
Abstract
In an effort to construct in vitro liver tissue models for drug screening, human induced pluripotent stem cell (hiPSC)-derived hepatocyte-like cells (iHeps) are a promising cell source alternative to primary human hepatocytes (PHHs). However, thus far, the functions of iHeps are significantly reduced when compared to those of PHHs. In this study, we aimed to determine the role of oxygen supply to the cell layer, which has not been investigated as thoroughly as other conditions, such as soluble factors, the extracellular matrix, or co-culture with nonparenchymal cells, in improving the hepatic function of iHeps. Polydimethylsiloxane (PDMS), which has a high oxygen permeability, was used for cell culture [O2 (+)], in addition to conventional tissue culture-treated polystyrene [TCPS; O2 (−)]. During endoderm differentiation, O2 (+) culture resulted in reduced expression of HIF-1α. As a result, we observed higher expression of the endoderm marker (GATA4). After hepatic differentiation, iHeps generated in O2 (+) culture showed about a threefold increase per cell in albumin production. Moreover, among the O2 (+) culture conditions, cells differentiating at a physiological oxygen concentration showed increased expression of several mature hepatic marker genes (ALB, CYP3A4, and CYP1A2) and increased ratio of ALB-positive cells. In O2 (+) culture, iHeps exhibited a per cell oxygen consumption rate two times higher than that observed in O2 (−) culture. Furthermore, in sufficiently oxygen-supplied culture systems, cells showed a self-organized, multilayered configuration, and the DNA content in the culture was two to three times higher than that in O2 (−) culture. Considering the amount of DNA and the ration of iHeps in each culture condition, we could achieve about 7.5 times higher per unit surface area productivity of iHeps in O2 (+) culture. Therefore, these results demonstrate that direct oxygenation through a PDMS membrane enhances the maturation and efficient production of iHeps. Thus, improving oxygen supply to the cell layer is important for iHeps culture, and direct oxygenation through a PDMS membrane is a promising alternative to conventional TCPS plates for the maturation of iHeps in vitro.
Impact Statement
Although oxygen is a vital nutrient for the hepatocytes in vitro, few reports have focused on its effect during hepatic differentiation of induced pluripotent stem cells (iPSCs). In this report, we performed the hepatic differentiation of human iPSCs (hiPSCs) under different atmospheric oxygen concentrations and oxygen supply fluxes to investigate the effects of oxygen in terms of both the concentration and the supply flux. Results demonstrate that direct oxygenation through a polydimethylsiloxane (PDMS) membrane enhances the maturation and efficient production of hiPSC-derived hepatocyte-like cells (iHeps). Thus, direct oxygenation through a PDMS membrane is a better alternative culture method over conventional tissue culture-treated polystyrene (TCPS) plates for the maturation of hiPSC-derived hepatocytes in vitro.
Introduction
T
Many hepatic differentiation protocols of hiPSCs have been reported in the last decade. In these protocols, iHeps are generated by the sequential addition of growth factors that are known to play important roles in liver organogenesis.10–13 Although the iHeps generated using these protocols show some hepatic functions, such as albumin (ALB) production and cytochrome P450 (CYP) activity, these functions are significantly reduced compared to those in PHHs. 14 To tackle this problem, many researchers have sought to improve culture systems for the further maturation of iHeps, including co-culture1,15 or three-dimensional (3D) culture.16–18 Berger et al. used micropatterned co-culture with stromal fibroblasts and reported improvements in the gene expression of several mature hepatic markers, as well as ALB and urea synthesis and several CYP activities. 1 Gieseck et al. reported that iHeps cultured in the Real Architecture for 3D Tissues (RAFT) system showed significantly improved functional maturation compared to iHeps cultured in a conventional two-dimensional (2D) system. 17 Although iHeps cultured using these culture methods showed higher functional maturity compared to those derived from conventional culture systems, these cells still showed a fetal phenotype or reduced function compared to PHHs.
