
Abstract
Background:
Glutaraldehyde (GLA) has been used to crosslink bioprosthetic heart valve (BHVs) tissues to enhance their stability, besides ensuring a satisfactory degree of immunological tolerance. Unfortunately, GLA fixation does not guarantee a complete tissue biocompatibility of BHVs in currently used devices. The interaction between preformed human anti-alpha-Gal antibody and alpha-Gal antigens promotes the calcification of GLA-treated alpha-Gal–positive tissue. Recently, an alarming correlation between the presence of the alpha-Gal epitope and a premature BHVs degeneracy was reported. This article presents the results of a novel treatment called FACTA, for the inactivation of the alpha-Gal epitopes in porcine aortic valve tissue and commercial BHVs.
Methods:
Evaluation of the alpha-Gal epitope inactivation was performed through a patented ELISA test, confirmed by western blot, immunofluorescence, and immunohistochemical analyses. Investigations were also conducted to assess the in vitro propensity to trigger thrombosis, calcification, and worsening of FACTA-treated tissue. To explain the mechanism of action through which the FACTA treatment acts, a specific experimental model, based on the mass spectroscopy approach, was performed.
Results:
The study confirms that GLA is able to ensure the inactivation of approximately half alpha-Gal epitopes originally present in both porcine aortic valve tissue and marketed BHVs. By subjecting tissues to the FACTA procedure, it was possible to obtain an alpha-Gal inactivation degree of about 95% alongside to a reduced propensity from 72.6% to 85.4% to the in vitro calcification for porcine aortic valve tissue and 80.5% for commercial treated BHVs. FACTA was effective in decreasing oxidative tissue damage and protecting collagen from degradation. Finally, FACTA could further mitigate or even abrogate the need for early anticoagulation therapies after BHV implantation.
Conclusion:
A novel treatment, called FACTA, is effective to produce biological tissues that are less susceptible to enzymatic and oxidative stress and structural degradation, calcification, and thrombus formation. FACTA-treated tissues display a clear improvement of their biocompatibility that is characterized by an almost complete inactivation of the alpha-Gal epitope. FACTA prevents the xenogeneic tissue antigens from reacting with the host immune system, ensuring an effective shield effect that makes the tissue surface less reactive and more biocompatible.
Introduction
H
The most common method of BHV preservation consists of subjecting the tissue to glutaraldehyde (GLA) fixation treatment (GFT). For more than 40 years, GFT has been the standard chemical process adopted to ensure tissue biocompatibility, increased mechanical strength, sterilization, and safe storage. BHVs express specific epitopes for their species of origin, which are not present in humans, and whose elimination or inactivation is essential to avoid tissue rejection. Unfortunately, GFT is not able to ensure a complete masking of the xenogeneic determinants. 9 In particular, the residual presence of the alpha-Gal xenoantigen significantly results in an increase of the human anti-galactose titers 10 specifically directed to the Galα1-3Gal residue, starting from day 10 after BHV implantation 11 and reaching a peak at around 3 months 12 for both IgM and IgG isotypes. The in vivo interaction between antigen and antibody was shown to exert a powerful pro-calcific effect.13–16
This article presents the results of a novel, patented-pending treatment called FACTA, for the inactivation of the alpha-Gal epitopes in porcine aortic wall tissue, both before and after GFT.
Treated commercially available porcine and bovine BHVs leaflets were also investigated.
Materials and Methods
Porcine native tissue treatment
Porcine hearts were harvested from Dutch Landrace × Large White pigs at a local abattoir. The aortic valves were isolated within 2 h of death and rinsed with cold saline for blood removal. A portion of the tubular ascending aorta (immediately above the sinotubular junction) was cut into small pieces weighing about 100 mg, gently blotted on filter paper to remove the excess of isotonic solution, and stored at −20°C until treatment. Samples were grouped as follows:
• N group: native tissue subjected to FACTA procedure; • G group: tissue fixed in GLA solution; • TG group: tissue subjected to FACTA procedure before GLA fixation; • GT group: tissue subjected to FACTA procedure after GLA fixation;
The choice of using porcine aortic wall samples to assess the efficacy of the FACTA treatment in the alpha-Gal inactivation investigations was contingent to the reliability of the procedure in critical situations, such as in a very compact and high-thickness tissue. A comprehensive summary of all samples and treatments abbreviations is reported in Table 1.
Porcine knockout tissue
Porcine alpha1,3-galactosyltransferase (alpha-GT) knockout (KO) animals do not express the alpha-Gal epitope as they are not able to synthesize the GT enzyme, which catalyzes the expression of the alpha-Gal residue on the cell surface glycoproteins and glycolipids. 17 Such animals were obtained by somatic cell nuclear transfer as previously described by using nuclei from a male fibroblast cell line alpha-GT−/− (provided by David Sachs, MGH). 18 Tissues from the alpha-GT−/− lines were used as an absolute negative control to validate our alpha-Gal inactivation procedure in treated samples. The alpha-GT KO feature of the adopted porcine tissues was previously confirmed by both long-range PCR and flow cytometry of porcine fibroblast from cloned pigs belonging to the same line.
Porcine samples of alpha-Gal KO tubular ascending aorta were carried to the laboratory in ice-cold sterile phosphate-buffered saline (PBS) with 1% penicillin and streptomycin (Sigma, St. Louis, MO). Subsequently, they were cut into small pieces weighing about 100 mg, gently blotted on filter paper to remove the excess of isotonic solution, and stored at −20°C until analysis. These samples were labeled as KO group.
Commercially available BHVs
Commercially available pericardial (Perimount PLUS; Edwards Lifesciences, Santa Ana, CA) and porcine (SAV; Edwards Lifesciences) BHVs, manufactured for clinical use, were analyzed. All BHVs were extracted from their packaging and washed in sterile PBS, pH 7.4, at room temperature (RT) for 15 min (three times), as required by the manual for “Preimplantation Directions for Use” provided by the manufacturer. BHV samples were grouped as follows:
S group: untreated tissue; ST group: tissue subjected to FACTA procedure;
A comprehensive summary of all samples and treatments abbreviations is reported in Table 1.
GLA fixation treatment
GLA fixation was performed as described by Stacchino et al. with an increasing GLA concentration up to 0.5% in PBS, pH 7.4, at RT in a dark room. 18 Subsequently, tissue samples were washed three times, 10 min each, in PBS.
FACTA tissue treatment
FACTA treatment is a novel, patent-pending procedure, designed to inactivate the alpha-Gal epitope in animal soft tissues, providing a starting material for the manufacture of BHVs intended for clinical use. Briefly, tissue was incubated in an isotonic solution consisting of a highly selected mixture of food-grade molecules, already used in the food industry (non-GMO) for two steps of 1 h each at RT in the dark. Subsequently, the treated tissue was rinsed with three steps of 10 min each in PBS at RT and stored at 4°C in PBS until use.
Alpha-Gal quantification test in sample tissue
Nine tissue samples for each group (N, G, TG, GT, S, ST, and KO) were gently blotted on Whatman filter paper, and their weight was recorded (weight range of about 100 mg). Subsequently, they were incubated with the primary anti-alpha-Gal antibody M86 [1:50] (mouse; LSBio, Seattle, WA) for 2 h at 37°C with gentle stirring and finally centrifuged at 14,750 g for 30 min at 4°C.
The quantification of alpha-Gal epitopes was performed by a patented ELISA test (EP2626701), and data were expressed as percentage of alpha-Gal epitope inactivation. Alpha-Gal inactivation percentage of samples belonging to the N, G, TG, and GT groups was obtained by comparison with untreated native porcine aortic wall tissue; for S and ST groups, by comparison with untreated native porcine aortic leaflets in the case of SAV bioprostheses or untreated native bovine pericardium in the case of Perimount PLUS bioprostheses. Briefly, a Polysorp 96-well plate (Nunc, Rochester, NY) was coated with 100 μL of alpha-Gal/human serum albumin (HSA; Dextra Laboratories, Berkshire, United Kingdom), 5 μg/mL, for 2 h at 37°C. After washing three times with PBS, the blocking procedure was performed by using 300 μL per well of 2% HSA (Sigma) for 2 h, at RT in darkness. Wells were then washed three times, as mentioned earlier. A set of wells was loaded with 100 μL of supernatant derived from samples of N, G, TG, GT, S, ST, and KO groups and incubated for 2 h at 37°C.
After washing, the secondary horseradish peroxidase (HRP)-conjugate antibody [1:500] (goat anti-mouse IgG; KPL, Gaithersburg, MD) was loaded. Finally, 100 μL of HRP substrate buffer was added to each well for 5 min, at RT, in darkness. The plate absorbance was measured by a plate reader at 450 nm.
Alpha-Gal quantification test for the identification of FACTA action mechanism
To assess and understand the possible action mechanism of the FACTA treatment in the inactivation of the alpha-Gal epitope, a simplified system, as described later, has been elaborated.
