
Abstract
The design paradigm of biomaterials has been changed to ones with favorable immunomodulatory effects, indicating the importance of accurately evaluating the immunomodulatory properties of biomaterials. Among all the immune cells macrophages receive most attention, due to their plasticity and multiple roles in the materials and host interactions, and thereby become model immune cells for the evaluation of immunomodulatory properties of biomaterials in many studies. Lipopolysaccharides (LPS), a polysaccharide in the outer membrane of Gram-negative bacteria, elicit strong immune responses, which was often applied to activate macrophages, resulting in a proinflammatory M1 phenotype, and the release of proinflammatory cytokines, including tumor necrosis factor alpha (TNFα), interleukin (IL)-1, and IL-6. However, there is no consensus on how to apply macrophages and LPS to detect the immunomodulatory properties of biomaterials. The lack of scientific consideration of this issue has led to some inaccurate and insufficient conclusions on the immunomodulatory properties of biomaterials, and inconsistences between different research groups. In this study, we carried out a systemic study to investigate the stimulatory effects of LPS with different times, doses, and conditions on the activation of macrophages. An experimental pathway was proposed accordingly for the activation of macrophages using LPS for assessing the immunomodulatory property of biomaterials.
Introduction
B
Among all the immune cells, macrophages received most attention due to its plasticity and diverse roles in the biomaterials and host body interactions.10,11 Macrophages are highly plastic and can change their phenotype and physiology for adapting in response to the environmental signals alteration, 12 which makes them a primer target for biomaterials-mediated immunomodulation. Lipopolysaccharides (LPS), also known as lipoglycans and endotoxin, are large molecules consisting of a lipid and a polysaccharide composed of O-antigen, outer core and inner core joined by a covalent bond; they are found in the outer membrane of Gram-negative bacteria, and elicit strong immune responses in animals. It is well known that LPS can switch macrophages into proinflammatory M1 phenotypes, resulting in the releasing of proinflammatory cytokines, including tumor necrosis factor alpha (TNFα), interleukin (IL)-1, and IL-6.13–16 Due to its strong proinflammatory effects, LPS has been generally used to activate macrophages for the applications of inflammation-related studies.17,18
To investigate the modulatory effects of biomaterials on macrophages, LPS is usually applied to activate the macrophage into a proinflammatory phenotype, which are then treated with candidate biomaterials to determine the anti-inflammatory capacity of biomaterials. One major issue underlying this approach is how to activate macrophages using LPS. After reviewing related studies (Table 1), we found out there are three major streams in applying LPS: (1) LPS activates macrophages together with biomaterials for the whole stimulation time19–22 ; (2) LPS is first applied to activate macrophages then removed and replaced by biomaterials23,24; (3) biomaterials are applied to activate macrophages without the involvement of LPS.25–27 However, there is no consensus on the indications of applying each specific stream. The LPS stimulation condition was designed without strong scientific rationales, and the activation time and dose in each stream were picked up randomly, which may lead to some inaccurate and insufficient conclusions on the immunomodulatory properties of biomaterials, and inconsistences between different research groups.
LPS, lipopolysaccharides.
Therefore, it is of great significance to do a systemic research on the interactions between LPS and macrophages, to provide basic knowledge for applying LPS to assess the immunomodulatory property of biomaterials. Based on the literature review, the doses of LPS range from as little as 10 ng/mL 28 to 1000 ng/mL or even more, 29 while the stimulation time varies greatly too, ranging from as little as 10 min 30 to 24 h or more.25,31 This follows a research question that what dose and time should we choose for each stream to achieve the research purposes. In this study, we first investigated the effects of stimulation time and doses on the LPS-mediated activation of macrophages, to serve for the applications of the first stream. As to the second stream, we activated macrophages using LPS with different doses and stimulation times. The LPS supplemented media were then removed and replaced by normal culture medium, which were subsequently cultured for 6 or 12 h before the inflammatory response was determined. Lastly, we tested the immunomodulatory property of a nano-scale material, mesoporous silica nanospheres (MSNs), using all the three streams to demonstrate the importance of choosing a suitable stream, dose, and time. It is the ambitions of the authors that this study can provide an experimental pathway for the activation of macrophages using LPS to evaluate the immunomodulatory properties of biomaterials, so that researchers can select a proper dose, time, and stimulation condition of LPS for the relevant studies.
Materials and Methods
Cell cultures
The inflammatory response of RAW 264.7 cells (RAW cell), a macrophage cell line, was applied to detect the proinflammatory capacities of LPS under different conditions, time, and doses. RAW cells culture medium (complete medium) was composed of 5% heat inactivated fetal bovine serum (FBS; Thermo Scientific), 1% (v/v) penicillin/streptomycin (Life Technologies), and Dulbecco's modified Eagle's medium (DMEM; Life Technologies). The cells were cultured in complete medium under a humidified incubator with 5% CO2 and constant 37°C. The cells were gently scraped off and passaged at around 90% confluence and expanded for two passages before applying for the subsequent experiments.
Inflammatory response of macrophages under LPS stimulation with different dose, time, and condition
The mRNA expression of the proinflammatory cytokine genes IL-1β, IL-6, and TNFα was assayed by RT-qPCR. RAW Cells were plated in 24-well plates at a density of 1.5 × 105 cells per well. After reaching 80% confluence, the medium was removed and replaced by medium containing different doses of LPS (0, 10, 50, 100, 250, 500, and 1000 ng/mL), with different stimulation time (10 min, 30 min, 2, 6, 12, and 24 h). The supernatants were collected for subsequent ELISA essay. As to the removal condition experiments, RAW cells were first stimulated by LPS with different concentrations (0, 10, 50, 100, 250, 500, and 1000 ng/mL) for 30 min, 2 or 6 h. The medium containing LPS was then removed and the cells were washed three times by fresh culture medium and kept cultured in normal culture medium for another 6 or 12 h before the total RNA was extracted and the supernatants were collected for ELISA assay.
