
Abstract
Mesenchymal stem cells (MSCs) have been widely used for tissue repair and regeneration. However, the inherent drawbacks, including limited cell survival after cell transplantation, have hindered direct MSC transplantation for tissue repair and regeneration. The aim of this study was to investigate if exosomes isolated from MSCs can promote the proliferation and differentiation of human primary osteoblastic cells (HOBs) and be potentially used for bone tissue regeneration. We showed that adipose tissue-derived MSC (ASC)-derived exosomes (ASC-EXO) were able to promote the proliferation and osteogenic differentiation in HOBs; and the trophic effects of ASC-EXO on HOBs were further harnessed when ASCs were preconditioned with tumor necrosis factor-alpha (TNF-α) for 3 days, which mimics the acute inflammatory phase upon bone injury. In addition, we showed that Wnt-3a content was elevated in ASC-EXO when ASCs were preconditioned by TNF-α, and inhibiting Wnt signaling decreased the osteogenic gene expression levels in HOBs which were cultured in TNF-α preconditioned ASCs conditioned medium. In conclusion, it was demonstrated that ASC-EXO, especially primed by TNF-α preconditioning on ASCs, offer a promising approach to replace direct stem cell transplantation for bone repair and regeneration.
Introduction
M
Exosomes are membrane vesicles with a diameter of 40–100 nm, a subfraction of extracellular vesicles that are secreted by many cell types, including MSCs, into the extracellular milieu. The functions of exosomes are still under investigation and current efforts focus on exploring the potential application of exosomes as biomarkers in the diagnosis of diseases. 8 Given the important role of exosomes in cell–cell communications, 9 increasing efforts are being directed to exploit their therapeutic potential.10–12 Using exosomes as therapeutic agent will provide significant advantages over the direct use of MSC transplantation such as limiting the arduous task of preserving MSC viability and function during manufacture and storage before their delivery to the patient.
MSC-derived exosomes have demonstrated their potential for promoting bone healing and/or bone tissue regeneration, but the therapeutic application of exosomes might be significantly compromised due to the low yield (around 1 μg/mL conditioned medium) of exosome production by MSC, as a great deal of exosomes would be needed for therapeutic application.13,14 Thus how we can further augment their efficacy remains explorable and of great interest for the scientist in the field of regenerative medicine. Different approaches have been utilized to harness the trophic function of MSCs, 15 such that genetic modification of MSCs by overexpression of one pivotal gene that acts as a “switch” can alter the profile of the released factors and, thus, maximize the therapeutic potential. 16 We have been searching for the potential compounds which are able to stimulate secretion of trophic factors from MSCs; as such a chemical approach for manipulating stem cell behavior has significant advantages in comparison to genetic manipulation. 17 We previously selected one of the main inflammatory factors (tumor necrosis factor-alpha, TNF-α) as a preconditioning agent, because the role of acute inflammation in response to tissue injury has recently been implicated in tissue repair and regeneration,18,19 and we demonstrated that 3 days of TNF-α preconditioning (mimicking the acute inflammatory microenvironment) direct adipose tissue-derived MSCs (ASCs) into osteogenic differentiation through a BMP-2 autocrine manner. 20 However, it is unknown if TNF-α preconditioning can serve as a feasible approach to promote the trophic function of stem cells.
In this study, we investigated if ASC-derived exosomes affect human primary osteoblastic cell (HOB) proliferation, migration, and differentiation. We further determined if 3 days of TNF-α preconditioning (mimicking the inflammatory phase upon bone injury) on ASCs would modulate the release of the trophic factors (e.g., exosomes) from ASCs. We demonstrated that the ASC-derived conditioned medium (ASC-CM) containing the trophic factors released from the ASCs was able to promote the proliferation and osteogenic differentiation in HOBs. Interestingly, the trophic function of ASCs was further potentiated when ASCs were preconditioned for 3 days with TNF-α. In support of the roles of exosome in mediating the trophic function of ASCs, we showed that the nanoparticles in the conditioned medium from ASCs, as well as TNF-α preconditioned ASCs, expressed exosome-specific markers (CD9 and CD81) by western blot. In addition, the removal of exosomes from the ASC-CM or TNF-α preconditioned ASC-CM (TNF-ASC-CM) largely diminished its effects on HOBs. Finally, we demonstrated that TNF-α preconditioning harnessed the trophic function of ASC-derived exosomes partially through a mechanism of increasing their Wnt-3a content.
Materials and Methods
ASC culturing and collection of ASC-CM
ASCs (Life Technology) were propagated according to the manufacturer's instructions in MesenPRO RS™ Basal Medium (Invitrogen) with the supplement of 2 mM l-glutamine and MesenPRO RS Growth Supplement (Life Technology). ASCs at passage 4 were used for all the studies and were cultured in complete growth medium (α-minimal essential medium [α-MEM]; Life Technology), supplemented with 10% (v/v) fetal calf serum (FCS; Life Technology), 100 units/mL penicillin, and 100 mg/mL streptomycin (Life Technology), and were maintained until they reached 80–90% confluence.