FACTA mixture was incubated with 5 μg/mL HSA/alpha-Gal in PBS for 2 h at RT in the dark under gentle stirring (n = 8). To evaluate the alpha-Gal inactivation percentage, the ELISA test was performed. A non-treated (NT) solution of 5 μg/mL HSA/alpha-Gal in PBS (n = 8) and 5 μg/mL HSA in PBS (n = 8) was adopted, respectively, as positive and negative controls. Solutions were subsequently incubated with the primary anti-alpha-Gal antibody M86 [1:50] for 2 h at 37°C with gentle stirring and finally centrifuged at 14,750 g for 30 min at 4°C.
After preparation of a Polysorp 96-well plate, as described earlier, a set of wells was loaded with 100 μL of supernatant derived from samples of FACTA-treated HSA/alpha-Gal, NT HSA/alpha-Gal, and HSA solutions. After washing, the secondary HRP-conjugate antibody [1:500] (goat anti-mouse IgG; KPL) was loaded. Finally, 100 μL of HRP substrate buffer was added to each well for 5 min, at RT, in darkness. The plate absorbance was measured by a plate reader at 450 nm.
To evaluate a potential direct interaction capacity by the FACTA solution toward the alpha-Gal epitope, a Polysorp 96-well plate (Nunc) was coated with 100 μL of α1–3 Galactobiose (Galα1,3Gal; Dextra Laboratories), 5 μg/mL, for 2 h at 37°C. A set of wells was incubated either with PBS or with the FACTA mixture solution for 1 h at 37°C. After washing, the primary anti-alpha-Gal antibody M86 [1:50] was incubated for 2 h at 37°C. The revelation of the M86 took place, as described earlier, through the use of the HRP-conjugate antibody [1:500].
Electrophoresis and western-blot analysis
Two samples of native porcine aortic wall (600 mg for each one) were minced and homogenized separately by using an Ultra Turrax Tube Drive (Ika, Staufen, Germany) in 5 mL of 100 mM phosphate buffer with 1.5 mM MgCl2. The homogenate was subsequently centrifuged at 17,000 g for 20 min at 4°C, and the two supernatants were retained for analysis and called NT and T samples. The T supernatant underwent FACTA treatment, and the NT sample was analyzed directly without further manipulation.
T and NT samples were loaded in two different 10% acrylamide/Tris-Glycine gels in a gel electrophoresis apparatus (Apelex, Lisses, France). HSA (Sigma) was adopted as a negative control, whereas HSA/alpha-Gal (Dextra Laboratories) was used as a positive control.
After the electrophoresis run, one gel was subjected to protein fixation in 30% ethanol (Sigma) plus 10% acetic acid (Sigma), stained with 10% acetic acid plus 60 mg/L of Coomassie Blue R-250 (Sigma), and finally destained in 10% acetic acid. The second gel was transferred to a nitrocellulose membrane by semidry blotters (Apelex). The membrane was incubated sequentially in blocking solution with 4% of HSA for 2 h at RT, primary M86 antibody [1:200] plus 1.5% HSA overnight at 4°C, and secondary peroxidase-labeled antibody [1:3000] (goat anti-mouse IgG; KPL) plus 1.5% HSA for 2 h at RT. The membrane was rinsed four times in PBS for 10 min before, and between, incubations.
Subsequently, the nitrocellulose membrane was incubated for 5 min in Supersignal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Waltham, MA) in darkness. Radiographic films were exposed for 10 min and developed.
Immunofluorescence and immunochemical analysis
To confirm the data obtained from the alpha-Gal ELISA test, and to better evaluate epitope distribution and inactivation by visual inspection, samples of G, TG, and KO groups were processed for immunofluorescence and immunohistochemical analysis. Tissues were embedded in OCT compound (Tissue Tek; Sakura Finetek, Tokyo, Japan), cryo-cooled in liquid nitrogen, and cut into 5-μm cryosections.
For immunofluorescence analysis, triple staining with anti-alpha-Gal M86 monoclonal antibody [1:10], anti-collagen I [1:25] (rabbit; GeneTex, Irvine, CA), and 4,6-diamidino-2-phenylindole (DAPI; Sigma) was performed. Recognition of M86 was carried out by a secondary rhodamine-labeled antibody [1:10] (goat anti-mouse; KPL) and for anti-collagen I by a secondary fluorescein-labeled antibody [1:10] (goat anti-rabbit; KPL).
To perform immunohistochemical analysis, sample slices were incubated with a PBS solution containing 0.3% of hydrogen peroxide for 10 min to inactivate endogenous peroxidase. The same anti-alpha-Gal primary antibody was used. Detection was carried out by peroxidase-labeled secondary antibody [1:100].
Accelerated oxidative and enzymatic damage model
Six tissue samples for each of G, GT, and TG groups were placed in 1% H2O2/100 μM FeSO4 for 8 days, with solution changes every 2 days. Three samples for each group were subsequently exposed to collagenase digestion (collagenase 600 U/mL) for 24 h at 37°C. A new set of five tissue samples for each of G, TG, and GT groups were weighted before and after collagenase digestion.
All the treated samples (oxidated, collagenase digested, and oxidated plus collagenase digested) were embedded in OCT compound (Tissue Tek; Sakura Finetek), cryo-cooled in liquid nitrogen, and cut into 5-μm cryosections. Finally, a staining with Picrosirius red was performed to evaluate the degree of collagen tissue damage. As a control, a sample from each group (G, GT, and TG) was analyzed by Picrosirius red staining without oxidation or enzyme digestion.
For the Picrosirius red staining, sample slices were incubated for 1 h in Picrosirius red solution (0.1% of Direct Red 80 in saturated aqueous solution of 1.3% picric acid; Sigma-Aldrich, St. Louis, MO). Subsequently, tissue slices were washed in two changes of acidified water (0.5% of glacial acetic acid in distilled water). Finally, samples were dehydrated in three changes of 100% ethanol (Sigma-Aldrich), cleared in xylene, and mounted in a resinous medium.
In vitro calcification propensity model
Five samples for each of G, GT, TG, S, ST, and KO groups were gently blotted on filter paper, and their weight was recorded (for S and ST specimens, only Perimount PLUS pericardial heart valve bioprosthetsis was analyzed). Samples were subsequently incubated in pooled normal human serum (Innovative Research, Peary Court Novi, MI) with 2% of penicillin and streptomycin (Sigma) for 14 days at 37°C. As a control, a set of samples was incubated just in PBS in the same condition.
After incubation, samples were washed twice in PBS for 10 min; four specimens for each human serum group and control samples were subsequently subjected to acid hydrolysis in HCl 6 N at 110°C for 12 h. Calcium evaluation was performed in hydrolyzed samples by inductively coupled plasma according to the directives of the EPA6010D method and expressed as ug Ca2+/mg of dry defatted weight (ddw). The remaining human serum specimens (one for each group) underwent Von Kossa histochemical staining.
ddw was determined by comparing lyophilized dry weight samples before and after delipidization treatment. After the lyophilization step, sample tissues were incubated for 36 h under 10 kPa over P2O5 at 37°C until a constant dry weight was attained. Defatted procedure was carried out by incubation of tissue specimens in ascending series of alcohols followed by two steps of chloroform/methanol (2:1 and 3:1), in a descending series of alcohols and finally in water.
Thrombin generation assay
Four samples of untreated commercially available porcine and bovine BHVs and four samples of the same BHVs treated with FACTA protocol underwent a Thrombin Generation Assay Test (Haemoscan, Groningen, Netherlands). Specimens were processed by following the instructions provided by the manufacturer. Briefly, tissues were incubated in modified human plasma (plasma was provided by the manufacturer) with subsequent withdrawals at different time points. Thrombin concentration of the samples was determined from a calibration curve of optical density at 405 nm. The thrombin generation curve for each specimen was constructed by plotting the thrombin concentration versus the time points at which the samples were taken. The curve is used to determine the speed of thrombin generation, expressed as per cm2 of sample. Reference materials were provided by the manufacturer, in particular:
• Polydimthylsoloxane (REF 1)—low propensity to thrombin generation; • Medical steel (REF 2)—high propensity to thrombin generation;
Complement convertase assay
Four samples of untreated commercially available porcine and bovine BHVs and four samples of the same bioprostheses treated with FACTA protocol were subject to a Complement Convertase Assay Test (Haemoscan). Specimens were processed by following instructions provided by the manufacturer. Briefly, tissues were incubated in modified human plasma (plasma was provided by the manufacturer); during incubation, the complement factors can bind the surface of the sample followed by the formation of a complement convertase complex. After incubation, specimens were washed and analyzed for complement convertase activity with a complement convertase-specific chromogenic substrate. The rate of color development determined at the optical wavelength of 405 nm was dependent on how much convertase was generated on the surface of tissue. Reference materials were provided by the manufacturer, in particular:
• Low-density polyethylene (REF 1)—low-complement convertase activity; • Medical steel (REF 2)—high-complement convertase activity;
Mass spectroscopy investigation for identification of FACTA action mechanism
The same solutions adopted in the Alpha-Gal quantification test for the identification of FACTA action mechanism (FACTA-treated HSA/alpha-Gal, NT HSA/alpha-Gal, and HSA) were investigated by mass spectrometry by use of a 40 nL C8 nano-precolumn coupled to a C8 160 nL analysis nano-column directly linked to a 6520 Q-TOF ESI-source (Agilent, LC-CHIP MS system). A switching valve between pre-column and column allowed a precise timing and extensive treatment of samples before chromatographic separation and sample ionization.
