
Abstract
Endothelial progenitor cell (EPC)-based therapy has immense potential to promote cardiac neovascularization and attenuate ischemic injury. Functional benefits of EPCs and other adult stem cell therapies largely involve paracrine mechanisms and exosomes secreted by stem cells are emerging as pivotal paracrine entity of stem/progenitor cells. However, modest outcomes after EPC-/stem cell-based clinical trials suggest that stem cell/exosome function might be modulated by stimuli they encounter in ischemic tissues, including systemic inflammation. We hypothesized that EPCs under inflammatory stress might produce exosomes of altered and dysfunctional content, which may compromise EPC repair in ischemic heart disease. We have previously shown that EPCs obtained from interleukin-10 knockout (IL-10KO) mice (model mimicking systemic inflammation) display impaired angiogenic functions. Whether IL-10KO-EPC-derived exosomes inherit their parental dysfunctional phenotype and whether inflammatory environment alters the cargo of their secreted exosomes are not known. After cell expansion from IL-10KO and wild-type (WT) mice, we isolated exosomes and compared their functions in terms of effect on cell survival, proliferation, migration, and angiogenic capacity in vitro. WT-EPC-Exo treatment enhanced endothelial cell proliferation and tube formation, and inhibited apoptosis, whereas IL-10KO-Exo exhibited impaired or even detrimental effects, suggesting that the reparative capacity of WT-EPC-Exo is lost in exosomes derived from IL-10-KO-EPCs. Deep RNA sequencing and proteomic analyses to compare WT and IL-10KO-Exo revealed drastically altered exosome cargo. Importantly, IL-10KO-EPC-Exo were highly enriched in microRNAs and proteins that promote inflammation and apoptosis and inhibit angiogenesis. Through modulation of a specific enriched miRNA (miR-375), we partially rescued IL-10KO-EPC-Exo dysfunction. Thus, our study revealed that EPC exosomes display impaired function under inflammatory stimulus through changed exosome contents, and the dysfunction can be rescued by modulation of a specific target packed in exosomes.
Introduction
S
Exosomes are microvesicles secreted by various cells and have been the subject of intense research in recent times. The function of exosomes is not fully known, but they are important for intercellular communication 6 and immune modulation, 7 and as biomarkers for disease progression. 8 Recent reports, including those from our laboratory, have suggested that exosomes secreted by stem cells can induce angiogenesis as effectively as their cell of origin and can enhance postinjury cardiac repair.9,10 Exosome-mediated repair occurs largely by transfer of exosome cargo, including parent cell-specific microRNA (miR), mRNA, and protein, to the target cell/tissue resulting in protection.11,12 However, the exact molecular mechanisms of exosome action are poorly understood and may differ depending upon parental cell population, exosome content, and modulation of those contents by various stimuli.
Systemic inflammation, a major symptom in patients with cardiac disease, is an important stimulus causing EPC functional deficiency.4,13 It is well established that stem cells obtained from patients with systemic inflammation have vastly reduced reparative activity.13,14 We have previously established the cardioprotective role of anti-inflammatory cytokine interleukin-10 (IL-10) in mouse models of acute myocardial infarction (MI) and pressure-overloaded myocardium.15,16 Using IL-10 knockout (IL-10KO) mice as a model of systemic inflammation, we reported that EPCs from IL-10KO mice have diminished ability to facilitate angiogenesis and post-MI recovery.15,16 Since the therapeutic efficiency of EPCs is largely due to paracrine effects, namely by exosomes, it is reasonable to predict that EPCs under inflammatory stimuli secrete dysfunctional exosomes with altered exosome contents, and ultimately compromise exosome reparative ability in ischemic cardiac tissue.
In this study, we provide evidence that EPCs under inflammatory conditions (IL-10KO EPCs) secrete dysfunctional exosomes with altered exosome contents (RNAs and proteins). We reveal that modulation of identified targets in IL-10KO EPC exosome cargo may enhance and/or rescue reparative properties of dysfunctional exosomes.
Materials and Methods
Vertebrate animals and reagents
All experiments conform to the protocols approved by the Institutional Animal Care and Use Committee at Temple University. Eight-week-old wild-type (WT) and IL-10 knockout (KO; IL10tm1Cgn) male mice of C57BL/6J background were procured from Jackson Research Laboratory (Bar Harbor, ME). Lipopolysaccharide (LPS) was obtained from Sigma-Aldrich, Inc. (St. Louis, MO).