Collection of ASC-CM and depletion/isolation of exosomes
TNF-α preconditioning procedure was as follows: ASCs (passage 4) at 80–90% confluence were cultured in α-MEM serum-free medium (control) or with the addition of TNF-α (preconditioning group, 1 ng/mL, human recombinant protein; Sigma-Aldrich, St. Louis, MO) for 3 days. At day 3, the medium was removed and the ASCs washed with warm phosphate-buffered saline (PBS) solution thrice and cultured again in α-MEM serum-free medium for 3 days. At day 3, the medium was collected and passed through 0.22 μm pore-sized filters to remove all the dead cells or cell debris to obtain ASC-CM or TNF-ASC-conditioned medium (TNF-ASC-CM).
Depletion of exosomes from the medium: exosomes were first pelleted by ultracentrifugation (Beckman Optima L-80XP) at 100,000 g for 4 h, and the supernatant (exosome-depleted CM) was collected and the pellets were resuspended in fresh PBS and further ultracentrifuged at 100,000 g at 4°C for another 60 min. The supernatant was then completely removed and the exosome pellets were collected for analysis. It was important to establish if the trophic function of ASC-CM was diminished by the removal of the exosomes. We first determined if the trophic function of ASC-CM was diminished by the removal of exosomes using exosome-depleted conditioned medium or we can regain a trophic function by adding exosome pellets into α-MEM culture media.
HOB isolation
Permission to use discarded human tissue was granted by the Human Ethics Committee of the University of Sydney, and informed consent was obtained. Human trabecular bone was chopped into 1 mm3 pieces and washed several times in PBS, followed by digestion for 90 min at 37°C with 0.02% (w/v) trypsin (Sigma-Aldrich) in PBS. Digested cells were cultured in complete media containing α-MEM (Gibco Laboratories), supplemented with 10% (v/v) FCS (Life Technology), 30 mg/mL penicillin, 100 mg/mL streptomycin (Life Technology), and 1 mM l-ascorbic acid phosphate magnesium salt (Wako Pure Chemicals, Osaka, Japan). The cells were cultured at 37°C with 5% CO2, and the medium was refreshed every 3 days until 80–90% confluence when cells were passaged. All HOBs used in the experiments were at passage 2.
HOB proliferation and migration assays
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay was used for assessing HOB proliferation after 3 and 7 days of culture in serum-free α-medium, or ASC-CM, TNF-ASC-CM, or osteogenic medium (α-MEM supplemented with 10% FCS, 50 μg/mL ascorbic acid, and 10 mM β-glycerophosphate). One hundred microliters of the reacted reagent from each well was transferred to a 96-well plate, and the absorbance was recorded using a microplate reader (PathTech) at a wavelength of 490 nm.
For the assay of cell mobilization, HOBs were trypsinized and then resuspended in serum-free α-MEM medium (3 × 106 cells/mL). The cell suspension was mixed 1:1 with the solution of Matrigel from Cultrex (Bio Scientific). From that suspension, 20 μL (containing 3 × 104 cells) was layered onto the surface of six-well tissue-culture dishes to form well-defined droplets. Dishes were placed at 37°C for 5 min for Matrigel to semisolidify. Two milliliters different media (serum-free α-medium, ASC-CM, TNF-α-ASC-CM, or osteogenic medium) were then added to the wells, and plates were incubated for up to 18 h. The number of cells that mobilized out of the droplet was scored by phase-contrast microscopy.
Exosome characterization and internalization analysis
The concentration and size distribution of the isolated exosomes were measured using NanoSight. Before sampling, the ASC-CM or TNF-ASC-CM was diluted in PBS to a final dilution of 1:500 to obtain the optimal nanoparticle density for measurement. A blank PBS was run as a negative control. Each sample analysis was conducted for 90 s. The NanoSight automatic analysis settings (high sensitivity, green laser) were used to process the data. All samples were evaluated in triplicate.
ASC-CM was collected and centrifuged at 3000 g for 30 min, followed by filtration through 0.22 μm filter to remove dead cells and cell debris. Exosomes were pelleted by ultracentrifugation at 100,000 g for 4 h and resuspended in PBS. The exosomes in PBS were then labelled by Exo-GLOW™ Exosome Labeling Kits (System Biosciences) following the manufacturer's instructions. The labelled exosomes isolated from 2 mL ASC-CM were added into HOBs (70% confluence) in 24-well plate, and the cells were visualized by fluorescence microscopy after 24 h.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from HOBs by adding TRIzol reagent (Sigma) after the medium was removed according to the manufacturer's instructions. First strand cDNA was synthesized from 0.7 μg total RNA using the Tetro cDNA Synthesis Kit (Bioline) according to the manufacturer's instructions. Real-time PCR was performed in Rotor-Gene 6000 (Corbett Life Science) using ImmoMix (Bioline) according to manufacturer's instructions, and the relative gene expression levels for Runx2, osteopontin, bone sialoprotein, and type 1 collagen were obtained by normalizing them to the housekeeping gene (18S) (primer sequences shown in Table 1).
