
Abstract
Islet transplantation has provided proof of concept that cell therapy can restore normoglycemia in patients with diabetes. However, limited availability of islet tissue severely restricts the clinical use of the treatment. Thus, there is an urgent need to develop new strategies to generate an abundant source of insulin-producing cells that could be used to treat diabetes. A potential approach is the in vitro expansion of pancreatic beta cells obtained from cadaveric organ donors. However, when human beta cells are expanded in vitro, they dedifferentiate and lose the expression of insulin, probably as a consequence of pancreatic islet dissociation into single cells. We have studied whether reestablishment of cell–cell and cell–matrix relationships with a biomimetic synthetic scaffold could induce redifferentiation of expanded dedifferentiated beta cells. Cells isolated from human islet preparations were expanded in monolayer cultures and allowed to reaggregate into islet-like cell clusters (ICCs). Afterward, ICCs were embedded between two thin layers of the noninstructive self-assembling peptide (SAP), RAD16-I or RAD16-I functionalized with the integrin-binding motif RGD (RAD16-I/RGD) (R: arginine, G: glycine, D: aspartic acid), which was expected to promote cell–extracellular matrix interactions. ICCs cultured with RAD16-I were viable, maintained their cluster conformation, and increased in size by aggregation of ICCs, suggesting a self-organizing process. ICCs cultured in RAD16-I/RGD showed enhanced cell adhesion to RAD16-I matrix and reexpression of the beta cell-specific genes, Ins, Pdx1, Nkx6.1, and MafA. Redifferentiation was caused solely by bioactive cues introduced to the RAD16-I peptide since no differentiation factors were added to the culture medium. The results indicate that RGD-functionalized SAP in sandwich conformation is a promising three-dimensional platform to induce redifferentiation toward a beta cell phenotype and to generate insulin-expressing cells that could be used in diabetes therapy.
Introduction
D
Pancreatic islets extracellular matrix (ECM) comprises the peri-islet and the vascular basement membranes (BMs). The main components of the islet BM are laminin (LN) and collagen type IV, although other ECM proteins such as fibronectin and other types of collagen are also present.19–21 As in many other tissues, ECM is one of the most important components of the islet microenvironment, playing a key role in islet survival,18,22–26 proliferation,26–28 and insulin secretion.18,29–34 Integrins, the main cell adhesion receptors, connect cells to specific sites of the ECM proteins, converting extracellular stimulus into a wide spectrum of intracellular signals.23,29,30 Ligand specificity is determined by the combination of alpha and beta subunits that conform to the integrin structure. 35
Complex matrices such as 804G, rich in LN,29,31 or purified proteins from the ECM, such as collagen type I,25,36 collagen type IV,34,36 LN-111, 36 LN-332, 23 LN-411, or LN-511, 21 have been used to culture pancreatic beta cells. Compared with conventional two-dimensional (2D) cultures, these matrices provide an improved microenvironment that increases cell viability and insulin secretion. However, cells cultured in matrices of natural origin are exposed to the influence of a large variety of unknown biochemical signals that make it difficult to attribute outcomes to specific matrix components.37,38 Furthermore, when animal-derived biomaterials are implanted into the human body, an immunogenic reaction may develop. 39 Synthetic matrices have a defined composition that can overcome these limitations.40,41
Self-assembling peptide (SAP) nanofiber scaffolds are synthetic biomaterials that can form hydrogels with excellent properties to create three-dimensional (3D) cell cultures.42–45 The SAP hydrogel RAD16-I (AcN-RADARADARADARADA-CONH2) is a peptide with a β-sheet structure that self-assembles under physiological conditions into a network of interweaving nanofibers of approximately 10–20 nm diameter. These nanofibers form hydrogel scaffolds with pore sizes of 50–200 nm and more than 99% water content, 44 which are structurally similar to the natural ECM. Moreover, RAD16-I allows the study of the effect of a 3D environment per se since it is a noninstructive material. 44 In addition, the lack of chemical signaling confers the possibility of functionalization with specific cell-binding motifs to create a BM analog. RAD16-I has been shown to promote cell adhesion, growth, proliferation, and differentiation of distinct cell types.46–57 Specifically, SAPs have increased survival and insulin secretion of cultured rodent58–63 and human islets. 64
To expand beta cells in vitro, pancreatic islets are dissociated and placed in 2D culture conditions, which entail the dedifferentiation of beta cells. Therefore, we hypothesize that reestablishment of cell–cell contacts and cell–matrix relationships in a biomimetic synthetic scaffold could promote redifferentiation of human beta cells expanded in vitro. We have investigated a 3D model of RAD16-I SAP based on the sandwich configuration, 49 in which dedifferentiated islet-derived cells were embedded between two thin layers of peptide. In addition, RAD16-I was functionalized by the covalent introduction of the integrin-binding motif RGD65,66 (R: arginine, G: glycine, D: aspartic acid), known to bind to a large number of integrins, aiming to promote cell-ECM interactions and enhance redifferentiation of the cells. To determine whether redifferentiation was promoted solely by the introduction of the RGD bioactive motif, no differentiation factors were added to the culture medium.
Materials and Methods
SAPs preparation
SAP scaffolds were made by diluting 1% (w/v in water) RAD16-I (PuraMatrix; Corning, Corning, NY) in 14% sucrose to achieve a final peptide concentration of 0.3%. The RGD recognition sequence [AcN-GP
SAPs characterization
Congo red staining
When RAD16-I is tailored with peptide recognition sequences, Congo red dye binding to β-sheet structures present in the SAP RAD16-I allows the study of these secondary structures by visual examination. RAD16-I and RGD peptides were diluted at 0.3% (w/v) and samples were prepared. Constructs were then incubated with 0.1% (w/v) Congo red (Sigma-Aldrich, St. Louis, MO) for 10 min and washed extensively with phosphate-buffered saline (PBS) to remove the excess of Congo red solution. Samples were analyzed under a stereoscopic microscope (Nikon SMZ660, Tokyo, Japan).
Circular dichroism spectroscopy
Circular dichroism (CD) spectra were acquired on a JASCO-715 spectropolarimeter (Jasco Corp., Tokyo, Japan) equipped with a Peltier system. Each peptide was dissolved at 1% (w/v; stock solution) in deionized water and sonicated for 30 min. Stock solution was then diluted to a working concentration varying from 6 to 50 μM, and samples were analyzed in a quartz cuvette with a path length of 1 cm. Spectra were obtained in a range of 195–250 nm and scan speed of 20 nm/min.
