
Abstract
Introduction:
Mesenchymal stem cells (MSCs) are considered a promising cell source for cartilage repair strategies due to their chondrogenic differentiation potential. However, their in vitro tendency to progress toward hypertrophy limits their clinical use. This unfavorable result may be due to the fact that MSCs used in tissue engineering approaches are all at the same developmental stage, and have lost crucial spatial and temporal signaling cues. In this study, we sought to investigate the effect of a spatial parathyroid hormone-related protein (PTHrP) signaling gradient on the chondrogenic differentiation of MSCs and progression to hypertrophy.
Methods:
Human bone marrow-derived MSCs were transduced with adenoviral vectors overexpressing PTHrP and seeded into fibrin-poly(ester-urethane) scaffolds. To investigate the effect of a spatial PTHrP signaling gradient, scaffolds were seeded with PTHrP-overexpressing MSCs positioned on top of the scaffold, with untransduced MSCs seeded evenly within. Scaffolds were cultured with or without 2 ng/mL transforming growth factor (TGF)-β1 for 28 days.
Results:
PTHrP overexpression increased glycosaminoglycan (GAG) production by MSCs irrespective of TGF-β1 treatment, and exerted differential effects on chondrogenic and hypertrophic gene expression when MSCs were cultured in the presence of a PTHrP signaling gradient. Furthermore, PTHrP-overexpressing MSCs were associated with an increase of endogenous TGF-β1 production and reduced total MMP-13 secretion compared to controls.
Conclusion:
The presence of a spatial PTHrP signaling gradient may support chondrogenic differentiation of MSCs and promote the formation of a more stable cartilage phenotype in tissue engineering applications.
Introduction
T
Commonly used protocols for the in vitro chondrogenic differentiation of MSCs involve high density three-dimensional (3D) culture systems with a defined medium containing members of the transforming growth factor (TGF)-β family.3,4 However, such strategies are limited due to the tendency of chondrogenically differentiated MSCs to express markers of hypertrophic chondrocytes following several weeks of culture in vitro, which recapitulates aspects of endochondral ossification and leads to an instable cartilage phenotype.5,6
Mature joints are composed of a clearly defined articular structure consisting of superficial, middle and deep zones, each associated with characteristic patterns of extracellular matrix structure and chondrocyte morphology and gene expression.7–9 Cellular crosstalk between superficial and deep zone chondrocytes has been proposed to play a role in the maintenance of a stable cartilage phenotype through regulating cartilage mineralization. 10 In contrast to cartilage in vivo, MSCs used in cartilage tissue engineering applications are all at the same developmental stage and have lost physiological spatial and temporal signals, which may govern the formation of stable articular cartilage. During skeletal development, the sequential proliferation and differentiation of chondrocytes is tightly controlled with multiple signaling pathways playing a vital role in regulating chondrocyte phenotype. 11
Parathyroid hormone-related protein (PTHrP) is a known regulator of chondrocyte maturation during endochondral bone development. 12 PTHrP is expressed by perichondrial cells and chondrocytes near the epiphyseal ends of growing bones, acting in a paracrine manner through binding to its receptor on neighboring chondrocytes.13,14 PTHrP maintains chondrocyte proliferation and delays chondrocyte hypertrophy through binding to the PTHrP receptor.15,16 Additionally, PTHrP is expressed postnatally by human articular chondrocytes in a hypoxia-dependent manner and positively regulates differentiated human articular chondrocyte phenotype.10,17
In light of its capacity to regulate chondrocyte maturation and hypertrophy, the ability of PTHrP to modulate chondrogenic differentiation of MSCs is an area under active investigation. The N-terminal fragment PTHrP(1–34) has been previously shown to attenuate chondrocyte hypertrophy. 18 However, constant application of soluble PTHrP was reported to attenuate the chondrogenic differentiation of MSCs. 19 In contrast to these findings, intermittent exposure of MSCs to PTHrP(1–34) has enhanced chondrogenic differentiation while reducing the expression of hypertrophic markers.20,21 Although current literature highlights the importance of temporal regulation of PTHrP signaling in chondrogenically differentiating MSCs, the impact of spatial PTHrP signaling on chondrocyte maturation, which more closely resembles physiological signaling cues in vivo, remains to be elucidated.
The aim of this study was to examine the effect of a spatial PTHrP signaling gradient on the chondrogenic differentiation of MSCs and progression to hypertrophy. Our findings highlight a chondro-inductive effect of PTHrP overexpression on MSCs, with differential effects of PTHrP signaling on MSC phenotype observed in the presence of a PTHrP signaling gradient. Additionally, PTHrP overexpression may enhance the paracrine activity of chondrogenically differentiating MSCs through increasing endogenous TGF-β1 production. In light of our findings, the presence of a spatial PTHrP signaling gradient may support chondrogenic differentiation of MSCs and promote the formation of a stable cartilage phenotype.
Materials and Methods
Isolation and culture of human MSCs
MSCs from four donors (52 ± 18 years) were isolated from bone marrow aspirates harvested during routine procedures and obtained with full ethical approval (KEK-ZH-NR: 2010-0444/0). MSCs were isolated based on their plastic adherence, plated at 3000 cells/cm2 and cultured in expansion medium composed of alpha minimum essential medium (Gibco, Carlsbad), 10% MSC screened foetal bovine serum (Pan Biotech, Aidenbach, Germany), 5 ng/mL basic fibroblast growth factor (Peprotech, Rocky Hill), and 1% penicillin/streptomycin (Gibco). MSCs cultured to passage 3 were used for all experiments.
