
Abstract
There is an emerging need for soft tissue replacements in the field of reconstructive surgery for the treatment of congenital deformities, posttraumatic repair, and cancer rehabilitation. Previous studies have shown that the bioactive adipose tissue extract can induce adipogenesis without additional stem cells or growth factors. In this study, we innovatively investigated whether exosome-like vesicles derived from adipose tissue (ELV-AT) could direct stem cell differentiation and trigger adipose tissue regeneration. In vitro, ELV-AT can induce adipogenesis of adipose-derived stem cells and promote proliferation, migration, and angiogenic potential of the aorta endothelial cells. In vivo, ELV-AT were transplanted to a chamber on the back of nude mice and neoadipose tissues were formed. Our findings indicated that ELV-AT could be used as a cell-free therapeutic approach for adipose tissue regeneration.
Introduction
T
Exosomes, one type of extracellular vesicles (EVs), 10 have gained the most attraction as a promising cell-free bioactive molecule that can be used in therapeutic applications due to their important roles of intercellular communications. 11 However, due to the lack of selective markers, state-of-the-art exosome isolation method yields a mixture of vesicles with similar morphology and size to exosomes, which are defined as exosome-like vesicles.12,13 As the proteins and nucleic acids contained in exosomes can reflect the original cell sources and modulate biological responses of the target cells, exosomes derived from stem cells have gained increasing attention in the field of regenerative medicine because of their multilineage differentiation capacity. Studies have shown that exosomes derived from mesenchymal stem cells (MSCs) repair bone defects and induce skeletal muscle regeneration through enhanced angiogenesis.14,15
The regeneration of adipose tissue involves various cells, signal pathways, and bioactive molecules. 16 Sarkanen et al. found that cell-free adipose tissue extract (ATE) can effectively induce adipogenesis.6,17 Lu et al. confirmed the inductive effect of ATE on adipose tissue regeneration in an engineering chamber model. 5 However, to date, there is no study about the effect of exosome-like vesicles derived from adipose tissue (ELV-AT), a kind of vesicle derived from tissue not from cells, on adipose tissue regeneration. In this study, we assume that ELV-AT can promote the specific homing of stem/progenitor cells and innovatively applied the ELV-AT to generate engineering adipose tissue.
Materials and Methods
Animals
Animals were obtained from Dashuo Experimental Animal Co. Ltd. (Chengdu, China). This study was reviewed and approved by the Ethics Committees of the State Key Laboratory of Oral Diseases, West China School of Stomatology, Sichuan University.
Isolation of adipose tissue-derived exosome-like vesicles
Inguinal fat pads from 4-week-old Sprague Dawley (SD) rats were minced into small pieces and transferred into a Celstir spinner flask (Wheaton). Serum-free α-minimal essential medium (α-MEM; Hyclone) supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin was added into the flask and the tissues were cultured at 37°C with speed at 100 rpm for 3 days. The debris of tissues and cells were removed by centrifugation (1200 rpm, 10 min). An additional centrifugation in Amicon® Ultra-50 Centrifugal Filter Units with Ultracel-3 membrane (3000 Mw cutoff membrane, Millipore) at 5000 g for 30 min was applied to concentrate ATE. Then, the ATE was mixed with the Total Exosome Isolation™ reagent (Life Technologies) at 4°C overnight and spun down at 10,000 g for 1 h at 4°C. The obtained pellet (ELV-AT) was resuspended in a serum-free solution and stored at −80°C with known concentration determined by the bicinchoninic acid method.
Cell culture
Aorta endothelial cells (ECs) were obtained from the thoracic aorta of 4-week-old SD rats. The vessel was rinsed with phosphate-buffered saline, cut into small pieces, supplemented with 3–5 mL digestion solution (1 mg/mL dispase, 0.075% type I collagenase), and incubated at 37°C for 1 h. Thereafter, the samples were centrifuged and the pellet was resuspended in growth medium (EGM-2MV; Lonza).
Adipose-derived stem cells (ADSCs) were isolated as described in our previous study.18,19 Inguinal fat pads were collected, cut into small pieces, and digested with 0.075% type I collagenase for 30 min at 37°C. The cells were cultured in α-MEM (HyClone) with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin.
Transmission electron microscopy
ELV-AT were fixed in 1% glutaraldehyde solution overnight. The sample was washed twice and loaded onto formvarcarbon-coated grids. Then, the grids were contrasted with 1% phosphotungstic acid for 60 s and were examined with a transmission electron microscopy (Hitachi H7500, Japan).