Because oxygen is essential during embryo development and liver organogenesis,19–21 the oxygen environment of the culture system is also an important factor in the differentiation of hiPSCs and maturation of iHeps in vitro. To enhance oxygen conditions in vitro, some efforts have focused on the atmospheric oxygen conditions. Si-Tayeb et al. reported a hepatic differentiation protocol in which they applied different atmospheric oxygen concentrations to each differentiation step. However, they did not discuss the advantage of applying different oxygen concentrations. 10 Moreover, although they focused on the atmospheric oxygen concentration, they did not control the amount of oxygen supplied to the cell layer, which is what cells utilize for their respiration and energy synthesis.
Oxygen supply to the cell layer in a conventional tissue culture-treated polystyrene (TCPS) plate depends only on diffusion through culture media. Because this diffusion-based oxygen supply is severely limited by the low solubility and diffusivity of oxygen, it is unable to meet the cellular oxygen demand. 22 Employing an oxygen-permeable membrane as the bottom of the culture plate is one of the simplest methods for enhancing oxygen supply to the cell layer. Xiao et al. reported that direct oxygenation through a polydimethylsiloxane (PDMS) membrane, which has a high oxygen permeability, could completely meet the cellular oxygen demands of rat hepatocytes in vitro. 24 Although many advantages of employing PDMS membranes at the culture plate bottom have been shown in the culture of hepatocytes (rat primary hepatocytes and human hepatoma HepG2 cells) according to previous reports,23–26 there have been few reports assessing the beneficial effects of direct oxygenation through a PDMS membrane on the hepatic differentiation of hiPSCs.
In this study, we performed the hepatic differentiation of hiPSCs using several different atmospheric oxygen concentrations in both TCPS and PDMS plates to investigate the effects of oxygen supply in terms of both the concentration at the cell layer and the oxygen supply flux. Comparisons of different conditions for differentiation revealed that the expression of hepatic genes, such as ALB, CYP3A4, and CYP1A2, was upregulated in cells differentiated on PDMS plates with physiological oxygen concentrations (5% or 10%). Interestingly, cells differentiated under low oxygen supply conditions exhibited significantly reduced hepatic functions, implying that hepatic differentiation failed under a low oxygen supply. Furthermore, in the PDMS culture system, cells exhibited a self-organized, multilayered configuration, and the DNA content of the culture was two to three times higher than that in TCPS culture. These results clearly demonstrate that improving oxygen supply to the cell layer using PDMS membranes is key to improving the hepatic differentiation of hiPSCs.
Materials and Methods
High- and low-oxygen cell culture
To enable direct oxygenation through PDMS membranes, hiPSCs (TkDN4-M), provided by the Center for Stem Cell Biology and Regenerative Medicine at the University of Tokyo, were cultured on commercially available polycarbonate 24-well plates assembled with a 0.35-mm thick PDMS membrane (Vessel) [O2 (+) condition]. In the low-oxygen control condition [O2 (−)], hiPSCs were cultured on PDMS discs inserted into the wells of a conventional TCPS plate (Iwaki) to keep the same surface characteristics both in O2 (+) and O2 (−) culture conditions (Fig. 1). Before cell inoculation, the PDMS surfaces were treated with oxygen plasma for 60 s using a YHS-GZA 200 (SAKIGAKE-Semiconductor) and coupled with 3-mercaptopropyltrimethoxysilane (Shinetsu Silicone). Amino groups introduced were subsequently reacted with a cross-linker, N-(4-maleimidobutyryloxy) succinimide (GMBS; Dojindo), activated by 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide, hydrochloride (WSC; Dojindo), and N-hydroxysulfosuccinimide sodium salt (sulfo-NHS; TCI). Finally, the PDMS surfaces were coated with Matrigel (Corning) to allow cells to attach. hiPSCs were inoculated and cultured on Matrigel-coated O2 (−) and O2 (+) plates with mTeSR1 medium (Stem Cell Research).

Schematic illustration of cell culture on PDMS discs inserted into the wells of TCPS plates [O2 (−), left] or on PDMS plates [O2 (+), right]. PDMS, polydimethylsiloxane; TPCS, tissue culture-treated polystyrene.