Preparation of adenoviral vectors
First generation, E1 and E3 deleted, serotype 5 adenoviral vectors containing cDNA for human PTHrP(1–34) were prepared utilizing the pAdEasy system (Agilent Technologies, CA) as previously described 22 (Fig. 1). Adenoviral vectors were propagated in AD293 cells and purified by cesium chloride density centrifugation. Following overnight dialysis, the concentration of virus particles was determined by UV/Vis spectrophotometry at 260 nm.
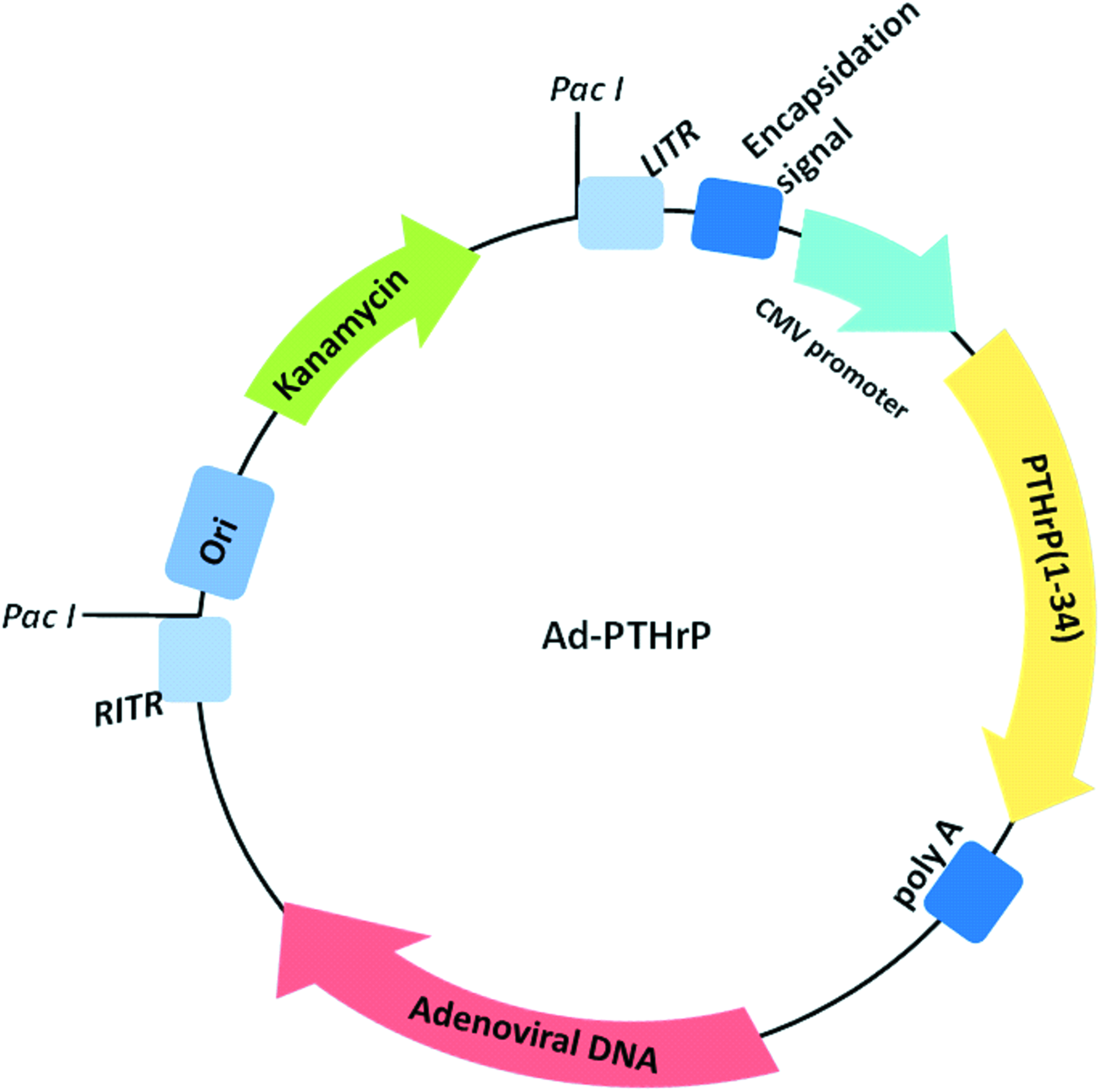
Schematic describing the adenoviral vectors utilized for adenovirus preparation. Adenoviral vectors containing cDNA for human PTHrP(1–34) were prepared utilizing the pAdEasy system (Agilent Technologies). Ad-PTHrP plasmid DNA was digested with Pac I to expose the ITR, and subsequently used to transfect AD293 cells for virus propagation. ITR, inverted terminal repeats; PTHrP, parathyroid hormone-related protein. Color images available online at www.liebertpub.com/tea
Adenoviral transduction of MSCs
Adenoviral transduction of MSCs in monolayer culture was performed utilizing the lanthanide-based method as previously described. 23 Calculated amounts of adenoviral vectors expressing PTHrP (1–34; AdPTHrP) were added to serum-free α-MEM medium to yield a final multiplicity of infection (MOI) of 20. Lanthanum(III) chloride heptahydrate (LaCl3; Sigma-Aldrich, Buchs, Switzerland) was dissolved in deionized water to generate a 0.4 M stock and further diluted in serum-free α-MEM medium to yield a 0.04 mM LaCl3 working solution.