Assessment of mineralization
The HOBs were cultured for 21 days in different media, including control (serum-free α-MEM, ASC-CM, TNF-ASC-CM, exosome-depleted ASC-CM, exosome-depleted TNF-ASC-CM, or osteogenic medium), at which time point Alizarin Red staining was carried out to assess calcium deposit by ASCs. In brief, after 21 days of culturing, the medium from each well was carefully aspirated, and cells were fixed by incubating in ice-cold 70% ethanol for 1 h at room temperature followed by rinsing the cells twice (5–10 min each) with water. The water was then aspirated, and 1 mL Alizarin Red Solution was added to cover each well of the 24-well plate and incubated at room temperature for 30 min. After 30 min, the Alizarin Red solution was removed, and the cells were washed four times with 1 mL water. One to 1.5 mL water was added to each well before visual inspection and image acquisition. ImageJ was used to quantify the staining area.
Blocking of Wnt-3a signaling
Blocking Wnt3a signaling was carried out to determine if Wnt3a signaling plays a role in ASC-CM or TNF-ASC-CM induced osteogenic gene expression in HOBs. The inhibitor (IWR-1; Sigma-Aldrich) was dissolved in DMSO and added to the control medium (serum free), ASC-CM, and TNF-ASC-CM at a final concentration of 50 μM. At days 3 and 7, osteogenic gene expression in HOBs was compared between DMSO group and IWR-1 treatment group.
Western blotting
HOBs or exosome pellets were washed with ice-cold PBS and lysed for 30 min in ice-cold RIPA lysis buffer [20 mM Tris–HCl, pH 7.5, 1 mM EDTA, 1 mM EGTA, 150 mM NaCl, and 1% Triton X-100, protease inhibitor cocktail (Sigma-Aldrich)]. Protein concentration was measured using the BCA Protein Assay Kit (Pierce). Equal aliquots of protein (10 μg) were heated at 70°C for 10 min in 4× sample buffer (WesternBreeze; Life Technology) and 10× reducing buffer and separated on 8–12% SDS-PAGE gels (WesternBreeze; Life Technology). Proteins were transferred to PVDF membranes, washed with 1× TBS-T (20 mM Tris–HCl pH 7.6, 137 mM NaCl, containing 0.1% Tween 20), and blocked for 1 h at room temperature in 1× TBS-T with 1% BSA. The membranes were washed thrice followed by incubation with primary antibody of anti-β-actin (1:5000; Abcam), anti-Wnt3a (1:1000; Millipore), anti-CD9, and anti-CD81 (1:1000; System Biosciences), in TBS-T containing 1% BSA overnight at 4°C. After three washes, the membranes were incubated with secondary antibody (WesternBreeze; Life Technology) for 60 min followed by another three washes before protein bands were visualized with chemiluminescent reagents (WesternBreeze; Life Technology) in ChemiDoc MP Imaging System (Bio-Rad Life Sciences).
Statistical analyses
Data in this study were obtained from at least four independent experiments and represented with mean ± standard error of the mean (SE). For statistical analysis, first Levene's test was performed to determine the homogeneity of variance for all the data, and then independent t-test was used for analyzing the data between two groups while Tukey HSD post hoc test was used for the data among three groups. SPSS 17.0 program was used for all statistical analyses, and differences were considered significant if p < 0.05.
Results
ASC-CM promoted the proliferation and osteogenic differentiation of HOBs
Compared to the control group (serum-free medium), HOBs cultured with ASC-CM had a significant increase in their proliferation at days 3 and 7 (Fig. 1A), but with no significant increase in the number of HOBs mobilized from the Matrigel after 18 h of culturing (Fig. 1B). In addition, ASC-CM significantly increased the gene expression levels of Runx2, collagen I, osteopontin, and bone sialoprotein in HOBs after 7 days of culturing on day 7 (Fig. 1C). Alizarin red staining was also performed to examine the mineral deposition and bone nodule formation by HOBs, and we showed that some bone nodules were observed in HOBs cultured in ASC-CM for 21 days, while no bone nodules were detected in the control medium (Fig. 1D). However, osteogenic medium (supplemented with 10% FCS, 50 μg/mL ascorbic acid, and 10 mM β-glycerophosphate) had much larger effects on HOB proliferation, migration, and osteogenic differentiation than ASC-CM (Fig. 1A–D).

ASC-CM promoted the proliferation and osteogenic differentiation of HOBs.
The effects of ASC-CM on HOBs were potentiated by TNF-α preconditioning on ASCs
We next examined if the trophic function of ASCs would be harnessed by their preconditioning with TNF-α (1 ng/mL) for 3 days. Compared with HOBs cultured in ASC-CM, TNF-ASC-CM significantly increased HOB proliferation at days 3 and 7 (Fig. 2A), migration at 18 h (Fig. 2B), and bone sialoprotein gene expression at day 7 (Fig. 2C). Alizarin red staining also demonstrated that more bone nodules (about two times increase) formed in the HOBs were grown in TNF-ASC-CM after 21 days of culturing (Fig. 2D).
