
Abstract
Mounting evidence indicates that neutrophils, first responders to an implanted biomaterial, prime the microenvironment for recruited immune cells by secreting factors and releasing neutrophil extracellular traps (NETs) through NETosis. In this study, we investigated the role of electrospun template architecture and composition in regulating NETosis. Electrospun polydioxanone (PDO), collagen type I (COL), and blended PDO-COL templates (PC) were fabricated with small-diameter (0.25–0.35 μm) and large-diameter (1.0–2.00 μm) fibers. Neutrophil–template interactions were evaluated in vitro for 3 and 24 h with human neutrophils, and the PDO templates were studied in vivo (rat subcutaneous model) for 1 and 7 days. Template-bound NETs were quantified by fluorescent microscopy and an On-cell Western assay. The in vitro results indicate that larger fiber diameters reduced NETosis on PDO templates, whereas the incorporation of COL attenuated NETosis independent of fiber diameter. The in vivo results similarly revealed a lower degree of NETs on large-diameter PDO templates at 1 day, resulting in marginal tissue integration of the templates at 7 days. In contrast, the small-diameter PDO templates, which were coated in a large amount of NETs at 24 h in vivo, were surrounded by capsule-like tissue at 7 days. These preliminary in vivo results validate the in vitro model and signify NETosis as a potentially significant physiological response and a critical preconditioning event for the innate immune response to templates. In conclusion, these results demonstrate the importance of characterizing the neutrophil's acute confrontation with biomaterials to engineer templates capable of promoting in situ regeneration.
Introduction
B
Neutrophils, the sentinels of the innate immune system, are highly mobile phagocytes that are recruited rapidly and in large quantities by chemotaxis to a site of acute inflammation.4,5 Historically, minimal attention was given to neutrophils and their role in tissue regeneration6–8 ; however, new evidence suggests that the neutrophil's activity and interaction with biomaterials deserve critical attention. Specifically, by adapting to microenvironmental cues and synthesizing and secreting an array of factors (i.e., cytokines, chemokines, and matrix metalloproteinases), neutrophils exhibit multifaceted physiological effects, including angiogenesis and regeneration.4,5,9–11 Furthermore, research in the field of tumor biology has revealed that tumor-associated neutrophils (TANs) have considerable plasticity of phenotype, displaying an N1 antitumor phenotype or an N2 protumor phenotype, which is regulated by the tumor microenvironment and its constituent factors.3,12 These contrasting TAN phenotypes are synonymous with the macrophage M1 and M2 paradigm, varying from the promotion of inflammation to the promotion of angiogenesis and tissue growth. 12
In addition to their ability to secrete potent factors (i.e., tissue inhibitor of metalloproteinase-free matrix metalloproteinase-9 13 ), neutrophils activated by microbial or chemical factors 6 are able to expel their DNA along with granular contents (i.e., antimicrobial proteins and proteases) to form neutrophil extracellular traps (NETs). 14 This process, deemed NETosis, was only recently discovered in 2004 and is not well understood,14,15 especially its implications in in situ template preconditioning and guided tissue regeneration.
On the decondensing of chromatin, aided by histone citrullination, 16 and the disintegration of granules, the neutrophil cell membrane ruptures to extrude a cloud-like, fibrillary network of DNA, decorated with proteins and histones, into the microenvironment. 17 Neutrophils dying through the NETosis mechanism do not display “eat me” signals like apoptotic or necrotic cells do before cell membrane disruption. This prevents their pre-emptive clearing by macrophages and facilitates complete NET extrusion. 18 Furthermore, it has been proposed that NETosis is a response to noxious stimuli, such as biomaterials that are too large to be phagocytosed,19,20 which is supported by findings that large particles such as cholesterol crystals induce NETosis.21,22 Taken together, this suggests that NETs may be extruded on the surface of implanted tissue regeneration templates and precondition the ensuing innate immune response.
This article evaluates the hypothesis that NETs are generated in response to electrospun tissue templates and that the degree of template-bound NETosis is a function of template architecture (i.e., fiber diameter) and composition. Polydioxanone (PDO), collagen type I (COL), and a blend of the two polymers (PDO:COL [PC] 90:10 v/v) were electrospun with varying yet controlled architecture (i.e., small-diameter [SD]) and large-diameter [LD] fibers and pores). NETosis was examined in vitro with freshly isolated, human peripheral blood neutrophils and in vivo with a subcutaneous implant rat model and analyzed based on template architecture, composition, and more specifically, architecture within a composition. Furthermore, NETosis was examined in vitro at multiple time points to elucidate its potential temporal regulation by template architecture and composition. Ultimately, understanding this aspect will facilitate design of electrospun templates and other biomaterials that harness the body's innate immune system as a bioreactor component to promote, guide, and enhance in situ tissue integration and regeneration.
Materials and Methods
Fabrication and characterization of electrospun templates
For this study, COL was isolated from rat tails following the protocol previously described by Rajan et al. 23 Briefly, the collagen type I obtained from rat tail tendons was dissolved in acetic acid, frozen at −80°C, and lyophilized. PDO (Sigma Aldrich Co.) and COL were dissolved overnight in 1,1,3,3,3 hexafluoro-2-propanol (HFP; Oakwood Products, Inc.) at ratios of 100:0, 90:10, and 0:100 (v/v) at low (60 mg/mL) and high (PDO 140 mg/mL and COL 110 mg/mL) concentrations to fabricate SD and LD regeneration templates, respectively.
Solutions of PDO, COL, and PC were placed in a syringe on a syringe pump (Model No. 78-01001; Fisher Scientific) with an 18-gauge blunt needle tip (2.5 cm length) attached to the positive voltage lead of a power supply (Spellman CZE1000R; Spellman High Voltage Electronics Corp.). Solutions were dispensed at a rate of 2–6 mL/h (2 mL/h for COL LD, 3 mL/h for COL SD, 4 mL/h for PC SD and PDO SD, 5 mL/h for PC LD, and 6 mL/h for PDO LD) with a voltage applied to the solution of +22 to +28 kV (+22 kV for PDO SD and PDO LD, +25 kV for PC SD, PC LD, and COL LD, and +28 kV for COL SD). The airgap distance was 12.7 cm for all solutions. Fibers were collected on a grounded stainless steel rectangular mandrel (200 × 750 × 5 mm), rotating at 1250 rpm and translating 6.5 cm/s over 13 cm.
Templates were coated with 5 nm of gold palladium by sputter coating in an argon gas field before imaging via scanning electron microscopy (SEM, FEI NN650 FEG, Nova NanoSEM™ with field emission gun) at +20 kV with a working distance of 5 mm. Templates were characterized regarding fiber and pore diameters by analyzing the SEM images with FibraQuant™ 1.3 software (nanoTemplate Technologies, LLC). The average fiber diameter and pore diameter with corresponding standard deviations were calculated by averaging a minimum of 200 semiautomated random measurements per image and 60 random manual measurements per image, respectively.