Atomic force microscopy
Peptide stock solutions (1%, 10 mg/mL) were sonicated for 30 min. Afterward, samples were diluted to a concentration of 0.005% (conventional RAD16-I) and 0.01% (RGD and RAD16-I/RGD) with deionized water, and 10 μL of each peptide was placed on a mica surface of 9.9 mm diameter. Images were obtained at a resolution of 256 × 256 pixels, using an XE-100 atomic force microscope (AFM; Park Systems, Suwon, Korea), operating in noncontact mode. A silicon cantilever (ACTA 10M; Park Systems) with resonance frequency of 300 KHz, force constant of 40 N/m, tip curvature radius <10 nm, and 125 μm length was used to perform the analysis.
Cell culture
Islet cell isolation and monolayer culture
Pancreatic islets were isolated from seven adult cadaveric organ donors (Table 1) by collagenase digestion (collagenase NB1 premium grade with neutral protease NB; Serva Electrophoresis GmbH, Heidelberg, Germany) using the Ricordi method with some modifications. 69 The islets were purified on a refrigerated COBE 2991 cell processor (COBE BCT, Lakewood, CO) with a continuous density gradient. Islet purity was determined by staining with dithizone (Sigma-Aldrich). Isolated islets were cultured in CMRL 1066 medium (Mediatech, Inc., Corning Cellgro, Manassas, VA) 5.6 mM glucose supplemented with 10% ABO-compatible human serum 70 for 24–48 h until they dissociated into single cells with 0.16 mg/mL trypsin (Sigma-Aldrich) and 0.1 mM EDTA (Sigma-Aldrich). Single endocrine cells were purified by magnetic cell sorting 71 using a MiniMACS magnetic cell separation system according to the manufacturer instructions (Miltenyi Biotech, Auburn, CA). Polysialylated neuronal cell adhesion molecule (PSA-NCAM) antibody (Miltenyi Biotech) was used as a positive selection marker of endocrine cells. The resulting endocrine-enriched fractions were seeded in monolayer and cultured in expansion medium with CMRL 1066 5.6 mM glucose, supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY). The medium was changed twice a week, and the islet cell population was split 1:2 once a week using 0.25% trypsin-EDTA (Sigma-Aldrich) and expanded in culture for 7–8 passages. At passage 7–8, expanded cells were trypsinized and seeded on nonadherent bacteriological Petri dishes (90 × 14 mm; Deltalab, Rubí, Spain) at a cell density of 100,000 cells/mL and cultured in expansion media. After 48 h, the cells clustered and formed independent cell colonies (islet-like cell clusters, ICCs) that were seeded into the cell culture inserts. The experiments were approved by the Ethics Committee of the Hospital Universitari de Bellvitge.
BMI, body mass index; SD, standard deviation.
RAD16-I sandwich culture system
Cell culture inserts (12 mm diameter; Merck KGaA, Darmstadt, Germany) were placed in 24-well culture dishes and coated with 40 μL of 0.3% pure RAD16-I or RAD16-I functionalized with the RGD motif. Since RAD16-I peptide forms a hydrogel when the ionic strength increases, 200 μL of expansion medium was added underneath the insert membrane, inducing gel formation due to a self-assembling process. This volume of peptide created a layer of 0.5 mm thickness. 49 After 30 min, 10 μL of expansion media was added to the cell culture inserts that were placed in a humidified 5% CO2 incubator at 37°C and, after a minimum of 30 min, they were loaded with the ICCs. After 48 h, a washing step with 10% sucrose was performed to remove unattached clusters, the expansion medium placed outside the insert was removed, and the well rinsed with 10% sucrose. A second layer of 0.3% RAD16-I or functionalized RAD16-I/RGD was added on top of the clusters, and 200 μL of culture medium (CMRL1066 5.6 mM glucose) supplemented with insulin-transferrin-sodium selenite (ITS) (Sigma-Aldrich) was added beneath the insert membrane to initiate the self-assembling process of RAD16-I. Therefore, ICCs were embedded between two layers of 0.3% RAD16-I or RAD16-I/RGD peptide (sandwich culture system, Fig. 1). The second layer of peptide was allowed to gel for 1 h in the cell incubator and a volume of 10 μL of culture medium was then added inside the insert and kept in the cell incubator for 15 min to equilibrate. A final volume of 120 μL of culture medium was carefully added on top of the second layer of RAD16-I or RAD16-I/RGD, and 400 μL was added outside the insert. The sandwich cultures were maintained for 18 days, and 60 μL of culture medium inside the insert and 300 μL in the well were removed and replaced with fresh medium every 2 days. As a control condition, ICCs were cultured in nonadherent bacteriological Petri dishes for 18 days.
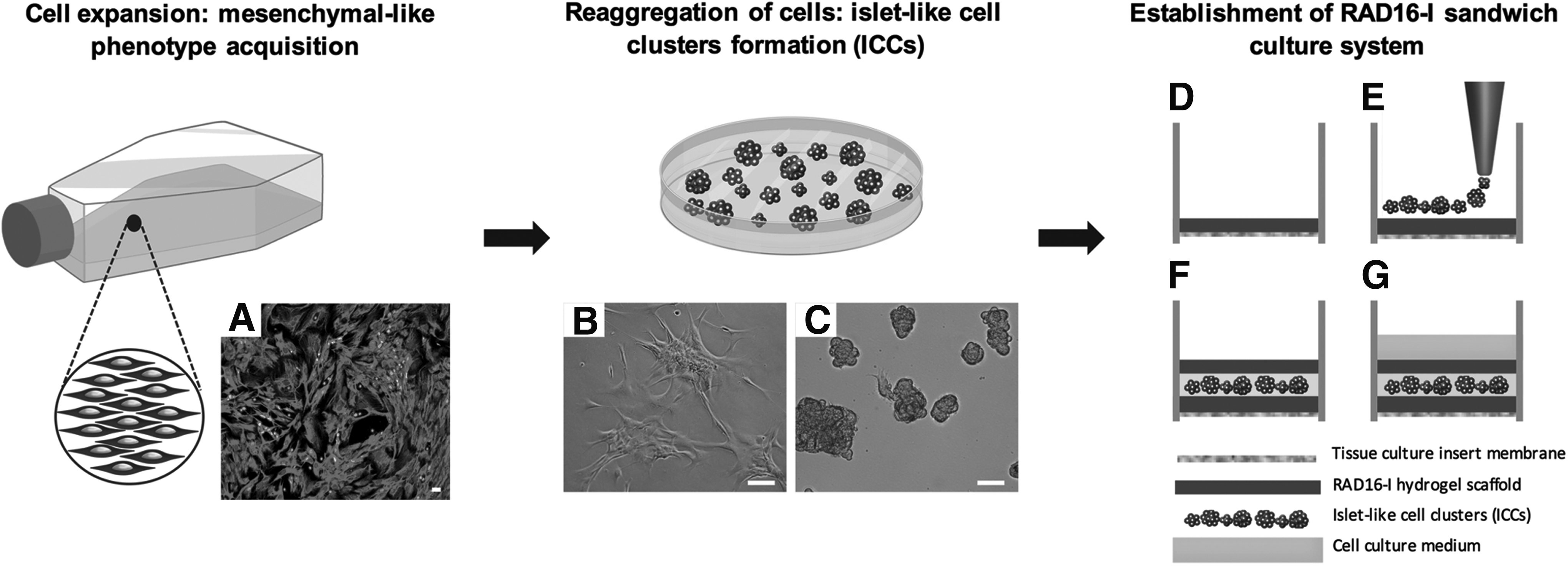
RAD16-I and RAD16-I/RGD sandwich culture model.