Cell isolation, culture, and characterization
EPC isolation, ex vivo expansion, and culture were performed as previously described.
1
Bone marrow extracted from the femurs, tibiae, and hip bones of 8–10-week-old C57BL/6J or IL-10KO mice were stained with labeled antibodies against lineage markers [CD3e (145-2C11), CD11b (M1/70), B220 (RA3-6B2), Ter119 (Ly76), Ly6G/C (RB6-8C5) Sca-1 (D7), and CD31 (MEC13.3)] and then sorted to retain lineage-negative cells on a FACS Aria-II (BD Biosciences). Sorted cells were cultured in phenol red-free endothelial cell basal medium-2 (EBM-2, Clonetics) supplemented with exosome-free 10% fetal bovine serum (FBS), vascular endothelial growth factor-A, fibroblast growth factor-2, epidermal growth factor, insulin-like growth factor-1, ascorbic acid, and antibiotics. Cells were cultured at 37°C with 5% CO2 in a humidified atmosphere. After 4 days in culture, nonadherent cells were removed by washing with phosphate-buffered saline
Exosome isolation and characterization
EPCs (WT and IL-10KO) become 80–90% confluent after 7 days of culture; cell-conditioned media are collected daily from day 7 to 10 for exosome isolation. The exosome isolation, purification, and storage were performed as previously described. 17 Collected exosomes were suspended in PBS, aliquoted, and stored at −80°C, or used immediately for experiments. The size and shape of exosomes were observed by electron microscopy supported by Electron Microscopy Resource Laboratory (EMRL) at University of Pennsylvania. Exosomes docking on Dynabeads (Sigma) coated with anti-mouse CD63 and CD81 were used to check exosome surface markers by flow cytometry. Three exosome markers, including HSP70 [HSP 70/HSC 70 Antibody (W27): sc-24], CD63 [CD63 Antibody (H-193): sc-15363], and Flotillin-1 [Flotillin-1 Antibody (K-19): sc-16640], were verified by Western blotting with antibodies from Santa Cruz. The exosome protein level was quantified with the Pierce BCA Protein Assay Kit (Thermo Fisher) as per manufacturer's instructions. Exosome particle number was quantified by NanoSight running Nanoparticle Tracking Analysis (NTA) software. The RNAs and proteins in exosomes were labeled and tracked by Exo-Glow (SBI) as per manufacturer's instructions to check the delivery function of exosomes.
Migration assay
The migration of SVECs toward a gradient of growth factors was performed in a 24-well Transwell Chamber (8.0 μm pore size, polycarbonate membrane, Corning Costar; Corning Incorporated Life Sciences, Acton, MA) as described earlier. 15 Briefly, SVEC culture media (DMEM with 10% exosome-depleted FBS; Life Technologies) were added to the lower compartment. SVECs (1 × 105 cells) in 200 μL of OPTIMEM (Thermo Fisher Scientific) were added to the upper compartment and simultaneously treated with 10 μg WT-EPC-Exo, IL-10KO-EPC-Exo, or 20 μL of PBS. After incubation at 37°C for 18 h, the filters were removed, and the cells that migrated through the pores of the membrane to the bottom chamber were stained with Giemsa stain (Sigma) and counted manually under phase contrast microscopy (Microscope model and manufacturer; × 10 magnification) in each well. Data are expressed as average number of cells per well that migrated through or invaded pores. All groups were studied, at least, in duplicate.
Tube formation assay
4 × 104 HUVECs were plated on 130 μL Matrigel (BD Falcon) per well in a 48-well plate and simultaneously treated with 5 μg of WT-EPC-Exo, IL-10KO-EPC-Exo, or 10 μL of PBS. After incubation at 37°C in an atmosphere of 5% CO2 for 16 h, gels were observed by using a phase contrast microscope ( × 10; model & manufacturer). The total branch points were counted in each well. Then, the average branch points per well in WT and IL-10KO groups were normalized against the PBS control group (whose value was set to 1). Results are represented as mean ± s.e.m. for three independent experiments.
Terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling staining assay
SVECs were seeded in chamber slides (Thermo Fisher Scientific) and simultaneously treated with 10 μg of WT-EPC-Exo, IL-10KO-EPC-Exo, miR-375-modified EPC-Exo, or 20 μL of PBS. After 16 h of culture, cells were stimulated with 100 ng/mL LPS or 100 nM H2O2 for 24 h. Cells were then washed with PBS and fixed with 4% paraformaldehyde (PFA) fixative. Survival of SVECs was determined by terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling (TUNEL) staining as per manufacturer's instructions (Cell death detection assay; Roche, Indianapolis, IN). DAPI staining was used to count the total number of nuclei. Apoptosis of SVECs was assessed by counting the total number of TUNEL+ cells and DAPI+ cells in each slide and calculated as percentage of TUNEL+/DAPI+ per slide.
Exosome RNA and protein content analysis
WT- and IL-10KO-EPC-derived exosomes (100 μg) in PBS (duplicates in each group) were sent to SBI for Exo-RNA Next-Gen Sequencing as per manufacturer's instructions. Deep sequencing data were analyzed with Maverix software provided by SBI. Fifty microgram of WT- and IL-10KO-EPC-derived exosomes in PBS was sent to Quantitative Proteomics Resource Core (QPRC) in U Penn for mass spectrometry to characterize and quantify proteins from exosomes.
Real-time polymerase chain reaction
Expression levels of miR-375 were measured using quantitative miRNA stem loop reverse transcription-polymerase chain reaction (RT-PCR) technology (TaqMan miRNA assays; Applied Biosystems). This assay uses gene-specific stem loop RT primers and Taq-Man probes to detect mRNA or mature miRNA transcripts. Transcription was performed using 2 μg or 10 ng total RNA and the TaqMan miRNA RT kit (Applied Biosystems). Real-Time PCR was performed on an Applied Biosystems 770 Step One Plus using the TaqMan Universal PCR Master Mix, No AmpErase UNG (Applied Biosystems). The amplification steps consisted of initial denaturation at 95°C, followed by 40 cycles of denaturation at 95°C for 15 s and annealing at 60°C for 1 min. The TaqMan specific primer 18S or U6 small nucleolar RNA was used for normalization with the threshold delta-delta cycle method [Gene Expression Macro; Bio-Rad, Hercules, CA].
miR-375 modulation in exosomes
Thirty nanomolar anti-miR-375 (Applied Biosystems) or a negative control (siRNA-NC; Applied Biosystems) was introduced using Lipofectamine™ RNAiMAX (Invitrogen) in IL-10KO-EPCs following the manufacturer's instruction, and the cells were transfected for 24 h. Subsequently, the knockdown efficiency in EPCs was determined by RT-PCR assays.
Statistical analyses
Data are presented as mean ± SE between two groups; unpaired Student's t-test determined significance. For >2 groups, ANOVA with Tukey post hoc was used. p values of <0.05, <0.01, and <0.001 were considered significant.
Results
Characterization of EPCs from WT and IL-10KO C57BL/6J mice
EPCs were isolated from WT and IL-10KO mouse bone marrows by FACS of lineage-negative (lin−), Sca-1+/CD31+ double-positive cells, which represent ∼4% of total mononuclear cells in BM of mice of both strains (Fig. 1A). These cells are referred to as EPCs henceforth, and they showed phenotypic characteristics consistent with their spindle-like morphology (Fig. 1B).

Characterization of EPCs from C57BL/6J mouse bone marrow:
Characterization of EPC-derived exosomes
We harvested exosomes from the conditioned media collected from cultured WT and IL-10KO EPCs by differential centrifugation with sucrose cushion. 17 Electron microscopy and NanoSight analysis of WT and IL-10KO EPCs exosomes exhibited a range of 60–100 nm particles, which is in accordance with the previously reported exosome size distributions 17 (Fig. 2A, B). In addition, Western blotting and flow cytometry indicated that these exosomes were positive for the characteristic exosome surface marker proteins, including HSP70, flotillin-1, CD63, and CD81 (Fig. 2C–E). Next, we determined EPC exosome uptake by SVECs. Cellular uptake and the delivery of both exosome RNA and exosome protein were confirmed using the Exo-Glow kit, which exclusively labels exosome RNA with Acridine Orange (red) and proteins with Carboxyfluorescein succinimidyl diacetate ester (green). Exosomes were internalized by cells (as early as 2–30 min after addition to the culture) and both RNA and protein were delivered to the target SVECs (Fig. 2F, G).

