
Abstract
Bacterial contamination during the healing of bone defects frequently compromises the effects of bone regenerative therapy. Human beta-defensin-2 (hBD2) and -3 (hBD3) are antimicrobial peptides of human innate immune system with a broad antibacterial spectrum and rare bacterial resistance. The purpose of this study was to determine the effect of hBD2 and hBD3 on the healing of bacteria-contaminated bone defects. Rat bone marrow stromal cells (BMSCs) were infected with adenovirus to overexpress hBD2 or hBD3. Treatment with the conditioned medium derived from the BMSCs overexpressing defensins could concentration dependently reduce the viable Staphylococcus aureus numbers in the colony formation assay. In addition, the antimicrobial effect of BMSCs overexpressing defensins was verified with a diffusion chamber model in rats. Furthermore, we established a S. aureus-contaminated rat calvarial defect model and demonstrated that S. aureus contamination significantly compromised the bone regenerative effect after treatment with wild-type BMSCs. When defensin-overexpressing BMSCs were implanted into the S. aureus-contaminated defect, the viable S. aureus numbers were dramatically reduced and the negative effects of S. aureus contamination on bone healing were significantly mitigated. In conclusion, application of hBD2 or hBD3 promotes the healing of S. aureus-contaminated bone defects.
Introduction
B
Staphylococcus aureus is the leading cause of osteomyelitis and joint infection. It is predominant in the bacterial cultures performed initially and between 3 to 7 days after surgery in open fractures. 2 The ever increasing occurrence of S. aureus with antibiotic resistance has caused serious clinical problems and has become an important issue.3,4
Antimicrobial peptides (AMPs) play a fundamental role in innate immunity. A sensitive microbial strain is surprisingly unlikely to acquire resistance against AMPs. 5 Furthermore, a major strength of AMPs is their antimicrobial activity against multidrug-resistant bacteria. 6 The rapid development of bacterial resistance to conventional antibiotics has stimulated interest in the application of AMPs for treating infections. 7
Human beta-defensin-2 (hBD2) and -3 (hBD3) are important human AMPs. Both hBD2 and hBD3 show killing activity against both methicillin-sensitive S. aureus (MSSA) and methicillin-resistant S. aureus (MRSA) in a concentration-dependent manner. In addition, the bactericidal effect in MRSA is less pronounced than in MSSA. 8 However, hBD2 and hBD3 can kill 100% S. aureus COL (a methicillin-resistant strain) at 10 and 5 μg/mL, respectively.9,10 It also has been reported that hBD3 shows potent antimicrobial activity against multidrug-resistant S. aureus.11,12 The secretion of hBD2 and hBD3 are upregulated after microorganism stimulation, 13 which indicates that they may play important protective roles during microorganism invasion.
hBD2 and hBD3 were first found produced by epithelial cells. Previous application studies of hBD2 or hBD3 were mainly focused on treating infected wounds. Furthermore, hBD2 or hBD3 was mostly delivered through gene therapy strategies.14–16 Interestingly, hBD2 and hBD3 may play an important protective role in bone to combat bacterial invasion. Both hBD2 and hBD3 are secreted by cultured osteoblasts17,18 and are increased after stimulation with S. aureus supernatants.18,19 The expression of hBD2 and hBD3 is detectable in noninfected bones and is upregulated in chronically infected bone.17,18 However, the application of hBD2 and hBD3 in treatment of bone defects with bacterial contamination or infection is still unexplored. The purpose of this study was to determine the effect of hBD2 and hBD3 on bone healing of S. aureus-contaminated bone defect.
Materials and Methods
Rats
Male Sprague Dawley rats (4-week-old rats for the isolation of bone marrow stromal cells (BMSCs), and 8-week-old rats for the diffusion chamber and the calvarial defect experiments) were used. All surgical procedures were performed under general anesthesia by intraperitoneal administration of xylazine (15 mg/kg BW) and ketamine (100 mg/kg BW). The animal research protocols of the present study were reviewed and approved by the Institutional Animal Care and Use Committee of the National Ying-Ming University.
BMSCs culture
Rat BMSCs were isolated from bone marrow aspirates by density gradient centrifugation method. 20 The cells were cultured in α-MEM (GIBCO-Life Technologies, Grand Island, NY), supplemented with 16.6% fetal bovine serum under hypoxic condition (94% N2, 5% CO2, and 1% O2). Nonadherent cells were removed 24 h after initial culture. The attached cells were passaged when the culture reached subconfluence after 10 days. Starting from passage 1, the BMSCs were cultured in low density (seeded at 100 cells/cm2) and under hypoxic condition according to a previous study. 21
Preparation of recombinant adenoviruses
Recombinant adenoviruses (replication incompetent [ΔE1/ΔE3] human adenovirus type 5), encoding human hBD2 (Ad-hBD2) or hBD3 (Ad-hBD3) gene, were generated with an adenovirus construction kit (Adeno-X™ Expression System 1, containing pShuttle2 cloning vector; Clontech, Mountain View, CA) as per the manufacturer's instruction. The gene fragments of hBD2 and hBD3 were cloned from cell lines (hBD2: HaCaT, hBD3: SAS) by reverse transcription–polymerase chain reaction (RT-PCR) (Table 1). Ad-LacZ, produced by pShuttle2-LacZ control vector encoding Escherichia coli beta-galactosidase gene, was used as a control virus.
hBD2, human beta-defensin-2; hBD3, human beta-defensin-3.
