
Abstract
Myogenic precursors sense and dynamically respond to mechanical stimulation through complex integrin-mediated mechanotransduction, in which focal adhesion kinase (FAK) is a fundamental intracellular signaling mediator. When skeletal myoblasts are exposed to uniaxial cyclic tensile strain (UCTS), they display uniform alignment and an enhanced rate of differentiation. In this work, we explored the role of FAK activation by using C2C12 myoblasts that were grown on flexible culture plates and exposed to UCTS during the early differentiation phase. After 24 h, the cells oriented perpendicularly to the direction of strain and exhibited an enhanced differentiation profile. Next, the cells were exposed to a strain field that was either kept in the same direction or rotated 90°, in the presence or not of an FAK phosphorylation inhibitor. On reorientation of the strain field by 90°, the cells reassembled their focal adhesions and actin cytoskeleton to regain the perpendicular position with respect to the engaging stress. After blocking the FAK, however, the cells failed to respond to the reoriented strain field and their differentiation was abrogated. Interestingly, when the strain field remained in the same direction, the FAK inhibitor compromised the differentiation, even though there was no evident change in cell orientation. Our data indicate that during exposure to UCTS, the activation of FAK is necessary for the myoblasts to undergo alignment and enhanced differentiation.
Introduction
D
Further, an enhanced understanding of the mechanisms that mediate the activation of myogenic precursors in response to mechanical loading may help in devising novel paradigms for ex vivo engineering of skeletal muscle. In this direction, significant efforts are being carried out worldwide to discover the optimal biophysical stimulation parameters leading to advanced maturation and assembly of skeletal myogenic precursors. 2
Mechanotransduction is a process whereby mechanical forces are sensed by specific cellular components and activate various biochemical intracellular signaling pathways. 3 In skeletal muscle cells, biophysical signals such as externally applied tensile stresses,4,5 substrate stiffness,6,7 and surface topography 8 have been shown to initiate mechanotransduction events by activating mechanosensors. These include stretch-activated ion channels, cell surface receptors, cytoskeletal proteins, and integrins. 9
Integrins, in particular, are believed to be primary sensors for relaying mechanical signals from the surrounding environment to the interior of the cell, as they constitute the direct physical link between the cytoskeleton and the extracellular matrix (ECM) and have the capacity to transduce the externally applied forces. 10 Integrin-mediated mechanotransduction signaling is initiated in intracellular adhesion complexes termed focal adhesions (FAs) on integrin transmembrane receptor activation 11 and subsequent recruitment and activation of focal adhesion kinase (FAK).12–14 FAK activation has been demonstrated to regulate responses both locally, by affecting the dynamics of focal adhesion assembly and disassembly15,16 and distantly, by activating the downstream pathways affecting cell growth,17,18 differentiation,19–21 and survival. 22
The effects of cyclic mechanical loading on myogenic differentiation are still not completely appreciated. The initiation of the myogenic differentiation program appears to be critically regulated by both magnitude and direction of the strain imposed to myogenic precursors.4,23,24 Studies have shown that myoblasts that are equally stretched from all horizontal directions (equiaxial strain) display an increased level of FAK activation and a reduced capacity of myogenic differentiation. 25
On the other hand, myoblasts that are predominantly stretched in one direction (uniaxial strain) display parallel alignment and an enhanced rate of differentiation.4,26,27 However, it is not yet known whether uniaxial cyclic tensile strain (UCTS) regulates FAK phosphorylation state, and to what extent FAK plays a role in UCTS-enhanced alignment and differentiation of myoblasts. Thus, in this study, we explore the FAK activation and role in skeletal muscle progenitors under cyclic unidirectional loading during the early differentiation phase.
Materials and Methods
Cell culture
Mouse myogenic cell line C2C12 from American Tissue Type Culture Collection (CRL-1772; LGC Standards, Boras, Sweden) was maintained in murine growth medium consisting of Dulbecco's modified Eagle's medium (Life Technologies Gibco, Naerum, Denmark) that was supplemented with 10% fetal calf serum (Helena Bioscience, Gateshead, United Kingdom), 100 U/mL penicillin, and 100 μg/mL streptomycin (all from Life Technologies Gibco). The myoblasts were cultured until 80–90% confluency, and then they were passaged into six-well flexible-bottom Bioflex culture plates precoated with collagen-I (Dunn Labortechnik, Asbach, Germany).
Mechanical stimulation and selective FAK inhibition
The C2C12 cells were subjected to vacuum-driven mechanical stimulation regimen by using a Flexcell FX-5000 Tension System (Flexcell International Corporation, Burlington, NC) that was fitted with custom-made rectangular loading posts to provide for a unidirectional strain field on the elastic growth surface. 28 In the control wells, the vacuum was prevented from affecting the membranes with the aid of the FlexStop valved rubber inserts (Flexcell). The strain was set to apply a peak magnitude of 15% at a frequency of 1 Hz for 48 h, and the cells were grown in murine growth medium. The cycling was interrupted after 24 h to reorient the designated pistons per experimental design.