An equal volume of LaCl3 working solution was added to the virus containing medium, mixed gently, and incubated at room temperature for 30 min. MSC expansion medium was aspirated from monolayer cultures and replaced with the LaCl3/virus solution. Following 3 h of incubation, MSCs were washed twice and received fresh expansion medium. MSCs were harvested for scaffold seeding at 24 h post-transduction. To achieve adenoviral transduction of MSCs in 3D, cells were untreated before scaffold seeding and adenoviral vectors were added directly to the MSC/fibrin mix during seeding to achieve a viral MOI of 10, as previously described. 24
Scaffold seeding and culture
Cylindrical poly(ester-urethane) scaffolds of 8 mm diameter and 2 mm high were prepared as previously described.25,26 Scaffolds were subjected to ethylene oxide sterilization and degassed before use. MSCs were seeded into 8 × 2 mm fibrin-poly(ester-urethane) composites (Baxter, Vienna, Austria) as described by Li et al. 27 The presence of AdPTHrP transduced MSCs and the position of MSCs within the scaffold varied according to experimental group (Fig. 2). To investigate the effect of PTHrP on MSC chondrogenesis, MSCs were transduced in 3D and seeded evenly throughout fibrin-poly(ester-urethane) composites at a cell density of 2 × 106 cells/scaffold. To vary the spatial signaling of PTHrP within cell-seeded constructs, 1.6 × 106 untransduced MSCs were seeded evenly within the construct with 4 × 105 pretransduced MSCs dropped on top of the scaffold as previously described. 26

Schematic describing the different seeding patterns applied for each experimental group. Untransduced (Group 1) or 3D-transduced (Group 2) MSCs were seeded evenly throughout fibrin-poly(ester-urethane) composites at a cell density of 2 × 106 cells/scaffold. To vary the spatial signaling of PTHrP within cell-seeded constructs, 1.6 × 106 untransduced MSCs were seeded evenly within the construct with 4 × 105 untransduced (Group 3) or pretransduced (Group 4) MSCs positioned on top of the scaffold. 3D, three-dimensional; MSC, mesenchymal stem cell. Color images available online at www.liebertpub.com/tea
Untransduced MSC-seeded constructs were included for each seeding strategy as a control as viral transduction under these conditions does not affect chondrogenesis (data not shown). All MSC-seeded scaffolds were cultured in chondropermissive medium composed of Dulbecco's modified Eagle's medium 4.5 g/L glucose (Gibco), 0.11 g/L sodium pyruvate, 50 μg/mL
Quantification of DNA content
MSC-seeded scaffolds were digested overnight at 56°C with 0.5 mg/mL proteinase K (PK) (Roche, Basel, Switzerland). Following digestion, samples were incubated for 10 min at 96°C for inactivation of PK. The DNA content of pellets was quantified using Höchst 33258 dye (Polysciences, Inc., Warrington, PA), using DNA calf thymus as a standard (LuBioScience, Luzern, Switzerland).
GAG measurement
GAG content of PK MSC-seeded scaffolds and harvested culture medium was determined spectrophotometrically at 530 and 590 nm following reaction with 1.9-dimethylmethylene blue (Polysciences, Inc.); using chondroitin 4-sulfate sodium salt from bovine trachea as a standard (Fluka).
Reverse transcription and polymerase chain reaction
MSC-seeded scaffolds were homogenized in TRI reagent (Molecular Research Centre, Inc., Cincinnati, OH) and RNA was isolated according to the manufacturer's instructions. Reverse transcription was performed using random hexamer primers and TaqMan reverse transcription reagents (Applied Biosystems, Carlsbad, CA). Quantitative real-time polymerase chain reaction (PCR) was performed in 10 μL reactions on cDNA using the Applied Biosystems QuantStudio 6 Flex Real Time PCR system (Applied Biosystems). Primers for collagen type II (COL2A1), collagen type X (COL10A1), aggrecan (ACAN), runt-related transcription factor 2 (RUNX2), and matrix metalloproteinase 13 (MMP13) were synthesized by Microsynth AG (Balgach, SG, Switzerland; Table 1). Gene expression assays for 18S ribosomal RNA (18S), SRY (sex determining region Y)-box 9 (SOX9), alkaline phosphatase (ALP), parathyroid hormone receptor 1 (PTH1R), and TGF beta 1 (TGFB1) were purchased from Applied Biosystems (Switzerland) (Table 2). Gene expression levels were normalized to 18S rRNA, and relative expression calculated via a ΔΔCT comparison.
Enzyme-linked immunosorbent assays
The levels of PTHrP released by MSCs during the entire culture period were determined by enzyme-linked immunosorbent assay (ELISA) of cell culture supernatant (Rabbit polyclonal anti-PTHrP [0.4 μg/mL, capture antibody] and biotinylated rabbit polyclonal anti-PTHrP [0.25 μg/mL, detection antibody]; Abcam, Cambridge, UK). The total amount of TGF-β1 in cell culture supernatant harvested during week 1 and 2 of culture was quantified using a human TGF-β1 DuoSet ELISA (R&D Systems, Minneapolis) according to the manufacturer's instructions. Levels of active TGF-β1 were quantified utilizing an optimized standard curve range (250–7.8 pg/mL) to achieve a lower limit of detection with this assay. The total amount of MMP-13 produced by MSCs at day 28 of culture was determined utilizing a human total MMP-13 ELISA (R&D Systems) according to manufacturer's instructions.