Characterization of EPC-derived exosomes:
IL-10 deficiency impairs EPC-derived exosome function in vitro
Since our published and preliminary data demonstrated functional impairment in EPCs obtained from IL-10KO mice,15,16 we next determined if exosomes isolated from these EPCs exhibited the same impairment as their parental cells. We performed TUNEL and tube formation and cell migration assays with endothelial cells (HUVECs or SVECs) treated with exosomes collected from WT or IL-10KO EPCs. Our results showed that WT-EPC-Exo treatment significantly decreased LPS-induced endothelial cell apoptosis, evident by decreased TUNEL+/DAPI+ cells, whereas IL-10KO EPC-derived exosomes showed decreased antiapoptotic effects (Fig. 3A, B) compared to untreated controls. Furthermore, in the in vitro angiogenesis assay, IL-10-KO-EPC-Exo strongly inhibited the ability of HUVECs to form tubular structures on Matrigel compared to WT-EPC-Exo and PBS treatments (Fig. 3C, D). Both WT and IL-10KO-EPC-Exo treatment can dramatically increase the number of migrating cells, but ability to promote cell migration was significantly decreased in the IL-10KO-EPC-Exo-treated group (Fig. 3E, F). Our data revealed that exosomes reflect their parental cell dysfunction: under IL-10 deficiency, EPC-derived exosomes are impaired in the functions of inhibiting endothelial cell death, promoting angiogenesis, and promoting cell migration in vitro.
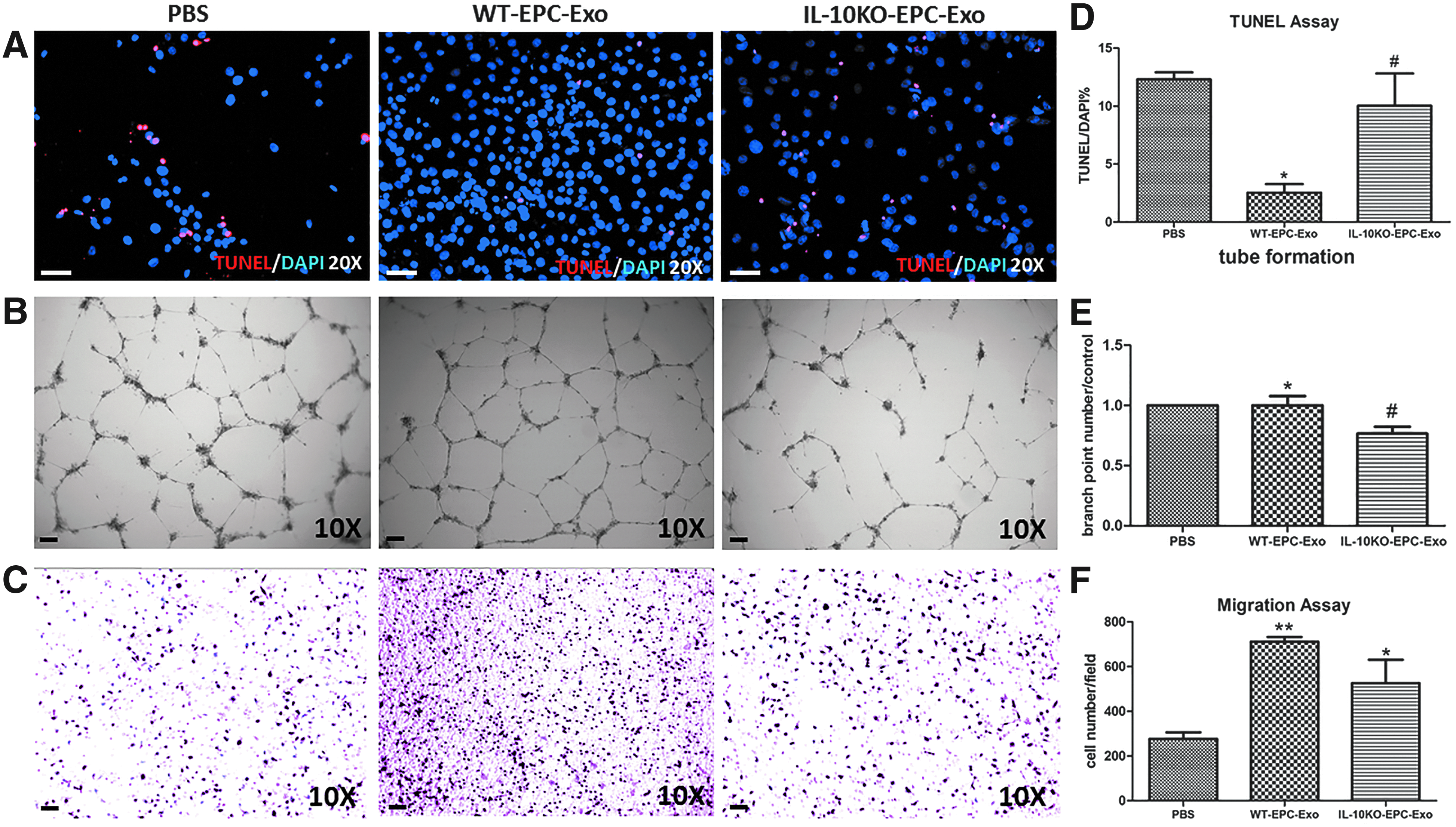
IL-10 deficiency impairs EPC-Exo functions:
IL-10 deficiency drastically modifies their EPC exosome contents
Several studies implicate that cells can exert functional effects over long distances by packaging RNA and protein in exosomes. 11 Therefore, we hypothesized that the functional differences in paracrine effects from WT or IL-10KO EPCs might be due to altered exosome content. Exosomes isolated from WT or IL-10KO EPCs were subjected to deep sequencing (SBI) as well as mass spectrometric analysis to reveal exosome contents. Our deep sequencing data identified various RNAs packaged in exosomes suggesting linkage to specific functions (Fig. 4A). We focused on those miRs that were differentially expressed between the two groups of exosomes, derived from EPCs of WT versus IL-10KO mice (Fig. 4B). IL-10KO-EPC-Exo are enriched with miRs involved in apoptosis, antiangiogenesis, and inflammation. Comparison of protein content between the two groups of exosomes also demonstrated great difference paralleling the RNA analysis (Fig. 5A). IL-10KO-EPC-Exo were enriched in proteins with proinflammatory, apoptotic, and anti-angiogenic functions (Fig. 5B). These data provide evidence that exosome contents determine the functions mediated by exosomes and these contents are dramatically affected by the status of their cells of origin.
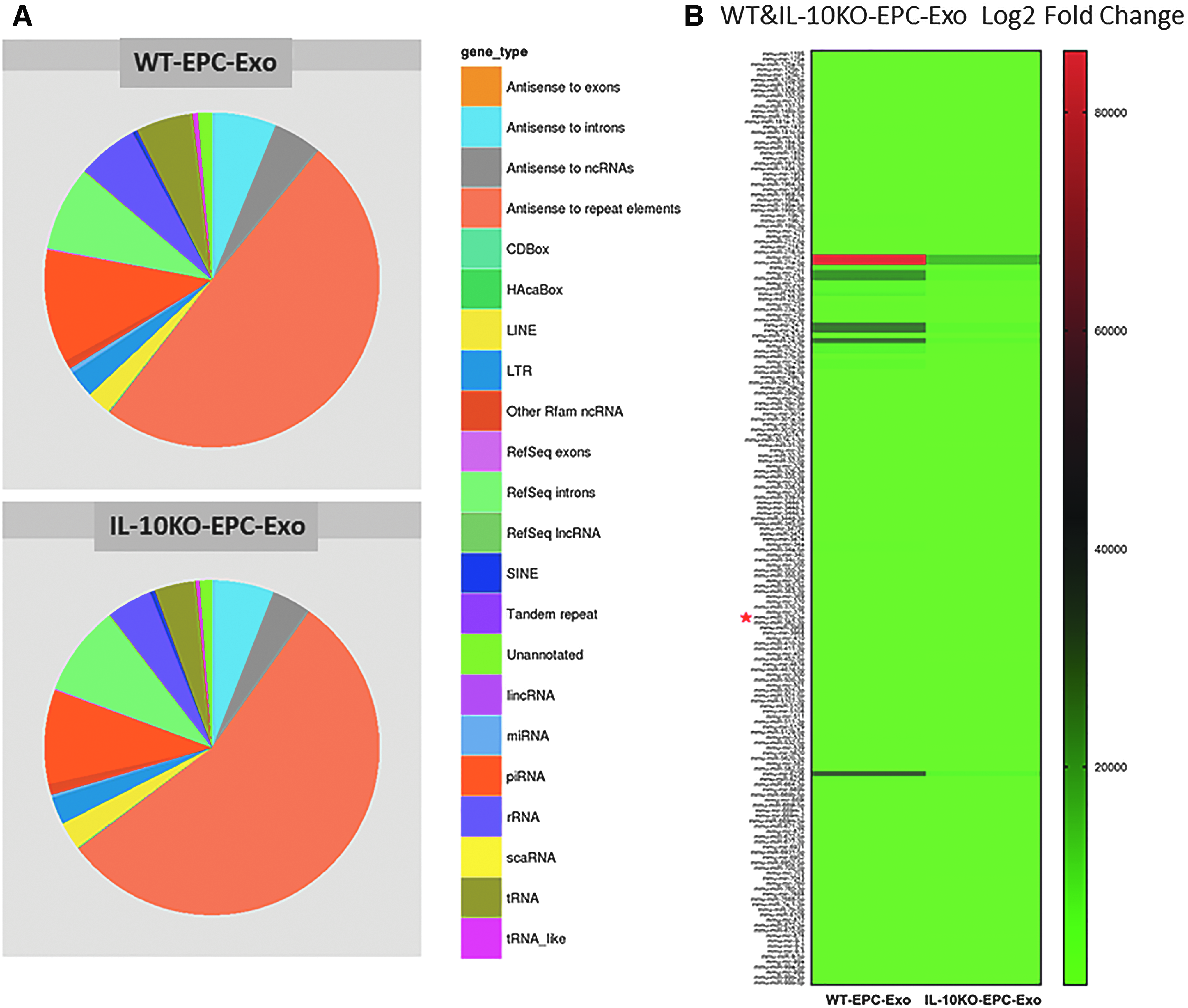
EPC phenotype modifies exosomal RNA profile: Equal amount of RNA from WT and IL-10KO EPC-derived exosomes were processed for deep sequencing by SBI and the results were analyzed by Maverix software.