To inhibit activation of FAK, FAK 14 blocker, 1,2,4,5-Benzenetetramine tetrahydrochloride (No. 3413; Tocris Bioscience, Bristol, United Kingdom), which is a specific inhibitor of FAK phosphorylation at the Y397 site, was used.
Western blot analysis
To determine the levels of FAK and phosphorylated FAK (pFAK), the cells were homogenized by sonication in a lysis buffer (20 mM Tris, pH 7.4; 150 mM NaCl, 1% Triton X-100), the lysate was clarified by centrifugation at 20,000 g at 4°C for 20 min, and the protein concentration was determined by using BCA protein assay kit (ThermoFisher Scientific, Waltham, MA). Proteins were resolved on a 10% Mini-PROTEAN TGX SDS polyacrylamide precast gel and transferred by wet blotting in an iBlot system to PVDF membranes (0.2 μm pore size) (all Bio-Rad, Berkeley, CA).
For the immunodetection, the membranes were first blocked with 5% dry milk in Tris-buffered saline/Tween-20 (TBS-T) consisting of 20 mM Tris, pH 7.5; 150 mM NaCl; and 0.3% Tween-20, and then they were reacted with the FAK (D-1) (No. sc-271126) and pFAK (Tyr 397) (No. sc-11765-R; both from Santa Cruz, Dallas, TX) antibodies in 500- and 150-fold dilutions in TBS-T and TBS-T, respectively, at 4°C for 16 h. After a wash with TBS-T, anti-mouse (No. A-9213), and anti-rabbit (No. A-2841a; both from Invitrogen, Waltham, MA), IgG horseradish peroxidase conjugates were utilized to tag FAK and pFAK, respectively. Both conjugates were diluted 2000-fold in TBS-T, and they were incubated at room temperature for 1 h.
After the final wash with TBS-T, the specific bands were visualized (ECL-plus; Amersham Biosciences, Little Chalfont, United Kingdom), and they were quantified by employing the LI-COR Odyssey Fc imager and the resident software bundle (LI-COR Image Studio; both from LI-COR, Lincoln, NE).
Immunofluorescence staining
For immunofluorescence analysis, the cell samples were first fixed with 4% buffered formaldehyde for 15 min, then permeabilized with 0.1% Triton X-100 in phosphate-buffered saline (PBS) for 5 min, and finally blocked with 1% bovine serum albumin in PBS (both from Sigma-Aldrich, St. Louis, MO) for 30 min, all at room temperature.
The exposure to primary antibodies involved the same reagents as with immunoblotting, except the dilution was 1:100 in PBS, and the incubation took place at room temperature for 1 h. Visualization of primary antibodies and F-actin was accomplished by incubation with Alexa Fluor 488 anti-mouse (No. A-21202) and Cy5 anti-rabbit IgGs (No. A-10523; both from Invitrogen) diluted 1:150 and 1:100, respectively, and BODIPY 558/568 phalloidin (No. B3475; ThermoFisher Scientific) diluted 1:500, all in PBS at room temperature for 1 h. The preparations were mounted in buffered glycerol and stored at 4°C until evaluation. The obtained micrograph images were pseudo-colored with actin in red, FAK in blue, and pFAK in green.
Microscopy and cytomorphometry
The phase-contrast and fluorescence images were obtained with the aid of the Zeiss Axio Observer.Z1 microscope (Carl Zeiss, Göttingen, Germany) that was equipped with 63 × /1.20 C-Apochromat objective, motorized stage and focus, and Hamamatsu ORCA-Flash4.0 CMOS camera (Hamamatsu Photonics K.K., Tokyo, Japan) by using Zen 2012 Blue Edition software (Carl Zeiss).
For each immunofluorescence sample, a representative z-stack of 20 high-resolution images was acquired. To equalize focal plane variance and improve quality of the images, the samples were three-dimensional (3D) deconvoluted by using Deconvolution plugin for Zen. 3D deconvolution was performed by using a theoretical point spread function with 10 iterations.
Cell directionality was evaluated, invoking ImageJ (NIH, Bethesda, MD) plugin OrientationJ (Daniel Sage, Swiss Federal Institute of Technology Lausanne, EPFL, Lausanne, Switzerland) based on the analysis of structure tensor in local neighborhood utilizing phase-contrast micrographs. Single cell area and the aspect ratio were determined on thresholded images by using ImageJ (NIH) measure function or fitted ellipse, respectively. The aspect ratio is an expression of the relationship between the dimensions along the major and minor axes.
Real-time reverse transcription-polymerase chain reaction
Transcriptional analysis was undertaken to assess the expression of early and intermediate myogenic markers, including myogenic factor 5 (MYF5) and myogenin (MYOG), respectively. To accomplish this, the total RNA was extracted by Aurum total RNA mini kit (Bio-Rad) and quantified by UV-Vis spectrophotometry and capillary electrophoresis (Agilent RNA 6000 Nano kit; Agilent Technologies, Santa Clara, CA). One microgram of RNA was used to synthesize the cDNA by iScript cDNA synthesis kit (Bio-Rad).