Quantification of ALP activity
Soluble ALP activity was determined in supernatant harvested at experimental day 28 in accordance with the Sigma Technical Bulletin Procedure No. 104 (Sigma-Aldrich). Cell culture supernatants were incubated with p-nitrophenyl phosphate substrate solution for 15 min at 37°C, and the resulting p-nitrophenol production measured spectrophotometrically at 405 nm. ALP activity was normalized to the DNA content of each scaffold and expressed as nmol of p-nitrophenol produced per minute per microgram of total DNA.
Statistical analysis
All data sets were analyzed by a Kruskal–Wallis test followed by Dunn's multiple comparisons test, using GraphPad Prism software version 6. For all analyses, differences were considered statistically significant at p < 0.05.
Results
Temporal regulation of PTHrP overexpression by MSCs is mediated by 3D versus LaCl3-based adenoviral transduction
PTHrP overexpression by transduced MSCs was confirmed by ELISA analysis of cell culture supernatant harvested throughout the 28 day culture period. PTHrP was overexpressed by all adenovirally transduced groups over the entire culture period, with levels reaching a maximum at day 7 and lowering by day 28 of culture (Fig. 3). Additionally, temporal differences were observed between the levels of PTHrP produced by 3D-transduced MSCs (3D Trans.) and pretransduced MSCs (Trans. Top). Levels of PTHrP produced by pretransduced MSCs were significantly higher compared to untransduced controls at day 1 of culture, whereas levels produced by 3D-transduced MSCs did not significantly differ at this time point. Conversely, 3D-transduced MSCs produced significantly higher levels of PTHrP at day 14, 21, and 28 of culture compared to untransduced MSCs.
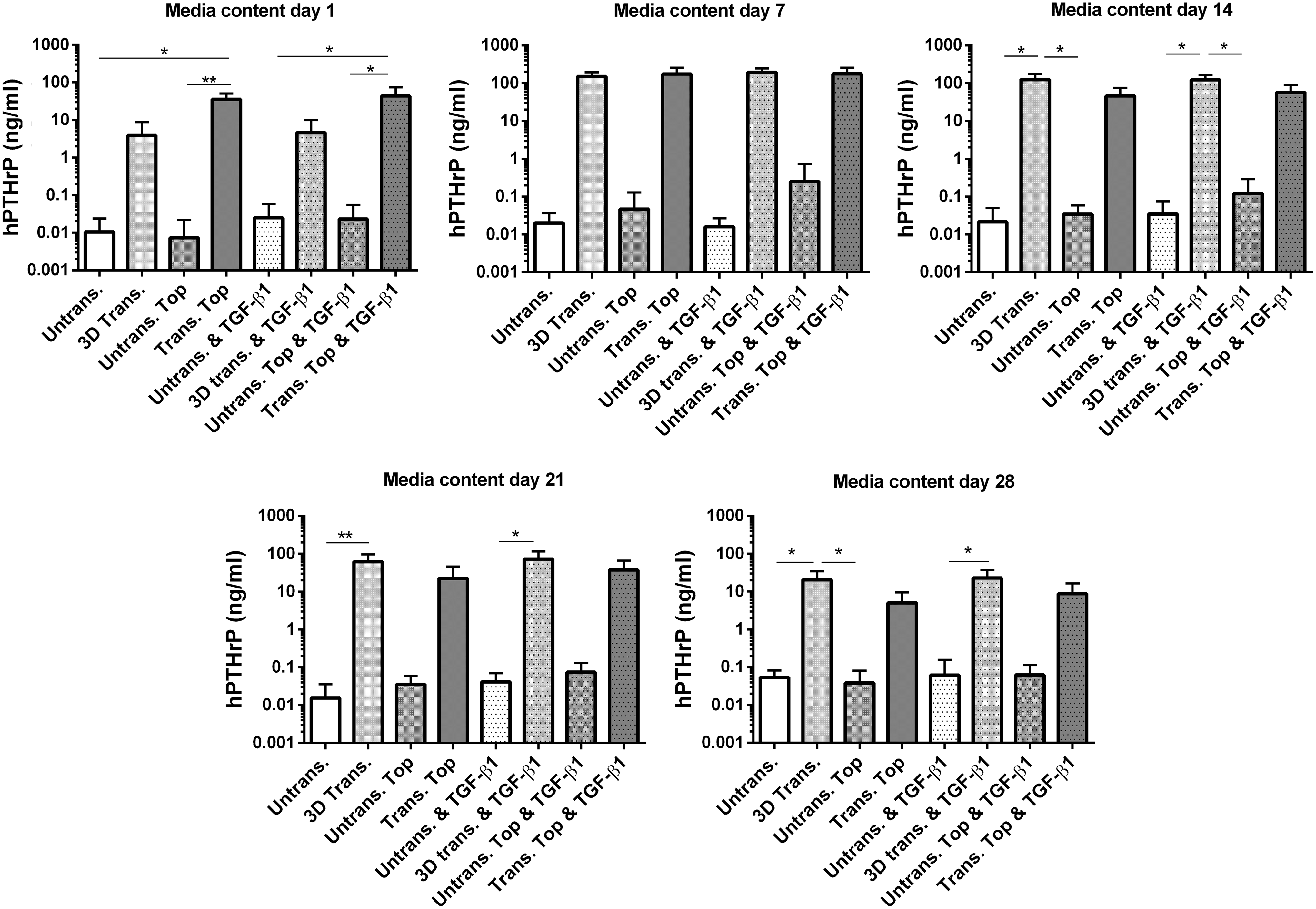
PTHrP expression levels by MSCs following adenoviral transduction. PTHrP production by MSCs at day 1, 7, 14, 21, and 28 of culture as determined by ELISA analysis of culture medium harvested at the experimental time point indicated. Values represent the mean ± SD of four donors, each assessed in experimental triplicate. Statistical significance was determined between groups at each experimental time point under evaluation by a Kruskal–Wallis test followed by Dunn's multiple comparisons test. *p < 0.05, **p < 0.005. ELISA, enzyme-linked immunosorbent assay; SD, standard deviation.