Generation of gene-modified BMSCs
The rat BMSCs were infected with adenoviruses to overexpress hBD2, hBD3, or lacZ. The infections were performed as previously described with minor modification. 22 In brief, passage 3 cells were grown in a 10-cm culture dish to 80% confluence, followed by 2 h of incubation with recombinant adenovirus (M.O.I. 1000) in fresh serum-free medium.
Preparation of conditioned medium
Conditioned mediums (CMs) were harvested from the gene-modified BMSCs (BMSC/hBD2, BMSC/hBD3, or BMSC/LacZ). Gene-modified BMSCs (1 × 106) were seeded in 10-cm culture dish. The culture medium was changed into 10 mL fresh serum-free medium the following day. CMs were collected after 48 h and concentrated 50-folds by a stirred ultrafiltration cell (Merck Millipore, Darmstadt, Germany) with ultrafiltration discs (PLGC, Ultracel regenerated cellulose, 1 kDa NMWL) (Merck Millipore) for further tests. The concentrations of hBD2 and hBD3 in the CMs were determined by hBD2 (Komabiotech, Seoul, Korea) and hBD3 (Alpha Diagnostic, San Antonio, TX) Enzyme-Linked Immunosorbent Assay (ELISA) Kits, respectively.
Colony formation assay
To determine viable bacterial numbers, colony-forming unit was counted after spreading the bacterial solution onto Tryptic Soy Broth (TSB) agar plates for 16 h at 37°C.
Preparation of collagen sponges
Disk-shaped collagen sponges were fabricated and utilized in the diffusion chamber model (10 mm in diameter and 2 mm in thickness) and the calvarial defect experiments (8 mm in diameter and 1 mm in thickness) described below. The sponges were prepared as previously described with minor modification. 23 In brief, type I rat collagen acidic solution was frozen at −80°C for 12 h and freeze dried to form the collagen sponges. The sponges were crosslinked by glutaraldehyde followed by glycine solution treatment.
Rat diffusion chamber model
Diffusion chambers (Merck Millipore) sealed with 0.22 μm PVDF hydrophilic membranes (Merck Millipore) were used. Diffusion chambers containing collagen sponge with 1 × 106 gene-modified BMSCs (BMSC/hBD2, BMSC/hBD3, or BMSC/LacZ) and 1 × 105 S. aureus (ATCC 25923) were randomly implanted into subcutaneous pockets on the back of three rats. Each rat received implantation of six diffusion chambers (two diffusion chambers per group). Three diffusion chambers per rat (one from each group) were retrieved after 24 and 72 h.
Rat calvarial defect model
Calvarial bone defects, 8 mm in diameter, were generated using a trephine bur under copious saline irrigation. Rats were randomly assigned to groups with different treatment. In groups with BMSC implantation, collagen sponge with 1 × 106 cells seeded 8 h before implantation was implanted. For S. aureus contamination, S. aureus (1 × 108 in 0.1 mL TSB medium) were spread evenly in the defect before wound closure.
Microcomputed tomography measurement
Calvarias were scanned, with an 8.6 μm image pixel size, by a compact μCT imaging system (SkyScan1174; SkyScan, Kontich, Belgium). The bone volume, bone surface density, and trabecular number of the centered 6-mm zone were determined by μCT. In addition, the healing area proportion of the 8-mm defect area was measured from μCT three-dimensional (3D) reconstruction images by an image analysis software (ImageJ 1.48; National Institutes of Health, Bethesda, MD).
Histological examination
Samples were fixed in 4% paraformaldehyde and embedded in paraffin. Five micrometer sections were cut perpendicular to the sagittal suture at the center of the created defect and stained with Hematoxylin and Eosin to evaluate the bone healing.
Statistical analysis
Statistical analysis was performed using unpaired t-test (ELISA data), repeated measure analysis of variance (diffusion chamber experiment data), or one-way analysis of variance (all other data). Statistical significance was designated at the level of p < 0.05. Data are shown as mean ± standard deviation.
Results
Generation of BMSCs overexpressing hBD2, hBD3, or lacZ
Adenoviral vectors carrying lacZ (control), hBD2, or hBD3 gene (Fig. 1A) were used to infect rat BMSCs. The RT-PCR data indicated that BMSCs infected with Ad-hBD2 or Ad-hBD3 expressed their respective mRNA (Fig. 1B). Furthermore, the ELISA data indicated that the secreted hBD2 and hBD3 peptides from BMSC were increased after Ad-hBD2 and Ad-hBD3 infection, respectively (Fig. 1C).