Immunostaining
Immunostaining was performed to characterize the cell population in magnetic activated cell sorting (MACS)- purified dispersed islet cells and in expanded cells. Islet-derived dispersed cells were cultured in flexiPERM micro 12 (Sarstedt, Nümbrecht, Germany) and fixed in 4% paraformaldehyde (PFA; Merck KGaA) at room temperature for 15 min. Antigen retrieval was performed using a microwave in citrate buffer (pH 6.0). Cells were washed, blocked for 1 h with 5% horse serum at room temperature, and then incubated overnight at 4°C with the primary antibodies: rabbit polyclonal anti-insulin antibody (1:100; Santa Cruz Biotechnology, Inc., Dallas, TX), mouse anti-vimentin antibody (1:50; Dako, Barcelona, Spain), rabbit anti-amylase (1:50; Sigma-Aldrich), and mouse anti-cytokeratin (1:80; Dako). Alexa Fluor-conjugated donkey anti-rabbit and goat anti-mouse (1:400; Invitrogen, Carlsbad, CA) were used as secondary antibodies. Nuclei were stained with 4′6-diamidino-2-phenylindole dihydrochloride (DAPI, 300 nM; Life Technologies). Images were acquired using a Leica DFC 310FX and Leica TCS-SL filter-free spectral confocal laser scanning microscope (Leica Microsystems, Mannheim, Germany). Results are expressed as percentage of positive cells over the total number of cells.
ICC characteristics
ICC morphology was assessed with an inverted microscope (Nikon Eclipse TS100). To perform fluorescence staining of the cytoskeleton and nuclei, samples were washed three times with PBS, fixed with 2% (w/v) PFA, permeabilized with 0.1% (v/v) Triton X-100, and covered with phalloidin–tetramethylrhodamine B isothiocyanate (Phalloidin-TRITC; Sigma-Aldrich) and DAPI (Sigma-Aldrich). Finally, the sandwich cultures were washed extensively with PBS and analyzed under a Zeiss Axiovert inverted microscope (Axiovert 200M; Carl Zeiss, Inc., Oberkochen, Germany) with the Zeiss ApoTome system, which allowed optical sectioning and subsequent 3D reconstruction (Z-stack images).
ICC number and area were sequentially analyzed on days 1, 10, and 18 of culture. The same inserts were analyzed at each time point. Pictures of clusters from at least three inserts from three different islet isolations were taken using an inverted microscope (Nikon Eclipse TS100), and the area of at least 80 clusters/insert was measured using ImageJ software (National Institutes of Health, Bethesda, MD).
Cell viability
MTT assay
ICC viability was measured using the quantitative colorimetric MTT assay. 72 Briefly, MTT was dissolved at a concentration of 5 mg/mL in PBS, filtered, sterilized, diluted in culture medium (0.5 mg/mL), and 500 μL was added inside the inserts on culture days 1, 10, and 18. After 3 h of incubation at 37°C, MTT solution was removed and one wash with PBS was performed. Then, 200 μL of dimethyl sulfoxide (DMSO) was added and the purple formazan crystals were solubilized by repeated pipetting. Groups of three samples were pooled, vortexed for 2 min, and the absorbance was read using a microplate reader (SpectraMax M2e) at a wavelength of 570 nm. Results were expressed as optical density (OD) and the OD of blank (DMSO) was subtracted from the samples.
Cell viability imaging
Cell viability was also analyzed with the LIVE/DEAD® Viability/Cytotoxicity Kit for mammalian cells (Invitrogen). On days 1, 10, and 18, sandwich cultures were washed three times with PBS, and a solution of calcein-AM and ethidium homodimer 1 (final concentration 2 μM) was added. After 15 min of incubation, constructs were rinsed with PBS and imaged. Afterward, samples were fixed with 2% (w/v) PFA, permeabilized with 0.1% (v/v) Triton X-100, and nuclei were stained with DAPI (Sigma-Aldrich) to properly count cell nuclei. Samples were examined under a Zeiss Axiovert inverted microscope (Axiovert 200M; Carl Zeiss, Inc.) with the Zeiss ApoTome system, which allowed optical sectioning and subsequent 3D reconstruction (Z-stack images). Results were expressed as percentage of viable cells per ICC. Cell viability images correspond to nonfixed cells.
RNA isolation, quantification, retrotranscription, and quantitative polymerase chain reaction
Total RNA was extracted from fresh isolated islets (purity of 50%) as well as from 2D cultures and ICCs in sandwich cultures on days 10 and 18, using the RNeasy Plus Mini kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. Groups of 10 and 15 inserts from day 10 to 18, respectively, were pooled. Once purified, RNA quantity and quality were assessed with the Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA). The RNA integrity (RIN) score ranged from 6.6 to 9.3. Complementary DNA (cDNA) was synthesized using the High-Capacity cDNA RT Kit (Applied Biosystems, Foster City, CA), followed by RNaseH treatment (Invitrogen). Quantitative polymerase chain reaction (PCR) was carried out in triplicates using the TaqMan Universal PCR Master Mix in 7900 Real-Time PCR System (Applied Biosystems). Primers used are listed in Table 2. Analysis of relative gene expression was calculated using the 2−ΔΔCt method, using human TATA box-binding protein (TBP) and human large ribosomal protein (RPLP0) as endogenous controls. Relative gene expression was calculated using ICCs maintained in nonadherent Petri dishes as controls. Cycle number of 40 was used for undetectable transcripts.