Total RNA extraction was done by using TRIzol reagent (Life Technologies) and the mRNA expression was analyzed by RT-qPCR. Complementary DNA was synthesized from 500 ng total RNA using the DyNAmo™ cDNA Synthesis Kit (Finnzymes, Thermo Scientific). Reverse transcription quantitative real-time PCR primers applied in this study were listed in Table 2. SYBR Green qPCR Master Mix (Life Technologies) was used for the qPCR assay in an ABI Prism 7500 Thermal Cycler PCR machine (Applied Biosystems). The relative expression was obtained by normalizing the average cycle threshold (Ct) value of each target gene with the Ct value of β-ACTIN (housekeeping gene).
For the ELISA assay, proinflammatory cytokines IL-1β, IL-6, and TNFα were quantified using mouse ELISA kits (R&D Systems) following the manufacturer's instruction. The IL-1β, IL-6, and TNFα concentration were calculated by correlation with a standard curve and the results were presented as the amount (pg) of IL-1β, IL-6, and TNFα per mL of supernatant.
The immunomodulatory property of MSNs under different LPS stimulation conditions, time, and doses
Preparation of MSNs
In a typical synthesis procedure, 1.82 g of cetyltrimethylammonium bromide (CTAB) and 3.0 g of ammonium fluoride (NH4F) were dissolved in 500 mL of distilled water at 80 °C under vigorous stirring, and then 9 mL of tetraethoxysilane (TEOS) was added drop by drop (CTAB, NH4F, TEOS, Sinopharm Chemical Reagent Co. Ltd.). After stirring for 2 h, the solution was cooled at room temperature overnight. The as-synthesized materials were centrifuged, washed with distilled water and ethanol, and then dried under vacuum. To remove the surfactant CTAB, the solid products were calcined at 600°C in air for 6 h at a heating rate of 1°C/min. The morphology and microstructure characteristics of MSNs were tested by transmission electron microscopy (JEM-2100F; JEOL).
Inflammatory response of macrophages stimulated by MSNs under different LPS stimulation conditions
RAW Cells were plated in 24-well plates at a density of 1.5 × 105 cells per well. According to different stimulation streams, three treatment groups were designed. In the first group, macrophages were treated with 200 μg/mL MSNs and 10 μg/mL LPS for 6 h. In the second group, RAW cells were first treated with 10 μg/mL LPS for 2 h and then LPS was removed and washed with PBS three times. Afterward, 200 μg/mL MSNs were further added to stimulate the cells for 6 h. In the last group, RAW cells were stimulated by 200 μg/mL MSNs only for 6 h. Then, total RNA was extracted and relative expression of proinflammation gene cytokines genes IL-1β, IL-6, and TNFα was assayed by RT-qPCR as described above.
Statistics
All data are presented as the mean ± SD or standard error (SE) of the mean, and n corresponds to the number of experiments. Statistical differences between groups were evaluated using paired Student's t-tests or one-way analysis of variance (ANOVA) followed by a Student's t-test with Bonferroni correction as indicated. Statistical analyses were performed using the GraphPad Prism 5.0 software (version 5.01 for Windows, GraphPad Software, Inc.). Differences were considered significant when p < 0.05. Statistical significances are between different tests in the same group. Data are represented–standard error of mean.
Results and Discussions
Constant LPS stimulation
Constant LPS activation represents a strong inflammatory in vivo environment that is usually led by the microorganism infections in situ. In this section, we have investigated how the LPS with different doses and stimulation time affected the inflammatory response of macrophages, aiming at determining the best stimulation dose and time for the material evaluation requiring constant LPS stimulation. The doses of LPS that we chose were 10, 50, 100, 250, 500, and 1000 ng/mL, which could almost cover the dose range applied in previous researches. Among these six doses, 10, 100, and 1000 ng/mL LPS appear to be more commonly applied in other studies.32,33
When the stimulation time was 10 min, LPS generally could not significantly enhance the gene expression of proinflammatory cytokines (IL-1β, IL-6, and TNFα), even when the concentration went up to 1000 ng/mL, indicating that 10 min was insufficient for the activation of macrophages (Fig. 1). Different expression pattern was observed when the stimulation time was increased to 30 min. All three proinflammatory genes expression was significantly enhanced by LPS with different concentrations (10–1000ng/mL). TNFα had the highest fold changes (~5), indicating that it had a more rapid response toward LPS compared with that of IL-1β and IL-6. IL-6 had a latest response toward LPS, with only slight increase in fold changes (~1.5) observed. The dose-dependent effect was not apparent at this time point, although LPS with high concentration (1000 ng/mL) showed a slightly higher fold change (Fig. 1).