Hepatic differentiation of hiPSCs
Using 90% confluent hiPSCs, hepatic differentiation was induced using a four-step protocol as previously reported 10 with minor modifications 27 : endodermal differentiation (step 1), specified hepatic cell differentiation (step 2), hepatoblast differentiation (step 3), and hepatic differentiation (step 4). Briefly, hiPSCs reached 90% confluence were cultured with RPMI medium supplemented with B-27 supplement (Gibco) and 100 ng/mL Activin A (R&D) for 5 days to differentiate into endodermal cells. After that, endodermal cells were cultured with RPMI medium supplemented with B-27 supplement, 10 ng/mL b-FGF (Peprotech), and 20 ng/mL BMP-4 (Humanzyme) for 5 days to induce hepatic progenitor differentiation. Then, cells were cultured with RPMI medium supplemented with 20 ng/mL HGF (Peprotech) for 5 days to induce hepatic differentiation. Finally, cells were cultured with hepatocyte culture medium (HCM; Lonza, without EGF) supplemented with oncostatin M (Peprotech) for 5 days to induce hepatic maturation. To evaluate the effect of oxygen supply during differentiation, we performed hepatic differentiation at various atmospheric oxygen concentrations (20%, conventional culture condition; 5% and 10%, physiological oxygen condition) under both O2 (−) and O2 (+) culture conditions (Table 1). We used cells differentiating in O2 (−) plates at 20% atmospheric oxygen as the control group, as these conditions were used in the original protocol. 10
Atmospheric Oxygen Concentration of Each Culture Condition During Hepatic Differentiation
Immunostaining
Cells were fixed with 4% paraformaldehyde at room temperature for 30 min and then incubated with 1% Triton-X100 (WAKO) for permeabilization. The cells were further incubated with 1% bovine serum albumin for blocking, followed by incubation with primary antibody overnight at 4°C. Cells were subsequently incubated with secondary antibody for 3 h and then counter-stained with 4′,6-diamidino-2-phenylindole (DAPI; Dojindo) for 3 min. The cells were rinsed with phosphate-buffered saline thrice for 5 min each after each step was finished. Fluorescent images were taken using a confocal microscope (PowerIX70; Olympus). Antibodies used for immunostaining are shown in Table 2. The ratio of marker protein-positive cells to DAPI-positive cells was manually counted using Image J (NIH) from three different areas of each sample.
List of Antibodies Used for Immunostaining
Real-time quantitative polymerase chain reaction
Total RNAs were isolated and purified from samples using TRIzol Reagent (Life Technologies) according to the manufacturer's instructions. The concentration and quality of each RNA sample were assessed with the NanoDrop system (BioSpec-nano; SHIMADZU). Subsequently, complementary DNAs (cDNAs) were reverse-transcribed from 0.5 μg total RNA using Rever Tra Ace qPCR Master Mix with gDNA Remover (TOYOBO) according to the manufacturer's protocol. Real-time quantitative polymerase chain reaction (qPCR) assays were performed with THUNDERBIRD SYBR qPCR Mix (TOYOBO) and assessed using the StepOnePlus qPCR system (Applied Biosystems). Primer sequences used for real-time qPCR are shown in Table 3. β-Actin (bACT) expression was used as the internal control.
List of Real-Time Quantitative Polymerase Chain Reaction Primers
Measurement of albumin production
The culture medium was changed to fresh HCM (Lonza) on day 20, the last day of differentiation, and differentiated cells were cultured for 24 h. After 24 h of incubation, the culture supernatant was collected for sandwich-type enzyme-linked immunosorbent assay to determine the amount of secreted albumin. Goat anti-human albumin antibody (Bethyl) was used as the capture antibody, horseradish peroxidase-conjugated goat anti-human albumin (Bethyl) was used as the detection antibody, and human reference serum (Bethyl) was used as the standard human albumin solution. The absorbance was measured by the microplate reader iMark (Bio-Rad) at 450 nm with an optical correction of 630 nm.