Statistical analysis
Statistical analysis was performed using SPSS 15.0 (SPSS, Chicago, IL) and GraphPad Prism 5.0 (GraphPad, La Jolla, CA). Data are expressed as mean ± standard error of the mean, and differences among means were analyzed using the one-way analysis of variance (ANOVA) and Tukey's test for post hoc analysis. Data not normally distributed are expressed as median with interquartile range and analyzed using Kruskal–Wallis one-way ANOVA and post hoc Dunn's test. A p value of <0.05 was considered significant.
Results
Design and characterization of the functionalized RAD16-I peptide
To assess whether the presence of the short RGD peptide motif interfered in the nanofiber self-assembling process, we performed a structural and physicochemical characterization of the SAPs based on Congo red staining, CD, and the AFM. The RGD construct was positive for Congo red staining, which binds to the β-sheet configuration present in the nanofibers 73 (Fig. 2A), indicating that the addition of the RGD functional motif to RAD16-I did not hinder the self-assembling process, allowing gelation of the constructs.
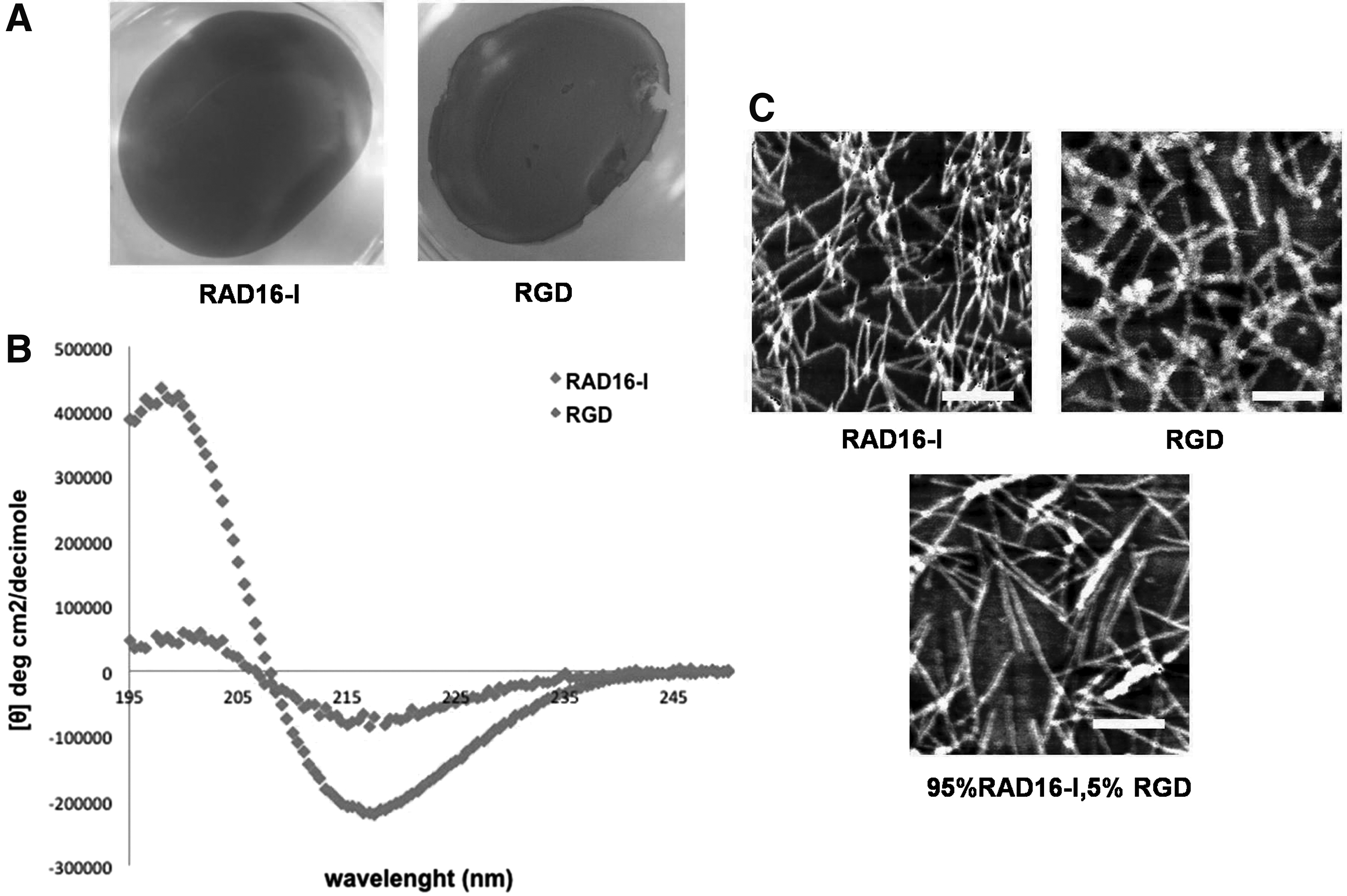
Structural and physicochemical characterization of RAD16-I and RAD16-I/RGD peptides.
CD spectroscopy was used to evaluate the influence of RGD peptide on the β-sheet secondary structure of RAD16-I. The far-ultraviolet CD spectrum of proteins and peptides gives information about their secondary structure in solution. 74 A negative band at 216–218 nm and a positive band at 198 nm characterize β-sheet structure profiles. 75 The CD spectra of RAD16-I showed the typical profile of a β-sheet structure, with minimum molar ellipticity (deg cm2/decimole) around 216 nm and maximum at 195 nm (Fig. 2B). The CD spectra of RGD peptide showed a similar spectrum as the nonmodified RAD16-I, but with a diminished β-sheet content due to the introduction of the two RGD groups. This is depicted as a decrease in the intensity of molar ellipticity at 216 and 198 nm (Fig. 2B).
Noncontact AFM studies were performed to study the formation of nanofibers and whether the insertion of RGD motifs modified the morphology of the scaffold nanostructure. Both peptides, pure RAD16-I and RGD, formed nanofibers (Fig. 2C). The analysis of a blend of 95:5 RAD16-I/RGD, the ratio used in sandwich cultures, showed that nanofibers were also present. The diameters of RAD16-I nanofibers, pure RGD, and RAD16-I/RGD blend were 19.58 ± 0.79 nm, 38.78 ± 2.04 nm, and 27.85 ± 1.48 nm, respectively (p < 0.001, RGD and RAD16-I/RGD compared with pure RAD16-I).