The polymerase chain reactions (PCRs) were performed on the basis of iQ SYBR Green Supermix in a CFX Connect Real-Time PCR Detection System (both from Bio-Rad). The amplification involved an initial step at 95°C for 3 min, followed by 40 cycles consisting of annealing and elongation at 63°C for 30 s and denaturation at 95°C for 15 s. The expression levels of the analyzed genes were normalized to those of the selected housekeeping genes tyrosine 3/tryptophan 5-monooxygenase activation protein (YWHAZ) and cyclophilin A (PPIA) (both from DNA Technology A/S, Risskov, Denmark). 29 The primer sequences can be reviewed in Table 1.
MYF5, myogenic factor 5; MYOG, myogenin; PPIA, cyclophilin A; YWHAZ, 3-/tryptophan 5-monooxygenase activation protein.
Statistics
The data originate from at least two independent experiments designed with multiple replicates, and they are presented as means + standard error of the mean. Differences between two independent groups were evaluated by using the unpaired t-test from the IBM SPSS statistics 24 package (IBM SPSS, Chicago, IL). Statistical significance was assigned to p < 0.05.
Results
FAK mediates cell reorientation in myoblasts subjected to a transversal strain field
The protocol, whereby the muscle progenitor cells were successively subjected to differentially oriented strain fields in the condition that the FAK activation beyond the steady-state level was inhibited, is outlined in Figure 1. It consisted of three stages, where in the first stage, the C2C12 myoblasts were expanded on BioFlex culture plates for 2 days; in the second 24 h, the preconditioning stage, the cells were aligned under the effect of UCTS, as previously described 4 ; and lastly, in the third 24 h period, the cells were continued under isodirectional straining or were subjected to UCTS reoriented by 90°, in the presence or not of FAK inhibitor.
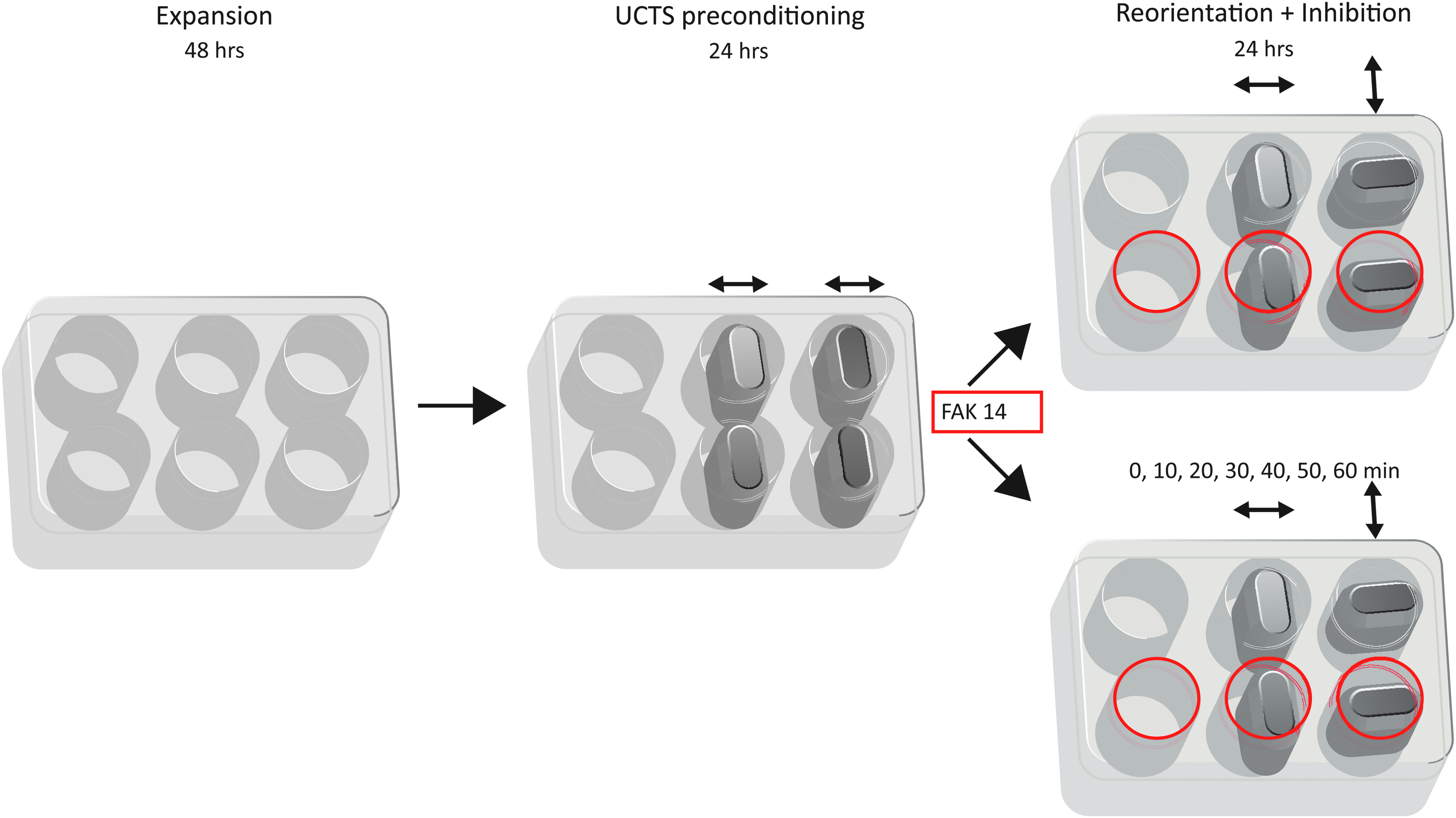
Schematic representation of the experimental setup. In the first stage, the C2C12 myoblasts were expanded on collagen I-coated flexible-bottom BioFlex culture plates for 2 days. After reaching confluency, in the second stage, the cultures were preconditioned by 15% cyclic strain at 1 Hz for 24 h to achieve unidirectional alignment. In the final third stage, the strain field was reoriented by 90°, and the FAK activation was inhibited beyond the steady state by using a specific blocker (FAK 14). The cultures were analyzed at the end or in 10 min intervals during the first hour of the last period. The orientation of the strain field is indicated by double-headed arrows, and the wells with the presence of FAK inhibitor are delineated in red. FAK, focal adhesion kinase.
