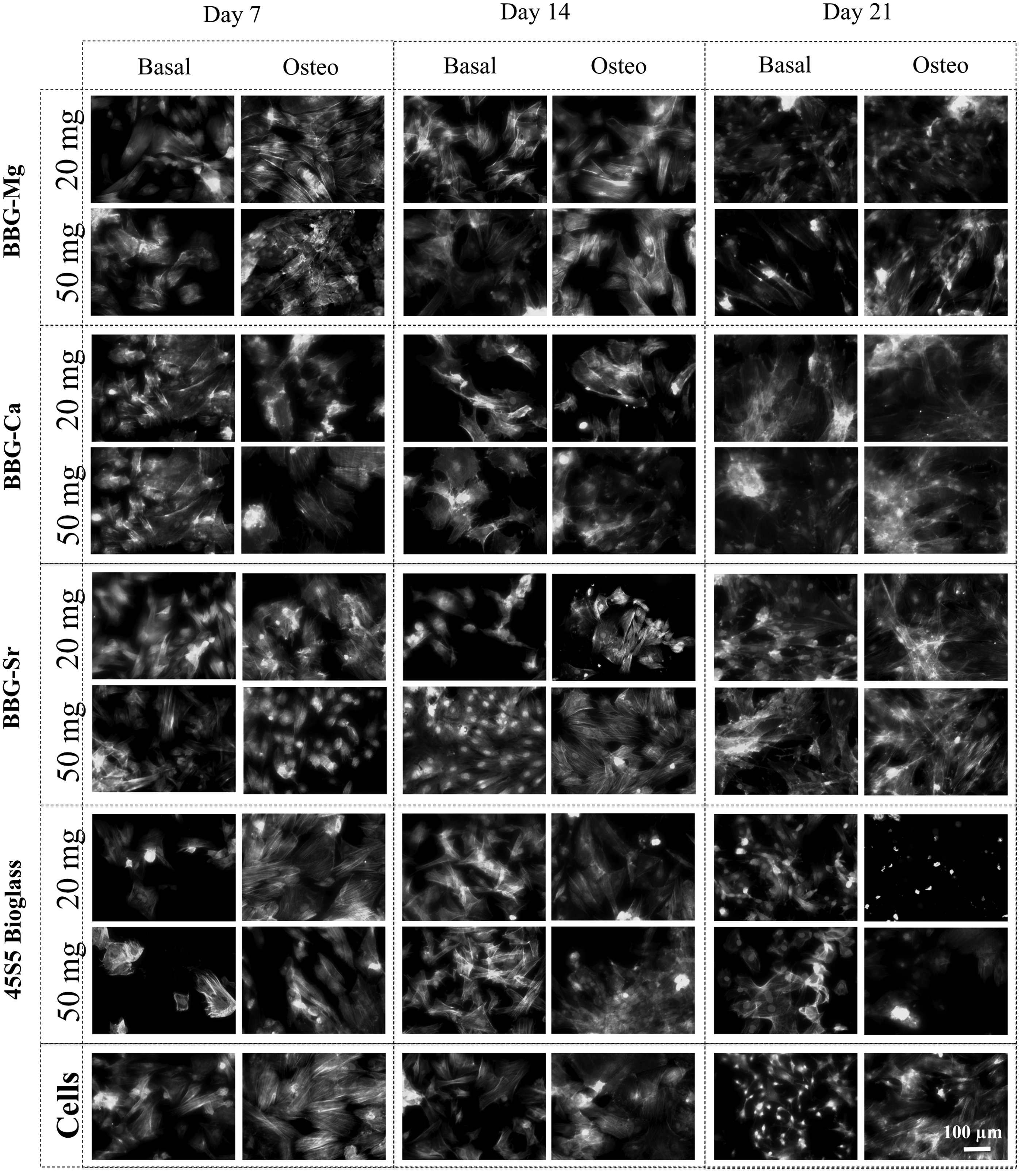
BM-MSC morphology observed by fluorescence microscopy after 7, 14, and 21 days of culture with BBGs either under basal or osteogenic culture medium. Each sample was incubated at two different concentrations (20 and 50 mg/mL). Cells cultured with basal and osteogenic media were used as negative control and 45S5 bioglass® incubated with medium was used as positive control. Nuclei stained blue by DAPI; Actin stained green by Phalloidin. BM-MSC, bone marrow mesenchymal stem cell.