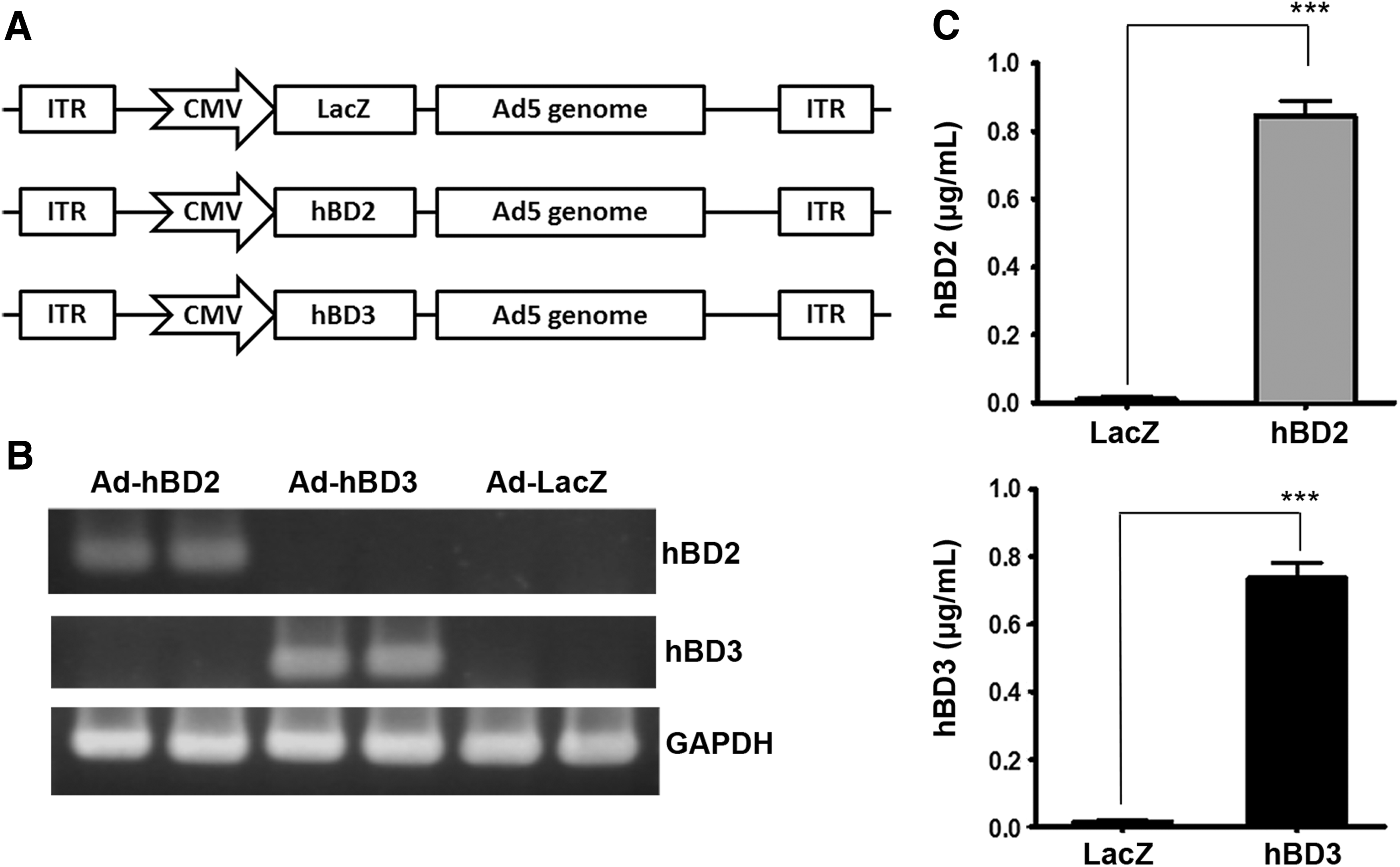
Generation of gene-modified BMSCs through adenoviral infection.
CMs from BMSC/hBD2 and BMSC/hBD3 inhibit the growth of S. aureus in vitro
S. aureus (1 × 103) were incubated with 100 μL CMs (1 ×∼50 × ) from different gene-modified BMSCs for 2 h at 37°C.13,24 The resulting S. aureus colonies in the colony formation assay were shown in Figure 2A. The quantified data showed that the percentage of colony numbers did not decrease with the fold's increase in the LacZ group. In contrast, the hBD2 and the hBD3 groups exhibited antimicrobial activity in a fold-dependent manner (Fig. 2B).

The CMs from BMSC/hBD2 and BMSC/hBD3 show antimicrobial activity in vitro.
BMSC/hBD2 and BMSC/hBD3 reduce viable bacteria in the diffusion chamber in vivo
The antimicrobial effect of gene-modified BMSCs was further tested in a subcutaneous diffusion chamber model (Fig. 3A). The S. aureus in the diffusion chamber were eluted by vortexing the content with phosphate-buffered saline (PBS). The viable S. aureus numbers were determined by the colony formation assay. The viable S. aureus numbers continually increased from implantation to 72 h in the LacZ group. In contrast, the number of viable S. aureus in the hBD2 and the hBD3 groups was dramatically reduced as the time increased (Fig. 3B, C).