Changes in cell characteristics along culture
The purity of islet cell preparations used in the studies ranged from 40% to 80% (Table 1). Magnetic cell sorting resulted in a significant enrichment in insulin+ cells in the PSA-NCAM-positive fraction (presorting: 27.4% ± 4.88%, postsorting: 50.1% ± 15.1%, n = 7 p = 0.02). The presence of amylase+, cytokeratin 19+, and vimentin+ cells was significantly reduced in the postsorting fraction (amylase: 16.5% ± 1.92% vs. 6.22% ± 4.03%, p = 0.02; cytokeratin: 25.3% ± 3.76%, vs. 9.42% ± 3.30%, p = 0.01; and vimentin: 20.3% ± 2.23%, vs. 10.5% ± 7.66%, p = 0.02). Once in monolayer culture, the percentage of cells expressing insulin declined rapidly: it was 9.2% ± 1.8% at the end of passage 1 and became undetectable beyond passage 4 (Fig. 3A). In contrast, the expression of the mesenchymal marker vimentin increased progressively (day 1: 10.5% ± 3.9%, passage 4: 98.0% ± 0.5%). Expanded cells were allowed to reaggregate through the formation of ICCs and were maintained in Petri dish cultures for 18 days to be used as controls (Fig. 3B). On day 1 of sandwich culture, ICCs had a compact and rounded shape with some indication of cell spreading (Fig. 3C). On day 10, ICC morphology had changed dramatically, with clear evidence of cell spreading and increased size. By the end of the culture, on day 18, ICCs conformation was maintained, while cells in the outer part of the clusters had adopted an elongated shape. No apparent morphological differences were identified between ICCs cultured in pure RAD16-I and in RAD16-I/RGD-functionalized matrices.
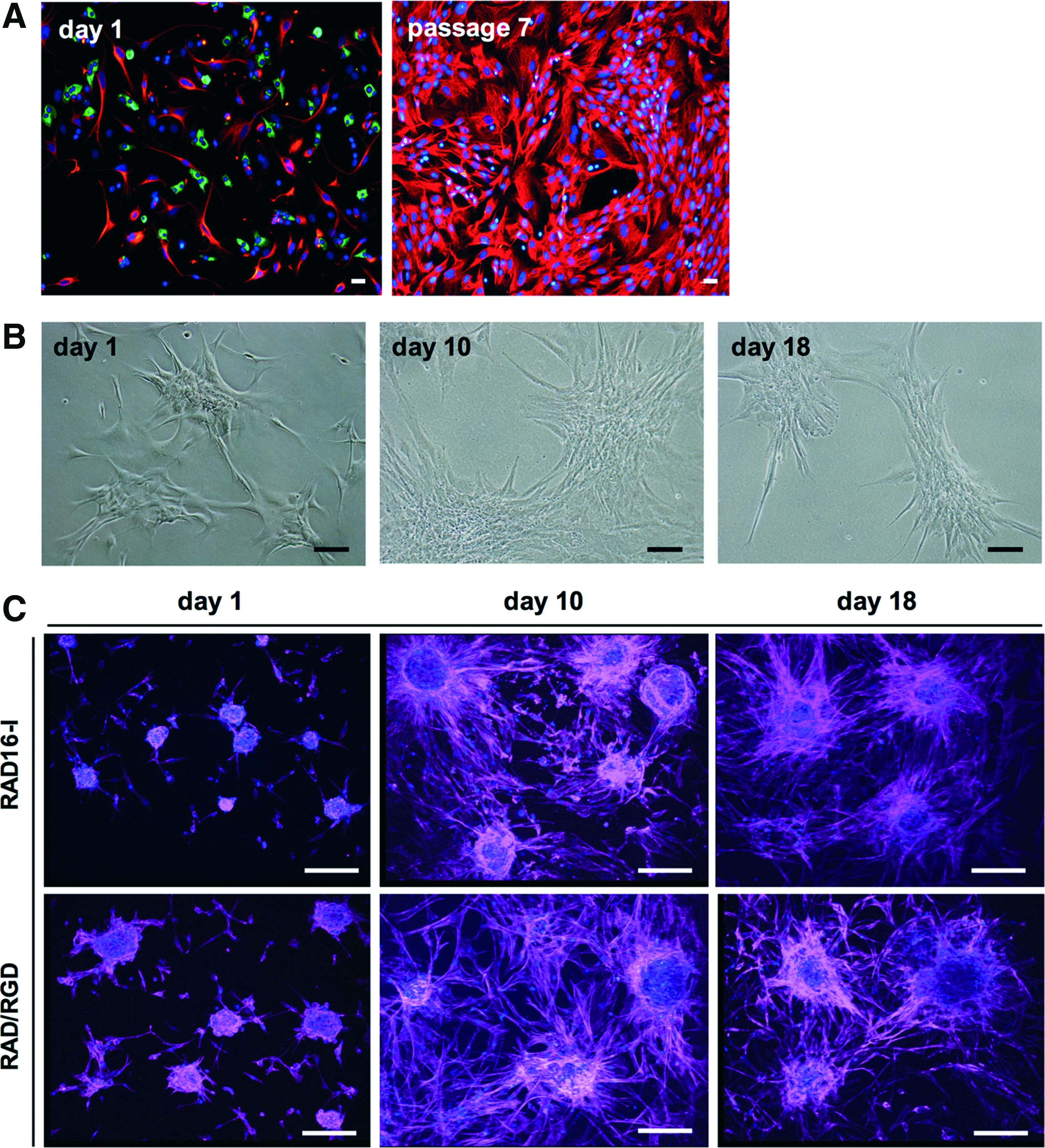
Cell characteristics along culture.
Expression of integrins
The gene expression of integrin subunits, αV, β1, and β3, was determined initially and at the end of the expansion in monolayer culture (passage 1 and passages 7–8, respectively) and in ICCs after 10 days in culture in Petri dishes and RAD16-I and RAD16-I/RGD sandwich cultures. Integrin expression was identified at all time points, supporting the rationale for the use of functionalized matrices in the sandwich cultures. No significant changes were found in the expression of integrins in any of the tested conditions (Fig. 4).