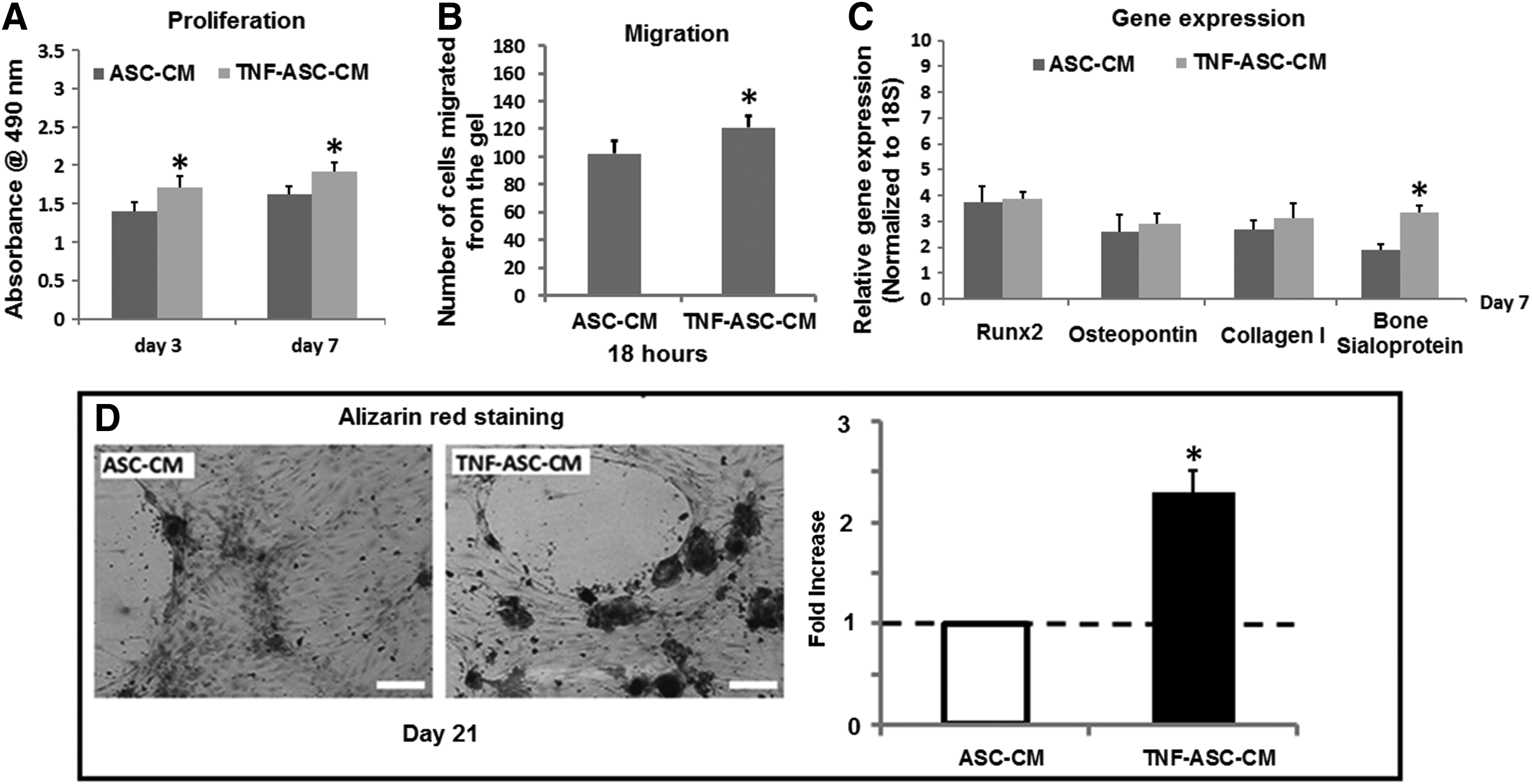
The effect of ASC-derived conditioned medium (ASC-CM) on HOBs was potentiated by TNF-α preconditioning on ASCs. Compared with ASC-CM, the conditioned medium from ASCs by 3 days of TNF-α preconditioning (TNF-ASC-CM) further increased HOB proliferation, migration, and bone sialoprotein gene expression
Characterization of exosomes in ASC-CM or TNF-ASC-CM and internalization by HOBs
We used the NanoSight to characterize the nanoparticles in the ASC-CM and TNF-ASC-CM and demonstrated that ASC-CM and TNF-ASC-CM contained a large amount of nanoparticles with a concentration of 3.04e + 008 ± 2.59e + 007 particles/mL and 2.02e + 008 ± 3.24e + 007 particles/mL, respectively. The average size of nanoparticle was 130 and 110 nm in ASC-CM and TNF-ASC-CM, respectively. The exosome pellets purified from ASC-CM and TNF-ASC-CM expressed the typical marker of exosomes (CD9 and CD81, Fig. 3A), while negative control (serum-free medium) did not. In addition, when the exosomes purified from ASC-CM or TNF-ASC-CM were labeled and added into HOB culture, there was no difference of exosome internalization by HOBs between the exosomes purified from ASC-CM and TNF-ASC-CM, and the internalization was apparent after 24 h of incubation (Fig. 3B).