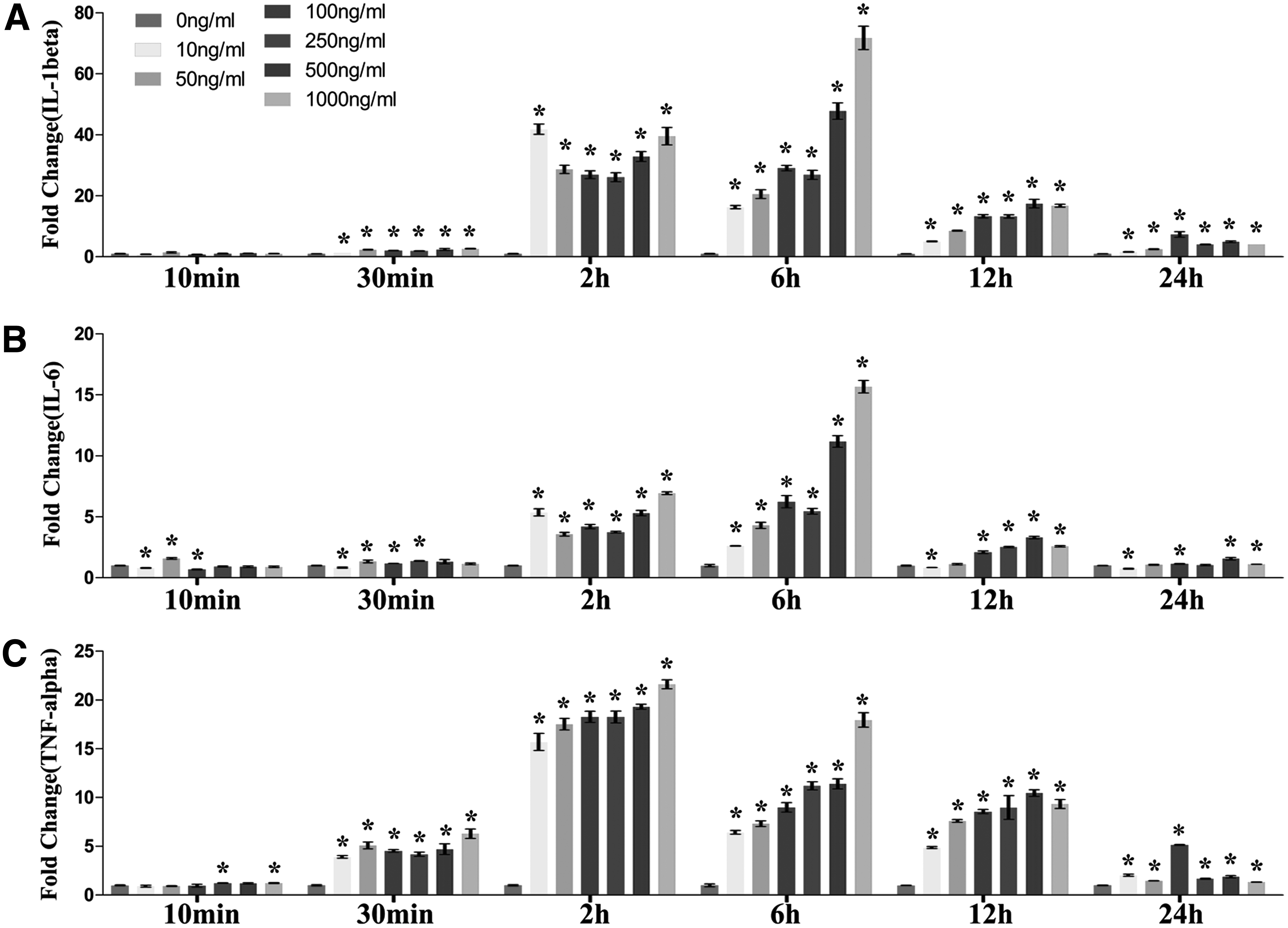
The effects of dose and time effects of LPS on activating macrophages. Macrophages (Raw cells) were treated by different concentrations of LPS (10, 50, 100, 250, 500, and 1000 ng/mL) for different stimulation time (10 min, 30 min, 2, 6, 12, and 24 h). The internal controls were macrophages under the same treatment of media without LPS (10 min, 30 min, 2, 6, 12, and 24 h after removal of culture medium). Relative mRNA expression of proinflammation cytokines IL-1β
With 2 h of LPS treatment (10–1000 ng/mL), IL-1β, IL-6, and TNFα gene expression has all dramatically upregulated, with fold changes of ~40, ~6, and ~20 respectively. It started to show a roughly dose-dependent manner. LPS with the concentration of 1000 ng/mL had the most significant stimulatory effect, within the range of 50–1000 ng/mL. However, LPS with the concentration of 10 ng/mL appeared to enhance IL-1β and IL-6 most significantly, which indicated a different response pattern in the low dose range (<10 ng/mL) (Fig. 1).
The dose-dependent manner was most obvious with the treatment time of 6 h. LPS with the concentration of 1000ng/mL had the most significant stimulatory effect, with the greatest upregulation of all three proinflammatory cytokine genes (IL-1β, IL-6, and TNFα). However, when the stimulation time was increased to 12 h, LPS with the concentration of 500 ng/mL showed the most significant stimulatory effect, and the one with 1000 ng/mL dropped to the second. Interestingly, LPS with the concentration of 100 ng/mL elicited most effective inflammatory effect as the stimulation time increased to 24 h, and the stimulatory effect of LPS (>100 ng/mL) on the IL-6, and TNFα gene expression by macrophages almost disappeared compared with the negative control (Fig. 1).
As to the time-dependent effects, when treated with LPS with the concentration of 10 ng/mL, the peak expression of all three proinflammatory genes was at 2 h. TNFα showed an early response and its gene expression started to go up significantly at 30 min, while IL-6 and IL-1β were delayed to 2 h. As to the expression pattern for 50 ng/mL, the peak expression of TNFα genes was still at 2 h. However, the peek expression of IL-1β was at 2 and 6 h, while that of IL-6 was delayed to 6 h. As the concentration went up to more than 100 ng/mL (100, 250, 500, and 1000 ng/mL), the peak expression of TNFα genes remained at 2 h, while the peek expression of IL-1β and IL-6 was at 6 h (Fig. 1).
The cytokine release profile showed a different pattern compared with gene expression profile. This may be due to the delayed protein expression compared with the gene expression, and ELISA shows the cumulative release of a specific cytokine for the period of observation time while RT-qPCR demonstrates the real-time gene expression in nuclei. For IL-1β, the dose-dependent manner started to be observed from 6 h of stimulation time, the 1000 ng/mL group had the highest release. Interestingly, the dose-dependent manner could still be observed at 24 h, which was different from the gene expression with the peak expression at 100 ng/mL under same treatment. As to TNFα and IL-6, no dose-dependent manner was observed throughout the period, it appeared LPS with 10 ng/mL had similar effects on enhancing the release of TNFα and IL-6 in comparison to one with 1000 ng/mL (Fig. 2).