Treatment of MSC constructs with 2 ng/mL TGF-β1 did not significantly alter PTHrP production by either AdPTHrP-transduced or untransduced MSCs throughout the entire culture period.
PTHrP overexpression enhances endogenous TGF-β1 production by MSCs in a temporal manner
To investigate the effect of PTHrP overexpression and the presence of a PTHrP signaling gradient on the paracrine activity of MSCs, ELISA analysis was performed to quantify the levels of total and active TGF-β1 released into the culture medium throughout the first and second week of culture when PTHrP overexpression levels were highest. MSC-seeded scaffolds containing a spatial PTHrP signaling gradient had significantly higher levels of both total and active TGF-β1 in the culture media from experimental week 1 (Fig. 4) in the absence of exogenous TGF-β1 treatment. This corresponds to the same time point when PTHrP release levels by this group had reached a maximum (Fig. 3). Conversely, 3D-transduced scaffolds produced significantly higher levels of TGF-β1 during week 2 of culture compared to untransduced controls, at the same time point when PTHrP production levels were higher by this group, highlighting temporal regulation of TGF-β1 production in response to PTHrP.
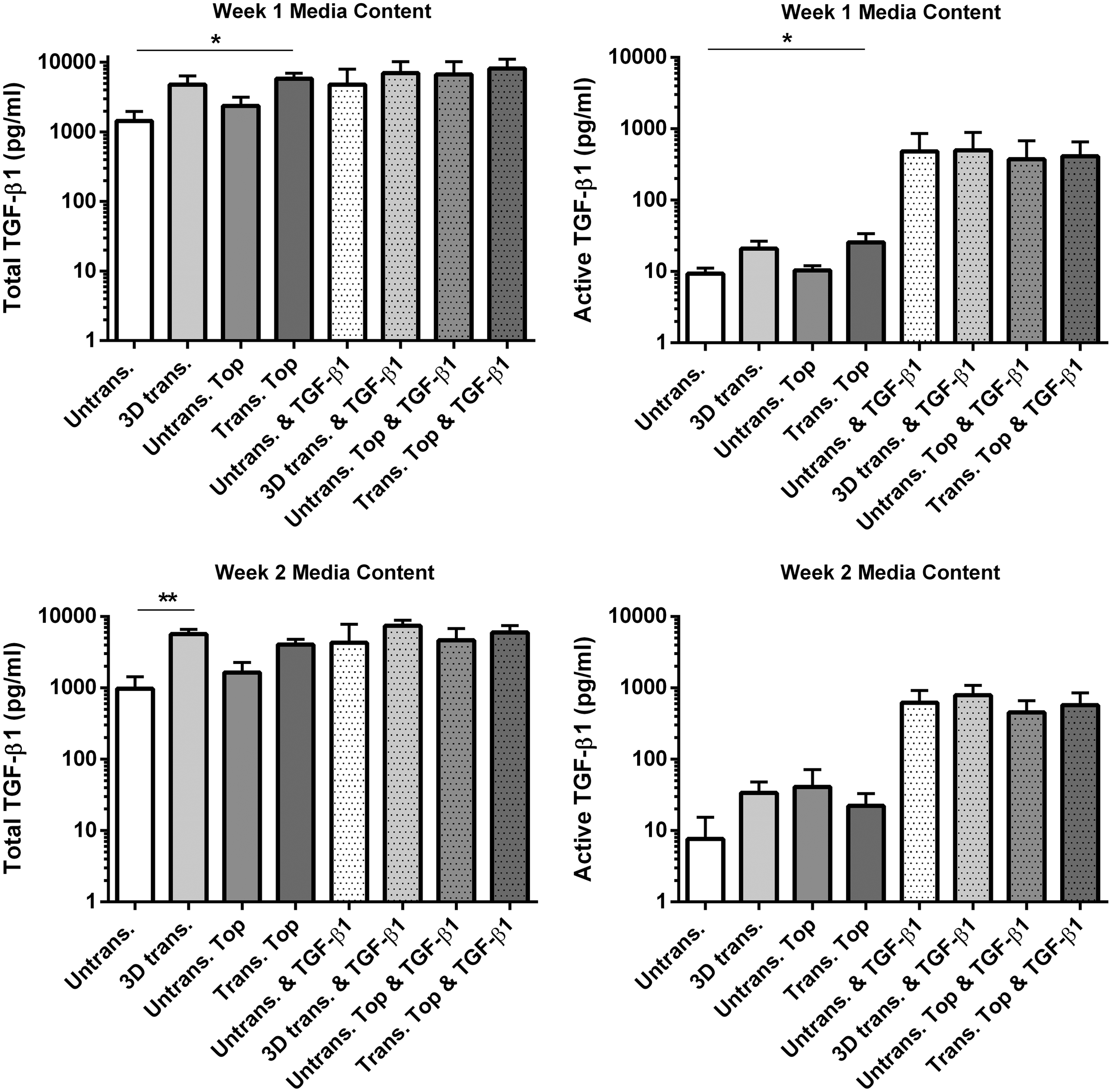
PTHrP overexpression enhances endogenous TGF-β1 production by MSCs. Levels of TGF-β1 produced by MSCs during week 1 and 2 of culture as quantified by ELISA. Values represent the mean ± SD of four donors, each assessed in experimental triplicate. Statistical significance was determined by a Kruskal–Wallis test followed by Dunn's multiple comparisons test. *p < 0.05, **p < 0.005. TGF, transforming growth factor.