In vitro study
Human neutrophil isolation
Neutrophils were isolated from blood obtained from healthy donors from Key Biological Services in accordance with protocols approved by the University of Tennessee Institutional Review Board. During disinfection and plating of the templates, fresh human peripheral blood neutrophils were isolated following a modified version of published protocols.24,25 Briefly, neutrophils were purified at room temperature from heparinized whole blood of healthy donors and collected from supernatant after dextran sedimentation and from the pellet of an Isolymph density gradient (Gallard Schlesinger Chemical) under endotoxin-free conditions. The contaminating erythrocytes were lysed in ice-cold hypotonic 0.2% sodium chloride solution for 30 s, and the solution was rendered physiological saline by addition of hypertonic 1.6% sodium chloride solution, resulting in neutrophil viability >98% as assessed by trypan blue dye exclusion. The neutrophils were washed once in 1× Hank's buffered salt solution (HBSS) and resuspended at a density of 4 × 106 cells/mL in HBSS with 0.2% heat-inactivated human serum, sodium-bicarbonate, and 10 mM HEPES.
Neutrophil seeding
Before disinfection, pure COL templates were crosslinked in 25 mM 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide hydrochloride (EDC; Thermo Scientific Pierce™) in ethanol for 18 h at room temperature. 26 Six 10-mm-diameter discs (thickness 500 ± 100 μm) of each template type were punched and disinfected with a 30-min ethanol soak, followed by three 10-min washes with HBSS. The disinfected templates were then placed in a 48-well culture plate and kept hydrated with HBSS before seeding of neutrophils. Immediately following neutrophil isolation, templates were seeded with 150 μL of the cell suspension (600,000 cells/well). After waiting for 10 min to allow the neutrophils to settle by gravity, an additional 150 μL of HBSS was carefully added to the wells. All templates were cultured under standard culture conditions (37°C and 5% CO2) for 3 and 24 h, after which the plates were placed on ice to inhibit further stimulation of the neutrophils. Supernatant was removed from the cellularized templates, and the cells were fixed in their wells on the templates with 10% buffered formalin. Fixed templates were stored in formalin at 4°C until analysis.
Fluorescent detection of NETs
The surface of fixed, cellularized templates (n = 3, biological and technical replicates represented by three different donors seeded on templates from three different fabrication runs) was imaged with fluorescent microscopy for quantitative analysis of the degree of NETosis in response to the templates. Templates were removed from formalin and stained with 100 nM SYTOX® green (SG) extracellular DNA stain (Cat. No. S7020; Life Technologies) in deionized (DI) water for 10 min at room temperature followed by 4′,6-diamidino-2-phenylindole (DAPI) fixed cell nuclei stain (Cat. No. R37606; Life Technologies) at stock concentration for 5 min at room temperature. Three DI water washes were performed between each step, and stained templates were covered with a glass coverslip.
Two images were taken randomly from different regions of each template with an Olympus microscope (Model BX43F) equipped with an Olympus DP73 high-performance digital color camera and fluorescent light source (DAPI and SG excited at 358 and 504 nm, respectively) at 20× magnification. Using cellSens® Standard 1.9 Digital Imaging software (Olympus Corp.), exposure times of each channel were kept at 200 ms to avoid saturated pixels. DAPI and SG images of the same region were overlaid, and the background was subtracted by defining a region of interest containing no cell nuclei or extruded NETs.
To determine the relative degree of NETosis, the blue to green (B:G) ratio, representing the area of an image occupied by intact cell nuclei compared to the area occupied by extruded NETs, respectively, was quantified. Using MATLAB® Release 2012a, the number of blue (DAPI), green (SG), and cyan (double staining of DAPI and SG) pixels in an image was quantified with a minimum threshold of 110 for all three colors. The number of cyan pixels was subtracted from the number of green pixels, and the modified number of green pixels was used to formulate a B:G ratio. An average B:G ratio and corresponding standard deviation were calculated for each template type.
Infrared On-cell Western blot
An On-cell Western assay was performed to quantify template-bound citrullinated histone H3 (CitH3). Fixed, cellularized templates were removed from formalin and rinsed with 1× phosphate-buffered saline (PBS; HyClone™; GE Healthcare). Free aldehyde groups were quenched with three washes of 25 mM glycine in PBS for 5 min at room temperature and blocked with Odyssey® Blocking Buffer (Part no. 927-40000; LI-COR®) for 1.5 h at room temperature.
A standard curve was generated to quantify template-bound CitH3 using human histone H3 (citrulline R2 + R8 + R17) peptide (Product code ab32876; Abcam) spotted on an Immobilon®-FL PVDF membrane (Cat. no. IPFL00010; EMD Millipore Corporation) at 23.5, 11.7, 5.96, 2.93, 1.46, 0.73, 0.37, 0.18, 0.09, and 0.00 ng per spot in DI water. After allowing it to dry overnight, the spotted membrane was blocked with Odyssey Blocking Buffer for 1 h at room temperature.
The templates (n = 3, biological and technical replicates) and membrane were incubated with the polyclonal anti-histone H3 (citrulline R2 + R8 + R17) antibody (Product code ab5103; Abcam) at a 1:200 dilution in Odyssey Blocking Buffer overnight at 4°C. All templates and membrane were washed five times with 0.1% Tween-20 in PBS for 5 min at room temperature with gentle shaking and subsequently incubated with IRDye 800CW donkey anti-rabbit (Part no. 926-32213; LI-COR) at a 1:20,000 dilution in Odyssey Blocking Buffer for 1 h at room temperature, protected from light. Noncellularized templates, serving as negative controls, were incubated with secondary antibody only. After four washes with 0.1% Tween-20 in PBS and a fifth wash with PBS, the templates and membrane were scanned on the 800 nm channel of the Odyssey CLx Infrared Imaging System (LI-COR) with automatic intensity adjustment to obtain full-thickness template fluorescence and analyzed using Image Studio Version 5.x (LI-COR).
For the standard curve, fluorescent intensities were measured using circular markers encompassing the area of a spot. The intensities and corresponding mass of CitH3 were plotted and fit with a line of best fit. Then, the fluorescent intensities of the templates were measured using circular markers placed on the center of the template. The intensities were extrapolated to give total fluorescence for a 10 mm punch and converted to the mass of template-bound CitH3 using the standard curve.
In vivo study
Implantation of templates in a rat subcutaneous pocket
Ten-millimeter-diameter discs of SD and LD PDO templates (to elicit the most drastic responses as determined by the in vitro model) were implanted on the dorsa of four Sprague-Dawley rats (250–300 g, male and female) following the protocol approved by the University of Memphis Institutional Animal Care and Use Committee. Templates were sterilized with peracetic acid (Product no. 77240; Sigma-Aldrich®), 27 and all materials were handled aseptically. Templates were hydrated for 10 min with sterile 1× PBS before implantation to minimize air within the template.