Briefly, 8 μL (about 600 femtomol) of each sample was loaded on the pre-column and flushed with 4 μL by using 0.1% formic acid in water (Eluent A) and 0.1% formic acid in acetonitrile (Eluent B), with a ratio of 95:5. Then, it was switched to the 160 nL analysis column and resolved in 25 min with a gradient from 10% to 70% eluent B at 0.35 μL/min flow rate.
Eluting proteins were ionized at 1700 V, 325°C source gas temperature, and 4.8 L/min gas flow rate. Acquisition parameters were set at MS scan rate of one spectra/s in the range between 130 and 1700 mz in high-resolution mode (4 GHz, resolution 20k).
All data were acquired in profile mode and analyzed with Agilent Mass Hunter Qualitative Analysis (release B.06.00) with pMod deconvolute (MS): protein tool, an improved version of maximum entropy algorithm. Deconvolution on MS signals from whole protein was performed in suitable limited m/z range encompassing at least seven differently charged clusters and setting baseline subtraction factor to minimum value, peak filters to 10% of largest peak relative height, and pMod significance filter to a value greater or equal to 20.00. Mass range considered for deconvolution was set from 30,000 to 130,000 Da; all other parameters were left at their default values.
Statistical analysis
Data are reported as mean ± standard deviation. For comparison between groups, Student's t-test and analysis of variance (ANOVA) were performed. A value of p ≤ 0.05 was considered statistically significant. Data were elaborated with Microsoft SPSS 11.0, SigmaStat 3.11.0, and G-Power software. The sample number for each single analysis is given in the caption of each individual graph/figure.
Results
Alpha-Gal quantification test in sample tissue
The ability of the FACTA treatment to prevent the binding between the alpha-Gal epitope and the M86 antibody was assessed directly on native tissues (N group) and compared with the degree of antigen masking obtained by the application of GLA treatment alone (G group). The percentage of alpha-Gal inactivation exhibited by FACTA was almost two times greater than GLA fixation, as shown in Figure 1A (n = 9 for each group samples, N vs. G p < 0.05).
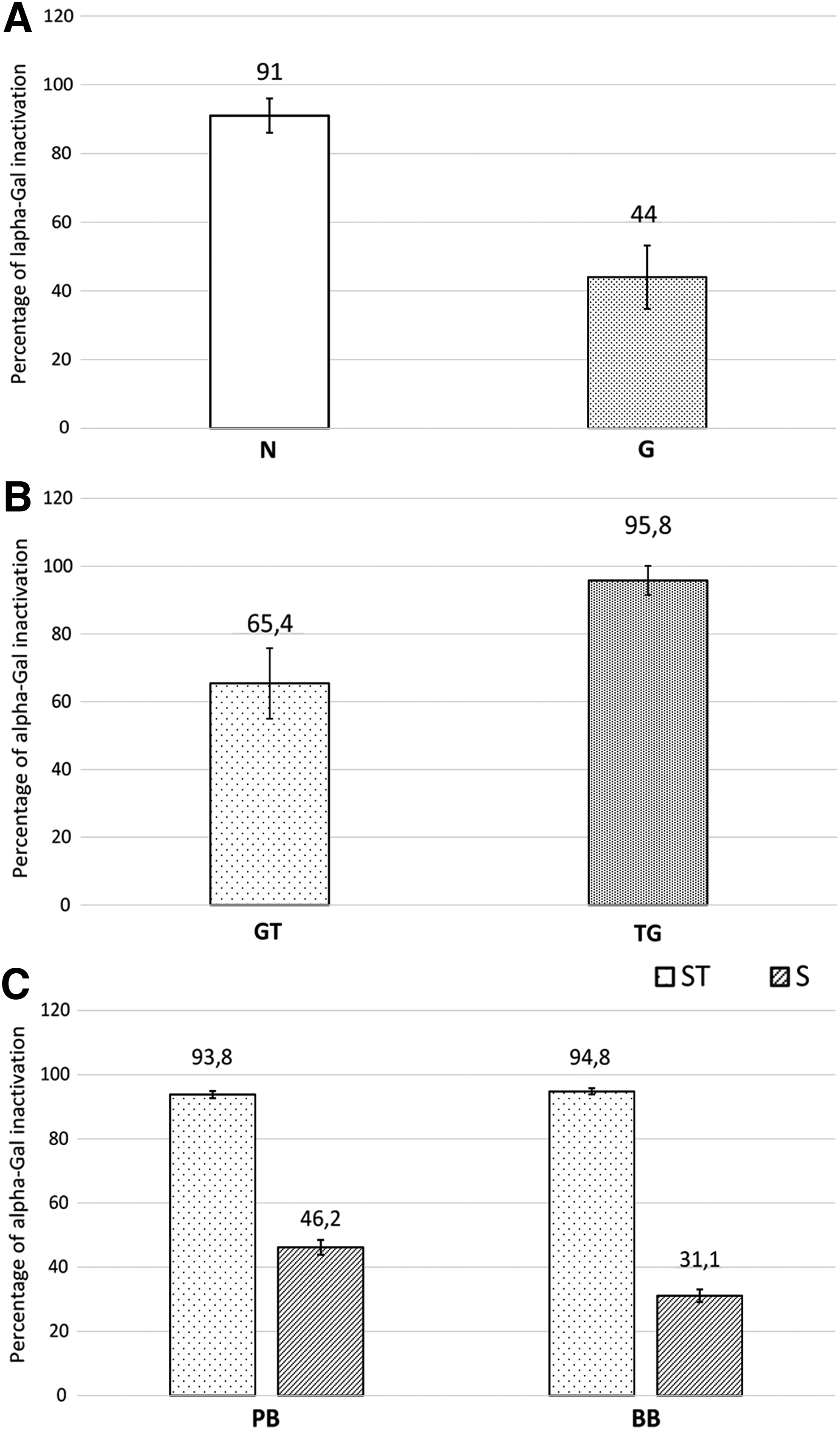
However, since BHVs are manufactured by using a crosslinking treatment, it is of fundamental importance to evaluate whether GLA fixation affected the efficacy of the FACTA protocol. In particular, it is necessary to determine any variation in performance caused by coupling FACTA treatment before or after GLA crosslinking. Coupling both FACTA and GLA treatments seems to achieve an improvement in alpha-Gal masking that ranges from about 20% to 51% compared with GLA alone (Fig. 1B, n = 9 for each group samples, GT vs. TG p < 0.05). Finally, the application of the FACTA treatment to both porcine and bovine commercially available BHV leaflets demonstrated an outstanding epitope inactivation of 93.8% ± 1.12% and 94.8% ± 0.96%, respectively (Fig. 1C, PB ST vs. PB S p < 0.05; BB ST vs. BB S p < 0.05; PB ST vs. BB ST p > 0.05). As previously reported, samples of the KO group do not display any presence of alpha-Gal epitopes (data not shown).10,13,15
Electrophoresis and western blot analysis
Two gels of 10% acrylamide/Tris-Glycine were prepared with the same type of samples and the same load sequence. The gel stained with Coomassie Blue showed the presence of the control specimens consisting of HSA and alpha-Gal/HSA distributed properly according to their molecular weight (Fig. 2). In fact, following the manufacturer's instructions, the reported molecular weights were 65 and 85 kDa for HSA and alpha-Gal/HSA, respectively. The protein content of porcine aortic wall extracts (NT and T samples) was separated with good resolution. The FACTA treatment does not seem to affect the protein set of the tissue extract.

10% acrylamide/Tris-Glycine gel stained with Coomassie Blue R-250 on the left and protein immunoblot (western) on the right. Samples were identified, as indicated later: (St) Protein ladder; (1) HSA; (2) alpha-Gal/HSA; (3) Untreated porcine aortic wall extract, NT sample −15 μL; (4) Untreated porcine aortic wall extract, NT sample −10 μL; (5) empty well; (6) FACTA-treated aortic wall extract, T sample −15 μL; (7) FACTA-treated aortic wall extract, T sample −10 μL. HSA, human serum albumin.
The immunoblot, incubated with the M86 antibody, revealed the ability to discriminate, in the nitrocellulose membrane, the presence or the absence of alpha-Gal epitopes (Fig. 2, column no. 1 negative control and column no. 2 positive control). The NT aortic wall extract (Fig. 2, NT sample, columns no. 3 and 4) exhibited a diffuse presence in the protein bands of the alpha-Gal xenoantigen, which was not detectable after the FACTA procedure (Fig. 2, T sample, columns no. 6 and 7).