A critical amount of FAK 14 that would not affect the short-term survival of the cells was determined by titrating an FAK 14 dilution series ranging from 2.5 to 10 μM against morphological changes (Fig. 2). First, changes were observed already after 10 min of exposure to 10 μM FAK 14; whereas, at 2.5 and 5 μM concentrations, the cultures appeared unaffected for at least 24 h. Based on the cytomorphometric data representing the total cell area, it was concluded that 7.5 μM was the maximal concentration at which cell spreading was not significantly affected. This concentration was used in all ensuing experiments.
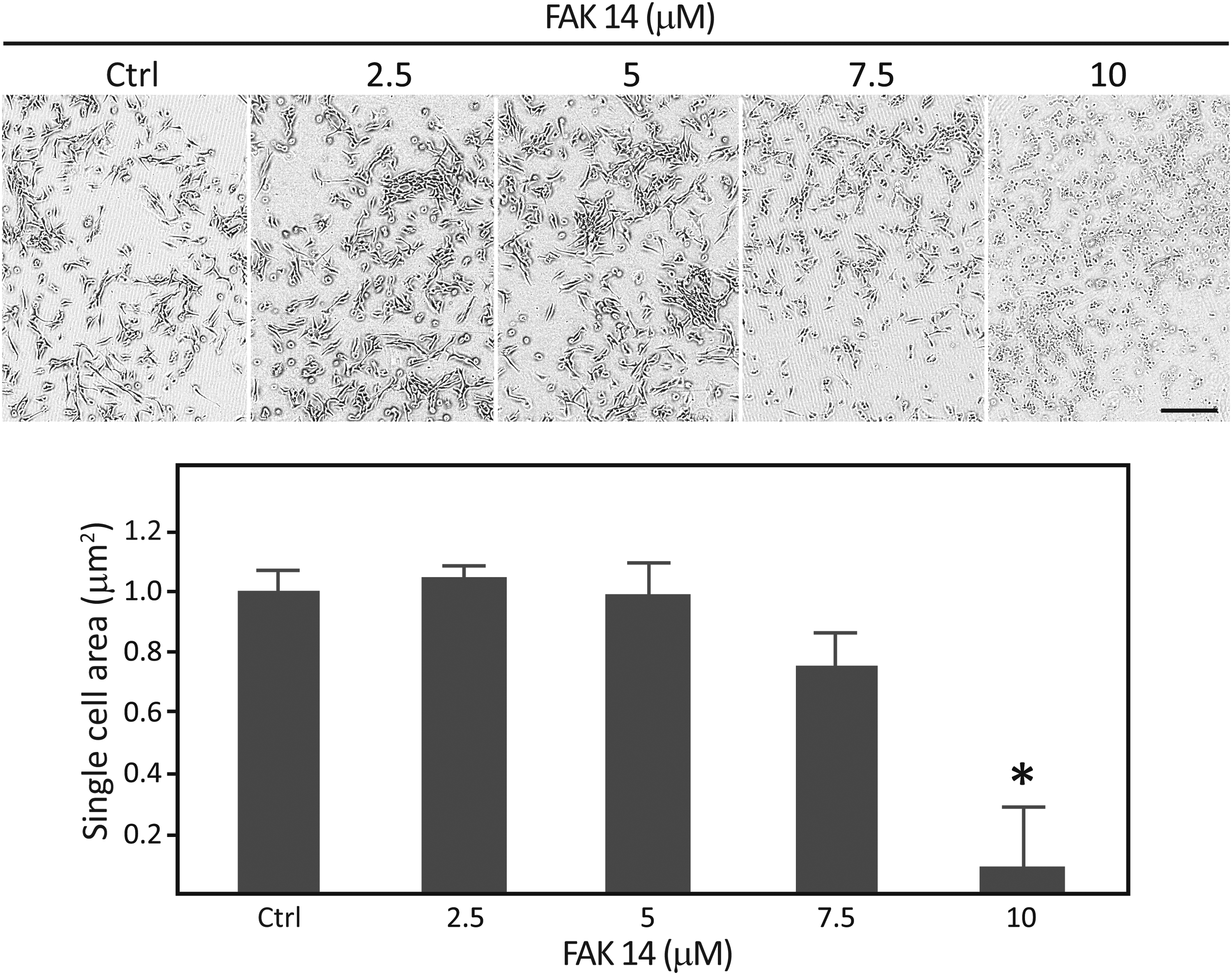
Determination of the maximal inhibitory concentration of FAK 14. The C2C12 myogenic precursors were grown on collagen I-coated flexible-bottom BioFlex culture plates and exposed to increasing concentrations of FAK 14. The effect of the inhibitor was assessed from the cell spreading normalized to that of the control, and 7.5 μM was selected as a concentration that still enabled steady-state phosphorylation. The data are presented as mean + SEM from two independent experiments carried out in duplicate (n = 4). *Significantly different from control at p < 0.05. The scale bar indicates 100 μm. SEM, standard error of the mean.