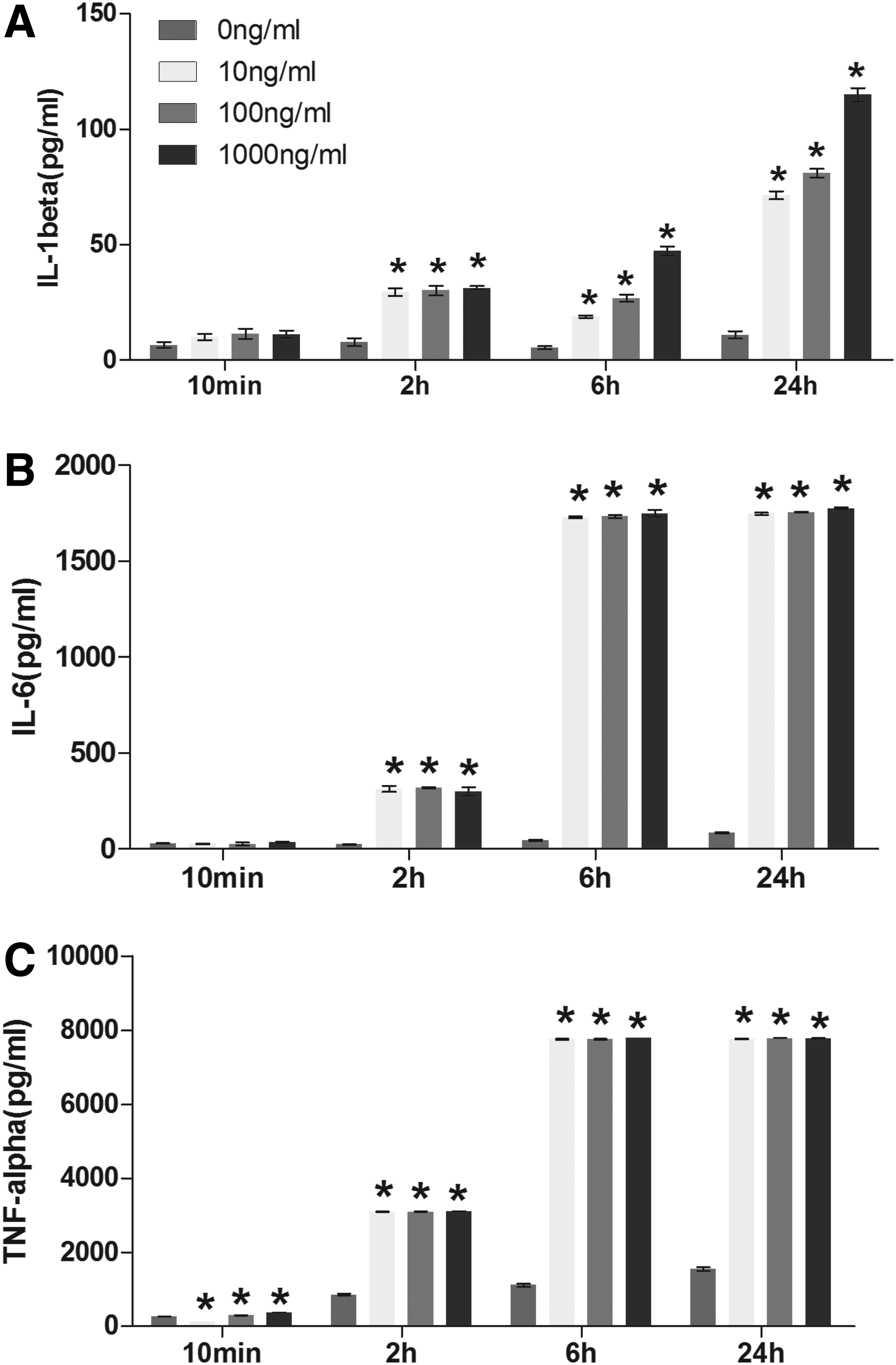
Release of proinflammatory cytokines from macrophages activated by LPS with different doses and stimulation time. Macrophages (Raw cells) were treated by different concentrations of LPS (10, 100, and 1000 ng/mL) for different time (10 min, 2, 6, and 24h).Then, the supernatants were collected and proinflammation cytokines IL-1β
As a summary, macrophages respond rapidly toward the stimulation of LPS. Gene expression changes can be observed as early as 30 min under proper LPS concentrations. However, dose-dependent manner shows up at 6 h, indicating that macrophages take around 6 h to distinguish the intensity of LPS. Afterward, the negative self-regulation is activated, implied from the downregulation of gene expression of 12 and 24 h groups compared to that of 6 h group and the shift of peak gene expression toward lower concentration group (500 ng/mL at 12 h and 100 ng/mL at 24 h). Therefore, 6 h could be the best time point to detect the gene expression. Stimulation time shorter than 6 h may result in failure of distinguishing the intensity of different stimulators, while stimulation time longer than 6 h may lead to misjudgment of some strong stimulators.
Due to the delayed protein expression compared with the gene expression, the stimulation time for the investigation of cytokine release profile should be more than 6 h, and 24 h can be a better time point. Especially for IL-1β, although the dose-dependent manner came up at 6 h, the release amounts of cytokines were not as much as that of 24 h. In the aspect of stimulation dose, we recommend that the candidate material is assessed with weak, moderate, and strong inflammatory response by LPS to demonstrate the immunomodulatory capacity under inflammatory environments with different intensities. However, among the tested doses, even with the lowest dose (10 ng/mL) LPS could cause similar effects on the release of TNFα and IL-6 cytokines with the stimulation time of 24 h, and the gene expression was significantly higher than negative control at 6 h although it was the lowest compared to that of other doses. This indicates that 10 ng/mL is already a strong stimulator for the inflammatory response of macrophages. Weak and moderate inflammatory response should be induced by LPS with the concentration lower than 10 ng/mL.