PTHrP overexpression increases chondrogenic gene expression by MSCs while decreasing markers of hypertrophy, at day 28 of culture
Three-dimensional-transduced MSCs had significantly higher gene expression levels of the chondrogenic markers ACAN and SOX9 PTHR1 at day 28 compared to untransduced controls, in the absence of exogenous TGF-β1 stimulation (Fig. 5A). Furthermore, 3D-transduced MSCs had significantly decreased levels of the hypertrophic marker MMP13 compared to controls when scaffolds were cultured in the presence of 2 ng/mL TGF-β1. A similar trend of an increase in chondrogenesis-related markers was observed for scaffolds containing transduced MSCs positioned on top, however, gene expression values were not statistically significant. MSC-seeded scaffolds containing a PTHrP signaling gradient had significantly lower levels of the hypertrophic marker COL10A1 when cultured in the presence of TGF-β1 compared to the control.

PTHrP overexpression increases chondrogenic gene expression by MSCs while modulating hypertrophic marker expression.
Gene expression levels of TGF-β1 and the hypertrophy-related marker RUNX2 did not significantly differ between any of the experimental groups at this time point. However, PTHrP overexpression by 3D-transduced scaffolds significantly increased ALP gene expression compared to controls, irrespective of TGF-β1 treatment. Conversely, this stimulatory effect of PTHrP on ALP gene expression was not observed in scaffolds containing a PTHrP signaling gradient. Three-dimensional-transduced MSCs were associated with a significant increase in the ratio of aggrecan to collagen type X compared to controls, in the absence of TGF-β1 stimulation (Fig. 5B). In experimental groups containing exogenous TGF-β1, MSCs exposed to a PTHrP signaling gradient had a significantly increased aggrecan to collagen type X ratio compared to untransduced MSCs. Collagen type II gene expression was undetectable in MSC donors 1 and 2 irrespective of exogenous TGF-β1 stimulation (Fig. 5C). However, collagen type II message was detected in donors 3 and 4 in groups containing 3D-transduced and pretransduced MSCs in response to PTHrP stimulation alone. Stimulation of MSCs with 2 ng/mL TGF-β1 was sufficient to induce collagen type II gene expression in donors 3 and 4, irrespective of PTHrP overexpression.
PTHrP overexpression supports chondrogenic differentiation and formation of a more stable phenotype at day 28 of chondrogenesis
The DNA content of scaffolds did not significantly differ between experimental groups at day 28 of culture (Fig. 6A). Three-dimensional-transduced MSCs had significantly higher levels of GAG secreted in to the culture medium compared to untransduced control, irrespective of treatment with 2 ng/mL TGF-β1 (Fig. 6B). Furthermore, scaffolds containing a PTHrP signaling gradient had a higher level of GAG retention in the scaffolds compared to the untransduced and 3D-transduced groups. Due to the large variation observed between donors in this group this finding was not statistically significant. Additionally, PTHrP overexpression by 3D-transduced MSCs significantly inhibited total MMP-13 protein expression by MSCs treated with TGF-β1 (Fig. 6C). In a similar manner to ALP gene expression, PTHrP overexpression by 3D-transduced MSCs statistically increased ALP activity irrespective of TGF-β1 treatment (Fig. 6D). This effect was not observed with scaffolds containing a spatial PTHrP signaling gradient.