From the PrestoBlue data (Fig. 4a, c, e, and g), it is possible to observe that the metabolic activity of BM-MSCs increased over the 21 days of culture. In the first 7 days of culture, the BM-MSCs under conditioned cultures presented reduced viability in respect to the cell culture under basal condition (negative control), especially in the presence of BBG-Ca, which may be due to the initial burst release of ions to the media. In contrast, from day 14, BM-MSC cultures under osteogenic media displayed a reduction of metabolic activity with respect to the control (absence of glass particles and cells cultured under basal medium). It is noteworthy that BM-MSC cultures at day 21 with 45S5 bioglass under osteogenic media showed very low metabolic activity when compared with the control experiment (osteogenic media), being consistent with the cell death observed by fluorescence microscopy (Fig. 3).
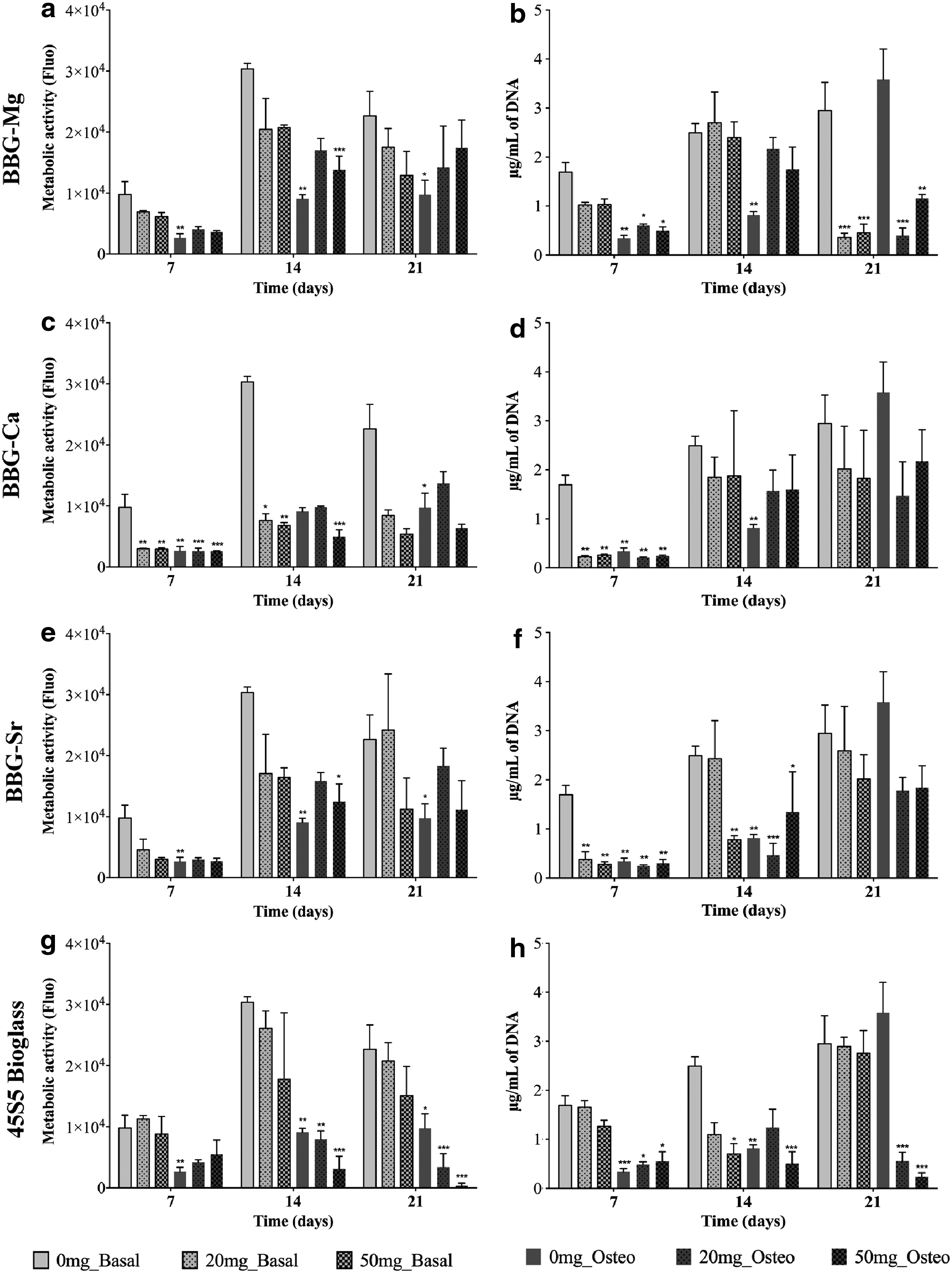
Metabolic activity (PrestoBlue® assay) and proliferation (PicoGreen® assay) of BM-MSCs cultured either in basal or osteogenic medium in the presence of different concentrations (20 and 50 mg/mL) of BBG-Mg
Complementary to the viability analysis, cellular proliferation was also assessed by measuring the total cell DNA (Fig. 4b, d, f, h). The PicoGreen data showed an increase in the number of BM-MSCs over time. However, under osteogenic media, the cells presented lower proliferation rates than cultures under basal media from day 14. Noteworthy is the prominent reduction of BM-MSCs for cultures with BBG-Mg (osteogenic and basal media) and 45S5 bioglass (osteogenic media) at day 21. To emphasize, BBG-Sr and 45S5 bioglass under osteogenic media showed a significant reduction in cell proliferation at day 14 when compared with cells cultured under basal media, suggesting an alteration of biological behavior. 33
Regarding the effect of concentration, there were no potential toxic effects with time except in the case of BM-MSCs cultured in the presence of BBG-Mg (osteogenic and basal media) and 45S5 bioglass (osteogenic media) at day 21, where a large reduction in cell number was observed in relation to the control (basal media).