In the next step, we proceeded to investigate whether FAK activation is necessary for the myoblast alignment in response to a uniaxial strain field. The cells that have not been strained displayed a random pattern of organization, independently of the presence of FAK 14 (Fig. 3A, B). As shown in the control condition at the end of the second stretching interval, the cells subjected to UCTS aligned, and their orientation was perpendicular to the direction of the strain field (Fig. 3C). These cells were able to rearrange themselves when challenged by a 90° rotation of the strain field (Fig. 3E).
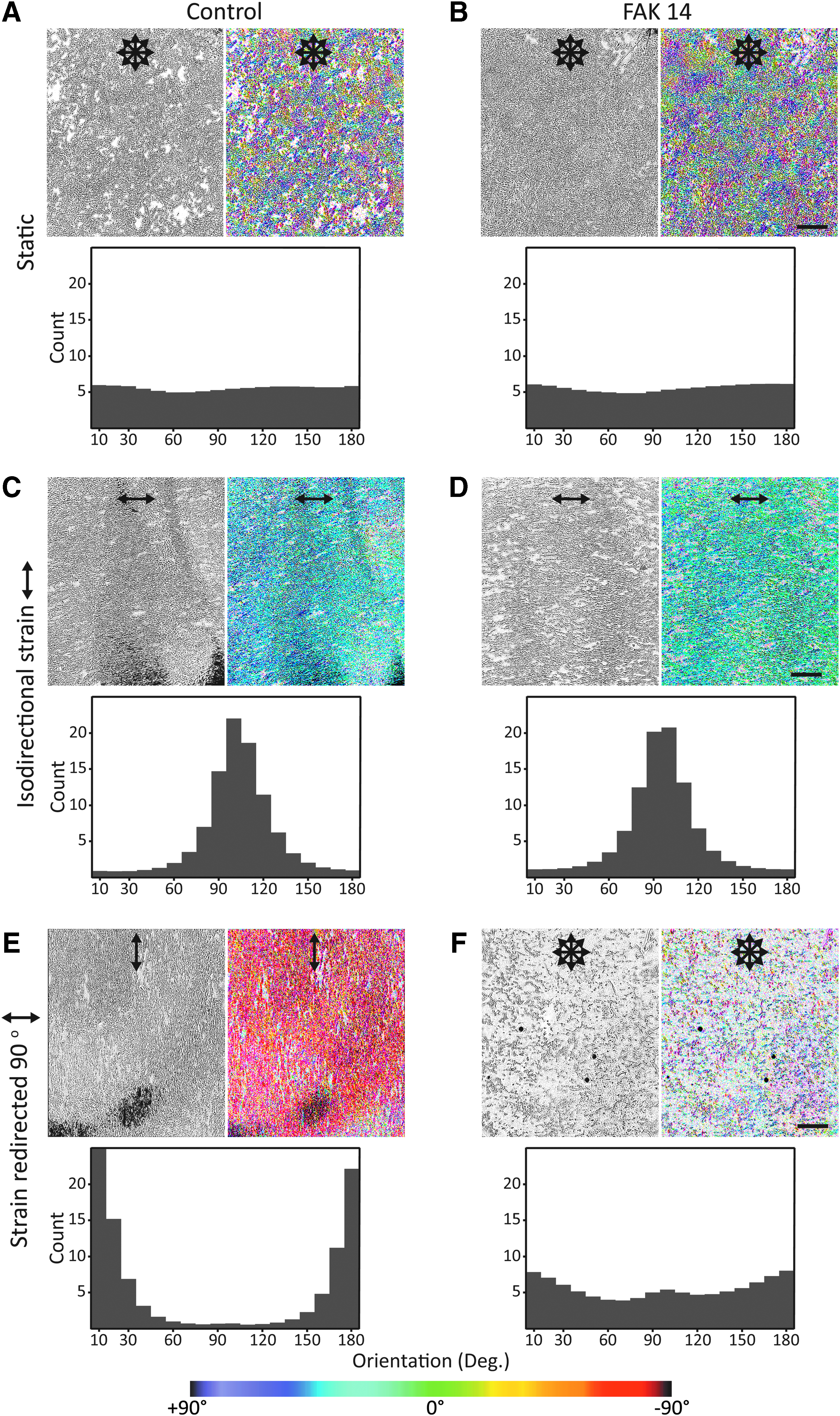
Role of FAK activation in cell responses to isodirectional and redirected strain loading. At the end of the last experimental stage, the cultures maintained omnidirectional appearance without strain loading, irrespective of FAK 14
A different scenario, however, became evident in the presence of FAK inhibitor. Here, as long as the orientation of the strain field remained unchanged, the cell polarization appeared intact (Fig. 3D) but on rotating the strain field by 90°, the cells shrank and detached (Fig. 3F). This experiment demonstrates that full FAK functionality is necessary for the cells to minimize through adjustment of the cytoskeleton the strain resulting from the externally applied stress.