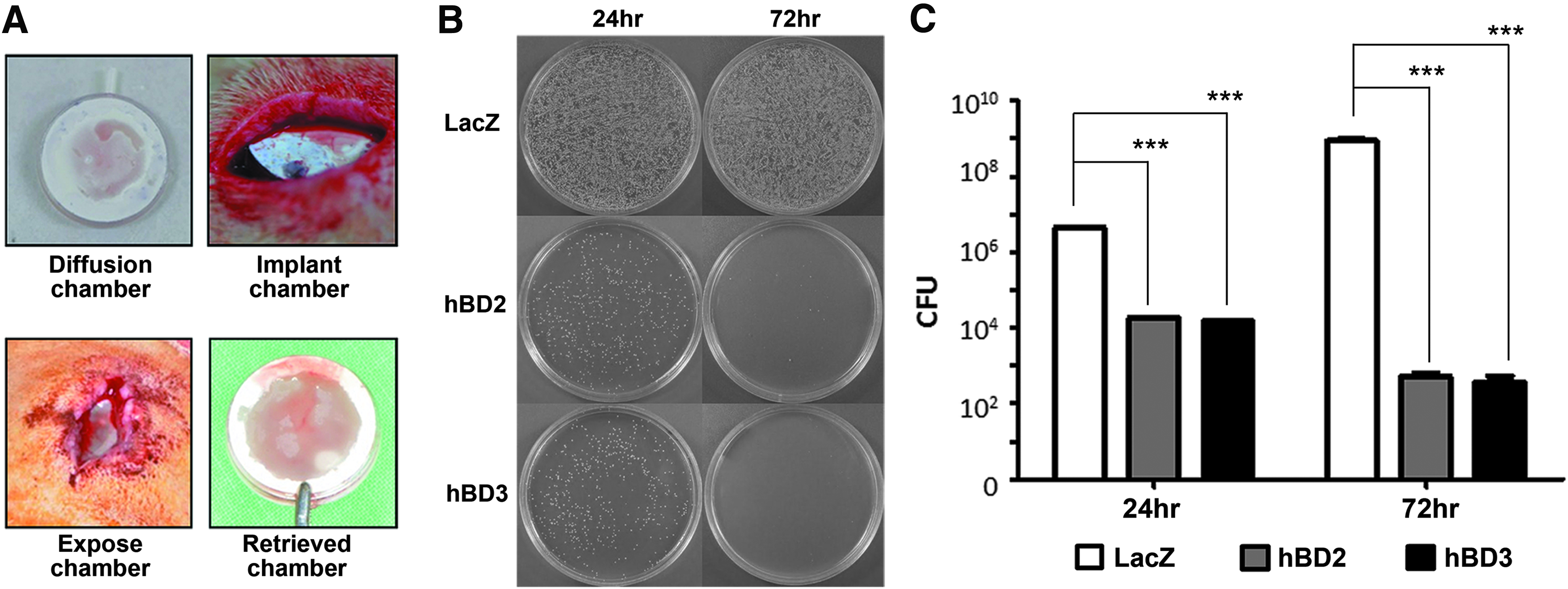
BMSC/hBD2 and BMSC/hBD3 demonstrate antimicrobial activity in the diffusion chamber model in vivo.
S. aureus contamination negatively affects the bone regeneration of rat calvarial defect
Collagen sponges preseeded with BMSCs were implanted into rat calvarial defects (Fig. 4A) and contaminated with S. aureus. The empty group showed barely any new bone formation in the dashed circle area (Empty group). The implantation of wild-type BMSCs effectively promoted the bone healing (BMSC group). However, the S. aureus contamination severely hindered the bone regeneration after BMSC implantation (BMSC+S. aureus group) (Fig. 4B). The quantitation data of bone volume of the centered 6-mm zone of defect indicated that the BMSC+S. aureus group showed significantly less bone volume when compared with the BMSC group (Fig. 4C, upper panel). The healing area proportion data showed a similar trend (Fig. 4C, lower panel).