The rats were anesthetized by 5% isoflurane in an induction chamber, and anesthesia was maintained with 1–2% isoflurane delivered through a nose cone. Antisepsis was provided with iodine, and after creating incisions, four isolated subcutaneous pockets were tunneled on the dorsa of each animal by blunt dissection. One template lying flat was implanted per pocket to minimize void space around the template. Four replicates per template type (n = 4) were implanted for the 24-h time point, and two replicates per template type (n = 2) were implanted for the 7-day time point. The skin was closed with a single suture. The animals were euthanized 24 h and 7 days after implantation. After euthanasia, the subcutaneous pocket was opened, templates were explanted (7-day templates excised with surrounding tissue), and subsequently fixed with 10% buffered formalin. Templates were stored in 10% buffered formalin at 4°C until analysis.
Template staining and evaluation
Fixed, excised templates were processed and stained to evaluate the in vivo response to the different template architectures. For cryosectioning, templates were soaked in 30% sucrose solution overnight, embedded in Tissue-plus® O.C.T. Compound (Fisher HealthCare), frozen at −20°C, cryosectioned at a thickness of 15 μm, and mounted on glass slides. Replicates from 24 h were stained with DAPI and SG following the same methods defined for the in vitro samples, as described in the Fluorescent detection of NETs section, and with Shandon Kwik-Diff™ stain kit (Cat. no. 9990701; Thermo Scientific™) following the manufacturer's protocol. In addition, 7-day samples were processed and stained with hematoxylin and eosin (H&E) following standard protocol. Sections were qualitatively evaluated for cell migration through the template and the overall inflammatory response.
Infrared On-cell Western blot
Template-bound CitH3 was quantified on 24-h templates (n = 4) following the On-cell Western assay protocol used for quantifying template-bound CitH3 on in vitro templates, as described in the Infrared On-cell Western blot section.
Statistical analyses
Statistical analyses were performed using Microsoft® Excel Real Statistics to determine significant differences at an a priori level of p < 0.05. For template characterization and quantification of template-bound CitH3, analysis of the data was conducted using a Kruskal–Wallis one-way analysis of variance (ANOVA) test and Tukey–Kramer HSD multiple comparison procedure. For the fluorescent detection of NETs, analysis of the data utilized a Welch's ANOVA test and the Games–Howell multiple comparison procedure.
Results
Template characterization
SEM image analysis confirmed that low and high polymer concentrations resulted in small and large fiber and pore diameters, respectively (Fig. 1). All LD templates resulted in fiber diameters that were statistically larger than their SD counterparts (1.0–2.0 μm vs. 0.25–0.35 μm, p < 0.05). Similarly, all LD templates resulted in pore diameters that were statistically larger than their SD counterparts (6.0–8.0 μm vs. 1.0–1.25 μm, p < 0.05).
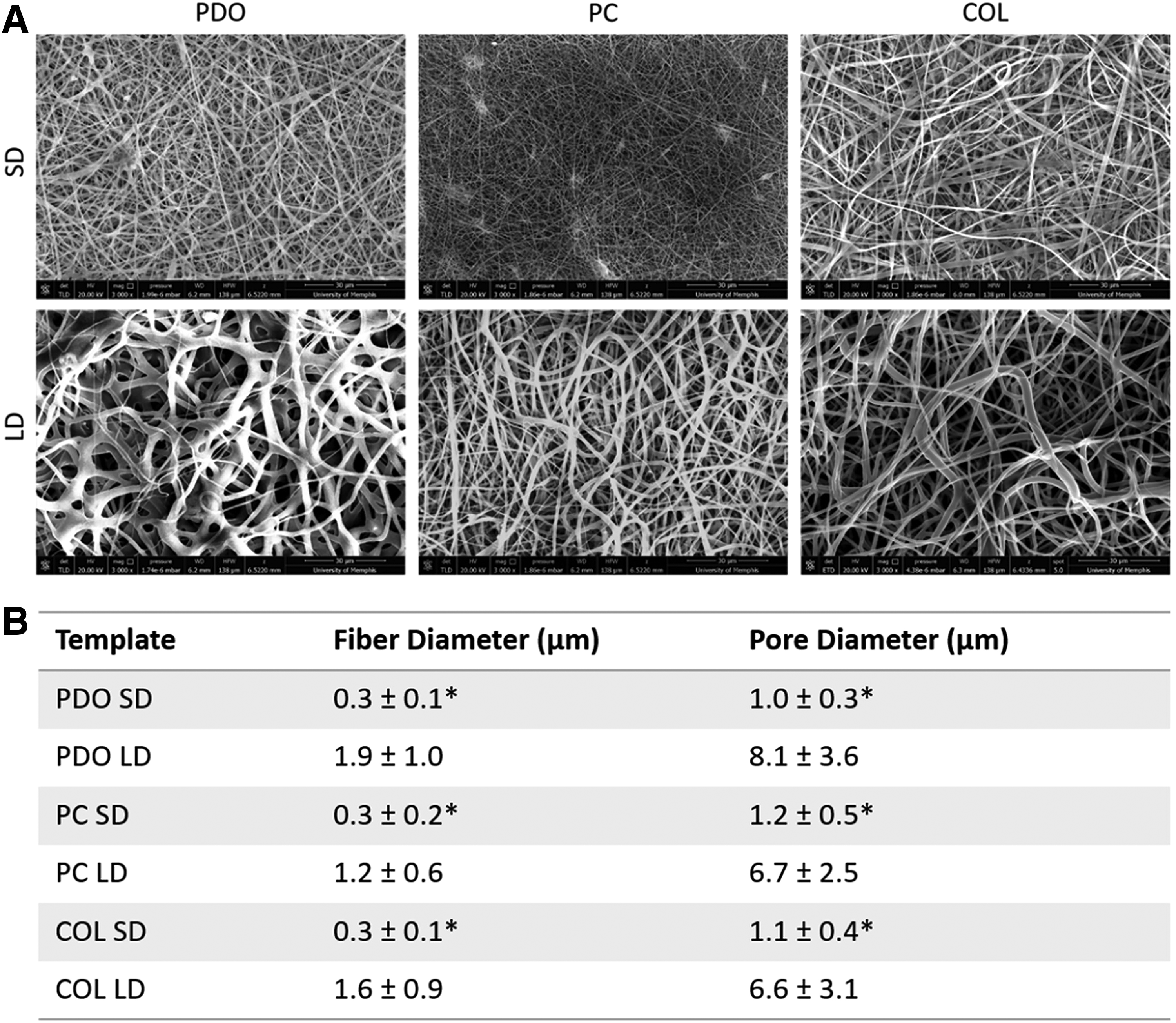
Processing solution concentration alters fiber diameter and pore diameter.