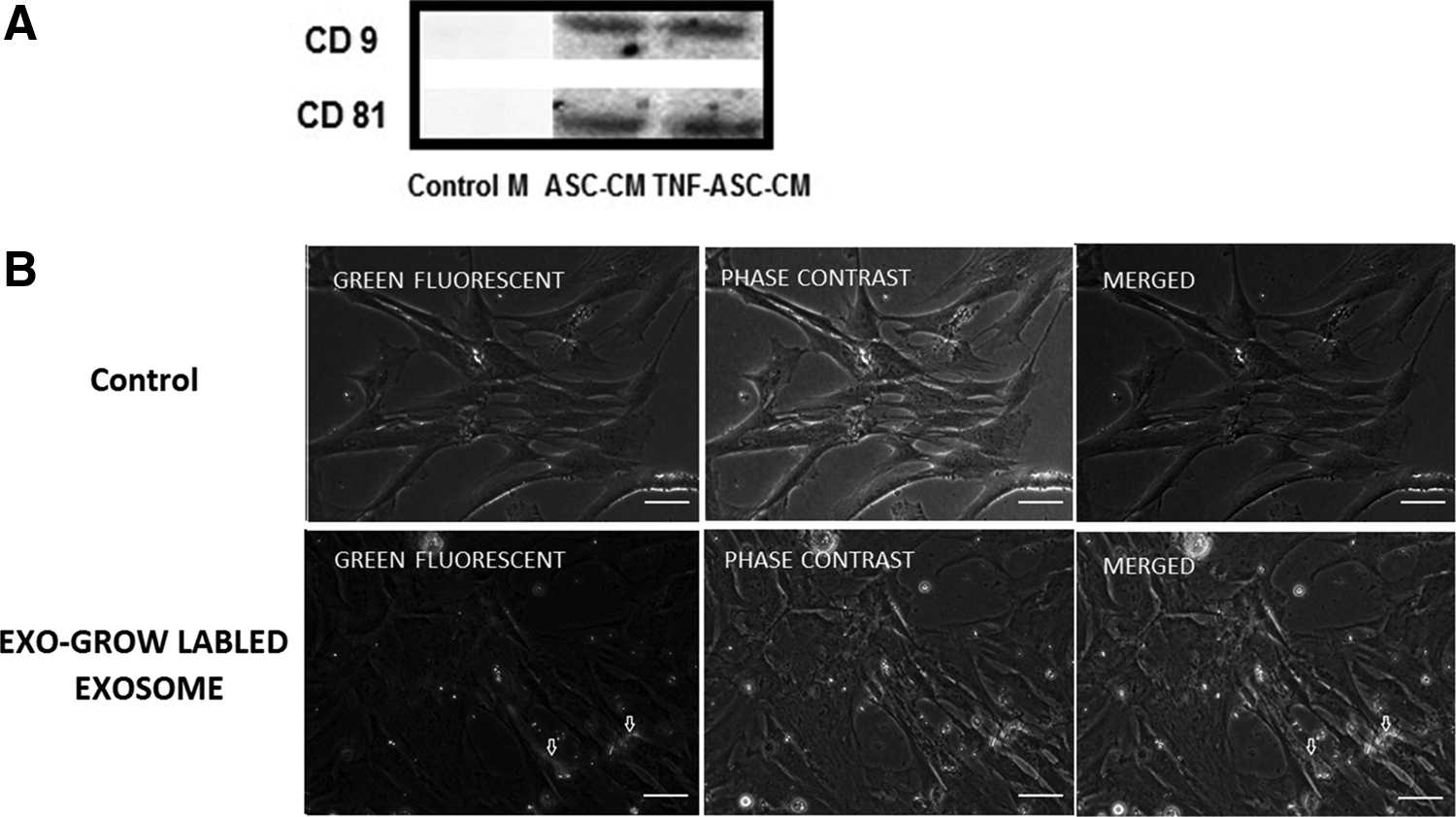
Characterization of exosomes in ASC-CM or TNF-ASC-CM and exosome internalization by HOBs.
The removal of exosomes from the ASC-CM or TNF-ASC-CM largely diminished its effect on the proliferation, migration, and osteogenic differentiation in HOBs
To confirm the role of exosomes in ASC-CM/TNF-ASC-CM in mediating the effects on HOBs, exosomes were depleted from the ASC-CM or TNF-ASC-CM by ultracentrifuging, and then HOBs were cultured in these exosome-depleted media for 7 days. We demonstrated that the depletion of exosomes from the culture medium significantly decreased HOB cell proliferation at days 3 and 7 (Fig. 4A) and migration at 18 h (Fig. 4B), compared to those seen for ASC-CM or TNF-ASC-CM. Moreover, in comparison to those seen for ASC-CM or TNF-ASC-CM, there was a significant reduction in the gene expression levels for Runx2, collagen I, osteopontin, and bone sialoprotein by HOBs cultured in exosome-depleted ASC conditioned medium at days 3 and 7 (Fig. 4C–F). There was no bone nodule formation at day 21 when HOBs were cultured in exosome-depleted ASC-CM or TNF-ASC-CM, which were seen in the HOBs cultured in exosome-contained ASC-CM or TNF-ASC-CM (Fig. 4G–J).