Measurement of DNA amount
Cells were detached by trypsin-EDTA treatment on day 20 and were homogenized with an Ultra Sonic Homogenizer UH-300 (SMT) before DNA measurement. DNA content was measured according to previously reported protocols. 28 DAPI was used as a fluorescent dye, and the intensity was assessed with the RF-5300PC spectrofluorophotometer (SHIMADZU). The excitation wavelength was set at 350 nm, and the emission wavelength at 450 nm.
Measurement of glucose production and lactate consumption
Culture supernatant was collected and stored at −20°C after every medium change. The concentrations of glucose and lactate in the samples were measured with the YSI 2950D Biochemistry Analyzer (YSI).
Measurement of oxygen tension
Oxygen concentrations at the cell layer were measured with a fluorescent oxygen probe (Firesting; Pyro Science) on day 20 as previously described. 24 Briefly, the probe was put about 30 μm above the cell layer. The concentrations were monitored continuously and read after they reached their steady state.
Statistical analysis
All data are represented as the mean ± standard deviation. Student's t-tests were used to determine the statistical significance of two experimental groups. Differences with p < 0.05 were considered to be significant.
Results
Direct oxygenation through PDMS enhanced endoderm differentiation in hiPSCs
To evaluate the effect of oxygen supply on endoderm differentiation in hiPSCs, endodermal cells generated under each condition were stained by labeling GATA binding protein 4 (GATA4), a known endodermal marker (Fig. 2A). Almost all cells differentiating at O2 (−)-20% and O2 (+)-5%, O2 (+)-10%, and O2 (+)-20% expressed GATA4 (89.4% ± 9.6%, 81.4% ± 16.0%, 87.6% ± 3.7%, and 77.2% ± 20.4%, respectively) (Table 4), indicating that these cells properly differentiated toward the endodermal lineage. In contrast, the number of cells expressing GATA4 decreased under O2 (−)-5% and O2 (−)-10% conditions (13.2% ± 9.1% and 54.3% ± 18.3%) (Table 4). Notably, almost all cells differentiating at O2 (−)-5% did not express GATA4. This result indicates that reduced oxygen supply to the cell layer inhibited the endodermal differentiation of hiPSCs.

The Ratio of GATA4-Positive Cells to DAPI-Positive Cells on Day 5
DAPI, 4′,6-diamidino-2-phenylindole.
Next, we performed real-time qPCR analysis to confirm the gene expression levels of both octamer binding transcription factor 4 (OCT4) and GATA4 in cells differentiating in each culture condition (Fig. 2B). When compared to cells differentiating at O2 (−)-20%, cells differentiating under lower oxygen supply conditions [O2 (−)-5% or O2 (−)-10%] showed higher expression of OCT4 and significantly lower expression of GATA4. In contrast, cells differentiating under O2 (+) conditions showed significantly higher expression of GATA4. However, there was no significant difference in the expression of OCT4 among cells differentiating at O2 (−)-20% and O2 (+)-5%, O2 (+)-10%, and O2 (+)-20%. The gene expression levels of OCT4 during differentiation at O2 (−)-20% decreased during endoderm differentiation, and this reduced expression was maintained to the last stage of differentiation (data not shown). The gene expression levels of OCT4 under these conditions may have reached the lower limit of expression, and thus, we were unable to observe significant differences. Based on these results, direct oxygenation through the PDMS membrane is effective for endoderm differentiation of hiPSCs.
Respiratory state and HIF-1α expression in endoderm cells
To determine the effect of direct oxygenation through PDMS on endoderm differentiation, we first determined the respiratory states of differentiating cells. The glucose and lactate concentrations in the culture supernatants on day 5 were measured, and the ratio of the lactate production (mol) to the glucose consumption (mol) was calculated (Fig. 3A). Cells differentiating under all conditions showed similar values close to 2 (mol/mol), showing that these cells were mostly performing anaerobic respiration. 23 For further evaluation, the gene expression of hypoxia inducible factor 1α (HIF-1α), which is highly expressed under hypoxic conditions, 29 was measured (Fig. 3B). As oxygen supply to the cell layer increased, the HIF-1α expression was reduced. These results confirm the relationship between HIF-1α expression and the endodermal differentiation of hiPSCs.