Immunofluorescence and immunohistochemical analysis
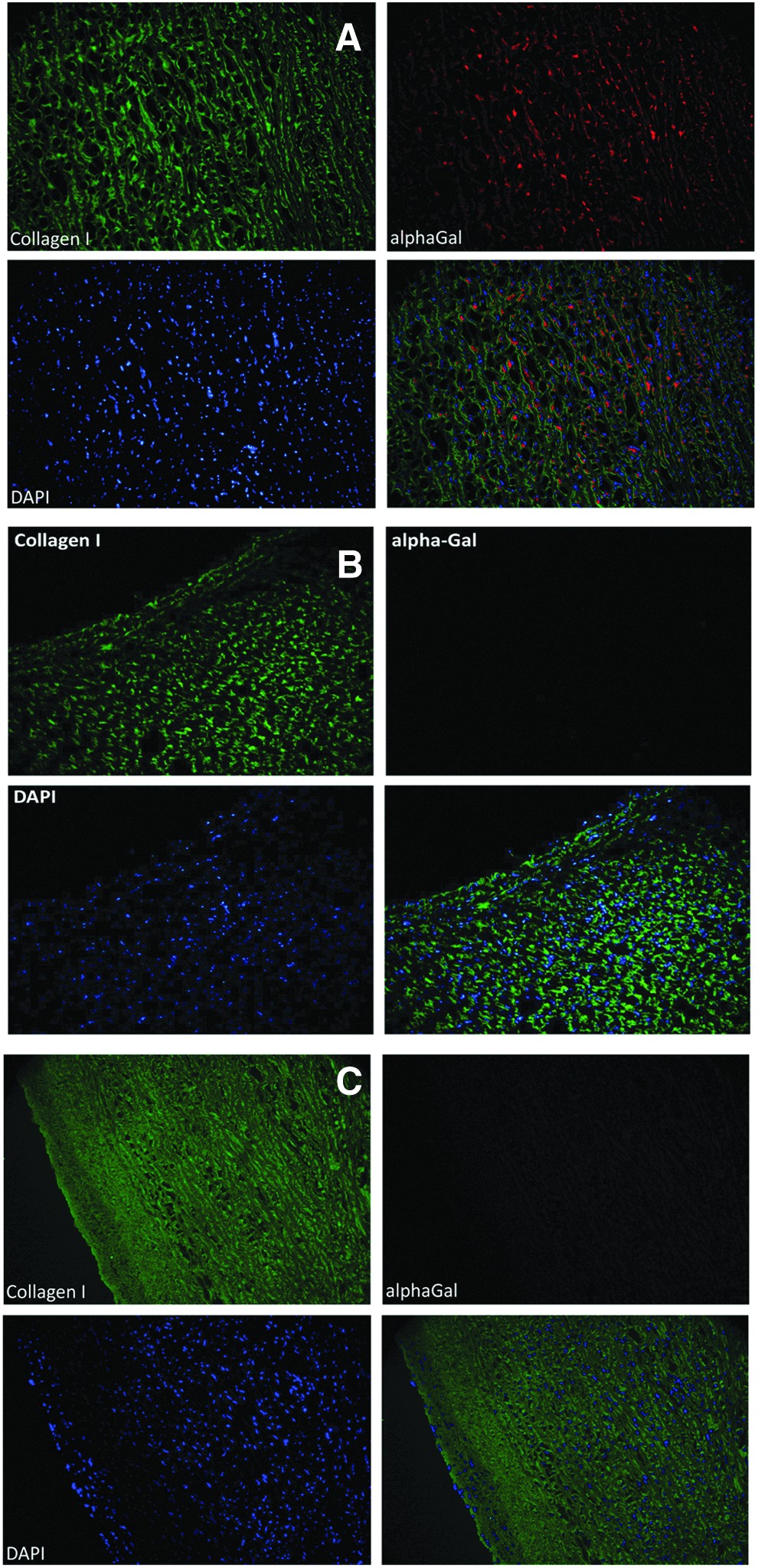
Porcine GFT aortic wall samples exhibited a diffuse presence of the alpha-Gal epitope in the inner stroma of extracellular matrix (ECM), as shown in Figure 3A, whereas the xenoantigen was not detectable in the porcine alpha-Gal KO sample (Fig. 3B). The FACTA treatment applied to the aortic wall before GLA fixation has demonstrated the ability to prevent epitope recognition and binding by the M86 antibody, making the treated tissue similar to that of KO ones (Fig. 3C).
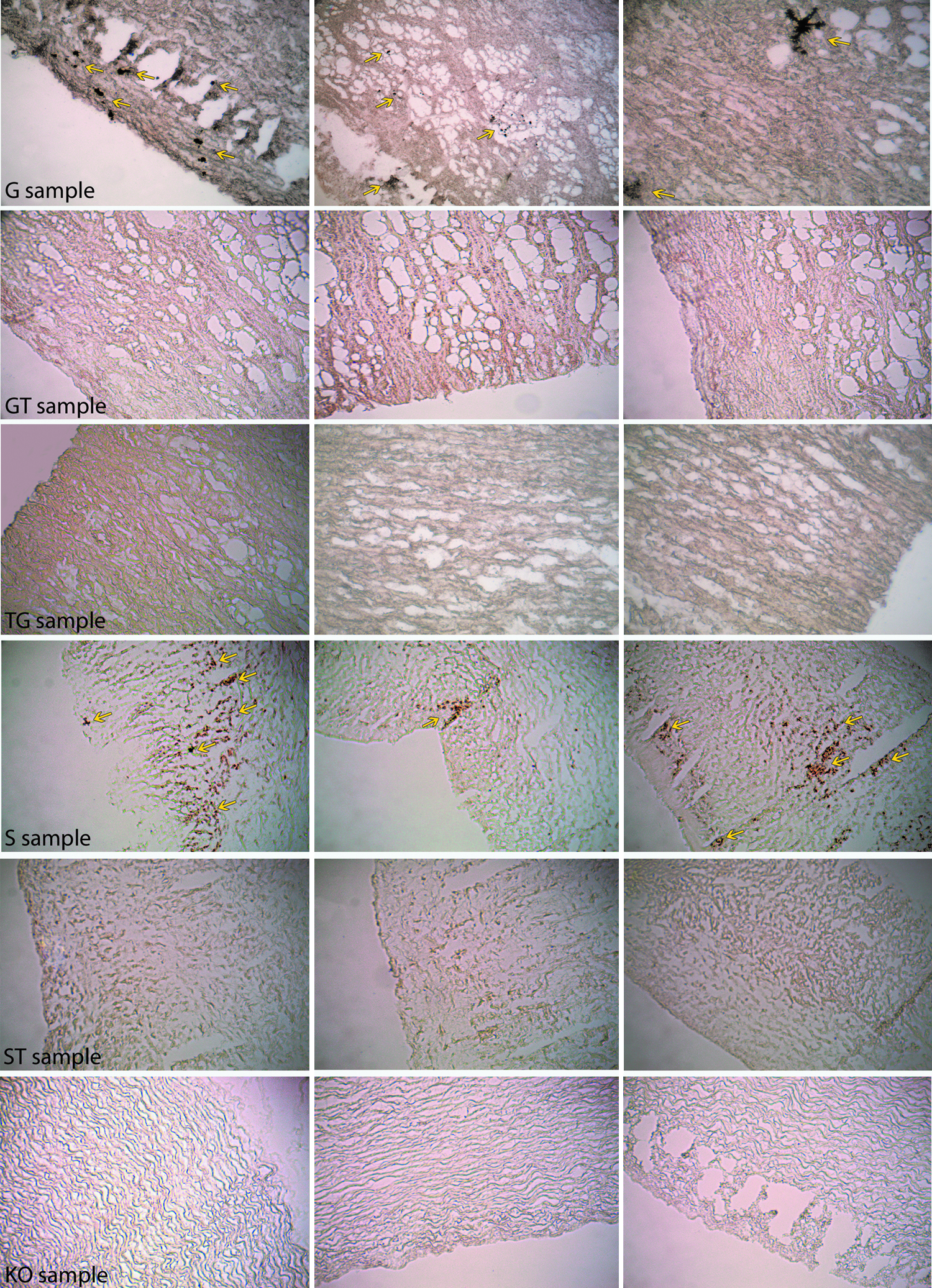
Immunohistochemical staining of GFT porcine tissue revealed a diffuse HRP reaction, which confirms a spread presence of the alpha-Gal epitope, particularly focused on the endothelial layer (Fig. 4A, G sample, brown color), unlike the alpha-Gal KO sample that does not exhibit any positivity (Fig. 4B, KO sample). The FACTA treatment seems to be able to guarantee full coverage, capable of masking the alpha-Gal epitopes, especially on the outer portion of the tissue samples (Fig. 4C). Porcine tissue treated with the FACTA protocol and subsequently crosslinked with GLA (Fig. 4D, TG sample) shows a very similar appearance to the alpha-Gal KO.