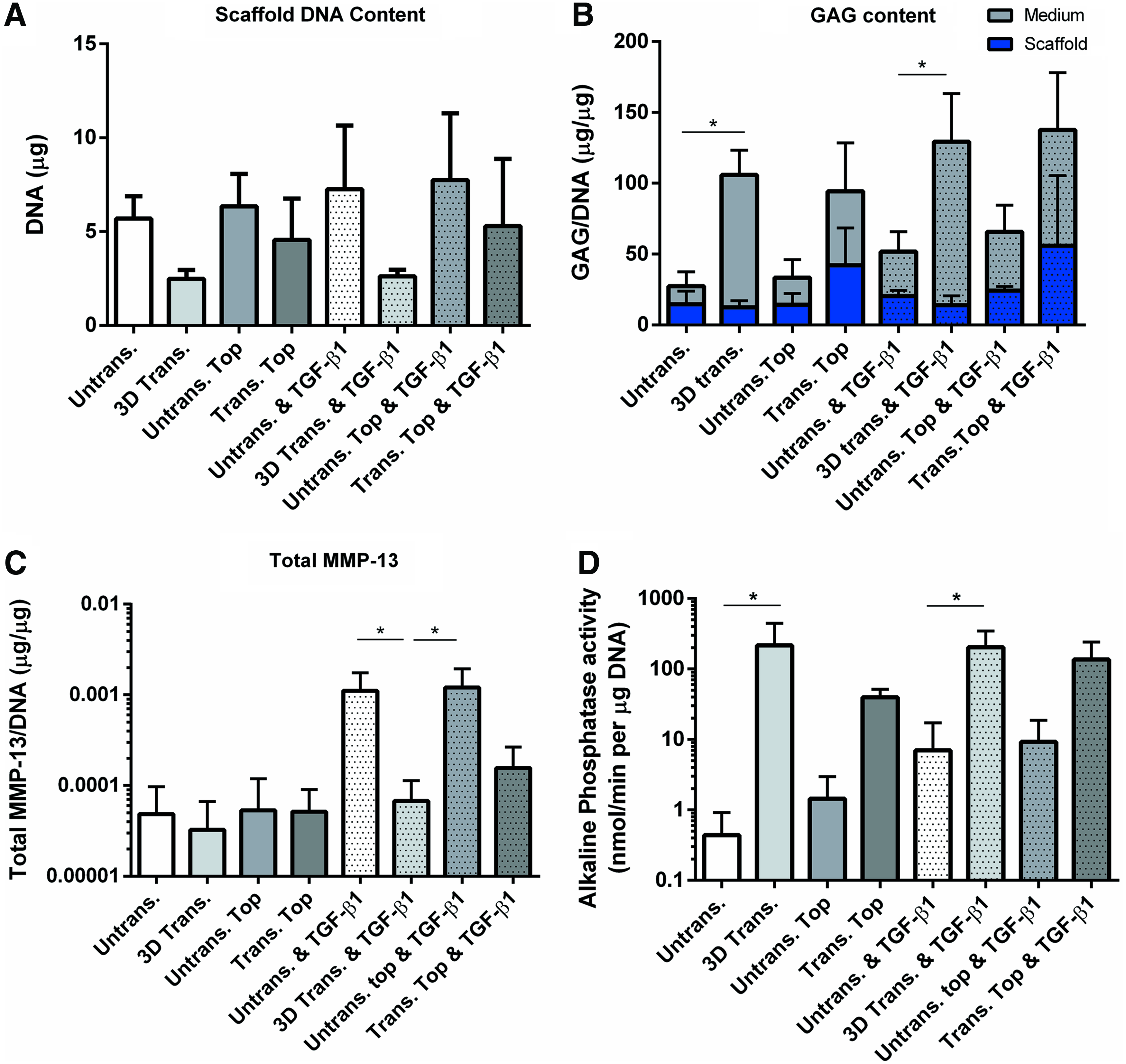
PTHrP overexpression increases GAG production by MSCs and modulates MMP-13 and ALP secretion at day 28 of chondrogenesis.
Discussion
In light of its capacity to regulate chondrocyte maturation and hypertrophy, the ability of PTHrP to modulate chondrogenic differentiation of MSCs is an area under active investigation. Current literature has highlighted the importance of temporal regulation of PTHrP signaling in chondrogenically differentiating MSCs.20,21 Although cellular communication between cartilage zonal layers via deep zone chondrocyte expression of PTHrP is known to maintain articular cartilage phenotype postnatally, the effect of spatial PTHrP signaling on MSC chondrogenesis has yet to be described. 10 The aim of this study was to examine the effect of a spatial PTHrP signaling gradient on chondrogenic differentiation and hypertrophic marker expression by MSCs. In this study we report that PTHrP overexpression enhances MSC chondrogenesis, with differential effects of PTHrP signaling on MSC phenotype observed in the presence of a PTHrP signaling gradient. Additionally, PTHrP overexpression enhanced endogenous TGF-β1 production by MSCs. In light of our findings, the presence of a spatial PTHrP signaling gradient may support chondrogenic differentiation of MSCs and promote the formation of a more resting chondrocyte phenotype.
PTHrP is a key regulator of chondrocyte differentiation in the growth plate, forming a feedback loop with Indian hedgehog (IHH), which is produced by prehypertrophic chondrocytes and stimulates PTHrP expression, preventing further chondrocyte hypertrophy. 29 Although the vital role of PTHrP and IHH in regulating chondrocyte maturation has been well established, additional reports have highlighted TGF-β as an intermediary factor regulating PTHrP expression in the periarticular region and in epiphyseal chondrocytes.30,31 In light of this previously established interplay between PTHrP and TGF-β signaling in chondrocytes, we sought to investigate the potential relationship between PTHrP and TGF-β expression by MSCs. In this study, stimulation of untransduced MSCs with 2 ng/mL of TGF-β1 was not sufficient to significantly enhance endogenous production of PTHrP.