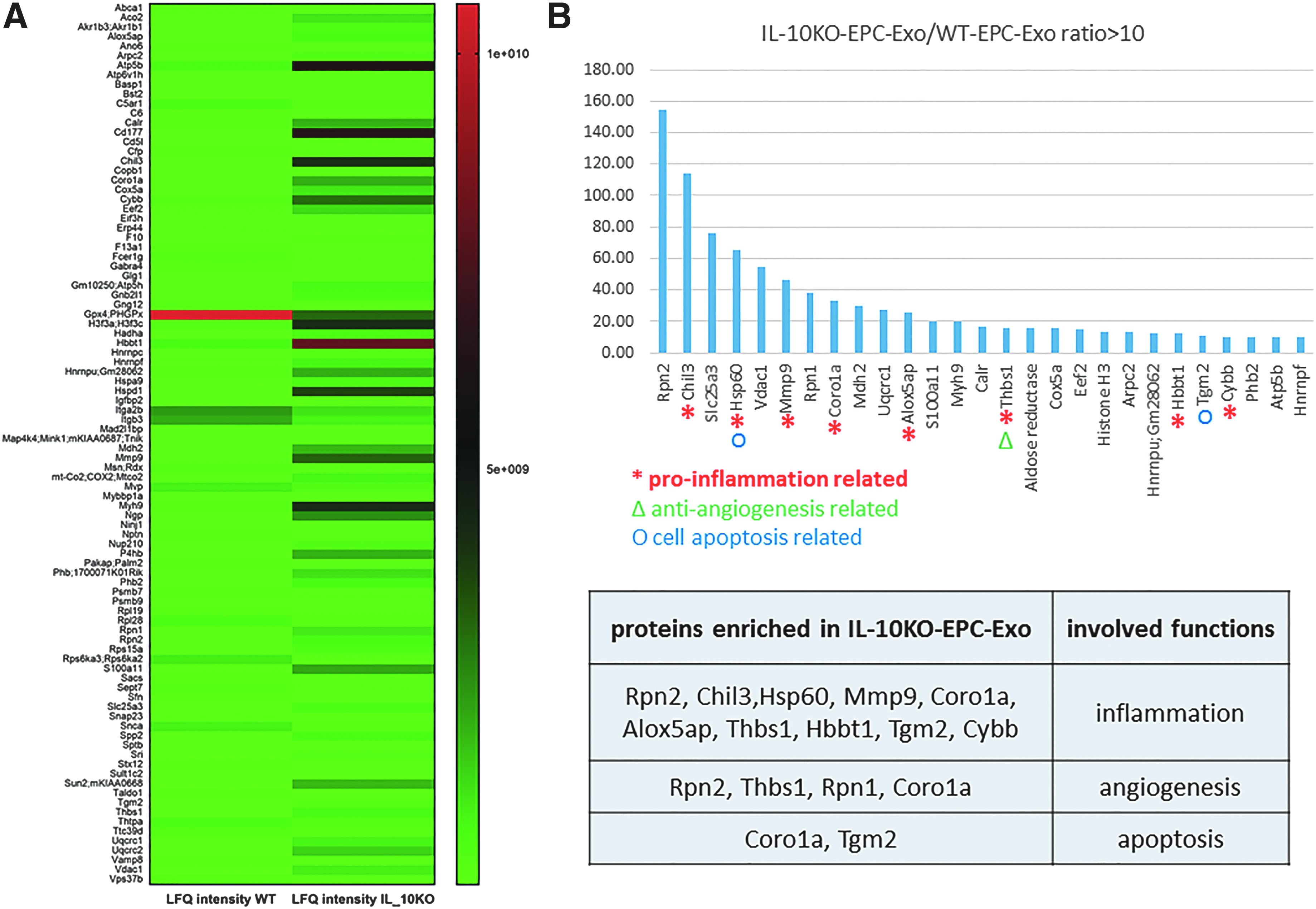
EPC phenotype modifies their protein profiling in exosomes: equal amount of protein from WT and IL-10KO EPC exosomes were processed for mass spectrometry analysis.
IL-10KO-EPC-Exo dysfunction is partly mediated by miR-375
We have recently published that untreated IL-10KO-EPCs and LPS-treated WT-EPCs exhibit significantly higher levels of miR-375 compared to untreated WT EPCs. 18 Inhibition of miR-375 in the IL-10KO-EPCs rescues their defective phenotype by enhancing the expression of PDK1 (a direct target of miR-375) and PDK1-mediated activation of PI3K/Akt pathway. 18 To establish that enrichment of stimulus-specific miRNA governs functional properties of exosomes, we assessed miR-375 (significantly upregulated in IL-10KO-EPCs) as a potential mediator. To modulate miR-375 levels in IL-10KO-EPC-Exo, IL-10KO-EPCs were transfected with miR-375 antagomirs before exosomes were isolated. Quantitative RT-PCR analysis of exosomes confirmed that inhibition of miR-375 in IL-10KO-EPCs significantly diminished miR-375 expression in IL-10KO-EPC-Exo. (Fig. 6A, B). Interestingly, experiments on cell survival (H2O2 treatment in the presence of miR-375-unmodified and miR-375-modified exosomes) showed that modulation of miR-375 in the IL-10KO-EPC-Exo partly rescued their cell protection activity (Fig. 6C–F). These exciting data suggest that cell-specific dysfunction observed in exosomes can be rescued.

IL-10KO-EPC-Exo dysfunctions are partly mediated by miR-375:
Discussion
Over the past few decades, stem cell-based therapy has emerged as a promising therapeutic approach for ischemic cardiac repair and regeneration. 19 Endothelial progenitor cells derived from bone marrow or peripheral blood have been tested in clinical studies. However, myocardial infarction, the leading cause of death in U.S. patients, triggers an intense immune response and greatly decreases the therapeutic effects of EPC transplantation. 13 Our published studies have established that EPCs are functionally impaired under inflammatory stimulus. Using IL-10KO mice as a model of systemic inflammation, we reported that EPCs from IL-10KO mice have a diminished in vitro angiogenic activity; when transplanted in vivo in an MI mouse model, IL-10KO EPCs do not activate angiogenic or protective responses and do not elevate postinjury left ventricle (LV) function. 20 Recent studies revealed that functional benefits of stem cell-based therapy are largely mediated by paracrine effects.10,21,22 We hypothesized that loss of IL-10 modifies EPC-Exo contents and further compromises reparative properties of exosomes. Our conclusions are based on the following experimental evidence: (1) IL-10 deficiency impairs EPC-derived exosome functions in vitro; (2) EPC phenotype drastically modifies their exosome content (both RNA and protein); (3) miR-375 levels are elevated in IL-10KO EPC-Exo; and (4) modulation of miR-375 in the IL-10KO-EPC-Exo partly rescues endothelial cell protection ability.