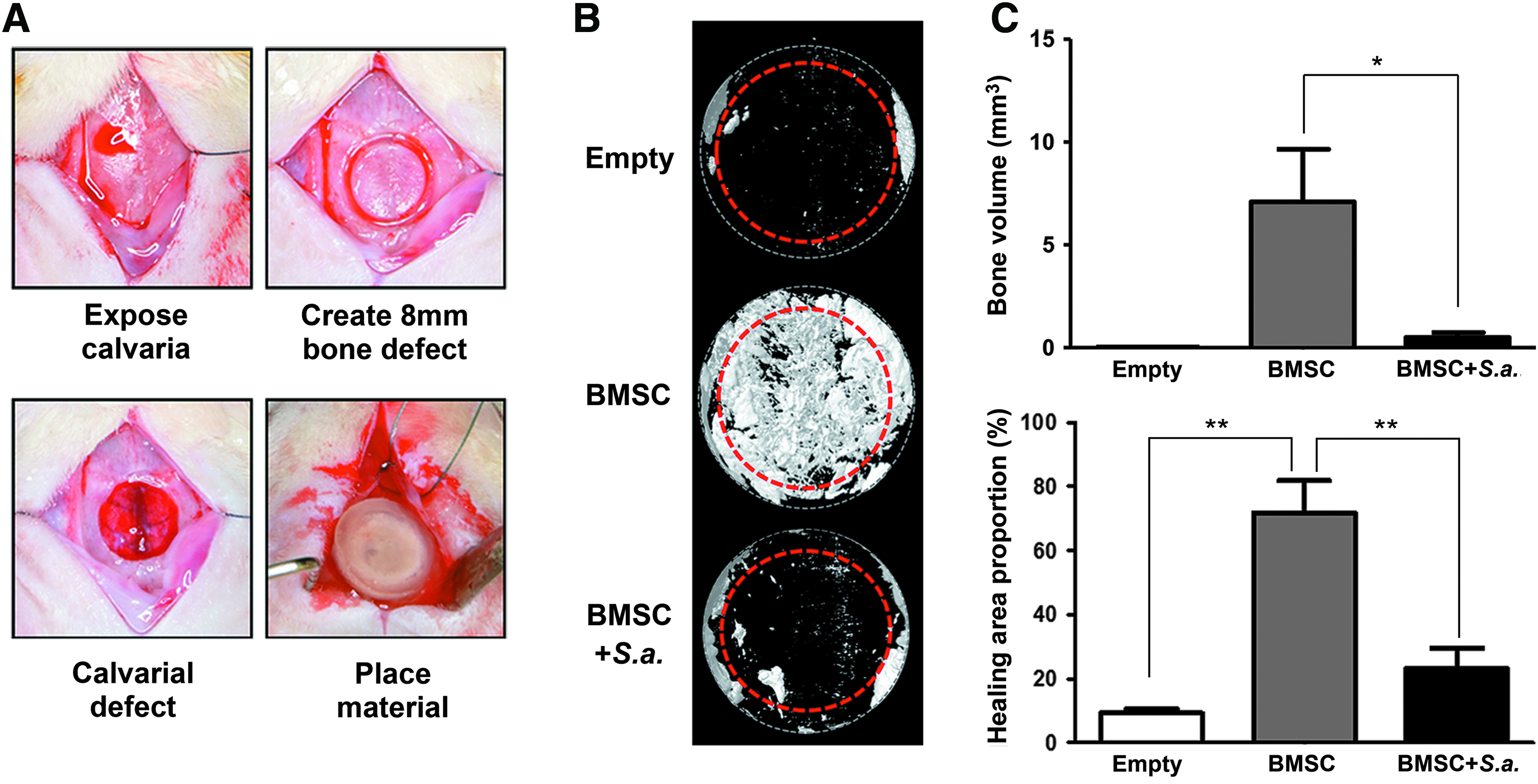
S. aureus contamination significantly impairs the BMSC-based bone regenerative therapy in rat calvarial defect model.
HBD gene-modified BMSCs decrease viable S. aureus numbers and promote bone healing in rat calvarial bone defects contaminated with S. aureus
The study design is shown in Figure 5. BMSC/hBD2, BMSC/hBD3, or BMSC/LacZ, were implanted into rat calvarial defects and contaminated with S. aureus (1 × 108). After 3 days, the S. aureus in the defects were eluted by vortexing the content with PBS. The viable S. aureus numbers were determined by the colony formation assay (Fig. 6A). The quantified data revealed that the viable S. aureus numbers were dramatically reduced in the hBD2 and the hBD3 groups, but not in the LacZ group (Fig. 6B).