Alkaline phosphate quantification
It is commonly accepted that ALP is a key player in the process of osteogenesis, being ALP known to be involved in early stages of normal and pathological calcification.34–36 In general, an increase of ALP activity is correlated with osteogenesis, increasing during the bone formation stage. 37 Not surprisingly, the ALP quantification data (Fig. 5) showed a significantly higher ALP activity when cells were cultured under osteogenic media rather than basal media. 38 However, at day 21, the levels of ALP activity on BBG-Mg, -Sr, and 45S5 bioglass cultured under osteogenic media are significantly higher than the cultures without glass addition (e.g., at day 21, under osteogenic media, the addition of 20 mg/mL of BBG-Sr (c) induced a highly significant (p < 0.001) enhanced ALP activity in relation to the cells cultured in the absence of glass particles).

ALP activity of BM-MSCs (cultured either in basal or osteogenic medium) in the presence of different concentrations (0, 20, and 50 mg/mL) of BBG-Mg
BBG-Mg, -Sr, and 45S5 bioglass were not capable of inducing the ALP protein expression alone; however, they were capable of increasing the ALP expression during the differentiation process of BM-MSCs into osteoblasts under osteogenic media (for 21 days of cell culture). It is relevant to point out that previous studies quantified the chemical species released from BBGs in solution, demonstrating a higher concentration of Mg2+ and Sr2+ ions (twofold) when compared with Ca2+ (from BBG-Mg, BBGs-Sr, and BBG-Ca, respectively).29,30
The presence of higher concentration of specific ions (i.e., Mg2+ and Sr2+) might facilitate cell differentiation. It is also relevant to highlight the fact that along with the increase of ALP activity for the case of BBG-Mg and -Sr glasses, an increase of cell proliferation was observed. In contrast, the viability and proliferation data presented a reduction of viable and live cells for BM-MSCs cultured in the presence of BBG-Ca and 45S5 bioglass. Remarkably, BBG-Sr promoted the increase of ALP activity for both concentrations of glass particles, that is, 20 and 50 mg/mL.