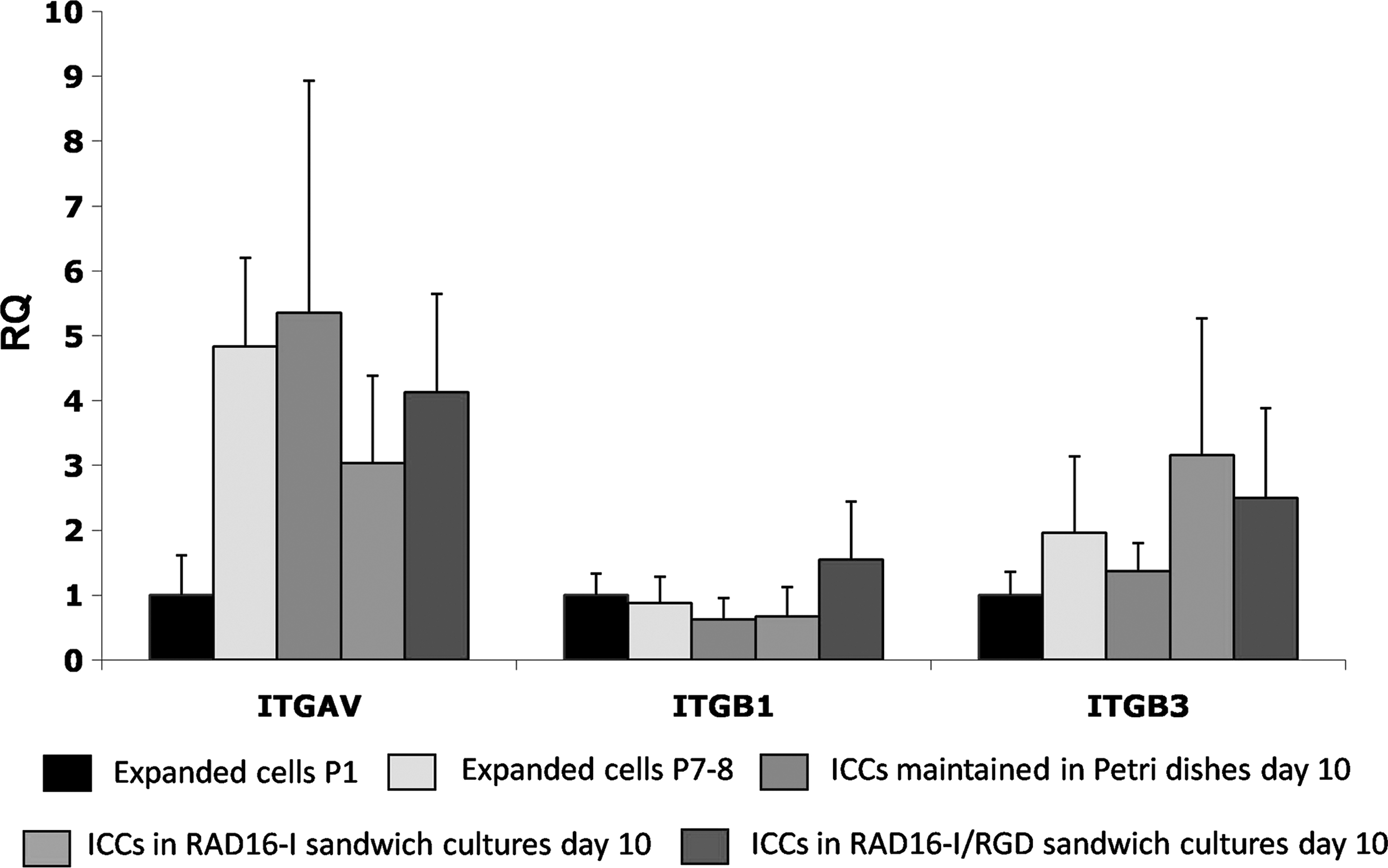
Gene expression of αV (ITGAV), β1 (ITGB1), and β3 (ITGB3) integrin subunits in expanded cells at passage 1, passages 7–8, and ICCs in nonadherent Petri dishes and cultured both in RAD16-I and RAD16-I/RGD sandwich scaffolds for 10 days. Data are expressed as RQ and are mean ± SEM of four experiments with cells from four different donors, except RGD condition (n = 3). RQ, relative quantification; SEM, standard error of the mean.
Behavior of ICCs in RAD16-I and RAD16-I/RGD sandwich cultures
The number of ICCs per insert was counted by visual inspection on days 1, 10, and 18 of culture. After removal of the unbound cells on day 1, a significantly larger number of ICCs adhered to RAD16-I/RGD matrix than to nonmodified RAD16-I (RAD16-I/RGD: 198 ± 25.0 ICCs/insert; RAD16-I: 123 ± 17.8 ICCs/insert, p < 0.001) (Fig. 5A), and the difference was similar throughout the culture. The total number of ICCs decreased progressively in both types of matrices (Fig. 5A), but ICCs size increased significantly from day 1 to 10 and it continued to increase from day 10 to 18, even though the difference did not reach statistical significance (Table 3). The size of the individual ICCs is shown in Figure 5B. The parallel reduction in the number of ICCs and the increment in size suggest that small clusters aggregated into bigger ICCs, as observed by visualization of whole sandwich constructs under the stereoscopic microscope (Fig. 5C).
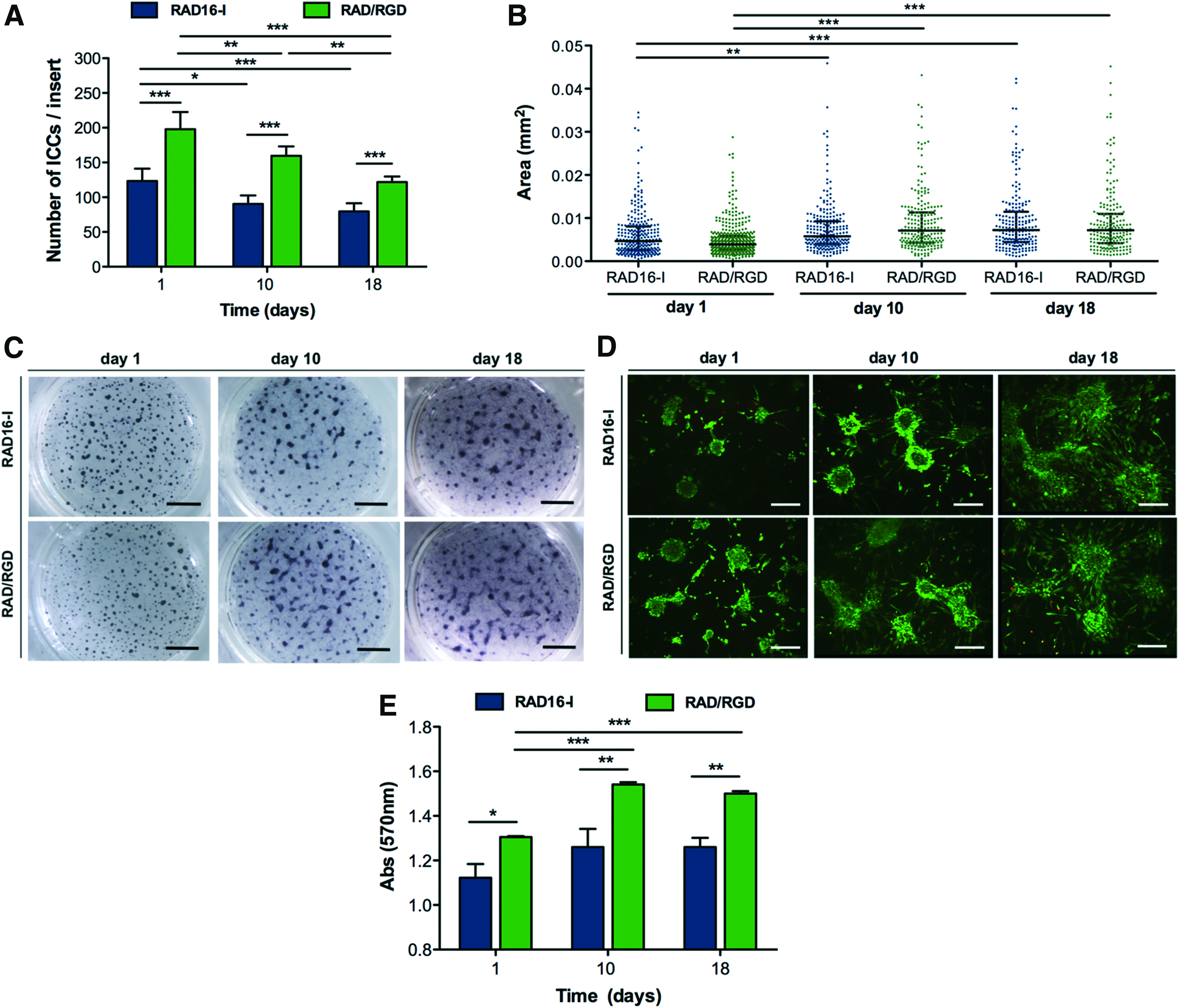
Size and morphology of ICCs cultured in RAD16-I and RAD16-I/RGD sandwich cultures for 18 days.