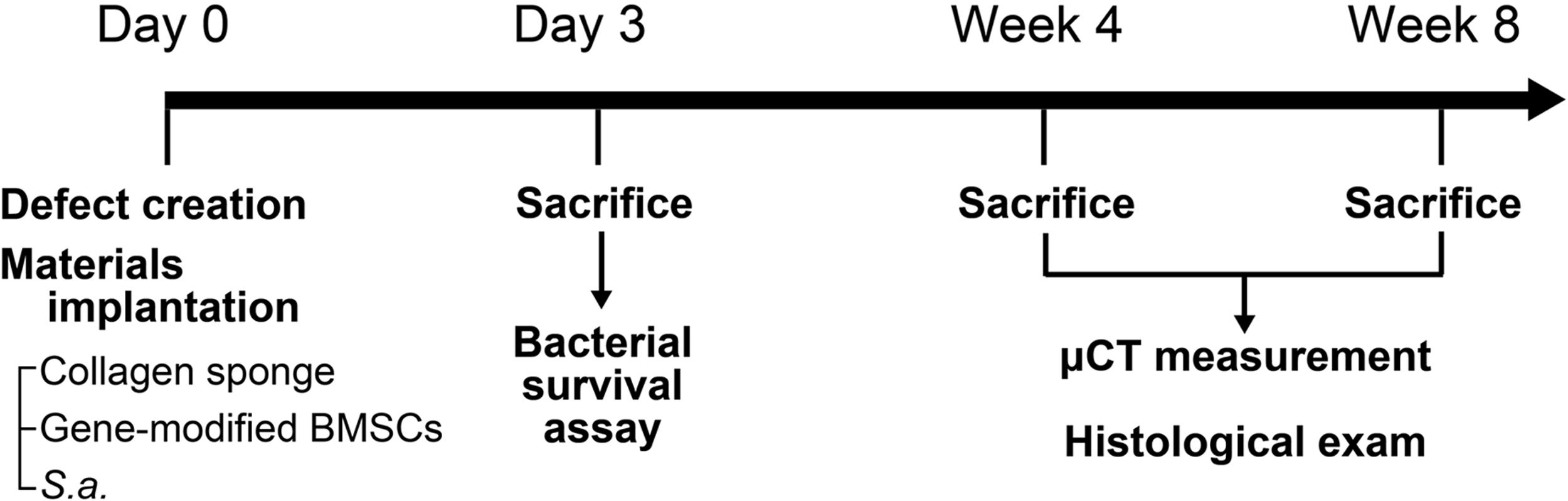
Experimental design to evaluate the antimicrobial and bone regenerative effects of gene-modified BMSCs in the S. aureus-contaminated calvarial defect model. At day 0, defect creation, implantation of collagen sponge preseeded with gene-modified BMSCs, and bacterial contamination with S. aureus were performed. After 3 days, the viable S. aureus number was determined by the colony formation assay. The bone healing was evaluated by μCT measurement and histological examination at 4 and 8 weeks.

BMSC/hBD2 and BMSC/hBD3 demonstrate antimicrobial activity in the S. aureus-contaminated calvarial defect model. The colony formation assay was performed 3 days after surgery.
Although S. aureus contamination was performed in all rats, no obvious signs of inflammation or abscess formation were found at the surgical sites during the healing period in any rat. The 3D reconstruction images of μCT indicated that the LacZ group only formed some new bone in the defect margin area and had barely any new bone formed in the central area at both 4 and 8 weeks (Fig. 7A). In contrast, the hBD2 and the hBD3 groups showed robust new bone formation in both areas. The bone volume of the hBD3 group was more than that of the LacZ group at 4 weeks. At 8 weeks, both the hBD2 and the hBD3 groups showed more bone volume when compared with the LacZ group. Furthermore, the bone healing area proportion, bone surface density, and trabecular number data showed a similar trend (Fig. 7B). Histological examination of the calvarial sections at 4 and 8 weeks revealed better bone formation in the hBD2 and the hBD3 groups than in the LacZ group (Fig. 7C). In addition, no obvious histological signs of inflammation or infection were noticed. Taken together, these results indicated that the hBD gene-modified BMSCs significantly reduced the viable S. aureus numbers at the early healing stage and promoted new bone formation in the S. aureus-contaminated rat calvarial defects.
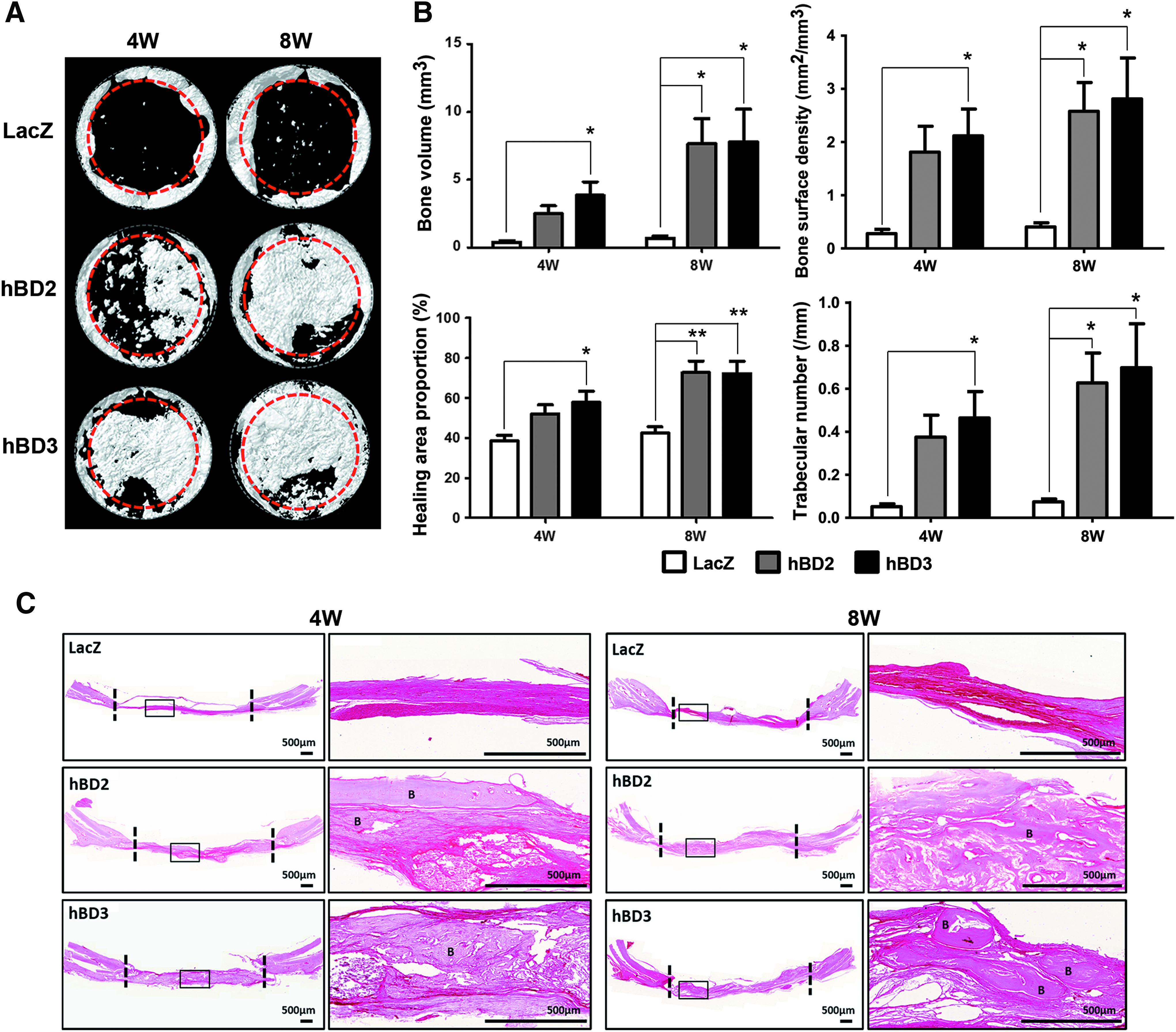
BMSC/hBD2 and BMSC/hBD3 promote bone regeneration in the S. aureus-contaminated calvarial defect model. Bone specimens were evaluated by μCT measurement and histological examination 4 and 8 weeks after surgery.
Discussion