Matrix mineralization
Similar results were found at day 21 after Alizarin red staining for mineral deposits (Fig. 6). In agreement with ALP activity data, cells cultured under osteogenic media yielded more bone-like nodules (intense red spots dispersed in the cell culture), resulting from ECM mineralization. 35 Of importance, and corroborating with ALP activity data, mineral deposits were more evident with BBG-Mg, -Sr, and 45S5 bioglass (Fig. 6a, c, and d) and an increase of red nodules with the increase of concentrations can be observed in the case of cells cultured in the presence of BBG-Sr (Fig. 6c, osteo 20 and 50 mg/mL).
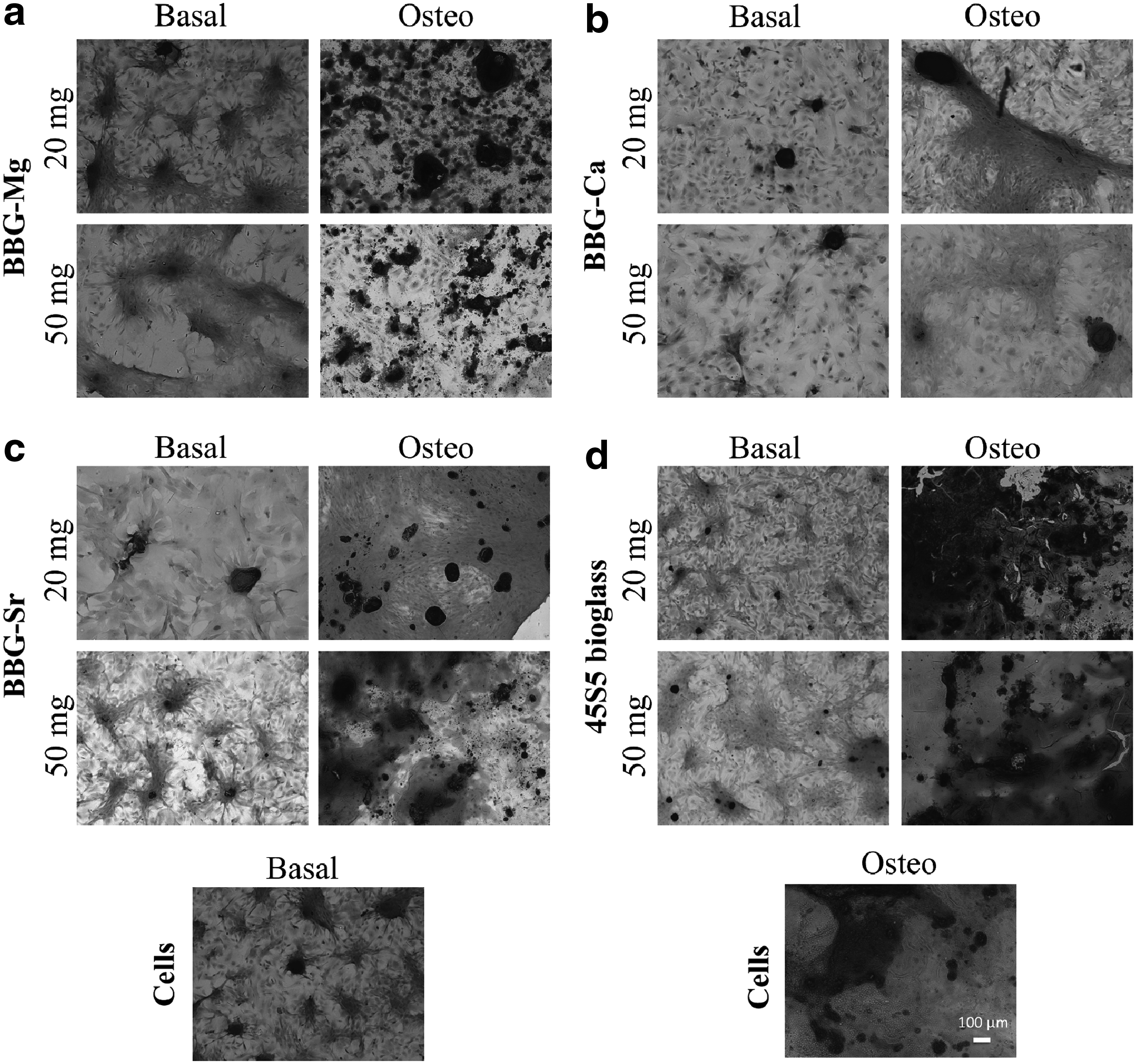
Alizarin red staining of BM-MSCs cultured during 21 days, either in basal or osteogenic medium in the presence of different concentrations (20 and 50 mg/mL) of BBG-Mg
Higher concentrations of BBG-Sr and 45S5 bioglass show the presence of red nodules even in cells cultured in basal medium. However, ALP activity data suggest that BBG-Sr and 45S5 bioglass (Fig. 6c, d basal for a concentration of 50 mg/mL) are capable to induce ECM mineralization by themselves. Therefore, the combination of ALP activity and mineralization results suggests the use of BBG-Sr to promote osteogenesis. 39