FAK activation in response to UCTS
The extent of FAK autophosphorylation was determined by immunoblotting. As seen in Figure 4A, after a total period of 48 h in static growth conditions, the cells exhibited a steady-state level of pFAK, irrespective of the presence of FAK inhibitor. The exposure to strain resulted in significant upregulation of FAK, and this was abrogated by FAK 14 no matter whether the strain during the last 24 h period was isodirectional or perpendicular with respect to the first preconditioning stage. The immediate response after the addition of FAK 14, at the beginning of the second straining stage, is presented in Figure 4B. It can be seen that the strain field reorientation resulted in FAK activation beyond the effect of continuous straining only during the initial short interval of about 30 min, after which the kinase activity remained at the same level in both straining scenarios. Similarly, the blocking by FAK 14 reached full effect between 30 and 40 min, from which point the autophosphorylation activity coincided with that of the static cultures.

FAK phosphorylation in response to isodirectional and redirected strain.
FAK recruitment to FAs in response to UCTS
The F-actin, FAK, and pFAK were visualized and evaluated to study the FAs assembly and activation (Fig. 5A). No usable micrographs were available from the end of the second straining interval, where the strain field was reoriented and FAK 14 added, since the cells succumbed to these conditions, as already shown in Figure 3F. Otherwise, in the UCTS conditions (strain), the actin filaments were predominantly arranged parallel to the major axis of the cells in patterns that were consistent with fusiform cell morphology; whereas in static cultures (static), the actin filaments were mostly randomly oriented. As for the FAK distribution, some perinuclear accumulation was noted but only in the static conditions; otherwise, it was associated with FAs. Similarly, pFAK was identified primarily at the FAs. Interestingly, on stretching, irrespective of FAK 14, the cells exhibited significantly elongated FAs with attached prominent actin stress fibers.
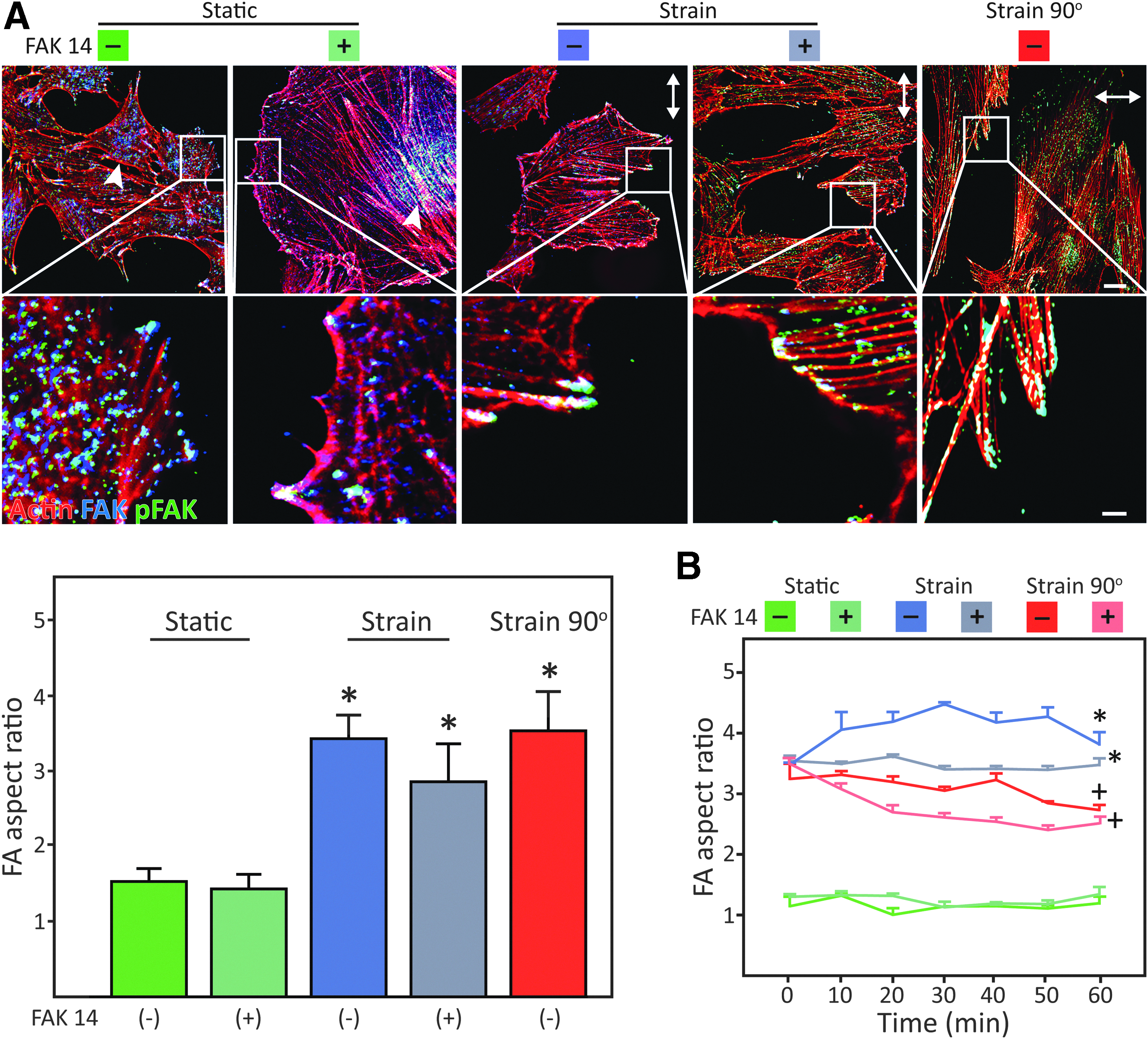
Role of FAK activation in the evolution of FA complexes after exposure to isodirectional and redirected strain.