Our findings illustrate that PTHrP stimulation alone increases endogenous production of TGF-β1 protein by MSCs. This stimulatory effect of PTHrP on TGF-β1 production was observed directly in response to PTHrP concentration, which varied in experimental timing according to the adenoviral transduction strategy applied. TGF-β is secreted by cells in an inactive, latent state bound to the latency-associated peptide (LAP). The release of TGF-β from LAP is required for TGF-β to bind to and activate its target receptor. 32 In this study, active TGF-β was only detected in cell culture media harvested from scaffolds containing a PTHrP signaling gradient during week 1 of culture.
Recent reports have suggested that supplementation of cell culture media with latent TGF-β1 leads to more uniform cartilage matrix deposition within chondrocyte-seeded tissue engineering constructs, compared to constructs cultured with active protein. 33 Albro et al. have shown that supplementation of cell culture media with exogenous active TGF-β results in the formation of concentration gradients within such constructs, leading to heterogenous cartilage formation. 33 Whether the increase of TGF-β1 production by PTHrP-overexpressing MSCs observed in our study may lead to cell-mediated activation and the formation of more uniformly distributed matrix within MSC-seeded constructs requires further investigation.
The effect of several isoforms of PTHrP on MSC chondrogenesis has been previously investigated, with PTHrP(1–34) identified as the superior isoform to enhance collagen type II gene expression while inhibiting expression of the hypertrophic markers collagen type X and ALP by MSCs. 34 However, current data are conflicting on whether PTHrP may enhance chondrogenesis and modulate hypertrophic differentiation. Weiss et al. have reported that PTHrP(1–34) inhibits both collagen type II and X expression by chondrogenically differentiating MSCs, when added continuously throughout the culture period. 19 Conversely, other groups have observed an increase in GAG and collagen type II and a decrease in hypertrophic marker expression by MSCs following 2 weeks of treatment or intermittent exposure to PTHrP.21,35
However, all of these aforementioned studies have assessed the ability of PTHrP to promote extracellular matrix deposition in the presence of chondro-inductive media containing TGF-β protein. In line with the findings of Weiss et al., PTHrP overexpression alone was not sufficient in our study to enhance the expression of collagen type II even in the presence of TGF-β. 19 We did observe a significant increase in gene expression levels of aggrecan, SOX9, and the PTHrP receptor by 3D-transduced PTHrP overexpressing MSCs compared to untransduced controls, in the absence of exogenous TGF-β1 stimulation. The PTHrP receptor is not expressed by undifferentiated MSCs, and it is upregulated during chondrogenic differentiation. 36
Further, 3D-transduced MSCs significantly increased GAG production by MSCs, in the absence of additional chondrogenic stimuli. These findings highlight a potential chondro-inductive effect of PTHrP-overexpression on MSC chondrogenesis, in the absence of additional stimuli. However, in contrast to previous studies describing an inhibitory effect of PTHrP on ALP expression by chondrogenically differentiating MSCs,19,37 we have observed a significant increase in ALP in response to PTHrP overexpression by 3D-transduced scaffolds, irrespective of TGF-β1 treatment. However, PTHrP expression by 3D-transduced MSCs had an inhibitory effect on the hypertrophy marker MMP-13, which was upregulated by MSCs in response to TGF-β1 stimulation.
Our study has some limitations. First, all MSC-seeded scaffolds were cultured in vitro for 28 days to evaluate the effect of PTHrP expression on MSC chondrogenesis and hypertrophy-related marker expression. A longer culture period may be required to more accurately access the effect of PTHrP on hypertrophic differentiation of MSCs and the long-term stability of MSC phenotype. Furthermore, MSC-seeded scaffolds were cultured with 2 ng/mL of TGF-β1 to provide a low-dose chondro-inductive stimulus, without concealing any chondrogenic activity of PTHrP.
However, this low dose of TGF-β1 was not sufficient to induce collagen II expression in all four MSC donors. Although the other chondrogenic genes investigated behaved similarly across all four donors, two did not induce collagen II. Such data need to be highlighted as these provide an indication of donor variability and reproducibility. It also suggests that there are differential regulatory mechanisms between collagen II and other genes such as aggrecan, which is in line with other MSC chondrogenesis studies. 38 Therefore, culture of MSCs with a strong chondrogenic stimulus such as 10 ng/mL of TGF-β may be required to fully evaluate the effect of PTHrP on actively differentiating MSCs.
Finally, the scaffold system used is not optimal for histological evaluation as much of the newly synthesized matrix is released into the medium. 27 However, a scaffold system was required to produce a gradient, meaning the commonly used pellet culture model was not possible. Whether the presence of a spatial PTHrP signaling gradient may have influenced the spatial organization of extracellular components requires further investigation.
In summary, we have demonstrated induction of MSC chondrogenesis in response to PTHrP overexpression in the absence of exogenous TGF-β stimulation. Furthermore, the presence of a spatial PTHrP signaling gradient may support the formation of a more stable chondrocyte phenotype in actively differentiating MSCs. In addition to promoting chondrogenic differentiation, PTHrP overexpression may enhance the paracrine activity of MSCs through increasing TGF-β1 production, which may also be advantageous for cartilage tissue engineering applications.
Footnotes
Acknowledgments
This work was supported by the Swiss National Science Foundation (Grant No. 31003a_146375/1). The authors thank Dr. D. Eglin (Musculoskeletal Regeneration Programme, AO Research Institute Davos, Switzerland) for producing the polyurethane scaffolds and Baxter Biosurgery, Vienna, Austria, for providing the fibrin components.
Disclosure Statement
No competing financial interests exist.