In Figure 7, we presented SEM micrographs of BM-MSCs cultured for 21 days in the presence of BBGs and 45S5 bioglass. In the images, it is possible to observe the deposition of minerals over the dense layer of cells when they were cultured in the presence of BBG-Mg, -Sr, and 45S5 bioglass (Fig. 7a, c, and d). The presence of these glass compositions in the culture medium promoted a mineralization typical to occur while BM-MSCs differentiate into osteoblasts. 38 This mineralization is in agreement with the Alizarin red data where the mineral deposits were more evident in the cells cultured in the presence of BBG-Mg, -Sr, and 45S5 bioglass (Fig. 6a, c, and d).
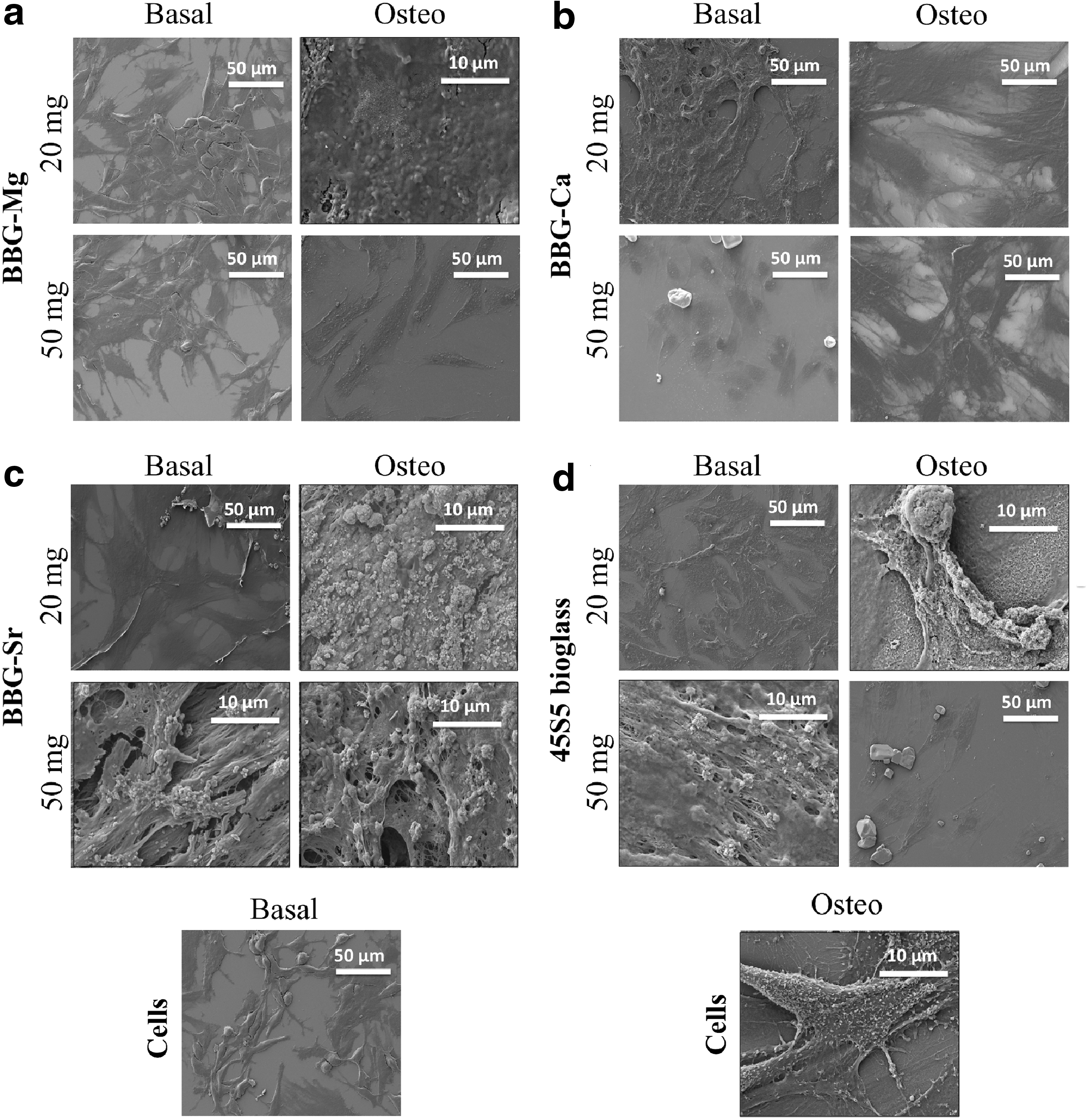
SEM micrographs of BM-MSCs in the presence of different concentrations (20 and 50 mg/mL) of BBG-Mg
Under these conditions, in the SEM/EDS images, calcium phosphate deposits were observed over the dense cellular layer. Once more, higher concentrations of BBG-Sr and 45S5 bioglass under basal culture conditions promoted the deposition of higher amounts of calcium phosphate structures, suggesting that BBG-Sr and 45S5 bioglass (Fig. 7c, d basal for a concentration of 50 mg/mL) are capable of inducing ECM mineralization by themselves, which could be beneficial for bone regeneration. 39
Protein expression (OP and OC)