Dynamic light scattering
Dynamic light scattering measurements were performed using Zetasizer Nano ZS90 (Malvern, United Kingdom) to determine the size of ELV-AT. The size distribution was plotted with an x-axis showing the distribution of estimated particle radius (nm) and a y-axis exhibiting the relative intensity.
Western blotting
Total vesicular or cellular proteins were extracted using the Total Protein Extraction Kit (KeyGEN, China) and separated on polyacrylamide gels, blotted onto a nitrocellulose membrane. Then, the samples were probed with a primary antibody followed by the horseradish peroxidase-coupled secondary antibody. The labeled proteins were visualized using ImageQuant LAS 4000 mini (GE Healthcare).
Cell proliferation assays
The proliferation of ECs was analyzed using the Cell Counting Kit-8 (Dojindo, Japan). ECs were seeded at 2 × 104 cells per well into 96-well plates and allowed to adhere overnight at 37°C. Then, cells were maintained in EM (EGM-MV medium) containing ELV-AT with 0, 10, 50, and 200 μg/mL. At 1, 2, 3, and 5 days, 10 μL cell counting solution was added into each well and incubated at 37°C for 1 h. The OD value was detected using the Multiskan Go Spectrophotometer (Thermo Fisher Scientific).
Cell migration assay
Cell migration was determined using a Chemotaxicell chamber (8 μm pore, Osaka, Japan). ECs were added to the upper chamber at a density of 105 cells per well, and ELV-AT (0, 10, 50, and 200 μg/mL) were added to the bottom well and incubated for 6 h. Cells that migrated to the lower surface of the membrane were fixed with 4% paraformaldehyde and stained with Giemsa staining solution (Solarbio, China).
Tube formation assays
ECs were pretreated with 0 and 50 μg/mL ELV-AT for 4 days, respectively, then collected, and seeded at 104 cells per well onto Matrigel-coated wells in 96-well plates. After incubation for 6 h, phase-contrast images were acquired on an inverted microscope (Olympus, Japan). The total number and length of all tubing within each field were measured using Image Pro Plus software.
Angiogenic differentiation of ECs
ECs were seeded in six-well plates at a density of 105 cells per well and then cultured with two different media: (1) EM medium as negative control and (2) EM medium supplemented with 50 μg/mL ELV-AT. After 4 days, the expression of angiogenic genes was determined by real-time polymerase chain reaction (PCR).
Adipogenic differentiation of ADSCs
ADSCs were seeded at 105 cells per well into six-well plates. Fifty micrograms per milliliter ELV-AT was used to investigate its adipogenic potential, with the basal medium alone as negative control. The expression of adipogenic genes was analyzed by real-time PCR after 5 days of induction. After 14 days in culture, adipogenic differentiation was determined by oil red O staining.
Real-time PCR
The RNAiso Plus (TaKaRa Biotechnology, Japan) was used to extract total RNA, which was reverse transcribed into cDNAs using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific). The synthesized cDNAs were amplified with SYBR Premix ExTaq (TaKaRa Biotechnology, Japan) using QuantStudio 6 Flex Real-Time PCR System (Life Technologies, Carlsbad, CA). The PCR cycling parameters were 95°C for 2 min, 44 cycles of 95°C for 5 s, and 60°C for 30 s. And the primers used for real-time PCR were shown in Table 1.
Animal experiments
All operations were performed on 8-week-old nude mice (25–30 g, n = 20) under general anesthesia (1% pentobarbital sodium, 10 mL/kg, intraperitoneal injection). A silicone tube (Dow Corning, Fisher Scientific, United Kingdom) with an internal diameter of 4.87 mm and a height of 5 mm was subcutaneously implanted into the transverse incision (about 1.5 cm) on the back of mice (Fig. 4). The incision was closed with 3/0 nylon suture. Each mouse was implanted with two tubes: one was filled with 200 μL Matrigel (Corning) containing 200 μg ELV-AT by injection and the other one was 200 μL Matrigel alone. Mice were sacrificed at 2, 4, 8, and 12 weeks, respectively (n = 5 per time point), for analysis of implanted tubes and their contents.