Removal after LPS activation
Short-term LPS activation represents a gradually diminishing inflammatory in vivo environment that may be due to the mammalian response to the surgery trauma. In this section, we investigated the inflammatory response of macrophages after removal of LPS with different removal time (30 min, 2 and 6 h), stimulation doses (10, 50, 100, 250, 500, and 1000 ng/mL) and postremoval time (6 and 12 h), aiming at determining the best parameters for the material evaluation requiring short-term LPS stimulation.
After the removal of LPS (including all three tested removal time periods 30 min, 2 and 6 h), the gene expression fold change of IL-1β and IL-6 dropped significantly comparing postremoval 6 h groups with 12 h groups. For the postremoval 6 h groups, the gene expression of IL-1β showed a roughly dose-dependent manner when the removal time was set at 30 min and 2 h, with higher dose having a greater expression. LPS with the concentration lower than 50 ng/mL was not strong enough to enhance the gene expression of IL-1β when the removal time was 30 min, however, when the removal time was increased to 2 h, the gene expression of IL-1β could be upregulated by LPS with the concentration as low as 50 ng/mL (Fig. 3A). LPS with removal time of 6 h had the most significant effect on postremoval inflammatory response of macrophages. The gene expression of IL-1β was significantly enhanced even when the stimulation concentration of LPS was only 10 ng/mL. Differently in terms of the expression of IL-6, the dose-dependent manner could be observed in all the postremoval 6 h groups, even when the removal time was set at 6 h, (Fig. 3B). LPS with the concentration of 50 ng/mL was able to enhance the gene expression of IL-6 when the removal time was 30 min. When the removal time was increased to 2 and 6 h, the gene expression of IL-1β could be upregulated by LPS with the concentration as low as 10 ng/mL (Fig. 3B
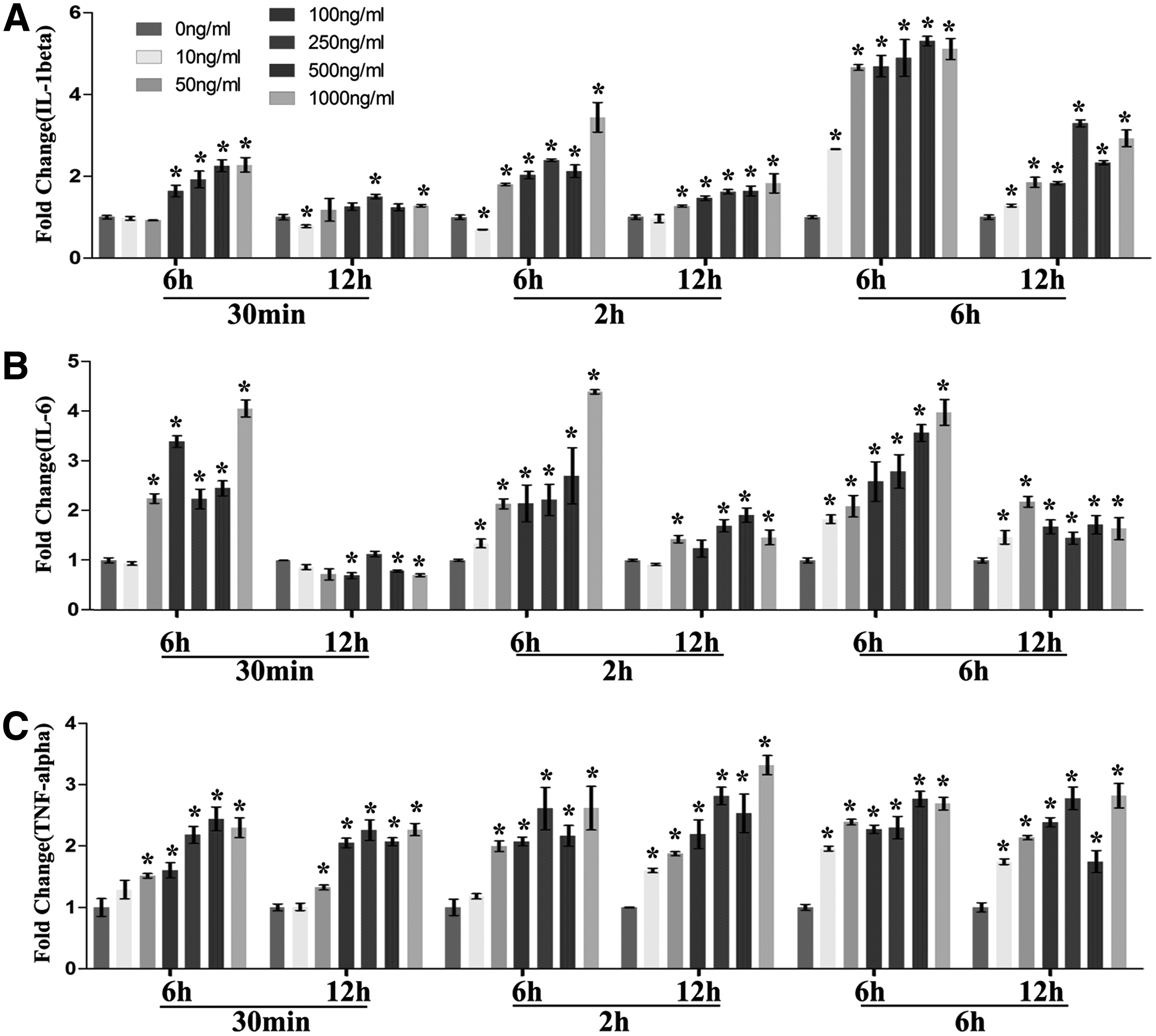
RT-qPCR results of LPS removal experiments. Raw 264.7 cells were first treated by different concentrations of LPS (10, 50, 100, 250, 500, and 1000 ng/mL) for 30 min, 2 or 6 h. Then, the medium was removed and replaced by fresh medium without LPS. RNA were extracted after 6 and 12 h. Relative mRNA expression of proinflammation cytokines genes IL-1β
The gene expression of TNFα showed a roughly dose-dependent manner when the removal time was set at 30 min and 2 h, with higher dose having a greater expression. Interestingly, LPS with the concentration of 10 ng/mL was not strong enough to enhance the gene expression of TNFα after 12 h when the removal time was 30 min, however, when the removal time was increased to 2 h, the gene expression of TNFα was upregulated at 12 h. When the removal time was set at 6 h, the dose-dependent manner disappeared, and the tested range (10–1000 ng/mL) demonstrated similar effects on enhancing TNFα expression (Fig. 3C).