In vitro results
Fluorescent detection of NETs
Fluorescent microscopy revealed varying degrees of NETosis in response to the different template architectures and compositions (Fig. 2), which were evaluated with B:G ratios quantified by an image analysis algorithm written in MATLAB® (Fig. 3). Blue and green signify SG and DAPI staining. During NETosis, the plasma membrane maintains its integrity, while the nuclear and granular membranes disintegrate. 28 Because SG is unable to penetrate an intact plasma membrane, 29 double staining of DAPI and SG, resulting in cyan, denotes cells that may be undergoing cellular changes, other than NETosis, that are associated with a compromised plasma membrane. 28 Therefore, blue, green, and cyan represent intact viable cells, extruded NETs, and intact cells with compromised cell membranes, respectively. To accurately quantify the degree of NETosis as the B:G ratio and obtain a true measurement of extruded NETs, cyan pixels were subtracted from the green pixels before calculating the B:G ratio. A B:G ratio greater than 1 signifies a larger area of the template occupied by intact cells than by extruded NETs (i.e., a relatively low degree of NETosis) with the inverse for a B:G ratio less than 1 (i.e., a relatively high degree of NETosis).
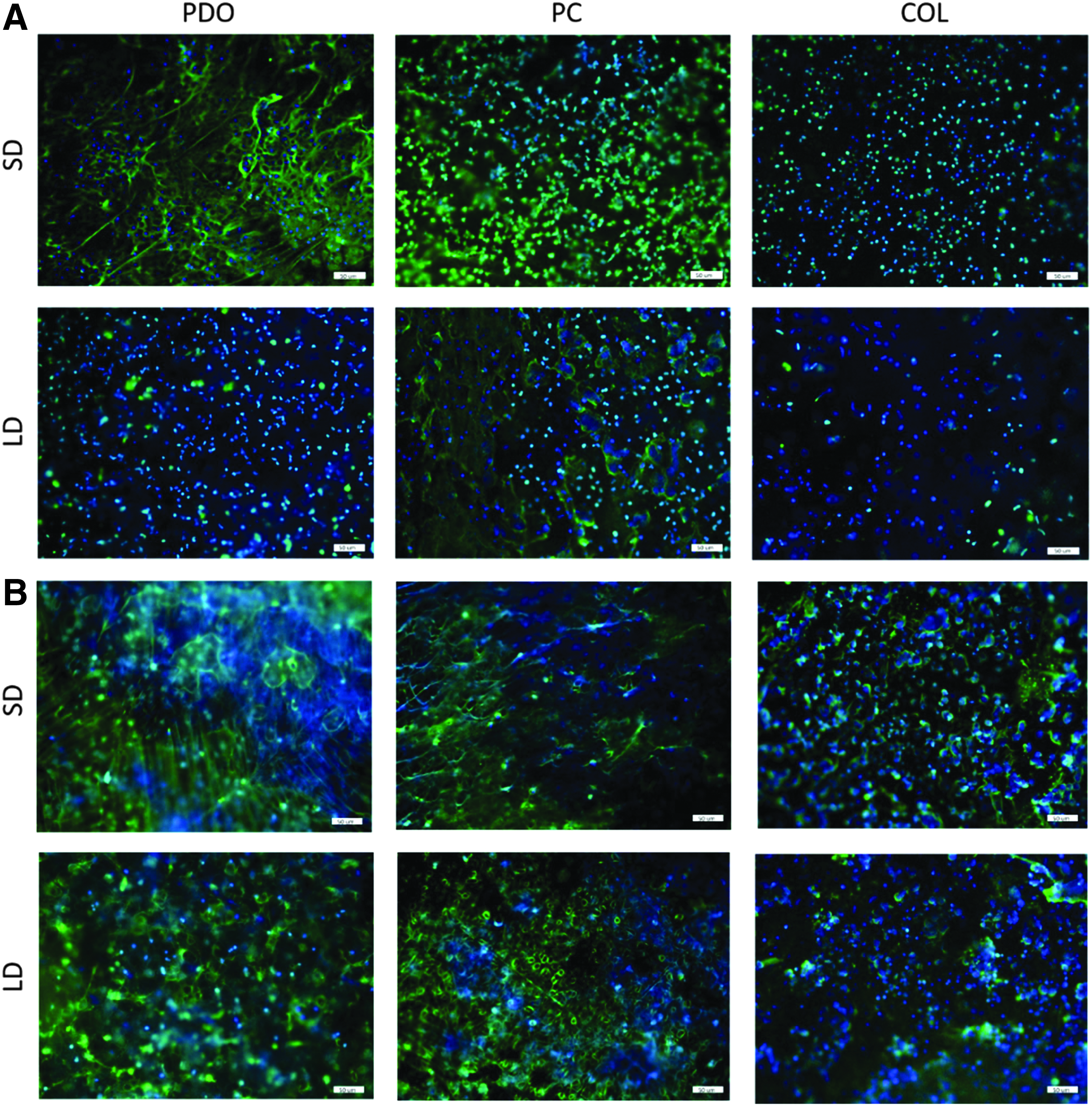
Fluorescent micrographs demonstrating varying degrees of NETosis in response to templates of different architectures and compositions. Images of in vitro cellularized templates at
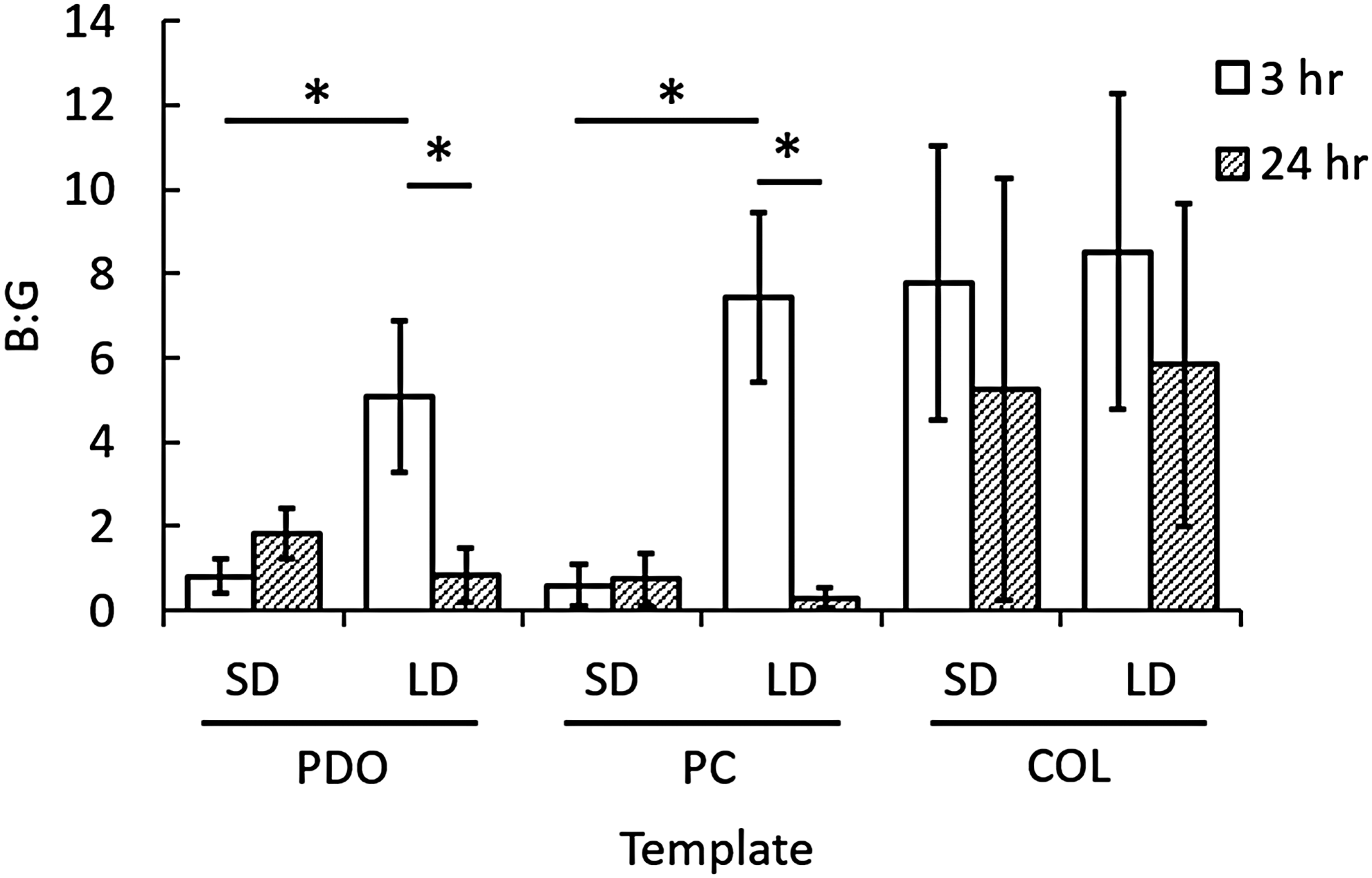
Template architecture and composition modulate NETosis as indicated by B:G ratios. B:G ratios quantifying NETosis at 3 and 24 h. *Indicates a significant difference (p < 0.05). No statistical differences were found between SD and LD template counterparts at 24 h.
Historically, extended in vitro experiments with isolated human neutrophils have not been considered feasible or reliable because of the neutrophil's brief half-life in vitro 30 ; however, fluorescent microscopy revealed that a portion of the template-adhered neutrophils at 24 h were stained with DAPI only (Fig. 2B), indicating the maintenance of an intact cell membrane and thus cell viability. In addition, previous work has relied on artificial stimulation with manganese to enhance neutrophil adhesion. 31 Our studies did not utilize manganese stimulation, and neutrophil adhesion on the templates was evident at both 3 and 24 h (Fig. 2). These results are similar to previous experiments with mast cells, which were also thought to require stimulation for adhesion, but were found to adhere unassisted in vitro. 32