Hematoxylin and eosin staining
The implanted tubes and their contents were fixed in 4% paraformaldehyde overnight at 4°C. After removing the tubes, remaining tissues were dehydrated using graded ethanol and paraffin embedded for hematoxylin and eosin staining.
Immunochemical staining
Samples were blocked for 2 h and then incubated overnight at 4°C with primary antibodies. Primary antibodies against perilipin A (ab3526; Abcam) and CD34 (sc-7324; Santa Cruz) were used in this study. The primary antibody was shown by the DAB kit (Gene Tech, Shanghai, China) or Alexa Fluor 555 goat anti-mouse (A21422; Invitrogen) or Alexa Fluor 555 goat anti-rabbit (A21428; Invitrogen). Nuclear staining was performed using DAPI (D1306; Life Technologies).
Statistical analysis
Results are expressed as mean value ± standard deviation. Statistical analysis was performed with Student's paired t-test. p < 0.05 was considered as statistically significant.
Results
Characterization of ELV-AT
ELV-AT displayed a round or cup-like shape with an average diameter of 100 nm. The marker for general EVs (HSP70) and exosomal markers (CD9 and CD63) were validated by western blotting, while the cellular protein ACTIN was detected exclusively in the cell lysates (Fig. 1). The vesicles used in this study qualified as exosome-like vesicles since they possessed the defining characteristics of exosomes. 20

Characterization of ELV-AT.
ELV-AT promoted proliferation and migration of endothelial cells