As a summary, inflammatory response of macrophages was active after the removal of LPS, even though the starting stimulation time was only 30 min. It should be noted that this was dose related, and the dose should be increased when the starting stimulation time was decreased. As aforementioned, we recommend that the material is assessed with weak, moderate, and strong inflammatory response to demonstrate the immunomodulatory capacity under inflammatory environments with different intensities. From this point of view, 2 h removal time should be the best option, because it demonstrated moderately high gene expression at 12 h (low expression for the 30 min removal time groups at this time point), while remained dose-dependent at 6 h (no significant dose-dependent effect observed in the 6 h removal time groups at this time point), so that the intensity of inflammation can be manipulated by using different doses. Therefore, the best protocol would be 2 h of starting stimulation, detecting gene expression at 6 h after removal and the doses of 10, 100, and 1000 ng/mL.
We then tested the cytokine release profile with the removal time of 2 h and the doses of 10, 100, and 1000 ng/mL. It showed a different pattern compared with gene expression profile. The release of IL-1β showed no significant increase at 6 h, and was significantly increased in a dose-dependent manner when the time was risen to 24 h. Differently, the release of TNFα and IL-6 was significantly increased in a dose-dependent manner at both 6 and 24 h, with the latter had higher expression. This indicates that macrophages experienced upregulated gene expression with stimulation of 2 h directly by LPS and the cells continued releasing proinflammatory cytokines even after the removal of LPS. Based on these results, the best detection time for cytokine release profile after removal should be 24 h, when all the cytokines were significantly upregulated in a dose-dependent manner (Fig. 4).

ELISA results of LPS removal experiments. Raw 264.7 cells were first treated by different concentrations of LPS (10, 50, 100, 250, 500, and 1000 ng/mL) for 30 min, 2 or 6 h. Then the medium was removed and replaced by fresh medium without LPS. Supernatants were collected at 6 and 24 h. Release of proinflammation cytokines genes IL-1β
Immunomodulatory effects of MSNs evaluated by different streams
To assay whether different streams could affect the conclusions on the immunomodulatory properties of biomaterials, we then used mesoporous silica nanoparticles (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea) to treat macrophages (RAW cells). Interestingly, MSNs demonstrated different immunomodulatory effects under different streams. When macrophages were treated with LPS (10 ng/mL) together with MSNs (200 μg/mL), the gene expression of IL-1β and IL-6 showed no significant change, while TNFα was downregulated (Fig. 5). However, when macrophages were pretreated with LPS (10 ng/mL for 2 h, then removed), the gene expression of IL-1β and IL-6 was significantly downregulated on macrophages stimulated with MSNs (200 μg/mL), while the expression of TNFα showed no significant change under the same treatment (Fig. 5). As to the third stream, the gene expression of IL-1β and IL-6 was downregulated on macrophages stimulated with MSNs (200 μg/mL, no LPS stimulation), while the expression of TNFα was upregulated under the same treatment (Fig. 5). MSNs demonstrated different immunomodulatory effects under different conditions, indicating the need for establishing an experimental pathway to accurately evaluate the immunomodulatory properties of biomaterials.