Bacterial contamination during bone healing often compromised the outcome of bone regenerative therapy. hBD2 and hBD3 are important AMPs without significant bacterial resistance problem. In this study, we established a S. aureus-contaminated rat calvarial defect model and demonstrated that the implantation of bone marrow mesenchymal stromal cells overexpressing hBD2 or hBD3 could effectively mitigate the negative effects of bacterial contamination on bone healing. To our knowledge, this is the first study applying AMPs in the treatment of bone defect with bacterial contamination.
BMSC-based adenoviral gene therapy strategy was chosen to deliver the AMPs in this study. In most bone regenerative studies, recombinant proteins/peptides are administered in a one-time high dosage manner. However, the therapeutic effects are often limited due to short half-life of the proteins/peptides in vivo. Furthermore, the initial supraphysiological dose may result in unwanted side effects. Recombinant adenoviral vectors can efficiently infect BMSCs. Furthermore, the transduced cells can express the transferred gene for up to 21 days. 25
In this study, the antimicrobial assays were performed both in vitro and in vivo. The in vitro assay determined the antimicrobial activity of the CMs from BMSCs without S. aureus stimulation. The results indicated that the secretion of BMSC/LacZ did not show significant antimicrobial activity. The 50-fold concentrated CMs of BMSC/hBD2 and BMSC/hBD3 only showed a moderate antimicrobial effect (Fig. 2). It may be due to the dilution of the secreted defensins by culture medium and a short treatment time (2 h) on S. aureus. In contrast, an excellent antimicrobial effect in the hBD2 and the hBD3 groups was shown in the diffusion chamber model (Fig. 3C) and the bacteria-contaminated calvarial defect model (Fig. 6B). It may have resulted from the accumulation of the defensins continuously secreted from the hBD gene-modified BMSCs and a longer interaction time with the implanted S. aureus (24 or 72 h).
Our diffusion chamber model can prevent the escape of implanted S. aureus and BMSCs from the chamber. Furthermore, it also blocked the entry of host immune cells into the chamber. Therefore, the strong antimicrobial effect of the BMSC/hBD2 and BMSC/hBD3 (Fig. 3C) in the diffusion chamber may be mainly due to the secreted hBD2 and hBD3. In contrast, the viable bacterial counts of the control group were increased after 24 and 72 h. It indicated that the secretion of the BMSC/LacZ under S. aureus stimulation cannot effectively withhold the growth of bacteria in the diffusion chamber.