Exosomes are the major functional units among stem cell-secreted paracrine factors. Exosomes have been reported to play a vital role in tissue repair by packaging and delivering RNA and protein and are gaining importance as an alternative approach for cell-based therapy. Our recently published study showed that exosomes derived from mouse embryonic stem cells enhanced neovascularization and cardiomyocyte survival, and reduced fibrosis postinfarction, consistent with a resurgence of the cardiac proliferative response. 9 One clinical study demonstrated that exosomes derived from human umbilical cord blood-derived EPCs possessed robust proangiogenic and wound-healing effects in both vascular injury and diabetic rat models. 23 However, exosomes are not all created equal and the constitution of their contents as well as their biological activity are largely dependent upon the physiological state of the parental cell of origin. 24 In this study, we used IL-10KO mice mimicking a systemic inflammation state, and compared the ability of exosomes derived from WT or IL-10KO EPCs to modulate endothelial cell function. We report that compared to WT EPC-derived exosomes, IL-10KO EPC-derived exosomes significantly decrease the angiogenic activity of HUVECs, greatly inhibit the mobilization of SVECs, and fail to prevent LPS-induced apoptosis. This evidence suggests a dysfunction of IL-10KO EPC exosomes in ischemic tissue repair, which is consistent with the dysfunction observed in IL-10KO EPCs.
The mechanisms of exosome-mediated repair are largely due to transfer of exosome cargo, including parent cell-specific microRNAs (miRs) and proteins to the target cell/tissue resulting in cell protection.11,12 Many studies have characterized exosome cargo using next-generation sequencing and mass spectrometry techniques, and found that exosome contents have a tight relationship with cancer and autoimmune diseases, revealing the role of exosome as biomarkers or signatures for typical disease diagnosis.11,22,25,26 One recent study compared the proteomic profiles of secretomes derived from human neonatal and adult cardiac progenitor cells and identified hundreds of proteins targeting myocardial recovery post-MI, suggesting an essential role for functional components of exosome. 22 In our study, we performed deep sequencing and proteomic analysis. Compared to the WT, IL-10KO EPC exosomes are enriched in proteins and miRs that mediate apoptosis, antiangiogenesis, and proinflammatory responses, including elevated miR-375 levels. We have recently published that EPCs derived from IL-10KO mice (a model of inflammation) are defective in their angiogenic and post-MI reparative activities, which are partially mediated by the miR-375/PDK-1 signaling axis. 18 In this study, an increased expression of miR-375 in exosomes from IL-10KO mouse EPCs can be associated with increased miR-375 levels in IL-10KO EPCs. 18 Furthermore, we have shown that knocking down miR-375 in IL-10KO-EPCs reduces miR-375 levels in exosomes derived from those EPCs and ultimately rescues the antiapoptotic effects of IL-10KO-EPC-Exo. These findings are very important and provide novel information for possible rescue and/or alteration of these exosomes through modulation of specific targets in exosomes. This approach proves the possibility and provides expectations toward designing “super-exosome” for possible clinical applications.
Conclusion
In summary, our observations demonstrate that IL-10 deficiency impairs EPC-derived exosome function in vitro. The alteration of function is largely due to changes in RNA and protein contents in exosomes. Thus, modulation of specific targets in exosome cargo is a feasible approach to enhance or rescue exosome function, which might prove to be an attractive therapeutic strategy for patients with MI.
Footnotes
Acknowledgments
This work was supported, in part, by National Institutes of Health grants HL091983, HL126186, and HL053354. Y.Y. is supported by American Heart Association predoctoral grant 17PRE33370001. V.N.S.G is supported by American Heart Association postdoctoral grant 15POST22720022 and S.V. is supported by AHA Scientist Development Grant 14SDG20480104. D.A.G. is supported with AHA—Grant in aid 17GRNT33400035.
Disclosure Statement
No competing financial interests exist.