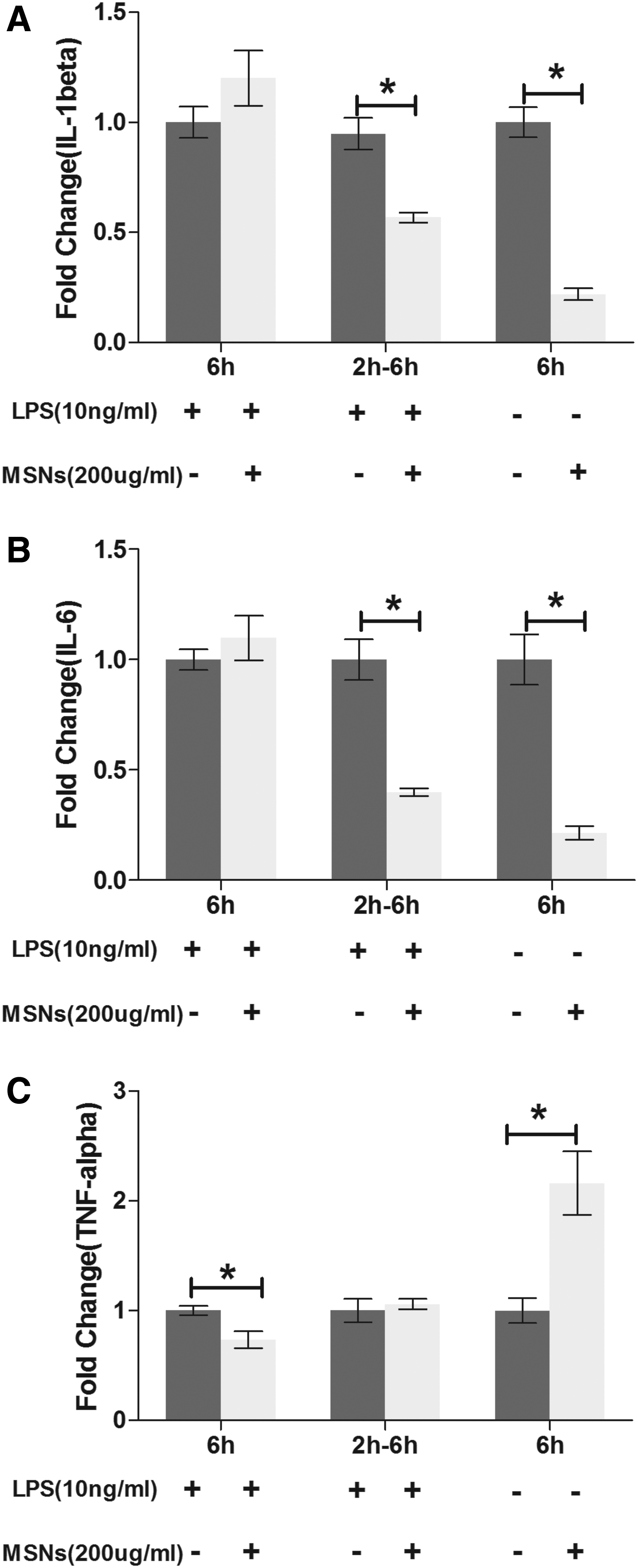
The immunomodulatory effects of MSNs on macrophages (Raw cells) were assessed under three different streams. For the first stream, cells were treated with 10 ng LPS and 200 μg/mL MSNs for 6 h. As to the second stream, cells were first treated with 10 ng/mL LPS, which was removed and replaced by 200 μg/mL MSNs for another 6 h. For the last stream, cells were just treated with 200ug/mL MSNs for 6 h, without any LPS activation. Relative mRNA expression of proinflammation cytokines genes IL-1β
Experimental pathway for LPS stimulation application
The experimental pathway proposed in this study is specially designed for the evaluation of immunmodulatory properties of biomaterials in vitro (Fig. 6). All biomaterials intended to be applied in regulating the immune reaction can follow this experimental pathway. Macrophages were applied as model immune cells. For other types of immune cells, researchers can follow the systemic protocol applied in this study to find the best stimulation of LPS. It is reasonable to choose any of the three streams applied in the previous studies, but great attentions should first be paid on what kind of immune environment the biomaterials will be implanted in. The first stream (constant activation) represents a strong inflammatory in vivo environment that may be led by the microorganism infections in situ. The second stream (short-term activation) represents a gradually diminishing inflammatory in vivo environment that may be due to the mammalian response to the surgery trauma when implanting biomaterials. The third stream (no activation) represents a normal physiological in vivo immune environment that has also been found to play key roles in maintaining local tissue dynamics. Unfortunately, the condition of immune environment is usually neglected in the current related studies. For example, when assessing the immunomodulatory effects of dental implants, the first stream is usually not suitable, because the dental implant always intends to be inserted into a germ-free bone socket with mild or moderate inflammatory response, indicating that the second or third stream is more suitable. However, it is not uncommon that the first stream is applied to evaluate the immunomodulatory effects of dental implants,19,34,35 which may lead to inaccurate conclusions.
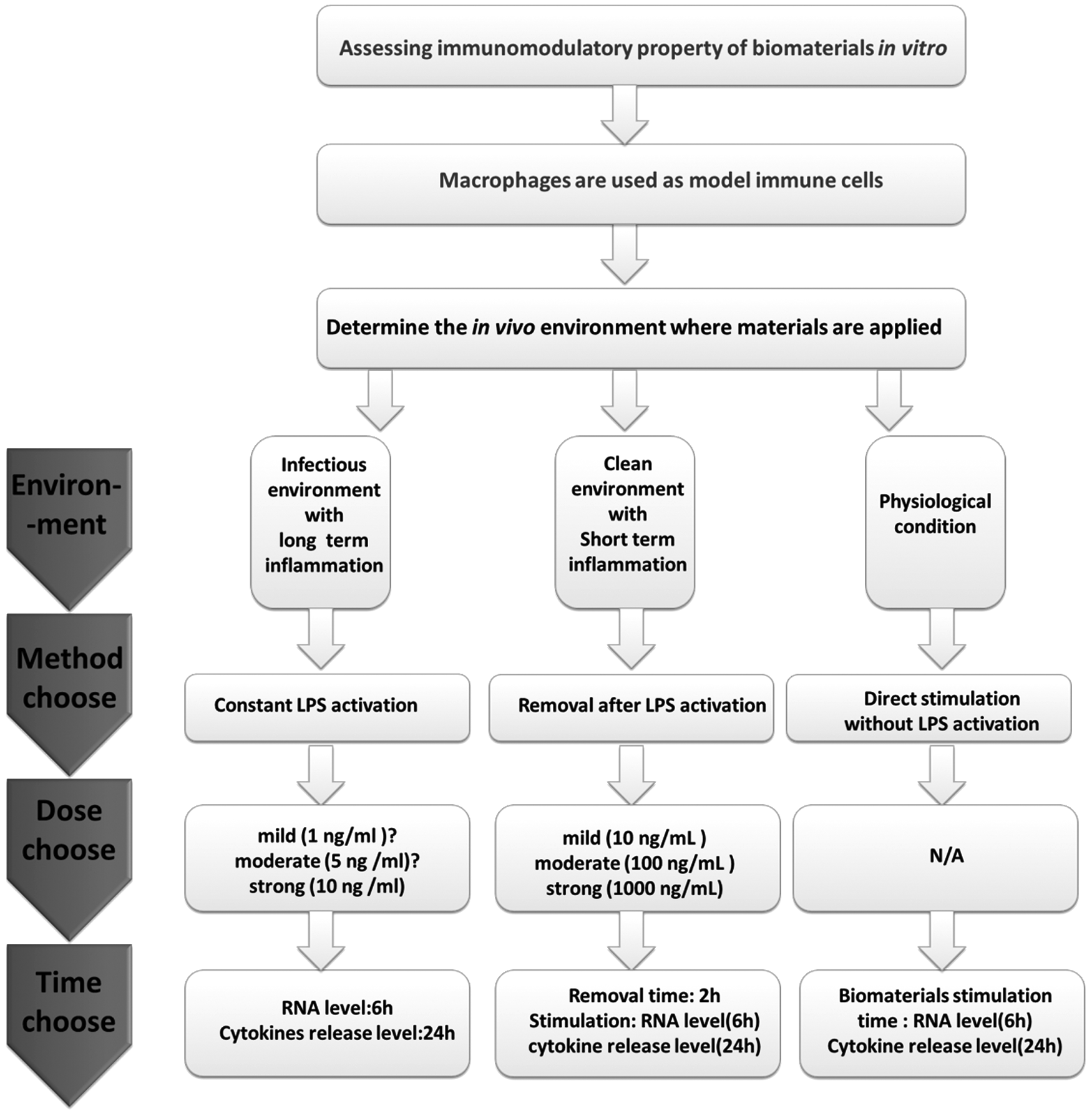
An experimental pathway of using LPS to assess the immunomodulatory property of biomaterials: According to different in vivo environment and stimulation dose, we recommend three pathways (long-term inflammation, short-term inflammation, or physiological condition) for assessing immunomodulatory property of biomaterials in vitro.
After confirming the streams, the next step would be to decide the suitable dose and stimulation time, which has been investigated in this study. LPS with impropriate dose may fail to activate macrophages or lead to an overreaction. For the first stream (constant activation), we found out that even low dose as 10 ng/mL could result in a strong inflammatory reaction by macrophages, with the release of proinflammatory cytokines similar to the dose as high as 1000 ng/mL. Therefore, 10 ng/mL is strong enough to activate macrophages, and for the moderate and mild inflammatory reaction, the dose should be lower than 10 ng/mL. As to the second stream (removal after activation), we demonstrated that macrophages continued an inflammatory reaction after the removal of LPS, however, the intensity went down