Horseradish peroxidase immunohistochemical analysis performed on porcine aortic heart valve wall tissue after GLA fixation
The complete absence of the alpha-Gal epitopes in the FACTA-treated samples (TG) evaluated both by immunofluorescence and HRP immunohistochemical analysis, resulted evident and shared to both investigation techniques (Fig. 3C, no signal detection in the red channel; Fig. 4D complete absence of brown color). Conversely, the GLA fixed samples (G) seems to exhibit a greater and widespread presence of the alpha-Gal xenoantigen if analyzed by HRP immunohistochemistry in comparison to immunofluorescence. However, this over-expression is only apparent because the amount of epitopes in different areas of the same tissue specimen are almost identical when quantified by the ELISA test.
Accelerated oxidative and enzymatic damage model
Picrosirius red staining of control tissue revealed a greater compactness of the collagen structure in the samples subject to FACTA procedure (GT and TG samples) compared with G specimens, which seem to be affected by a high degree of porosity (Fig. 5). In GT and TG samples, a densely colored area is visible corresponding to the intima layer, probably attributable to the effect of the FACTA treatment. After 8 days in a pro-oxidant environment (1% H2O2/100 μM FeSO4), the G sample appears heavily deconstructed, with cracks and a general thinning. The GT sample exhibited diffuse collagen collapse; conversely, TG specimens maintained good tissue structure with a preserved appearance of the collagen fibers (Fig. 5, yellow arrow) and limited cracking. Interestingly, a clearer area with a distinguishable fibrous component was revealed, corresponding to the intima layer of the TG sample, probably as a result of FACTA treatment performed before the GLA crosslinking step.

Picrosirius red staining of G, GT, and TG samples after incubation in an oxidative environment, collagenase digestion, and both. (Magnification 10×; collagen fibers with a good mainteinance of the fibrous structure are identified by yellow arrows).
Collagenase digestion is responsible for the onset of cracks that leads to a decrease of tissue density in G and GT samples (Fig. 6, −25.44% ± 1.6% and −11.9% ± 0.4% of wet weight loss, respectively, n = 5 for each sample group, G vs. GT p < 0.05). Conversely, after enzyme degradation, TG specimens maintained their collagen structure with a physiological fibrous appearance (Fig. 5, yellow arrow) and with a limited wet weight loss (Fig. 6, −2.97% ± 0.4%, n = 5, TG vs. G p < 0.05, TG vs. GT p < 0.05). Once again, the intima layer is visible as a densely colored area similar to that of the TG control sample.
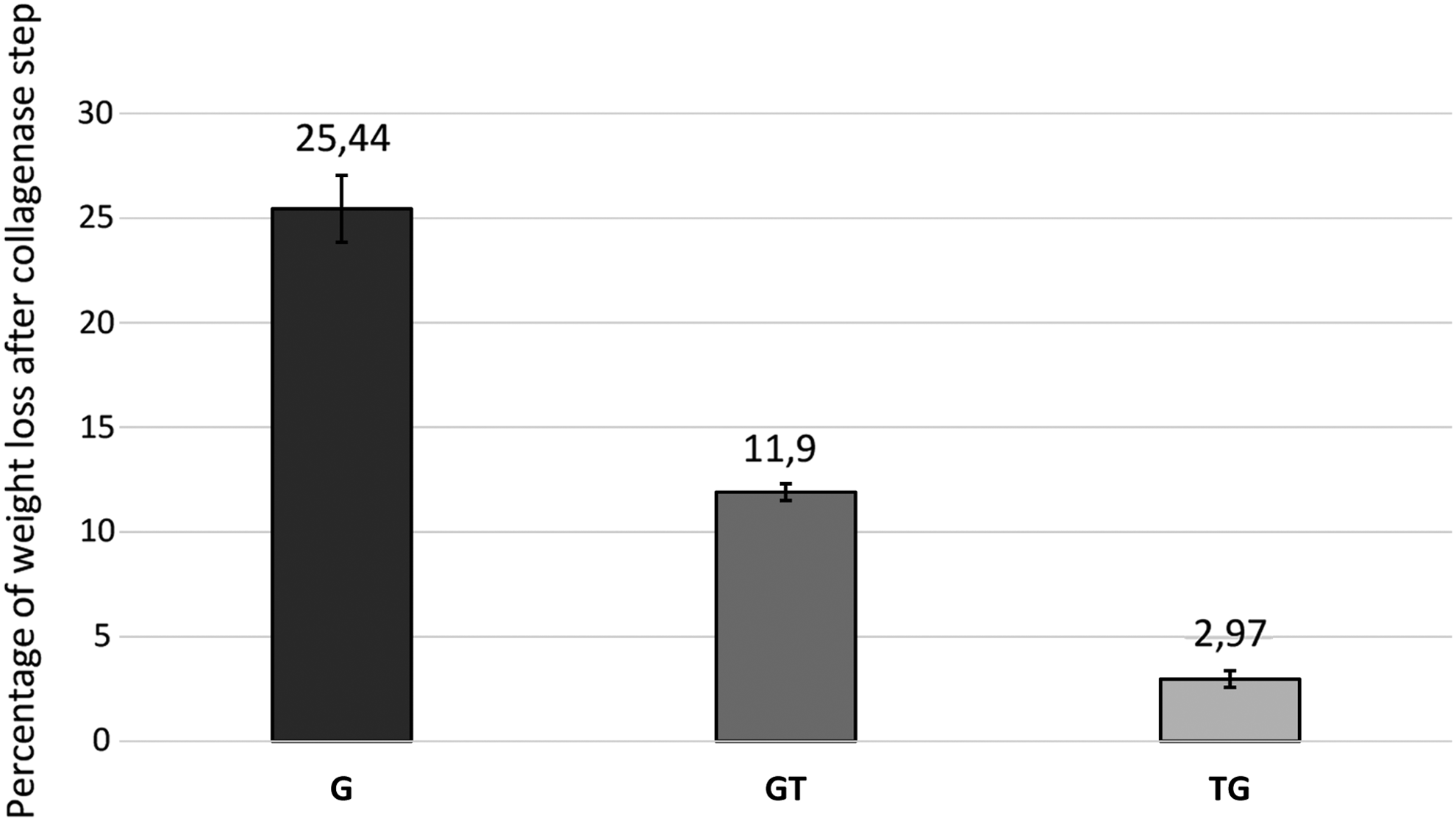
Comparison between percentage of wet weight loss after collagenase digestion in G (25.44% ± 1.6%), GT (11.9% ± 0.4%) and TG (2.97% ± 0.4%) samples (n = 5 for each sample, all the differences between the values of the samples were statistically significant, p < 0.05).
Incubation in an oxidative environment followed by collagenase digestion caused substantial destruction of the G and GT sample tissues with cracks and intimal relaxation. FACTA treatment performed on tissues before the GLA fixation step resulted in a better-preserved ECM with a normal fibrous appearance (Fig. 5, yellow arrow), despite the heavy degradative stress to which the sample was subjected.
In vitro calcification propensity model
FACTA treatment applied to GFT porcine aortic wall tissue has shown the ability to decrease the propensity of in vitro calcification to about 72% (GT sample) and 85% (TG sample) compared with G samples (Fig. 7). Treatment performed before GLA fixation was twice as effective in preventing the precipitation of calcium salts in tissue (TG 7.5 ± 2.3 vs. GT 14.1 ± 2.9 μg Ca2+/mg of ddw, GT vs. TG p < 0.05). Leaflets of bovine pericardial bioprostheses (S sample) exhibit a calcium concentration that is five times greater compared with cusps treated with the FACTA procedure (ST sample; 92.3 ± 33.5 vs. 18 ± 7.8 μg Ca2+/mg of ddw, respectively, ST vs. S p < 0.05). The alpha-Gal KO has reported a calcium content that is similar to that of TG ones (9.0 ± 2.8 and 7.5 ± 2.3 μg Ca2+/mg of ddw, respectively, KO vs. TG p > 0.05).