The antimicrobial effect demonstrated in the S. aureus-contaminated rat calvarial defect may be derived from both the implanted BMSCs and the rat immune response. Besides the direct antimicrobial effect, hBD2 and hBD3 might promote the rat adaptive immune response. It was reported that hBD2 and hBD3 can chemoattract immature dendritic cells, CD45RO+ CD4+ T cells, monocytes, macrophages, and neutrophils.26,27 In addition, hBD3 activates professional antigen-presenting cells through Toll-like receptors 1 and 2. 28 The viable S. aureus number after 3 days increased in the control group; this indicated that BMSC/LacZ and the rat immune response were unable to successfully control the S. aureus contamination in the early stage of bone healing. The absence of histological signs of inflammation or infection at 4 and 8 weeks in the LacZ group (Fig. 7C) may be the result of the effective antimicrobial effects of rat immune system at the later stage of bone healing. In contrast, the significant decrease in viable bacterial numbers of the hBD2 and the hBD3 groups suggested that BMSC/hBD2 and BMSC/hBD3 were effective against the contaminated S. aureus in the early stage of bone healing (Fig. 6B).
The antimicrobial effect against S. aureus was comparable for hBD2 and hBD3 in this study. BMSC/hBD2 and BMSC/hBD3 overexpressed a similar level of respective defensins (Fig. 1C). Their CM showed a similar antimicrobial effect in vitro (Fig. 2). Furthermore, the antimicrobial effect of BMSC/hBD2 and BMSC/hBD3 were not different in vivo (Figs. 3B, C, and 6).
Multiple mechanisms may be responsible for the negative effects of S. aureus contamination on bone healing (Fig. 4). S. aureus can stimulate bone resorption 29 and inhibit the synthesis of bone matrix protein. 30 The effects of S. aureus on osteoblast include inhibition of proliferation and mineralization, induction of apoptosis, and promotion of RANKL expression on osteoblast to enhance osteoclastogenesis. 31 In addition, BMSCs may secrete factors, such as interleukin (IL)-6, IL-1β, and tumor necrosis factor (TNF)-α, which are detrimental to bone healing after S. aureus stimulation. 32
The bone healing patterns of the experimental and the control groups are significantly different in the S. aureus-contaminated rat calvarial defect experiment (Fig. 7A). Bone formation seemed to be derived from two sources. One grew inwards from the defect margin and formed new bone mostly limited to the peripheral 1 mm ring area. In contrast, the new bone formation in the centered 6-mm diameter area may mostly result from the implanted BMSCs. The central bone formation of the hBD2 and the hBD3 groups were almost equivalent to that of the BMSC group (without S. aureus contamination) at 8 weeks (Figs. 4 and 7). It may be due to the rapid and excellent antimicrobial activity against S. aureus shown after implantation of BMSC/hBD2 and BMSC/hBD3 during the initial bone healing. In contrast, the minimal bone formation of the LacZ group may be due to the severe suppression of bone formation of BMSC/LacZ by high S. aureus count at the early stage of healing (Fig. 6).
The bone healing was significantly better in the hBD2 and the hBD3 groups when compared with the control group in the S. aureus-contaminated rat calvarial defect experiment. Multiple factors may contribute to this difference. First, viable S. aureus number in the defect was much higher in the control group when compared with the hBD2 and the hBD3 groups (Fig. 6). Previous reports indicated that use/application of stem cell therapies with the existence of infection may have a limited therapeutic effect. 33 BMSCs cocultured with S. aureus or treated with S. aureus CM significantly compromise their viability and osteogenic potential.33,34 The implanted BMSC/LacZ might not be able to survive and form new bone under the negative influences of S. aureus. The minimal new bone formation of the Empty group in Figure 4B suggested that the rat critical-sized calvarial defect heals poorly without the beneficial effects of the implanted BMSCs. Thus, the LacZ group showed very limited bone regeneration at 8 weeks, and even the pathogen might be cleared at 4 weeks, as evidenced by the lack of signs of inflammation and infection (Fig. 7C). Second, hBD2 and hBD3 may facilitate bone healing. hBD2 increases the proliferation of both primary osteoblasts, osteoblast-like cells (MG63 cells), and bone marrow mesenchymal stem cells. In addition, hBD2 and hBD3 can increase the osteogenic differentiation and mineralization of MG63 cells.35,36 Furthermore, hBD2 may also promote bone healing through its proangiogenic activity. 37
In conclusion, we established a rat S. aureus-contaminated calvarial defect model to simulate the compromised bone healing in bone defect with bacterial contamination. The model may be valuable for studies involving the bone healing in nonsterile bone defects. Furthermore, the negative effects of bacterial contamination on bone healing can be significantly mitigated after hBD2 and hBD3 antimicrobial therapy. The results of this study support the application of hBD2 and hBD3 in the promotion of bone healing in bacteria-contaminated bone defects.
Footnotes
Acknowledgments
This study was funded primarily by the Taipei Veterans General Hospital, Taipei, Taiwan [Grant No. V99E1-007] and the Ministry of Science and Technology of Taiwan (National Science Council) [Grant No. NSC 101-2314-B-010-034-]. The authors thank the Taiwan Mouse Clinic [Grant No. MOST 105-2325-B-001-010], which is funded by the National Research Program for Biopharmaceuticals (NRPB) at the Ministry of Science and Technology (MOST) of Taiwan for μCT technical support in the rat calvarial defect experiment.
Disclosure Statement
No competing financial interests exist.