At 3 h, all LD templates, regardless of composition, resulted in B:G ratios greater than 1 (Fig. 3). For PDO and PC, SD counterparts resulted in B:G ratios less than 1. PDO LD and PC LD had significantly higher B:G ratios (6× and 12× increase, respectively) compared to their SD counterparts (p = 0.003, not indicated in Fig. 3). These results suggest that a LD template primarily composed of PDO induces a lower degree of NETosis in vitro compared to SD templates.
In addition, all COL templates at 3 h resulted in B:G ratios greater than 1. The B:G ratio for COL SD was significantly higher (10× increase) than the other SD templates (p = 0.003). Moreover, although not statistically greater (p = 0.34), the B:G ratios for all COL templates were higher than those for PDO LD and PC LD templates, which suggests that incorporation of COL, regardless of architecture, minimizes the degree of NETosis beyond that of PDO template architecture. Together, these data indicate that the degree of NETosis exhibited in response to a tissue regeneration template in the critical, initial hours following implantation does not solely depend on the template architecture free from its composition, but rather depends on a specific template architecture for a given composition.
For all templates that had a B:G ratio greater than 1 at 3 h, the B:G ratio decreased at 24 h with significant differences between time points for PDO LD (p = 0.002) and PC LD (p = 0.03) (Fig. 3). The decreased B:G ratios for templates exhibiting a low degree of NETosis at 3 h suggest that the templates may be temporally modulating NETosis. In other words, these templates may be slowing the onset of NETosis. Inversely, for all templates that had B:G ratios less than 1 at 3 h, the B:G ratio increased at 24 h with no statistical differences. This may be the result of the delicate NETs degrading or detaching from the cellularized templates as time progresses.
Infrared On-cell Western blot
To quantify template-bound CitH3, the PVDF membrane spotted with CitH3 was scanned on the Odyssey system and analyzed. The standard curve revealed a linear relationship (y = 3998.8x, R 2 = 0.9165) between intensity and CitH3. Using the equation obtained from the standard curve, the total amount of template-bound CitH3 was quantified for each template type.
The amount of extracellular CitH3 expressed on the templates appears to be largely dependent on template architecture and composition (Fig. 4). Citrullination of histone H3 is a critical step during NETosis, allowing for the extrusion of an NET, and CitH3 is known to be contained within NETs after extrusion. 33 Because the cells were not permeabilized for the On-cell Western, increased CitH3 expression can be correlated to a high degree of NETosis, while decreased CitH3 expression can be correlated to a low degree of NETosis.