Values are median and range.
Cell viability showed that on day 1 of sandwich culture, a large majority of cells (98%) within each cluster were alive, both in RAD16-I and in RAD16-I/RGD scaffolds (Fig. 5D). On days 10 and 18, the percentage of viable cells was 85%, slightly decreased compared with day 1 (p: not significant [NS]). Remarkably, the majority of cells located in the center of clusters remained alive for the whole culture period. No differences in viability were observed regarding ICC size. Viability of ICCs maintained in Petri dish cultures was not studied since the main motivation was to assess the influence of ICC entrapment in RAD16-I or RAD16-I/RGD matrix. Before self-assembling, the pH of RAD16-I hydrogel is around 3, which may be harmful to cells. Moreover, when the construct's thickness is higher than 300 μm, diffusion problems may arise, 76 having an impact on ICC viability. The thickness of our construct was 500 μm, therefore viability of ICCs during the culture period could have been affected due to poor oxygenation and nutrition.
On day 1 of sandwich culture, MTT assay showed significantly higher absorbance values in RAD16-I/RGD-functionalized cultures than in pure RAD16-I, probably reflecting the higher cellular adhesion to the RAD16-I/RGD scaffold, and the difference between the two matrices was maintained throughout the culture period (Fig. 5E). In addition, ICCs cultured in RGD-based matrix showed higher MTT values on days 10 and 18 than on day 1. In ICCs cultured in pure RAD16-I scaffold, the MTT values tended also to increase on the first 10 days of culture, but the difference did not reach statistical significance. No significant changes in MTT values were observed in ICCs cultured in RAD16-I nor in RAD16-I/RGD matrices between culture days 10 and 18. Considering that cell viability was slightly lower on day 10 than on day 1, the increased values of the MTT assay suggest that the total number of cells in the ICCs was higher on days 10 and 18, probably due to cell proliferation.
Expression of beta cell markers in ICCs cultured with SAPs
Gene expression of beta cell-specific markers was markedly reduced and in the limits of detection in expanded cells on passages 7–8 and in newly formed ICCs (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea). After 10 days in RGD-based sandwich cultures, the expression of the specific beta cell marker Ins and the beta cell transcription factors Pdx-1, Nkx6.1, and MafA was significantly increased compared with both control condition (ICCs cultured in Petri dishes) and RAD16-I scaffold (Fig. 6A). On day 18, similar results were obtained, and Ins, Nkx6.1, and MafA gene expression levels were increased compared with control ICCs cultured in nonadherent conditions (Fig. 6B). In addition, the transcription factors, NeuroD1 and Sox9, showed a tendency to increase (p = 0.06 and p = 0.07, respectively). The expression of epithelial and mesenchymal markers, E-cadherin and N-cadherin, respectively, as well as vimentin, the hallmark of the mesenchymal phenotype, was similar in all groups, confirming the specificity of the increased expression of beta cell markers in cells cultured in the RAD16-I/RGD sandwich system. The persistency of vimentin expression in 3D cultures indicates that ICCs still maintain a mesenchymal phenotype. However, the steady expression of vimentin in 3D cultures seems to be balanced by the increased expression of pancreatic beta cell markers.

Gene expression analysis of ICCs cultured in RAD16-I and RAD16-I/RGD sandwich cultures for
Discussion
The scarcity of human islets available for transplantation severely limits the clinical use of cell therapy in the treatment of diabetes. In vitro expansion of human beta cells is a promising approach to provide an almost unlimited source of insulin-expressing cells that could be used in beta cell replacement therapy.11,14 However, human beta cells have a low replicative capacity in vivo and in most in vitro conditions. Although beta cells may be induced to expand massively when islet cells are cultured in monolayer conditions, they lose the beta cell phenotype and dedifferentiate into a population of mesenchymal-like cells that do not secrete insulin.11,12,16 The disruption of cell–cell and matrix–cell interactions is considered a crucial contributor to the dedifferentiation of endocrine islet cells. When pancreatic islets are isolated, the majority of cell–cell and cell–matrix interactions are disrupted, and disruption is more severe if islets are subsequently dispersed into single cells.16,18 We have induced the reaggregation of islet-derived cells into clusters (ICCs) and used a 3D culture model with functionalized SAPs to stimulate redifferentiation of these expanded cells. The 3D culture model was based on the SAP hydrogel RAD16-I in a sandwich configuration and functionalized with the integrin-binding motif RGD (RAD16-I/RGD). Previous studies have demonstrated the ability of RAD16-I SAP to maintain islet function in vitro and after transplantation in induced diabetic mice. 64
After expansion in the monolayer, the dedifferentiated islet-derived cells were induced to form cell clusters (ICCs) to mimic the in vivo configuration of pancreatic islets characterized by close cell contacts and cell–cell and cell–matrix interactions.20,38 Then, the ICCs were embedded between two thin layers (sandwich configuration) of RAD16-I peptide or RAD16-I functionalized with the RGD integrin-binding motif, a well-studied adhesive peptide used to recreate the BM conditions. ICCs were also kept in culture in nonadherent conditions (control).