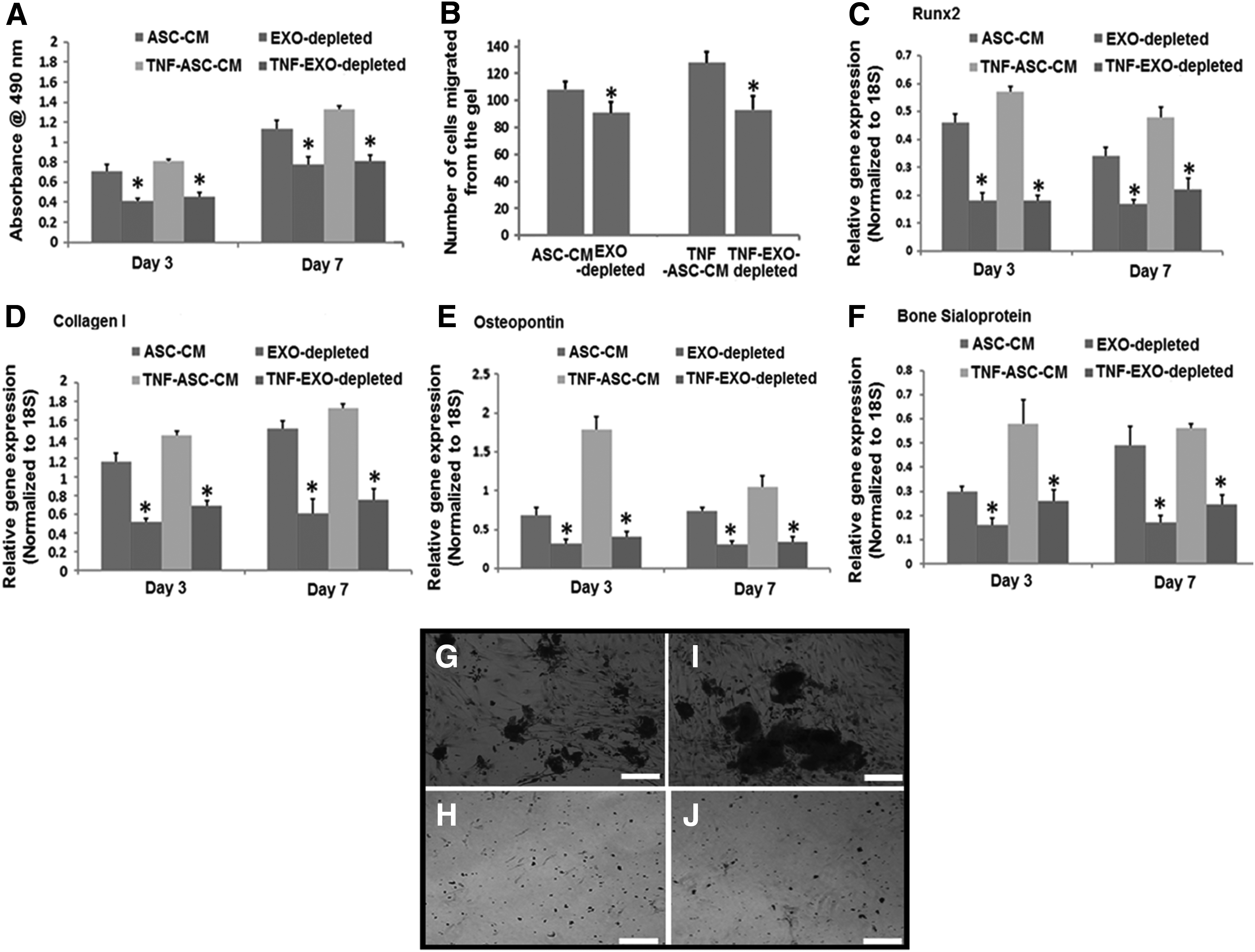
The removal of ASC-EXO from the ASC-CM or TNF-ASC-CM largely diminished their effects on the proliferation, migration, and osteogenic differentiation in HOBs.
We also recovered the exosome pellet followed by resuspending them in the control medium (serum-free medium) and demonstrated that the serum-free medium gained the trophic function of exosomes and had the similar effects of ASC-CM or TNF-ASC-CM on HOB proliferation and differentiation as shown in Figure 1.
TNF-α potentiated the function of ASC-EXO partially through a mechanism of increasing Wnt-3a content in ASC-EXO
To gain more insight into the mechanism(s) by which TNF-α potentiates the function of ASC-EXO on HOBs, we measured Wnt-3a protein contents in ASC-EXO and TNF-ASC-EXO. Exosomes purified from the conditioned medium by TNF-α preconditioned ASCs had higher expression of Wnt-3a than that in the conditioned medium from ASCs without TNF-α preconditioning (Fig. 5A). Moreover, the HOBs cultured in TNF-ASC-CM showed a significant increase in total β-catenin expression level compared to that in the HOBs cultured in ASC-CM (Fig. 5B). The total β-catenin expression level in HOBs was decreased when they were cultured in exosome-depleted TNF-ASC-CM (Fig. 5B).