Complementary to the reported biological data, the differentiation level of BM-MSCs cultured in the presence (20 and 50 mg/mL) and absence of BBG-Mg, -Ca, -Sr, and 45S5 bioglass (either in basal or osteogenic medium) was assessed by quantification of the expression level of two major bone-specific proteins, that is, OP and OC. The relative expression of these proteins was normalized in relation to the number of cells, that is, amount of dsDNA. It is well known that osteoblasts are differentiated cells that mineralize the bone matrix.
OP is a phosphoprotein synthesized by bone-forming cells, which present calcium-binding domains, and is responsible for cell attachment, proliferation, and ECM mineralization. 40 In the case of OC, it is a bone-specific glycoprotein capable of binding to calcium, which promotes ECM calcification. 40 Not surprisingly, the OP and OC quantification data (Figs. 8 and 9) showed a significantly higher protein expression when BM-MSCs were cultured under osteogenic media rather than basal media. 41

OP protein content of BM-MSCs cultured either with basal or osteogenic medium in the presence of different concentrations (20 and 50 mg/mL) of BBG-Mg
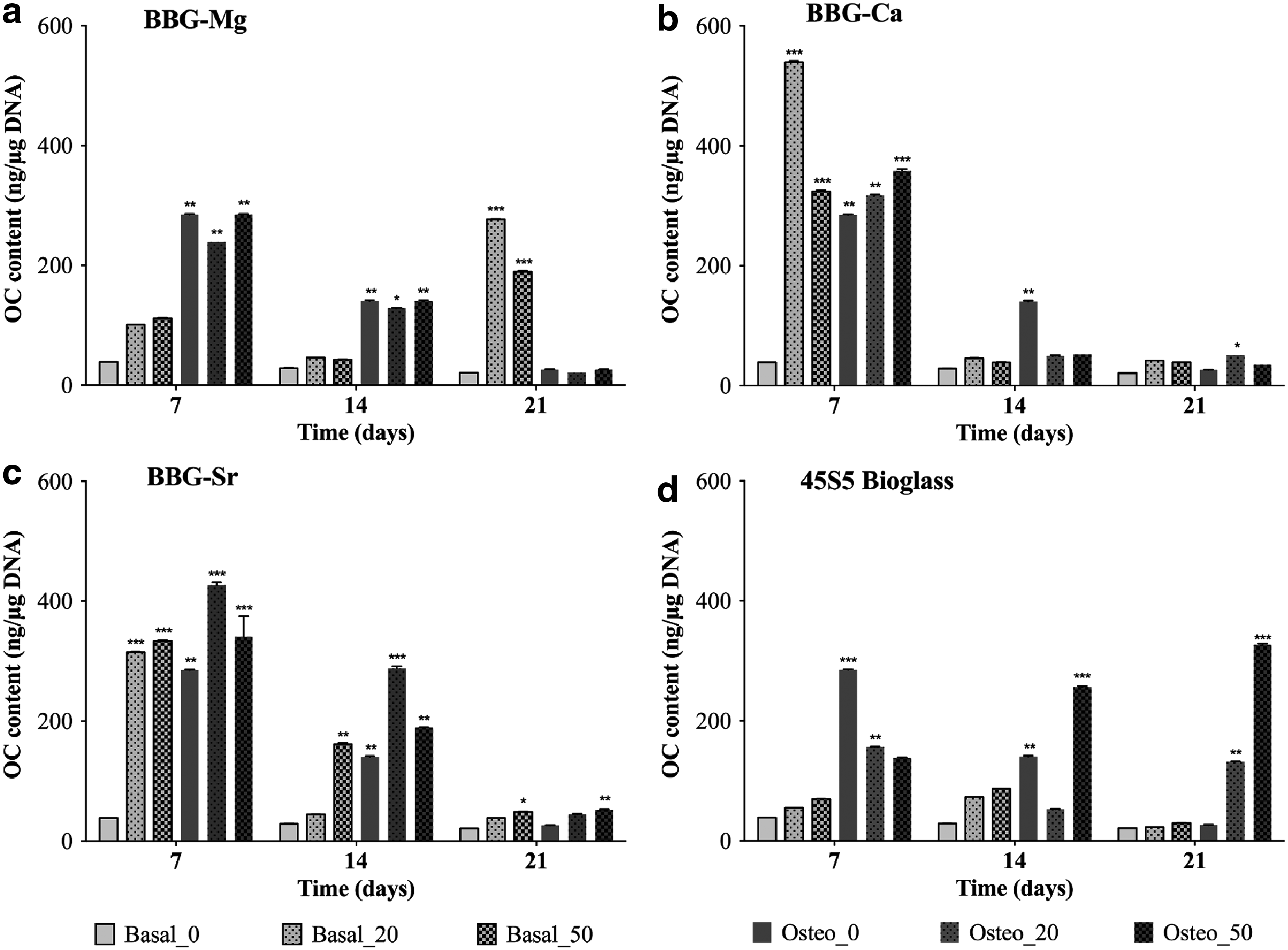
OC protein content of BM-MSCs cultured either with basal or osteogenic medium in the presence of different concentrations (20 and 50 mg/mL) of BBG-Mg