At a concentration of 50 μg/mL, ELV-AT treatment increased the number of migrated cells (Fig. 2A), which was threefold compared to the control group (Fig. 2B). However, the migration capacity of ECs decreased when cells were treated with high concentrations of ELV-AT (200 μg/mL). Likewise, the proliferation of ECs determined by CCK-8 showed an increase in a dose-dependent manner in the groups of 10 and 50 μg/mL ELV-AT but a decrease in the group of 200 μg/mL. Especially on day 3, ELV-AT at a concentration of 50 μg/mL significantly affect the proliferation of ECs.
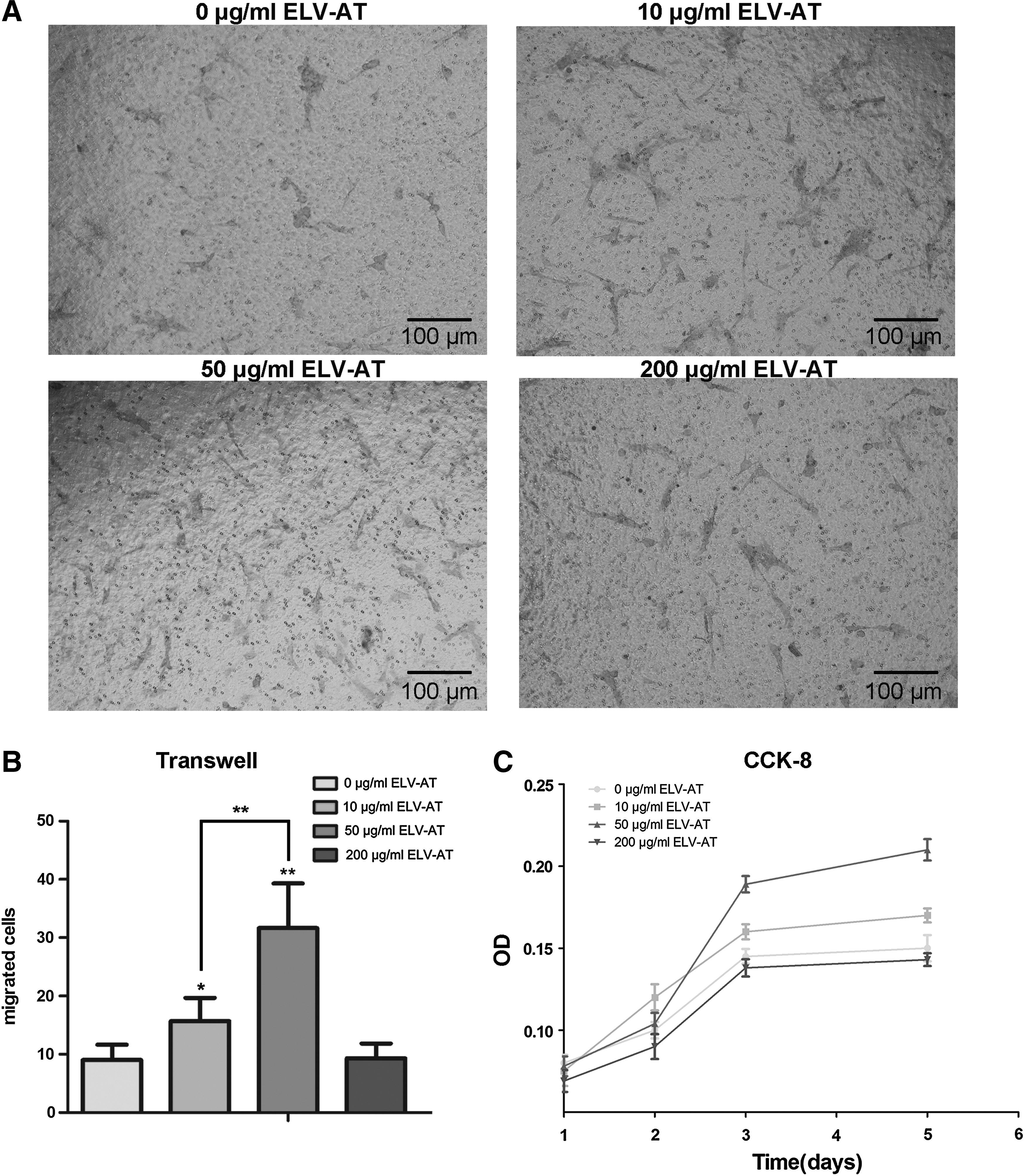
Migration and proliferation of rat aorta ECs enhanced by ELV-AT.
ELV-AT induced angiogenesis and adipogenesis in vitro
Angiogenic and adipogenic effect of ELV-AT was evaluated on ECs and ADSCs, respectively (Fig. 3). After cocultured with 50 μg/mL ELV-AT, the total number and length of all tubing within each field in ECs were twofold higher than that in the control group (Fig. 3A, B). The expression of angiogenesis markers (VEGF, CD31, and angiogenin) was also upregulated in ELV-AT-treated cells with angiogenin expression increased by threefold (Fig. 3E).
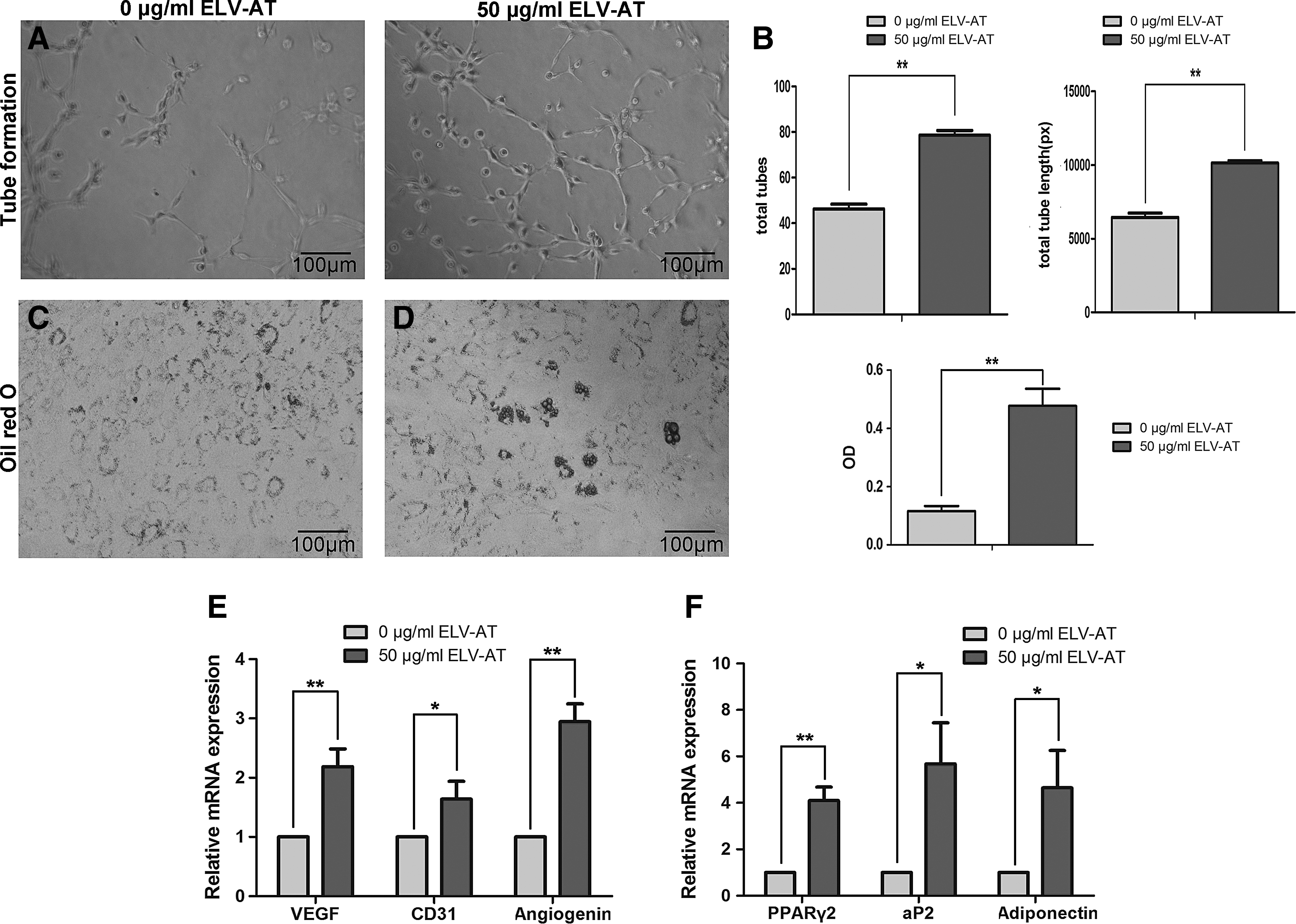
Angiogenic and adipogenic differentiation enhanced by ELV-AT. ECs were cocultured with 50 μg/mL ELV-AT before the analysis of angiogenic capacity.
Lipid droplets showed at 14 days when ADSCs were treated with 50 μg/mL ELV-AT (Fig. 3C). The expression of adipogenic genes at 5 days showed that the expression of PPARγ2 in 50 μg/mL ELV-AT group was fivefold higher than that in the control group, along with increased expression of aP2 and adiponectin.
Adipose tissue regeneration induced by ELV-AT in vivo