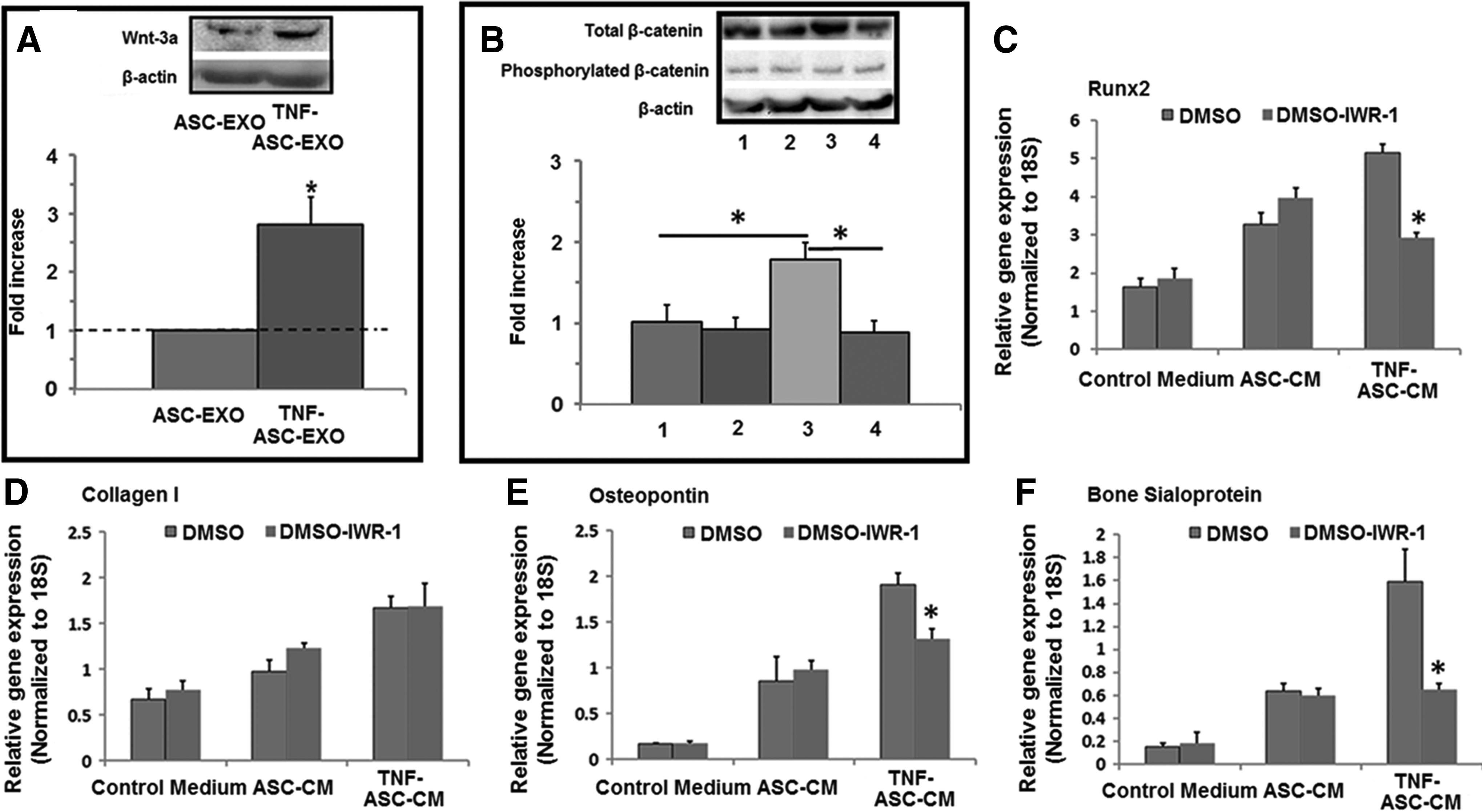
TNF-α potentiated the function of ASC-EXO partially through a mechanism of increasing Wnt-3a content in ASC-EXO.
To confirm the role of Wnt signaling in exosome-mediated osteogenic induction in HOBs, we added IWR-1 (Wnt signaling inhibitor) into the different culture media: control medium, ASC-CM, or TNF-ASC-CM. The presence of IWR-1 significantly decreased the levels of Runx-2, osteopontin, and bone sialoprotein gene expression in HOBs only when they were cultured in TNF-ASC-CM, but not for control medium or the ASC-CM at day 7 (Fig. 5C–F).
Discussion
In this study we explored the potential of using the exosomes produced by ASCs for use in bone tissue regeneration. We demonstrated that ASC-derived exosomes are able to efficiently promote the proliferation and osteogenic differentiation of primary human osteoblasts. In addition, we showed that 3 days of TNF-α pretreatment on ASCs further potentiated the function of exosomes released from ASCs partially through a mechanism of increasing the exosome Wnt-3a content.
It was originally thought that the mechanisms of using MSCs for tissue repair are their homing to the injured tissue, to differentiate, replace, and restore the function of the damaged tissue. 21 Increasing evidence, however, indicates that MSC engraftment and differentiation to injured sites are low and transient. 22 Instead, MSCs exert their therapeutic effects through secreted paracrine or trophic factors. 23 The studies using MSC-conditioned media demonstrated that MSCs are able to induce wound healing through the mechanism of paracrine.24,25 In agreement with these studies, current study showed that ASC-derived conditioned medium significantly enhanced HOB proliferation and osteogenic differentiation, suggesting that the trophic factors are released from ASC which then act on HOBs to exert their therapeutic effects.