In the case of OP, as expected, a delay in the protein synthesis is observed (Fig. 8). At day 7, there was no significant difference of OP expression in relation to the control (absence of glass particles and cell culture in basal medium). However, at day 14, there is a high expression peak by BM-MSCs cultured in osteogenic medium (in the presence of BBG-Mg, -Sr, and 45S5 bioglass), which determines the decay of the matrix deposition phase and the beginning of the mineralization phase. Moreover, BBG-Sr and 45S5 bioglass continue to induce a significant overexpression of OP over time (e.g., at day 21), supporting the mineralization demonstrated by ALP and Alizarin red analysis (Figs. 5 and 6, respectively).
In the OC case, there was a high protein expression up to day 14, indicating bone ECM maturation (Fig. 9). 42 At day 7, there is a significant difference in OC expression in relation to the control (cell culture in basal medium and in the absence of glass particles). After day 7, there was a reduction of OC expression, consistent with matrix mineralization. Noteworthy is the observation that BBG-Sr under basal medium induced the BM-MSCs to exhibit a peak of OC expression at day 14. These data suggested that the BBG-Sr glass particles (at a concentration of 50 mg/mL) induced the OC protein expression, which is in agreement with the ALP and Alizarin red data. In addition, BBG-Sr and 45S5 bioglass prolonged the OC overexpression over the 21 days of culture.