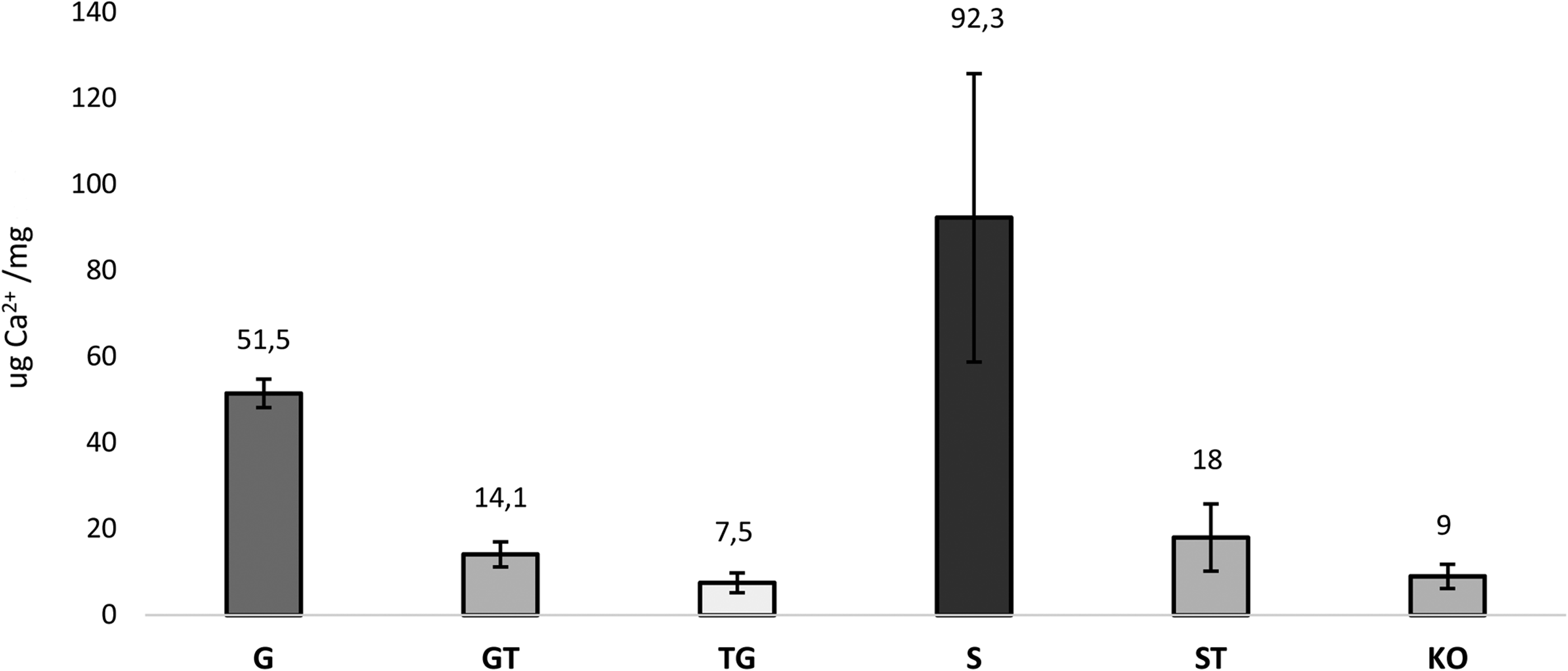
Comparison of calcification express as μg/mg of dry defatted weight between different samples (n = 4 for each ones) after 14 days of incubation in pooled normal human serum plasma at 37°C. For S and ST specimens, Perimount Plus pericardial heart valve model was investigated. GT (14.1 μg/mg ± 2.9) versus TG (7.5 μg/mg ± 2.3) p < 0.05, GT (14.1 μg/mg ± 2.9) versus ST (18 μg/mg ± 7.8) p > 0.05, TG (7.5 μg/mg ± 2.3) versus ST (18 μg/mg ± 7.8) p < 0.05, TG (7.5 μg/mg ± 2.3) versus KO (9 μg/mg ± 2.8) p > 0.05, and ST (18 μg/mg ± 7.8) versus KO (9 μg/mg ± 2.8) p > 0.05.
Von Kossa histological staining was consistent with the analysis of calcium quantification (Fig. 8): G and S samples exhibited a diffuse and important presence of calcium deposition (yellow arrow). Calcium aggregates were not detectable in the remaining analyzed samples (GT, TG, ST, and KO).
Visual investigation of calcium deposition (yellow arrow) by Von Kossa staining. Magnification 10 × .
Thrombin generation assay
Leaflets from commercially available porcine bioprostheses subjected to FACTA treatment demonstrated a statistically significant decrease in thrombin production when incubated in human plasma (Fig. 9A, PB T sample n = 5) compared with untreated bioprostheses (Fig. 9A, PB NT sample n = 5). Notwithstanding the similar amount of thrombin in the first minute of reaction for FACTA-treated and -untreated PB samples, the thrombin formation time curve of the PB T specimens remains always lower than the PB NT ones and the relative standard deviation intervals never intersect with each other.
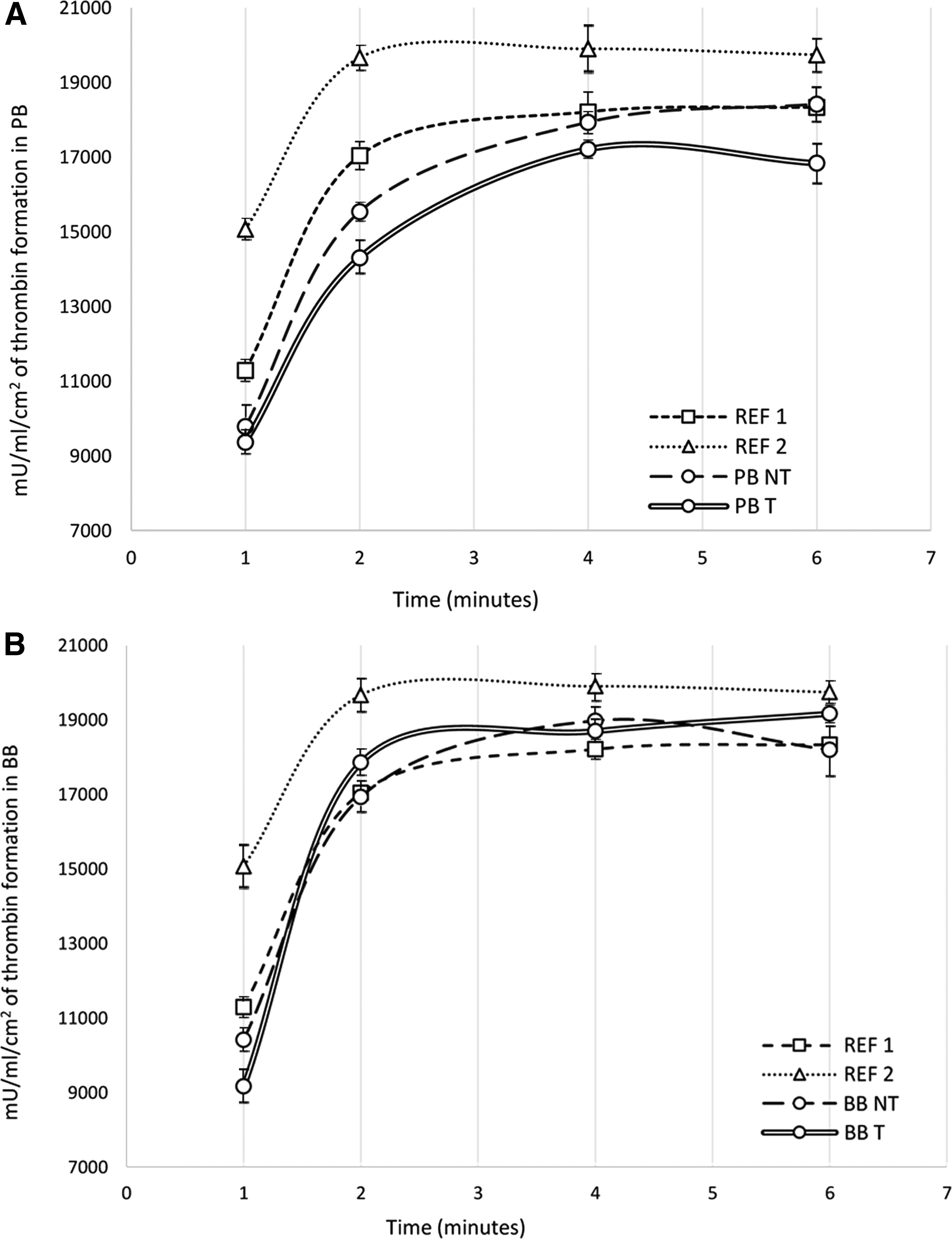
Thrombin generation assay performed on porcine commercially available bioprostheses
Cusps from commercially available bovine pericardial heart valves treated with the FACTA procedure (Fig. 9B, BB T sample n = 5) reach a peak of thrombin production equal to that of the untreated bioprostheses (Fig. 9B, BB NT sample n = 5); however, in the FACTA samples, the highest peak is reached later, thus suggesting a degree of protection toward the potential of thromboembolic events.
Complement convertase assay
Cusps from commercially available bioprostheses, of both porcine (PB) and bovine (BB) origin, show a distinct ability to activate complement. In Figure 10, the NT bioprosthetic samples were positioned in the area corresponding to a high-complement convertase activity. Despite the FACTA treatment, specimens showed no significant improvement, and this did not worsen an already suboptimal outcome.