MSC-conditioned medium contains a number of cytokines and growth factors such as interleukin-6 and fibroblast growth factor.26,27 In addition to the soluble factors in the medium, exosomes produced by MSCs contain a number of potential regulatory components, including miRNAs, mRNAs, and proteins, which can be transferred as a type of “physiological lipofection” to recipient cells to modify their characteristics. 28 For example, MSC exosomes have been found to transfer growth factor receptor mRNA, which was translated to the corresponding protein in renal tubular cells. 29 In our study, we hypothesized that exosomes released by ASCs play a crucial role in ASC-CM-mediated effects on HOBs. To test this hypothesis, we depleted the exosomes in the CM by ultracentrifuge to test if exosome-depleted CM would lose its trophic functionality. We demonstrated that HOBs cultured in exosome-depleted ASC-CM had significantly lower capability to proliferate, migrate, and osteogenic differentiation, compared to the HOBs cultured in exosome-contained ASC-CM. Moreover, resuspending the isolated exosomes into the plain medium (serum-free) resulted in the regain of its trophic function, further confirming the key role played by the exosomes released by ASCs in ASC-CM mediated trophic functions on HOBs.
The identification of critical roles of trophic factor secretion for the functionality of MSCs sparked the possibility that we might be able to modify the quality and/or concentration of secreted trophic factors by MSCs within the injured area. One of the ways to achieve the higher concentration of trophic factors would be by direct injection/transplantation of more MSCs into the injured organ (and hence more bioactive factors). However, one main limitation for direct cell transplantation as discussed above is the extensive early death of grafted cells. Studies performed in solid organs showed that more than 80–90% of grafted cells die within 72 h after injection.30–32 A plausible approach would be to modify and isolate the concentrated trophic factors in vitro and then administer them (in vivo) into the site of injury. In this study, we showed that the trophic function of ASCs (promoting proliferation and osteogenic differentiation in HOBs) was significantly potentiated when ASCs were pretreated for 3 days with TNF-α. These results showed that there appears to be no significant difference in the number of exosomes generated in response to TNF treatment, but rather the osteogenic activity of these exosomes is increased, suggesting that TNF-α preconditioning can be served as a promising approach to harness the release of trophic factors (e.g., exosomes) from stem cells, consolidating the potentiality of stem cell-derived exosomes as a cell-free therapeutic approach for bone tissue repair and regeneration.
The exact mechanisms by which exosomes exert their therapeutic efficacy are not yet fully understood, but it is thought to be dependent on their composition, including proteins and miRNAs, 33 which are released then internalized by neighboring cells, and subsequently exert their systematic effects. We showed that ASC-EXO indeed could be internalized by HOBs. We further investigated the mechanism by which ASC-EXO modulates HOB function by focusing on Wnt-3 signaling, as Wnt family has been shown to play a very important role in new bone formation and bone homeostasis, 34 and Wnt proteins exist in exosomes released by MSCs and act as key components for their therapeutic function.35,36
In agreement with these findings, we showed in this study that exosomes purified from ASC-CM contained Wnt-3a protein. However, the presence of IWR-1 (Wnt signaling inhibitor) did not significantly decrease the osteogenic gene expression levels in HOBs cultured in the control medium or ASC-CM, indicating that activating Wnt signaling is not the key mechanism of ASC-EXO-mediated trophic function on HOBs. We therefore proposed that the exosomes from ASCs are more likely to exert their trophic function through a systematic route rather than through a single signaling pathway, which is in agreement with other researchers.37,38 Very interestingly, however, the exosomes from TNF-ASC-CM contained significantly higher levels of Wnt-3a, and inhibiting Wnt signaling resulted in significant decrease in osteogenic gene expression levels by HOBs cultured in TNF-ASC-CM. These results suggested that 3 days of TNF-α pretreatment on ASCs potentiated the trophic function of exosomes released from ASCs through a mechanism of increasing the Wnt-3a content in exosomes. It is therefore plausible to imply that it would be a feasible approach to produce more potent exosomes for therapeutic application by pharmaceutical pretreatment (e.g., TNF-α) on stem cell to modify their exosome components, and such an approach would be of great importance for translating the modified ASC-EXO for clinical application.
In summary, the overarching goal of this study would be to use exosomes as a systematic therapeutics for bone tissue repair or regeneration, so that we are able to circumvent the drawbacks of direct MSC transplantation for bone tissue engineering. The results demonstrate that ASC-derived exosomes promote the proliferation and osteogenic differentiation of HOBs, which can be further potentiated by preconditioning ASCs with TNF-α, suggesting that the exosomes from TNF-α preconditioned ASC might offer a promising approach to replace direct stem cell transplantation for bone repair and regeneration. However, to ultimately develop the approach of using ASC-EXO for treating bone regeneration-related diseases, more studies will be needed to further reveal the effective components in exosome and how these components would coordinate to contribute to bone regeneration in a systematic manner.
Footnotes
Acknowledgments
The authors acknowledge the Australian National Health and Medical Research Council (NHMRC), the Rebecca Cooper Foundation, Australian Research Council and the facilities provided by School of AMME, and the Bosch Institute at the University of Sydney.
Disclosure Statement
No competing financial interests exist.