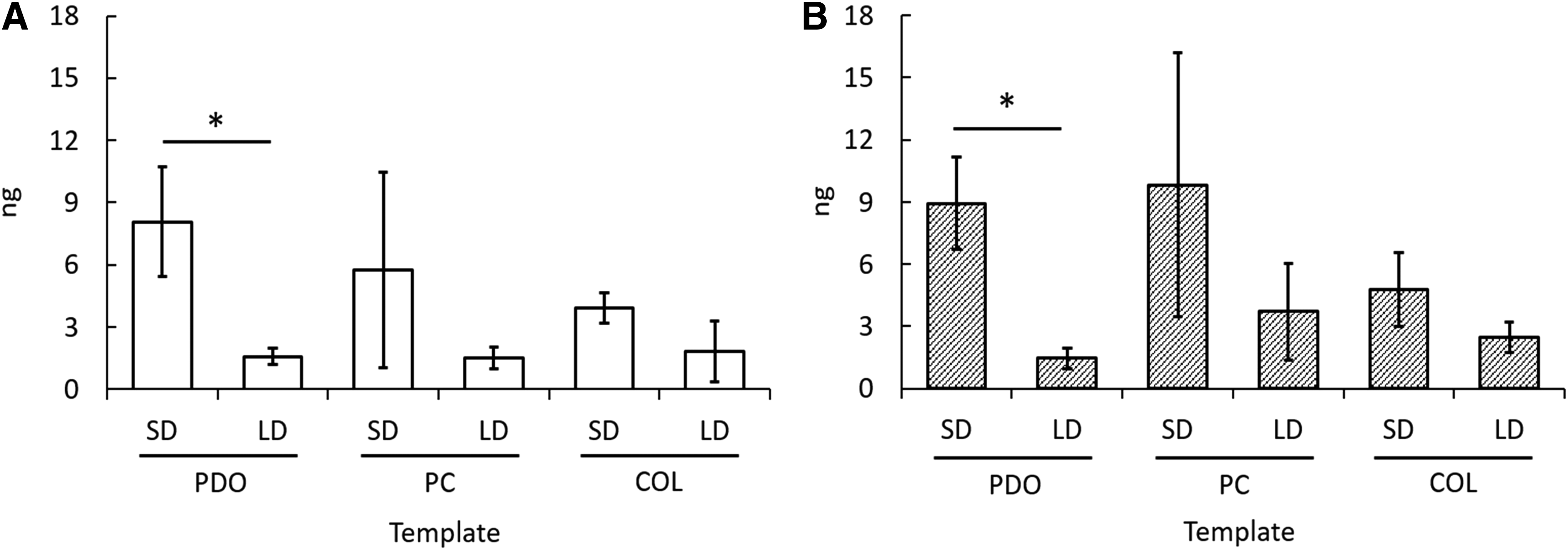
Template architecture and composition modulate NETosis in vitro as indicated by template-bound CitH3. Quantified template-bound CitH3 on templates at
At both time points, templates with smaller fiber diameters and pore diameters resulted in higher amounts of template-bound CitH3 expression compared to templates with larger fiber diameters and pore diameters, with a significant difference between PDO SD and PDO LD at both 3 and 24 h (p < 0.05). Specifically, PDO SD expressed 8.06 ± 2.64 ng of CitH3 at 3 h, which is five times the expression on PDO LD, and 8.94 ± 2.24 ng of CitH3 at 24 h. This stark contrast again demonstrates, via quantification of extracellular, template-bound CitH3, that the degree of NETosis may be regulated by template architecture. Furthermore, the COL SD template expressed 3.9 ± 0.73 ng of CitH3, significantly less than PDO SD, at 3 h (p < 0.05), and 4.0 ± 1.8 ng of CitH3, also less than PDO SD, at 24 h. This decrease in CitH3 expression again suggests that template composition also plays a crucial role in the degree of NETosis.
Interestingly, the amount of template-bound CitH3 did not decrease from 3 to 24 h for PDO SD and PC SD, as anticipated based on the results from fluorescent microscopy (Fig. 4B). This interesting finding suggests that the CitH3 may remain template bound at later time points, even after the NETs have detached or been degraded. Considering the strong positive charge of histones and the largely negative charge of adsorbed proteins on the templates, it is possible that the attraction between CitH3 and the proteins is great enough to conserve the presence of CitH3 for extended periods of time.
In vivo results
Template staining and evaluation
A preliminary in vivo study was executed to evaluate the physiological significance of the in vitro model. Cryosectioning and staining of the PDO SD and PDO LD templates from the rat subcutaneous pocket show that the overall response to the templates in vivo, as orchestrated by the acute confrontation of the neutrophil, varies significantly based on fiber diameter of the template. DAPI and SG staining (Fig. 5A) and Shandon Kwik-Diff staining (Fig. 5B) highlight the extreme differences between templates at 24 h. A dense, cell layer can be seen surrounding the PDO SD templates, whereas the PDO LD templates possess an even distribution of cells throughout their thicknesses. Moreover, at 7 days, the H&E staining (Fig. 5C, D) reveals