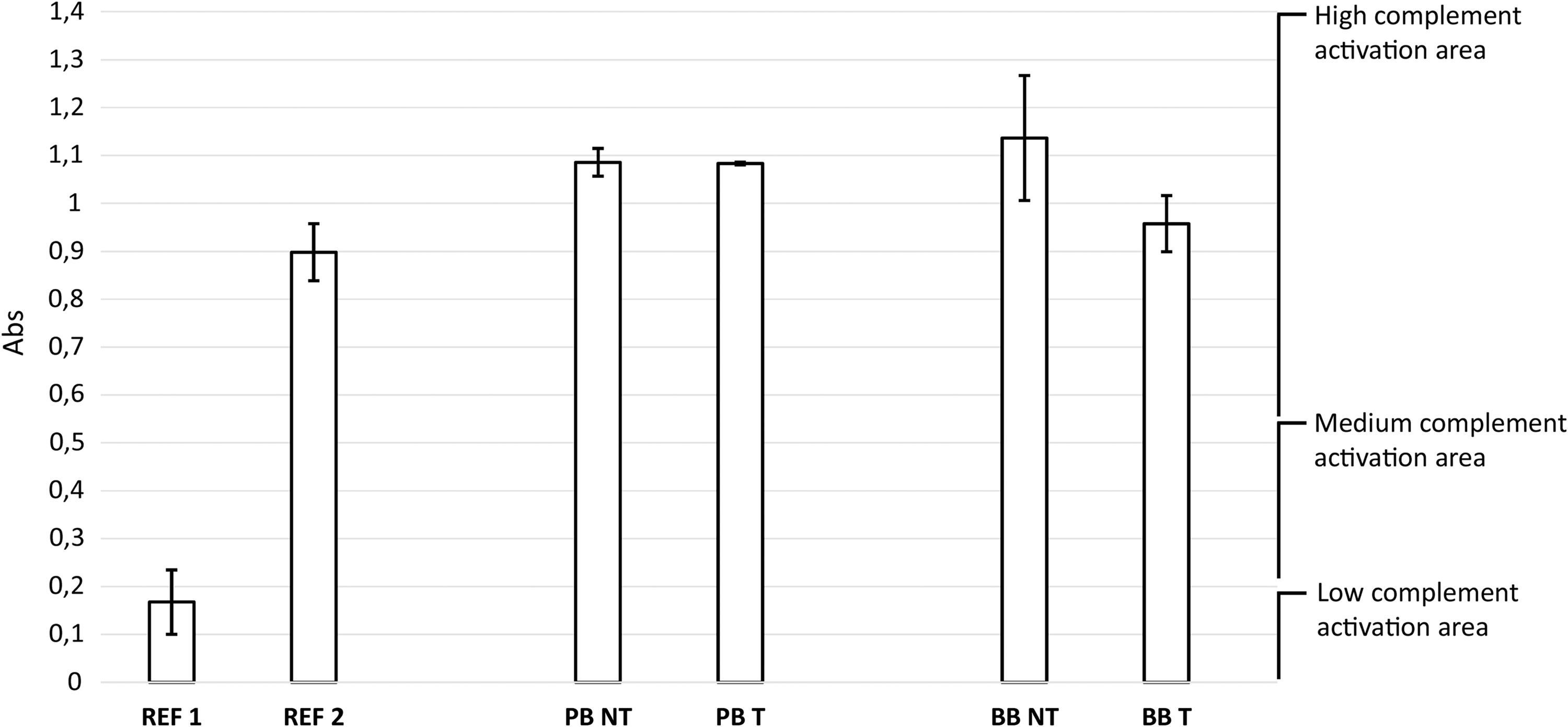
Complement convertase activity exhibited by commercially available porcine (PB sample) and bovine pericardial (BB sample) bioprostheses with (T sample) and without (NT sample) FACTA treatment (n = 4). Samples were compared with reference materials: low-density polyethylene (REF 1) and medical steel (REF 2). PB NT versus PB T, p > 0.05, BB NT versus BB T, p > 0.05.
Alpha-Gal quantification test for the identification of FACTA action mechanism
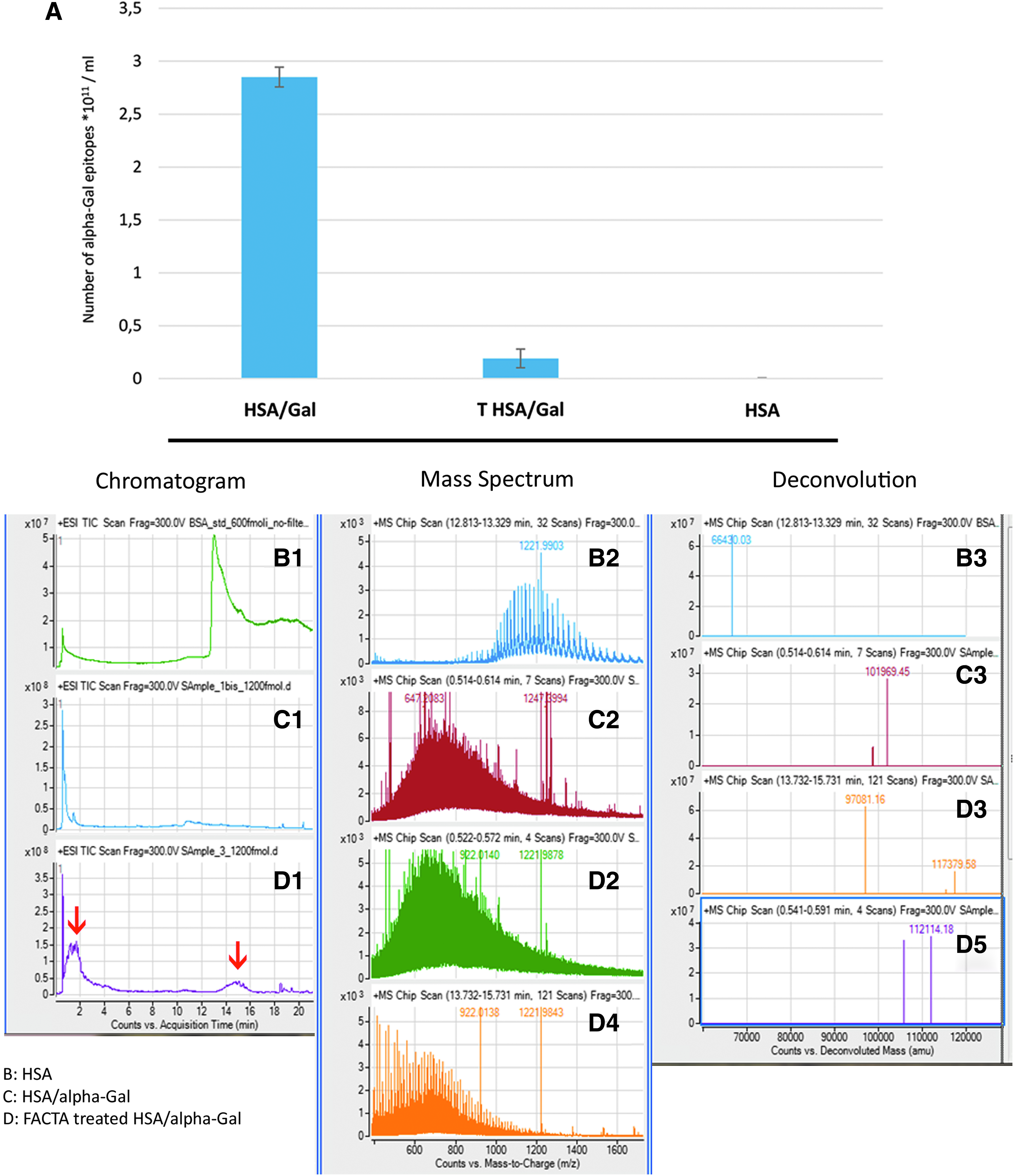
As visible from Figure 11A, the ELISA test reported the presence of 2.85 × 1011 ± 0.09 alpha-Gal epitopes for each milliliter of solution for untreated HSA/alpha-Gal. After FACTA treatment, the detectable alpha-Gal antigens were limited to 0.191 × 1011 ± 0.08, corresponding to a percentage of inactivation equivalent to 93.3 ± 1.62. The negative control constituted by HSA solution does not report the presence of epitope.
The sets of wells loaded with α1–3 Galactobiose and incubated with either PBS or FACTA solution before adding the primary M86 antibody have reported an absorbance of 0.805 ± 0.021 and 0.714 ± 0.016, respectively (n = 16 for each group, p < 0.05), indicating a potential, although limited, capacity of the FACTA in the removal of the isolated antigen.
Mass spectroscopy investigation for identification of FACTA action mechanism
Data exhibited from mass spectroscopy investigation reported the presence of proteins in all the analyzed samples. The solution of HSA was found to be eluted in 14 min (Fig. 11B1). The deconvolution of the mass spectrum showed an estimated mass of 66,430 Da, in line with the specifications provided by the manufacturer (Fig. 11B3). The NT solution of HSA/alpha-Gal exhibited an elution peak within 2 min (Fig. 11C1), with an estimated mass corresponding to the sum of the HSA mass plus the conjugated di-galactose residues (the upper range of galactosylation provided by the manufacturer is 29 residues of alpha-Gal for each HSA molecules, Fig. 11C3).