Matrigel mixed with ELV-AT was injected into the silicone tube at the back of nude mice, and Matrigel alone served as the control (Fig. 4). In the early stages, the gross appearance revealed an active angiogenesis in the ELV-AT group with growth of vessels from host into the tubes, while in control group, tubes filled with Matrigel were wrapped by a fibrous capsule (Fig. 5A). In the ELV-AT group, immature adipocytes (stained with adipocyte marker, perilipin A) with smaller diameters comparable to the endogenous adipocytes started to show up after 2 weeks of injection (Fig. 5A), a significant increase of newly formed vessels and adipocytes and loosened tissues were detected at 4 weeks (Fig. 5B). In the control group, Matrigel almost has been completely absorbed at 4 weeks (Fig. 5B). By the end of 8 weeks, the control group showed fibrous capsule alone because of the complete absorption of Matrigel (Fig. 6A). The adipose neotissue gained a further increase by 8 weeks in the ELV-AT group. However, there was no significant difference of the neotissue between 8 and 12 weeks (Fig. 6B). Maturity of the adipose neotissue and a long-term stability of the adipose neotissue were further confirmed by comparing the structure of the adipose neotissue with endogenous adipose tissue. At 8 and 12 weeks, the mean size of neoadipocytes reached the level of endogenous adipocytes (Fig. 6C, E). In addition, staining of CD34, a marker of endothelial cells, showed that the structures of the neoadipose tissues were supplied with well-organized blood vessels, similar to that of the endogenous one (Fig. 6D).

The nude mice chamber model.