Figure 5B displays a further dissection of the initial 60 min after the addition of FAK 14, immediately after the UCTS preconditioning stage. It was found that cells evidence a temporary increase in FA elongation as a reaction to the resumption of the isodirectional strain (strain−). As expected, the cells with inhibited FAK activation (strain+) were not able to emulate this process. On strain field redirection, the FA aspect ratio dropped as an indication of the cytoskeletal readjustment (strain 90°−) or failure to do so when the FAK inhibitor was present (strain 90°+).
FAK mediates myogenic differentiation induced by UCTS
As shown in Figure 6, the transcriptional activity of MYF5 was significantly upregulated after 24 h of UCTS. This effect became even more pronounced after the second 24 h stretching period, and it was here also obvious with MYOG. The inhibition by FAK 14 effectively suppressed the inducing effect of mechanical stimulation on myogenesis.
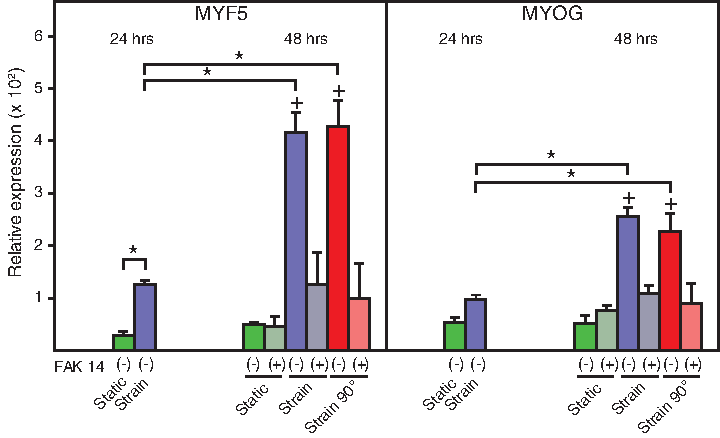
Role of FAK activation in the myogenic differentiation of C2C12 cells after exposure to isodirectional and redirected strain. The data were obtained after 24 h of straining during the preconditioning stage and at the end of the following 24 h period with straining in the presence of FAK inhibitor. The data are presented as mean + SEM from two independent experiments carried out in duplicate (n = 4). *Significantly different at p < 0.05; +significantly different from FAK 14 counterpart condition at p < 0.05.
Discussion
In this work, we have employed uniaxial cyclic tensile stimulation to investigate the role of FAK in the dynamic modulation of FA, cytoskeletal assembly, and differentiation of murine myoblasts. Our UCTS approach is based on an accurate knowledge of the distribution of the strain in the elastic growth substrate using a dedicated finite element model.6,21 It is well described that in response to cyclic uniaxial strain, adherent cells adjust actin fiber orientation and length to a threshold where fiber strain is minimized, 30 which drives the cells to a uniform orientation pattern.31,32
Here, after the 24 h preconditioning period, the UCTS field directed the myoblasts to achieve a parallel orientation. Previous studies have revealed that disruption of FAK significantly affects migration of myogenic precursors, as FAK regulates the assembly and disassembly of adhesion complexes.33,34 Indeed, our results show that FAK activation is necessary for FA reassembly and actin remodeling in myoblasts that are exposed to a sudden change of direction in the strain field.