The chromatogram related to the FACTA-treated HSA/alpha-Gal solution reported the coexistence of two different protein forms (Fig. 11D1, red arrows). A first peak eluted within 2 min (Fig. 11D2), in which an increase of the molecular weight compared with the untreated solution is determinable (deconvolution Fig. 11D3) and a second peak at 14 min (Fig. 11D4) is comparable with that obtained from the solution of HSA (Fig. 11B2).
Unlike the HSA sample, the remaining solutions represent a non-homogeneous population of proteins, since there are different levels of derivatization and subsequent changes due to treatments (FACTA or di-galactose conjugation). This involves a considerable increase in the complexity of the related spectra as it appears from the comparison between Fig. 11B2, C2, D2 and D4. As a consequence, the weights obtained by deconvolution are approximate and are primarily related to the most abundant and/or more ionizable species.
The conjugation of the alpha-Gal epitope with HSA can be evaluated by a change in the retention time (2 min, Fig. 11C1) if compared with the unconjugated HSA (14 min, Fig. 11B1). This difference demonstrates, in agreement with that expected from the addition of oligosaccharides, the effectiveness of the analytical method to distinguish the presence of the alpha-Gal molecule linked to a protein component.
Discussion
GLA has been used to crosslink tissues to enhance their stability, in addition to ensuring a satisfactory degree of biocompatibility, allowing implantation of xenogeneic biological material into humans. Unfortunately, treatment with GLA is also responsible for side effects that lead to calcification. Although GLA is cytotoxic to xenogeneic resident cells, it cannot remove them from the ECM. Consequently, the physiological mechanisms of intracellular calcium elimination are compromised and, trapped in the cell cytoplasm, they themselves become a calcium nucleation site. Cell membranes and phosphorous-containing intracellular structures (e.g., nucleic acids) exposed to body fluids may also bind calcium, thus contributing to its deposition. 19 Heart valve matrix proteins such as collagen contain about 11.5% proline, 10.1% hydroxyproline, 0.65% glutamine, 0.48% aspartic acid, 0.24% serine, 0.17% hydroxylysine, and 0.15% threonine. 20 All of these have the potential to attract Ca2+, inducing hydroxyapatite nucleation. GLA, reacting only with the ɛ-amino groups of lysine residues, leaves other functional groups (such as hydroxyl terminals) free to bind calcium. 21 Moreover, GLA has previously been shown to partially achieve immunological tolerance. Specifically, half of the alpha-Gal epitopes originally present in a porcine heart valve tissue are able to react even after GFT.10–12,22 Recently, an alarming correlation between the presence of the alpha-Gal epitope and premature bioprosthetic aortic valve degenerations, within 2 years of implantation, was reported. 23
The present study confirms that GFT is able to ensure inactivation of approximately half of the alpha-Gal epitopes originally present in both porcine aortic wall tissue (Fig. 1A) and commercial available BHVs (Fig. 1C). By subjecting the xenogeneic tissue to FACTA procedure before GFT, it was possible to obtain an alpha-Gal inactivation degree of about 95% (Fig. 1B); data were further confirmed by western blot (Fig. 2), immunofluorescence (Fig. 3C), and immunohistochemical (Fig. 4) analysis. Surprisingly, in addition, FACTA treatment of commercially available porcine and bovine BHVs demonstrated similar inactivation results, with no statistically significant differences between the two models considered (Fig. 1C, PB ST vs. BB ST, p > 0.05). This finding potentially paves the way to improving the performance of currently available BHVs without necessarily changing the production process, but simply by treating them before implantation. It has been well demonstrated that the interaction between preformed human anti-alpha-Gal antibody and alpha-Gal antigen is sufficient to accelerate calcification of GLA crosslinked alpha-Gal–positive tissues in both rat and rabbit subcutaneous implantation models.13,14 Consistent with these findings, FACTA-treated tissue, in addition to demonstrating a high degree of alpha-Gal epitope inactivation, also showed a reduced propensity to calcification (determined by an in vitro model), ranging from 72.6% to 85.4% for porcine heart valve wall tissue (depending on whether the treatment is applied before or after the GFT) and 80.5% for commercial treated BHVs (Fig. 7).
The main advantage of BHVs, with respect to mechanical ones, is their freedom from anticoagulation therapies. The need for early anticoagulation after BHV implantation remains a controversial topic, with a high degree of institutional and surgeon variability. The majority of medical companies continue to recommend antithrombotic therapy for 90 days postoperatively; however, this recommendation is based on reports with conflicting results and significant limitations. 24 The treatment of commercial BHVs with FACTA (especially those of porcine origin) revealed a clear protective action against the production of thrombin (Fig. 9), either by directly inhibiting its generation or by slowing the formation mechanism (particularly for bovine pericardial BHV). Thus, FACTA treatment could further mitigate or even abrogate the need for early anticoagulation after BHV implantation.
Another important cause of bioprosthetic material failure is oxidative stress. Inflammatory cells that are recruited to the site of the prosthesis produce reactive oxygen and nitrogen species that are able to lead oxidative alteration of the proteins conformation. Reactions of oxidants with collagen result in specific structural modifications, including o,o′-dityrosinedityrosine (a tyrosyl radical-mediated crosslink), the non-physiological isomers o-tyrosine and m-tyrosine formed by hydroxyl radical modification of phenylalanine, as well as an increase in the susceptibility to degradation by proteolytic enzymes.25–27
In clinically explanted BHVs, meta and ortho isomers of tyrosine were significantly elevated. Dityrosine crosslinks in BHVs may disrupt normal collagen fiber organization, increasing matrix stiffness and contributing to valve failure through impaired mechanical function. 28 Furthermore, the exposure to oxidizing conditions results in the loss of GLA crosslinking: Previous studies have shown that rat subdermal implantation of GLA-treated BHV has decreased GLA content in explanted materials.29,30 The combination of dityrosine crosslinks formation and the loss of GLA network could affect the hemodynamic properties of the valve, resulting in clinical failure or impaired function. 28 The FACTA procedure applied before the GFT was effective in both reducing BHV oxidation (Fig. 5) and protecting collagen from degradation (Fig. 6).
The FACTA treatment resulted in an ability to interact in a stable manner with the component of the ECM since the inactivation of the alpha-Gal xenoantigen has been maintained despite some of the conducted tests involving the incubation of the samples in denaturing environments and conditions (SDS electrophoresis and mass spectrometry). The mass spectroscopy investigation has detected the increasing of the protein mass (Fig. 11D1 vs. C1, D3 vs. C3) as well as a partial removal of the alpha-Gal molecule from the protein in which it was conjugated, also confirmed by the ELISA test performed with α1–3 Galactobiose.
According to these results, the FACTA mixture seems to have been able to reach the alpha-Gal inactivation by two possible different ways: On the one hand, the FACTA molecules can interact through the protein component to create a coverage effect that is able to prevent the interaction between epitopes and antibody. This mechanism is not effective in the isolated epitope, suggesting the necessity of the presence of a proteic support for ensure the shielding effect.
The second method showed the unexpected ability for a limited removal of the alpha-Gal molecule, regardless of the presence of a proteic support.
Recently, treatment with the α-galactosidase enzyme has effectively demonstrated the removal of the alpha-Gal epitopes that are expressed on both bovine and porcine tissues,30,31 leading clinicians and researchers to consider tissue from alpha-Gal KO animals as the best source for bioprostheses manufacture.13,32,33 However, despite the guarantee of the absence of the alpha-Gal epitope in tissues, other xenogeneic antigens such as Neu5Gc are expressed and reactive.34,35 It might be interesting to know whether the FACTA treatment, applied to an alpha-Gal KO tissue, is able to inactivate the non-Gal antigenic residues, in addition to making tissue less susceptible to enzymatic and oxidative structural degradation, calcification, and thrombus formation.
The main limitations of this study are inherent to the use of an in vitro model, restricting the generalizability of the results to the clinical setting. The next step, currently being planned, will be the confirmation of these findings by implanting FACTA-treated heart valves in an in vivo large animal model.
In conclusion, a novel treatment called FACTA has been shown to be effective in biological tissues intended for BHV manufacture that are less susceptible to enzymatic and oxidative structural degradation, calcification, and thrombus formation. Furthermore, FACTA-treated tissues display a clear improvement in biocompatibility, characterized by an almost complete inactivation of the alpha-Gal epitope, a result that is currently not possible with the classical GFT. Of particular importance is the fact that it can be applied to currently available BHVs, potentially making a new generation of more durable and biocompatible BHVs.
Footnotes
Acknowledgments
This work was financed by the company BioCompatibility Innovation. This article concerns part of the work carried out under the European Union Seventh Framework Programme Collaborative Project Translink (Grant agreement No. 603049) to Avantea. The authors express their special gratitude to Prof. David H. Sachs (MGH) for the supply of the Gal KO pig cell line.
Disclosure Statement
No competing financial interests exist.