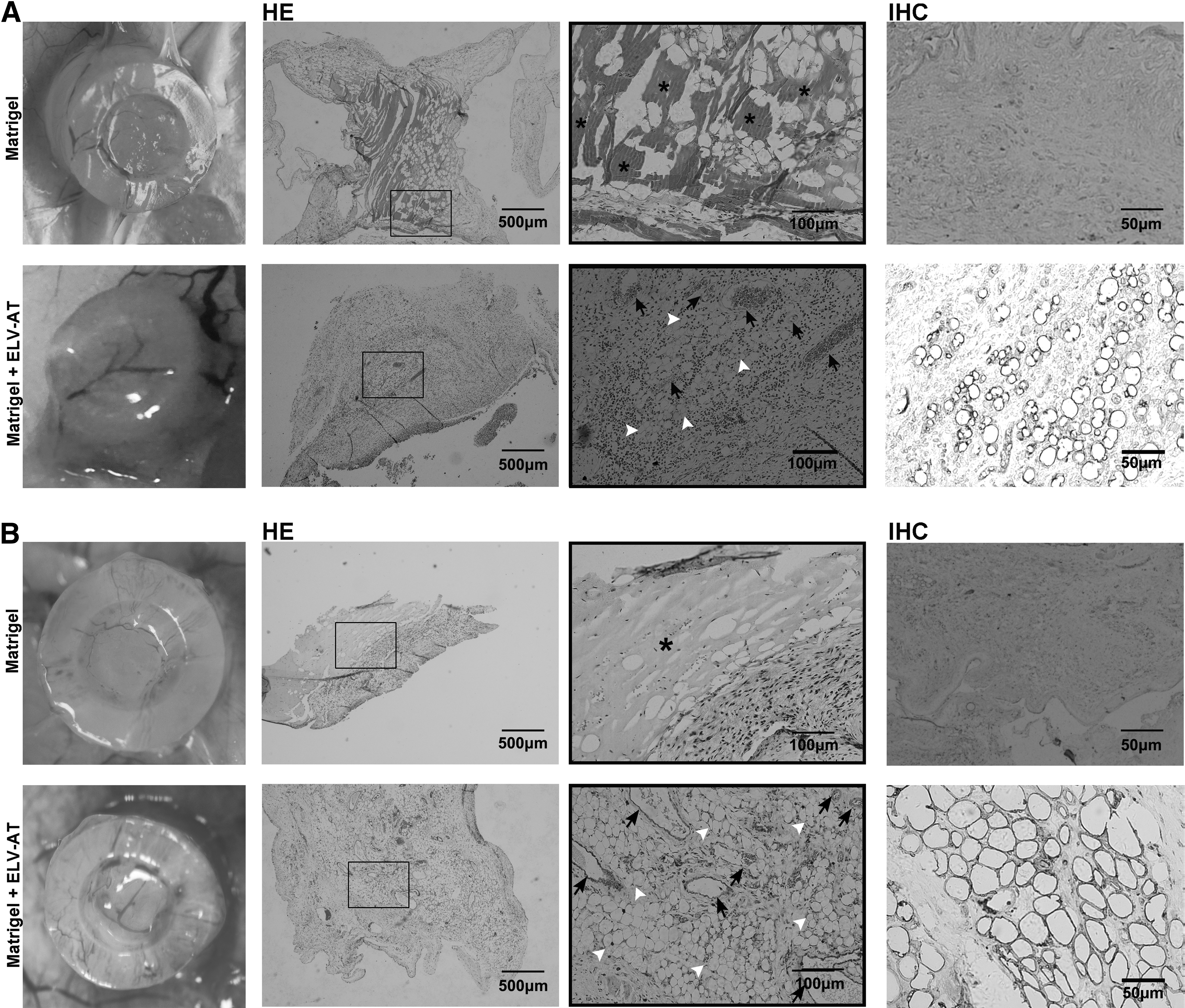
In vivo studies of ELV-AT adipogenic capacity at the early points (2 and 4 weeks, five mice at each time point). Analysis of adipose neotissue generated in groups treated with and without ELV-AT at

In vivo studies of ELV-AT adipogenic capacity and adipose neotissue long-term stability at 8 and 12 weeks (five mice at each time point).
Discussion
Studies using synthetic or natural biomaterial scaffolds loaded with cells and/or growth factors had successfully obtained adipose tissue regeneration.1,4,21,22 However, problems such as foreign body reactions4,23–25 make it difficult to apply in clinic extensively. Another strategy of tissue engineering is to induce de novo adipogenesis by providing a microenvironment suitable for originally existing cells in the body to migrate, proliferate, and differentiate to form adipose tissues, which avoid the transplantation of cells.