the formation of a capsule-like tissue (i.e., pink staining) separating the SD template from the surrounding tissue. In contrast, the PDO LD template is partially integrated with the surrounding tissue and has high cellularity around its borders at 7 days. Since the only factor that varied in this initial in vivo study was template architecture, these results emphasize the importance of better understanding the innate immune response to a material and subsequent healing as modulated by material design.
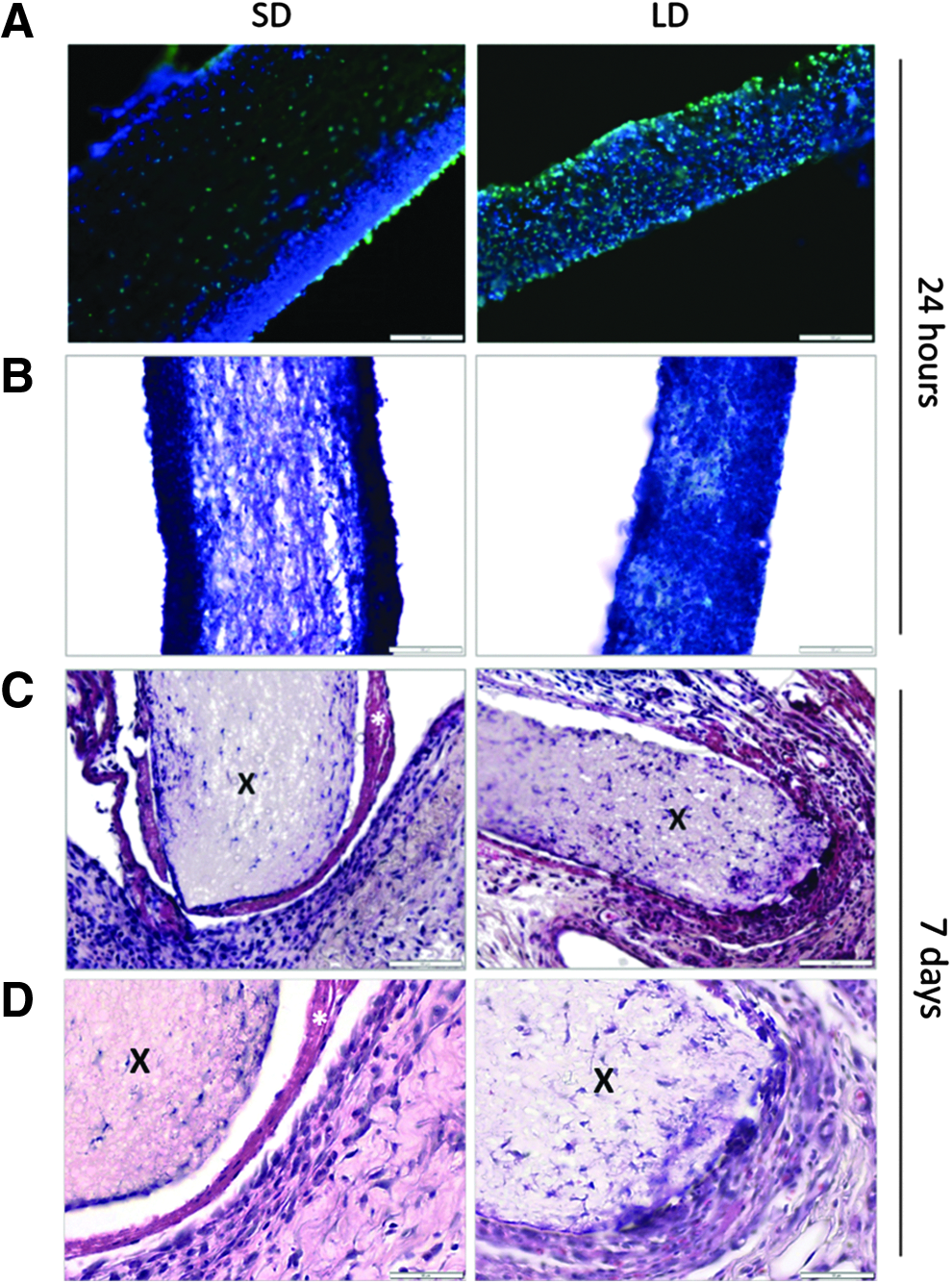
Fiber diameter modulates evoked cell distribution through PDO templates and marginal tissue integration in vivo. The white space between the template and surrounding tissue is an artifact of sectioning. Cross sections of SD and LD templates stained with
Infrared On-cell Western blot
As seen with the in vitro templates, the expression of extracellular, template-bound CitH3 in vivo appears to be dependent on template architecture (Fig. 6). PDO SD expressed 1376 ± 286 ng of CitH3, which is significantly greater than the expression of 422 ± 30 ng of CitH3 on PDO LD (p = 0.02). The three and a half-fold increase in CitH3 expression is indicative of a high degree of NETosis in response to PDO SD templates.
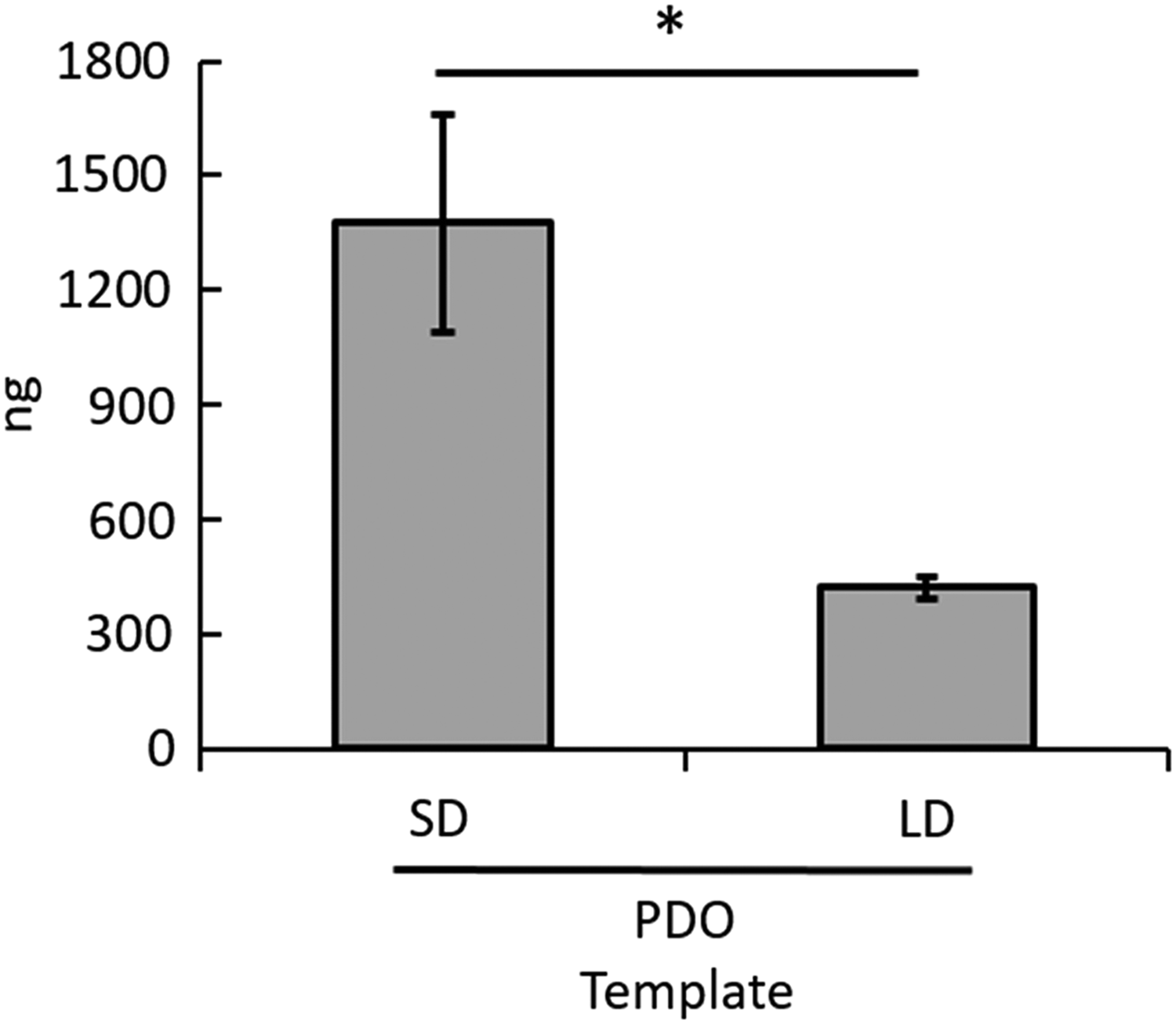
Electrospun PDO template architecture modulates NETosis in vivo as indicated by template-bound CitH3. Quantified template-bound CitH3 on templates at 24 h in vivo. *Signifies a significant difference between templates (p < 0.05).
In addition, the in vivo expression of extracellular template-bound CitH3 is one and two orders of magnitude greater than the in vitro expression on LD and SD PDO, respectively. In the in vitro model, the number of neutrophils was restricted to the seeding density, which inherently limited the expression of CitH3. In the in vivo model, the templates were implanted subcutaneously by blunt dissection, which resulted in an acute inflammatory response at the implant site and allowed for the recruitment of readily available, circulating neutrophils, thus giving rise to the increased expression of CitH3. Most notably, even with the restricted number of neutrophils in vitro and the complex response involving multiple cell types in vivo resulting in CitH3 expression of varying magnitudes, the in vitro and in vivo models follow the same significant trends. In both cases, the expression of CitH3 is four to five times higher on PDO SD than PDO LD.