Further, studies have shown that FAK inhibition does not affect FA orientation or its assembly rate in cells that are situated perpendicular to the direction of stretching. 35 In agreement with these results, here we show that FAK activation above the steady state does not appear to be necessary to maintain a feasible myoblast orientation, since continued isodirectional strain maintained the stretch-induced orientation, actin fiber arrangement, and focal adhesion aspect ratio, even in the presence of FAK 14. Interestingly, a previous study using osteoblasts under cyclic uniaxial strain has shown that FAK activation appears to display an initial orientation specific response and a long-term orientation independent response. 35
Our results show a gradual increase in the levels of FAK phosphorylation in cells that were challenged by a sudden rotation in the strain field, which later converged to a level equivalent to that in the isodirectionally strained cells. Therefore, myoblasts also appear to display two phases of FAK activation, which are dependent on the orientation of the strain field.
Several studies further suggest that FAK functions as a scaffolding protein where cellular signaling molecules are recruited in maturing FA,36,37 suggesting that FAK is a mediator of cell/ECM signaling events. FAK is present in the cytoplasm and is recruited to clustering of integrins during formation of FA as part of cell spreading and migration or in response to external mechanical stimulation. 38 On incorporation into FA, FAK is tyrosine autophosphorylated and serves its function through downstream effectors, such as cyclin D1 39 and SH2. 36
We observed that all cells subjected to strain display an elongation of FAs perpendicular to strain direction with a migration of cytoplasmic FAK toward maturing FA, where an increased turnover to phosphorylated FAK takes place. This activation can, in turn, be rapidly reverted by inhibiting FAK. With reengagement of strain, after the short period required for media change, the cells reacted with a short-lived elongation of FA. This short-lived effect was absent in FAK-impaired cells.
Change in strain field direction induced a short period of shrinking FA as the cells realigned, where blocking of FAK phosphorylation effectively disabled cell realignment and the cells succumbed. This again strongly suggests that FAK is crucial in the initial response to changes in external stresses, and if the directional changes are diverting too much cells would effectively succumb when they are not capable of engaging in FAK signaling.
It has previously been described that UCTS enhances assembly and maturation of myogenic mammalian progenitors, which is evident through early myogenin activation, enhanced formation of myosin-rich myotubes,4,20,40 and enhanced fusogenic capacity of the cells. 26 In particular, myogenin activation is a key landmark in the myogenic differentiation pathway, representing the so-called “point of no return.” 41 Thus, the enhanced regulation of MYOG displayed by cells under UCTS clearly indicates exit from the cell cycle and commitment to the final differentiation program.
Our data confirm the positive effect of UCTS in myogenesis and indicate that the enhancement of myogenic differentiation is indifferent of sudden changes in strain direction. However, impairment of FAK activation can abolish the effect to such an extent that continuous UCTS no longer increases the myogenic differentiation rate. This indicates that a continuous engagement of FAK signaling is needed to drive myogenic differentiation.
Other studies have suggested that impairment of FAK signaling might compromise myogenesis by reducing the ability of the myoblasts to undergo fusion, which is a crucial event in the myogenic differentiation process. 22 It is worth noticing that we did not find evidence of myotube formation in any of the wells assessed after 48 h (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/tea).
This is not unexpected, as our experiments were performed by using growth medium to prevent the formation of myotubes, which would have detached on rotation of the piston due to their inability to form new FAs. This is in agreement with the study of Yoshiko et al., which has shown that myotube formation is still negligible in growth conditions during the 48 h after confluence. 42 Even though we focus on the early differentiation and did not attempt to assess the subsequent myoblast fusion events, it remains to be investigated whether UCTS-mediated FAK activation might modulate myoblast differentiation through increased fusogenic activity.
The mechanisms involved in FAK phosphorylation in transducing mechanical stimuli are not fully understood. However, our findings suggest that most activation of FAK takes place in large FAs, indicating a strong association between FAK phosphorylation and transduction of external mechanical loads. Further, our findings indicate that FAK activation above the steady-state level has a significant effect on the myogenic differentiation program. It is worth mentioning that our results might not be directly transferrable to primary myoblasts, since the C2C12 cell line appears to behave differently in response to biophysical environmental cues as compared with primary muscle progenitor cells.43,44 Further, extrapolation of these results to more realistic 3D environments is not straightforward, which warrants further research efforts in the subject.
In conclusion, we showed that in myoblasts subjected to cyclic uniaxial strain, FAK activation is of paramount importance for cell motility responses leading to cell reorientation. Further, a persistent activation of FAK appears to be necessary to promote stretch-induced myogenic differentiation in C2C12 cells.
Footnotes
Acknowledgments
The technical assistance of Ole Jensen and Lisa Engen is appreciated.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.