Exosomes can modulate proliferation, migration, and differentiation of the target cells by transferring proteins, mRNAs, and miRNAs.26–30 For example, matrix metalloproteinases were found in exosomes 31 and transferred into 3T3L1 and primary rat preadipocytes to promote adipogenesis. 32 Yamada et al. found that MFG-E8, an exosomal protein, 33 can increase the expression of VEGF and ET-1 in MSC and induce angiogenesis. 34 Reduction of miR-126 levels in MSCs led to the impairment of angiogenic ability through the upregulation of Spred 1 and inhibition of mitogen-activated protein kinases pathway. 35 Delivery of miRNA-125b contained in neuronal cell line-derived exosomes can induce neuronal differentiation of MSCs. 36 Our results showed that ELV-AT significantly increased proliferation and migration ability of ECs at 50 μg/mL (Fig. 2). However, the mechanism underlying the phenomenon that high concentration (200 μg/mL) of ELV-AT showed a reverse effect remained elusive. One of the possible explanations might be that the cells are overloaded with proteins and nucleic acids in the exosome. Coculturing with ELV-AT rapidly enhanced the angiogenic and adipogenic ability of the cells at early time points (Fig. 3). Ahabbit et al. showed that the exosomes could activate Akt, Erk 1/2, and STAT3 pathways in target fibroblast and increase their angiogenic response. 37 And for adipogenesis, exosomes can positively regulate the terminal differentiation of adipocyte lineage through pathways such as MAP2K5-ERK5. 38 Besides, exosomes could induce pro-osteogenic effects on MSCs through the activation of PI3K/AKT pathway.39,40 To conclude, from a perspective of paracrine effect, ELV-AT can be delivered into target cells and give rise to angiogenesis and adipogenesis ultimately. The exact proteins or miRNAs involved in ELV-AT-mediated differentiation are still needed to be determined in the future.
We further investigated the potential effects of ELV-AT in vivo over a long-term period of 12 weeks, which is sufficient for complete adipogenesis and neotissue stabilization.41,42 We created silicone-support structures to provide a mechanically stable space capable of bearing the compressive forces in vivo for volume-stable adipose tissue formation. The neotissue formation began from 2 weeks with an infiltration of host vessels into the tubes; the progressive development was reflected by the increase of adipose tissue areas, along with the maturity of the neotissues and replacement of Matrigel by neovascularization. The adipocytes were first detected at 2 weeks and mainly appeared around the newly formed blood vessels (Fig. 5A). Due to the close relationship between angiogenesis and adipogenesis,43,44 we found a positive correlation between adipocyte density and capillary density at 4 weeks (Fig. 5).
It has been shown that the increased resorption of a fat graft is usually related to debilitated neovascularization. 45 Therefore, the mature blood vessels (stained with CD34) on 8 weeks (Fig. 6D) ensure that regenerated adipose tissue could maintain till 12 weeks.
To summarize, our in vivo studies demonstrated that ELV-AT suspension in Matrigel could be an efficient and biocompatible system. It expanded the application ranges of engineering adipose tissue. This cell-free system will benefit from the time-saving isolation procedures, long-term storage at −80°C, and the less potential risk of tumor formation of cell therapy. However, as the possible contamination such as free-floating factors were coprecipitated by the isolation kit, our results need to be further proved with purer exosome samples isolated by developed techniques to exclude the potential contaminants. Recent reports showed that EV number, RNA content, or miRNA levels decrease after two cycles of freezing and thawing, 46 and EVs can be degraded by proteases. 47 So, an optimized preservation method needs to be established to guarantee further application of exosomes. In this study, we only demonstrated the effectiveness of ELV-AT by adding them into a culture system, and further studies to ablate the components of the ELV-AT, such as protein denaturation, RNase digestion, or high-temperature incubation, can also be applied to decode ELV-AT. The study to identify the cell types that were homed to the preset sites is expected as well to expand our understanding on the role of exosome in cell–cell communication.
Conclusion
In this study, we demonstrate the angiogenic and adipogenic potential of ELV-AT both in vitro and in vivo for the first time. ELV-AT could promote proliferation, migration, and angiogenesis of ECs and induce adipogenesis of ADSCs in vitro. The in vivo transplantation results indicated that ELV-AT could effectively induce adipose tissue regeneration and that the neotissue could maintain viability for 12 weeks. These results demonstrated that ELV-AT could serve as an alternative approach for generating adipose tissue, providing a promising application prospect in reconstructive surgeries.
Footnotes
Acknowledgments
This work was supported by the National Nature Science Foundation of China (81300848) and the National Key Research and Development Program of China (2016YFC1101400).
Disclosure Statement
No competing financial interests exist.