We first tested the characteristics and suitability of the SAP. The addition of the two RGD motifs decreased the content of the β-sheet structure present in RAD16-I, but the capacity to form nanofibers was maintained, as demonstrated by AFM studies, which also showed that pure RGD formed thicker nanofibers than RAD16-I. The influence of RGD peptide on the structural organization of RAD16-I could be observed when RAD16-I and RGD peptides were blended at the same ratio used in sandwich cultures (95:5, respectively). Specifically, RAD16-I/RGD matrix provided a network of thicker nanofibers than RAD16-I alone, showing that the RAD16-I/RGD scaffold was structurally different from the conventional RAD16-I scaffold. Since it is well known that cells sense the biological and structural traits of the ECM through integrin receptors, the chemical and structural modifications induced by the RGD motif could cause the change in ICC behavior, including cell adhesion and gene expression profiles that we identified when the RAD16-I/RGD scaffold was used. However, the integrin gene expression was not expected to change under the RAD16-I condition since it is a noninstructive SAP. The introduction of the RGD signaling motif did not cause any change in the expression of integrins either. Lam and Segura also showed that αV and β3 integrin expression levels were not affected by introduction of the RGD bioactive motif in hydrogel-based cultures. 77 One possible explanation is that cells were expressing enough integrin receptors to promote the proper response during redifferentiation.
Our findings indicate that RAD16-I sandwich cultures are dynamic cultures in which ICCs were able to connect to one another as the culture progressed. Preservation of the cell cluster conformation implies that cell–cell contacts within each cluster were maintained over the culture period. This trait seems to be critical for the redifferentiation process. 14 Cell viability was also preserved along the culture period, both in pure RAD16-I and in RGD-modified RAD16-I. In contrast, ICCs number and area changed from day 1 to the end of the culture on day 18. The progressive reduction in the number of ICCs in the absence of a reduced viability and the increment in ICC area suggest the presence of a self-organizing process where initial small ICCs clumped together to give rise to larger aggregates. Proliferation could also be involved in the increase of ICC area during the first period of culture (days 1–10), as indicated by higher absorbance values at day 10.
The addition of the RGD motif to RAD16-I increased the adhesion of ICCs to the matrix, as shown by the higher number of clusters attached to the RGD-functionalized matrix compared with adhesion to pure RAD16-I surface. The higher MTT values on day 1 of culture in the RAD16-I/RGD scaffold probably reflect this increased adhesion. Furthermore, MTT values in ICCs cultured in RAD16-I/RGD, but not in RAD16-I, matrix increased on days 10 and 18 compared with day 1. These results suggest that the number of cells in the ICCs increased overtime and that RGD addition to RAD16-I matrix promoted cell proliferation. Thus, the results suggest that cells specifically recognize and respond to the RGD peptide sequence present in the modified RAD16-I matrix, creating a more favorable microenvironment that improves cell attachment conditions.
The gene expression results show that addition of the matrix-derived RGD sequence to the noninstructive RAD16-I induced a change in phenotype involving the overexpression of several beta cell-specific genes, indicating the induction of some degree of redifferentiation toward a beta cell phenotype. Specifically, the Ins gene expression, as well as that of the beta cell transcription factors Pdx1, Nkx6.1, and MafA, was upregulated on day 10 in ICCs cultured in the RAD16-I/RGD system. Similar results found on day 18 of culture provide additional support to the presence of this redifferentiation process. On the other hand, the absence of statistically significant differences between days 10 and 18 suggests that a shorter, 10 days, protocol may be just as good if not better than prolonging the incubation by 8 further days. Gene expression was similar in ICCs maintained in nonadherent culture conditions and ICCs cultured in RAD16-I, indicating that nanofibers were not sufficient to induce redifferentiation. Addition of the integrin-binding motif RGD to the RAD16-I scaffold was required to induce redifferentiation of expanded cells. The expression of nonendocrine markers such as vimentin, E-cadherin, or N-cadherin did not change, confirming specificity of the increment in beta cell markers. However, a reduction in expression of mesenchymal markers could have been expected to occur in parallel with the redifferentiation process. It is worth noting that to better analyze the effect of the structural and biomechanical microenvironment provided by the SAP on the redifferentiation process, the culture medium used in this study was very simple, and specifically no differentiation factors were added to it. Therefore, we suggest that the addition of new signaling motifs, in combination with a redifferentiation cocktail, would enhance the redifferentiation process.
Conclusions
We have studied whether the reestablishment of cell–cell and cell–matrix relationships with a biomimetic synthetic scaffold could induce the redifferentiation of pancreatic human beta cells dedifferentiated after in vitro expansion. The expanded cells were allowed to reaggregate into ICCs and then embedded between two thin layers of the noninstructive SAP RAD16-I or RAD16-I functionalized by the covalent addition of the integrin-binding motif RGD (Arg-Gly-Asp) (RAD16-I/RGD). The addition of the RGD sequence to the matrix structure was expected to promote cell-ECM interactions that could stimulate the upregulation of beta cell markers and the redifferentiation toward a beta cell-like phenotype. ICCs cultured with RAD16-I were viable, maintained their cluster conformation, and increased in size by aggregation of small ICCs, suggesting a self-organizing process. ICCs cultured in functionalized RAD16-I/RGD showed enhanced cell adhesion to the RAD16-I matrix and the reexpression of the beta-specific genes, Ins, Pdx1, Nkx6.1, and MafA. Therefore, ICCs have the capacity to respond to the microenvironment stimuli, modulating their behavior and gene expression levels. The results indicate that RGD-functionalized SAP in sandwich conformation is a promising 3D platform to induce redifferentiation toward a beta cell phenotype and to generate insulin-expressing cells that could be used in cell therapy of diabetes. To achieve complete redifferentiation toward a mature beta cell phenotype, different combinations of other specific beta cell integrin-binding motifs could be developed to create a biologically functional scaffold more similar to the beta cell microenvironment.
Footnotes
Acknowledgments
This research project was supported by a Fundació La Marató TV3 grant PR084/12 (E.M. and C.E.S.); by a Carlos III Health Institute (ISCIII) grant PI13/00108; ERDF “A way to build Europe” (E.M.); and by ACCIÓ (ADVANCECAT), an initiative of the Program FEDER Catalunya 2014–2020. The authors are also grateful to the Materials Science Laboratory of IQS School of Engineering for helping with AFM studies.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.