Discussion
Understanding the role of NETosis in priming electrospun templates may act as a guide for designing templates that promote synergy between neutrophils and other innate immune cells leading to innate immune system-guided in situ regeneration. Our objective was to determine a relationship between template architecture and composition and the degree of NETosis. By electrospinning PDO, COL, and PC at low and high concentrations, resulting in statistically different templates with anticipated fiber morphologies and size distributions,34,35 any variation in the degree of NETosis seen on SD and LD template counterparts can be considered a function of template architecture. Likewise, any variation in the degree of NETosis seen on templates with the same architecture, but varying compositions, can be attributed to the template composition.
Using freshly isolated human peripheral blood neutrophils, we demonstrated that it is possible to study neutrophil interactions with templates in vitro in non-tissue culture plastic microenvironments that are more physiologically relevant and do not rely on artificial stimulation to facilitate neutrophil–template interactions. Notably, we demonstrated that both template architecture and composition affect the degree of NETosis in response to tissue templates, as evidenced by the variations between B:G ratios and extracellular, template-bound CitH3.
In general, we demonstrated that template architecture plays a significant role with more hydrophobic, synthetic polymer templates and that the incorporation of a more hydrophilic, natural polymer attenuates NETosis regardless of architecture. With both fluorescent microscopy and the On-cell Western blot, we showed that the incorporation of COL decreases NETosis, potentially by enhancing the bioactive surface functionality of the templates,35,36 and thus biocompatibility. Furthermore, the incorporation of COL, segregated to the external surface of blended composition electrospun fibers, 37 increases the hydrophilicity of the template, which has previously been shown to improve the biocompatibility of electrospun templates. 38
For primarily PDO templates, an increased fiber diameter (i.e., micron-size) induces a significantly lower degree of NETosis compared to a decreased fiber diameter (i.e., nano-size), which is potentially the result of the associated nonrestrictive pore sizes allowing for uninhibited cell migration into the template. Our data also suggest that there is a potential temporal modulation of the neutrophil's response via template architecture and composition, demonstrated by templates with a low degree of NETosis at 3 h and a slightly increased degree of NETosis at 24 h. Most importantly, the significant and notable trends regarding synthetic PDO templates were magnified in vivo using a rat subcutaneous pocket model, thus validating the methods used for the in vitro model.
Considering the in vitro and in vivo results, a high degree of NETosis appears to correlate to inflammation and encapsulation/rejection of a biomaterial, which is not surprising since excessive NET formation has been linked to a number of pathologies and diseases in recent years.39–43 The PDO LD templates, which were integrated with the surrounding tissue after 7 days in vivo, stimulated a low degree of NETosis in vitro, while the PDO SD template, coated in CitH3 and other potent proteins contained within NETs, led to minimal marginal tissue integration. Thus, a low degree of NETosis and the associated neutrophil phenotype (i.e., N1 or N2) may better prime the microenvironment for tissue integration and regeneration by reducing inflammation and the presence of harmful factors, such as histones and granule proteins. Characterizing the cell population surrounding and infiltrating these templates may also provide valuable information regarding NETosis and the attenuation or promotion of the foreign body response. Most importantly, the preliminary in vivo study suggests that there is a critical link between the neutrophil's acute confrontation and degree of NETosis, its temporal modulation, and the overall response to and integration of an electrospun template.
The mechanisms of NETosis are not yet fully understood because previous research has relied on pharmacological stimuli, such as phorbol 12-myristate 13-acetate, or microbes to trigger NETosis; nonetheless, significant insight has been discovered in recent years. In general agreement, various stimuli such as microbes and their by-products (i.e., lipopolysaccharide), cytokines (i.e., interleukin-8, tissue necrosis factor, and others), and immune complexes can induce NETosis via binding to toll-like receptors (TLRs, i.e., TLR2 and TLR4), Fc receptors, cytokine receptors, complement receptors, 44 and integrins (i.e., β2 integrin). 45 Binding of stimuli to these receptors leads to an intracellular Ca2+ influx from the endoplasmic reticulum stores and opening of membrane channels, resulting in protein kinase C activity, assembly of NADPH oxidase, and rupture of granular and nuclear membranes. Simultaneously, histones are citrullinated to facilitate chromatin decondensation, mixing of cellular contents, and final extrusion of NETs. 46
Clearly, there are many factors to consider and characterize in the complex neutrophil response to other biomaterials, including how the variation of template hydrophilicity and surface-area-to-volume-ratio 34 may be affecting the adsorption of serum proteins and binding of surface receptors (i.e., TLRs, Fc receptors, complement receptors, and integrins44,45), triggering different degrees of NETosis. More direct approaches to modifying NETosis through the neutrophil's acute confrontation may be prevalent in the near future and would likely assume an important role in the subsequent design of biomaterials for optimal integration.
Nonetheless, one looming question that remains unanswered is the effects of the preconditioning event orchestrated by the initially interacting neutrophils. For this study, we identified that PDO templates with a larger fiber and pore diameter reduce the degree of template-bound NETs. Because a PDO template with larger fiber and pore diameters is also known to shift macrophage phenotype toward the regenerative M2 phenotype and encourage angiogenesis, 47 we conclude that a low degree of NETosis may correlate to a neutrophil N2 phenotype, subsequent macrophage M2 phenotype, and ultimately, angiogenesis and regeneration.
Conclusion
Taking this into consideration, it is evident that this preliminary, first-of-its-kind study yielding significant insight into neutrophil NETosis in response to biomaterials only scratches the surface of the highly complex innate immune response that drives inflammation and rejection of a biomaterial or integration and regeneration. Over the last decade, the macrophage has received significant attention as the key cell determining success or failure of tissue regeneration templates. In contrast, this work suggests that we must focus on the acute confrontation between the neutrophil and biomaterial if we wish to achieve tissue regeneration.
Most significantly, we have demonstrated that NETosis is acutely modulated by electrospun template architecture and composition in vitro and in vivo and shown that a high degree of NETosis in vivo may result in a proinflammatory microenvironment that inhibits marginal tissue integration and angiogenesis. Further characterization of extruded NETs by immunoreactivity assays that colocalize CitH3 with other neutrophil granule contents contained within NETs may provide additional valuable information for further characterizing the response to electrospun templates and the microenvironment it creates. Our results thus far suggest that investigating the neutrophil's specific response to electrospun templates and other biomaterials is an avenue of utmost importance.
Footnotes
Acknowledgment
This work was supported by the Helen Hardin Honors Summer Research Fellowship Program at the University of Memphis and the Memphis Research Consortium.
Disclosure Statement
No competing financial interests exist.