In addition, the 45S5 bioglass promoted a high deposition of OC at day 21 (Fig. 4h) compared with high BM-MSC density in the case of BBG-Sr (Fig. 4f). However, it is important to consider that in the case of the cultures in the presence of 45S5 bioglass, a very low BM-MSC cell density was observed, which might be related with the cytotoxicity of 45S5 bioglass. 43 Therefore, and overall, our data suggest that the BBG-Sr glass particles are able to induce the BM-MSCs to express higher levels of OP and OC, while maintaining the BM-MSC cell density.
Hence, combining the obtained biological data, that is, viability, proliferation, mineralization, and protein expression analysis (ALP, OC, and OP), we were able to identify BBG-Mg and -Sr (at concentrations of 20–50 mg/mL) as relevant promoters of osteogenic differentiation of BM-MSCs. Several authors reported that Mg2+ ions significantly enhanced osteoblast adhesion by the altering cell–matrix interactions, which modulate the function of integrins related with cell differentiation.19,44 Our results suggest that BBG-Mg also promoted BM-MSC differentiation, which might be related with the presence of Mg2+ in the culture medium.
Remarkably, BBG-Sr (at a concentration of 50 mg/mL and 21 days of culture) presented the capacity to induce osteogenic response in BM-MSCs in the absence of osteogenic medium. Other authors reported Sr-containing glasses to stimulate osteoblast metabolic activity, inhibiting osteoclast differentiation as well as promoting the increment of ALP activity. 24 For instance, Hurtel-Lemaire et al. 45 have shown that Sr induces osteoclast apoptosis at concentrations higher than 9 mM. This is in accordance with our previous studies 29 that demonstrated a concentration of Sr in the culture media of about 20 mM after 3 days of culture. The ALP activity results showed the shift of BM-MSCs to a more differentiated state, while the Alizarin red analysis demonstrated that the cells in the presence of BBG-Mg and BBG-Sr glass particles present intense and dispersed red spots in the cell culture, corresponding to the mineralization promoted by the BM-MSCs.
Finally, Santocildes et al. 25 demonstrated that the dissolution of Sr-containing glasses stimulated the upregulation of genes associated with the process of osteogenic differentiation, such as Bglap (OC) and Spp1 (OP). In agreement with these data, we also observed that BBGs-Mg and BBG-Sr might also influence the ECM maturation and mineralization, through the promotion of the OP and OC protein overexpression, which suggests that these glass compositions may be effective in inducing and sustaining the osteoblastic phenotype. 46
Conclusion
BBGs with different substituted divalent cations (Ca2+, Sr2+, or Mg2+) were successfully synthesized by melt quenching. In vitro studies demonstrated that the studied BBGs exhibit the capability to improve the osteogenic differentiation of BM-MSCs with no deleterious effects over cell viability and proliferation. Specially, BBG-Mg and BBG-Sr (at 20 and 50 mg/mL) provided favorable conditions for BM-MSCs to differentiate to osteoblast-like cells and induce the formation of a high amount of mineralized nodules. The phenotypic expression of two major bone-specific proteins, namely OP and OC, confirmed the osteogenic potential of the BBGs.
The findings that the BBGs are able to promote in vitro cell differentiation into an osteogenic lineage support their potential application in regenerative medicine. Based on these promising results, we propose the incorporation of these BBGs into biomaterials for bone regeneration. The proposed BBGs are also relevant candidates for further in vivo evaluation.
Footnotes
Acknowledgments
The authors gratefully acknowledge financial support from Portuguese Foundation for Science and Technology (PhD grant BD/73162/2010), the European Commission Seventh Framework Programme (FP7/2007–2013) under Grant No. REGPOT-CT2012-31633-POLARIS, and Horizon 2020 Programme under Grant No. WIDESPREAD-2014-2-668983-FORECAST. This work was also supported by the European Research Council grant agreement ERC-2012-ADG-20120216-321266 for the project ComplexiTE and is associated with the UK EPSRC Centre for Innovative Manufacturing of Medical Devices-MeDe Innovation (EPSRC grant EP/K029592/1) where Fernandes was a visiting researcher.
Disclosure Statement
No competing financial interests exist.