as time went up. When the removal time was set at 2 h, 10 ng/mL could not significantly enhance the proinflammatory gene expression, however, ELISA results indicated the improved release of the same cytokines. This indicates that 10 ng/mL could be a starting dose in enhancing the inflammatory reaction for this stream, therefore, 10 ng/mL was defined as a mild stimulator. A dose-dependent manner could be observed in the range of 10–1000 ng/mL, 100 ng/mL, and 1000 ng/mL leading to a moderate and strong reaction respectively. Accordingly, 100 and 1000 ng/mL of LPS was defined as a moderate and strong inducer for the second stream (Fig. 6).
As to the time-dependent effect, the peak hour of proinflammatory gene expression was 6 h when macrophages were activated by LPS constantly, while release of proinflammatory cytokines was higher in 24 h under the same treatments. Therefore, we recommend that detection of gene expression can be done after 6 h stimulation, and after 24 h for the cytokines release level. In terms of the removal time for the second stream, 2 h appears to be suitable time, which is sufficient for activating macrophages even after 12 h of removal (Fig. 6).
The mechanisms underlying these different inflammatory responses by macrophages toward different LPS treatment pathways are of great interest. The effects of LPS on mediating inflammation have been well understood. LPS can be transferred to CD14 by LPS-binding protein (LBP) and recognized by Toll-like receptor 4 (TLR4) on the cellular surface of macrophages. 36 Interaction of LPS with TLR4 could lead to the activation of both nuclear factor-κB (NF-κB) and MAPK cascades, which are mediated by MyD88- and TRIF-dependent pathways. The LPS/TLR4 signal induces activation of IκB kinase (IKK), and phosphorylation and degradation of IκBα, which masks the nuclear localization signal of NF-κB. The degradation of IκBα causes nuclear translocation of NF-κB, which induces gene transcriptions of various proinflammatory cytokines.37,38 The change on LPS treatment pathways may affect the activation of these signaling pathways, thereby influencing the inflammatory response. Future studies should target more on elucidating these mechanisms and how the biomaterials affects them, so that the LPS-mediated inflammatory response can be precisely controlled and serve better for evaluating the immunomodulatory properties of materials.
Conclusions
Macrophages respond differently under different stimulation time, doses, and conditions of LPS. This implies the need for careful consideration of these parameters when applying LPS to activate macrophages. Different stimulation time, doses, and conditions of LPS might lead to a different conclusion on the immunomodulatory capacity of biomaterials. For example, MSNs demonstrated different immunomodulatory effects under different LPS and macrophage conditions. It is of great importance to apply a proper stimulation time, dose, and condition of LPS, accordingly, an experimental pathway was proposed.
Footnotes
Acknowledgments
The authors would like to thank Prof. Chengtie Wu from Shanghai Institute of Ceramics for his assistance with the preparation of MSNs. This study was supported by the Queensland-Chinese Academy of Sciences Collaborative Science Fund 2014, One Hundred Talent Program of Sun Yat-sen University (Z. C.), the Osteology Foundation (2015, young investigator) and the Prince Charles Hospital Foundation (2015, new investigator).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.