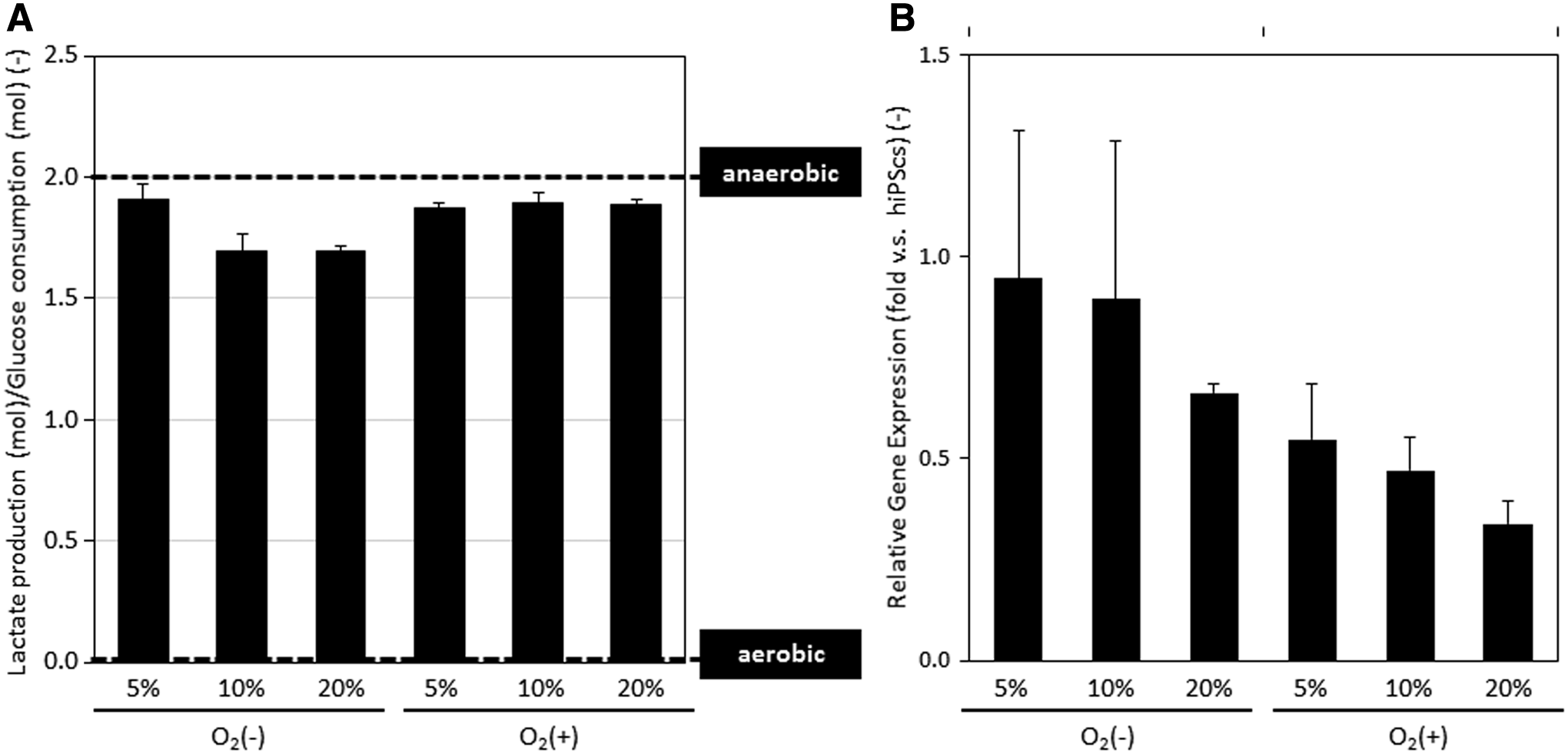
Cellular oxygen consumption was enhanced by direct oxygenation on day 20
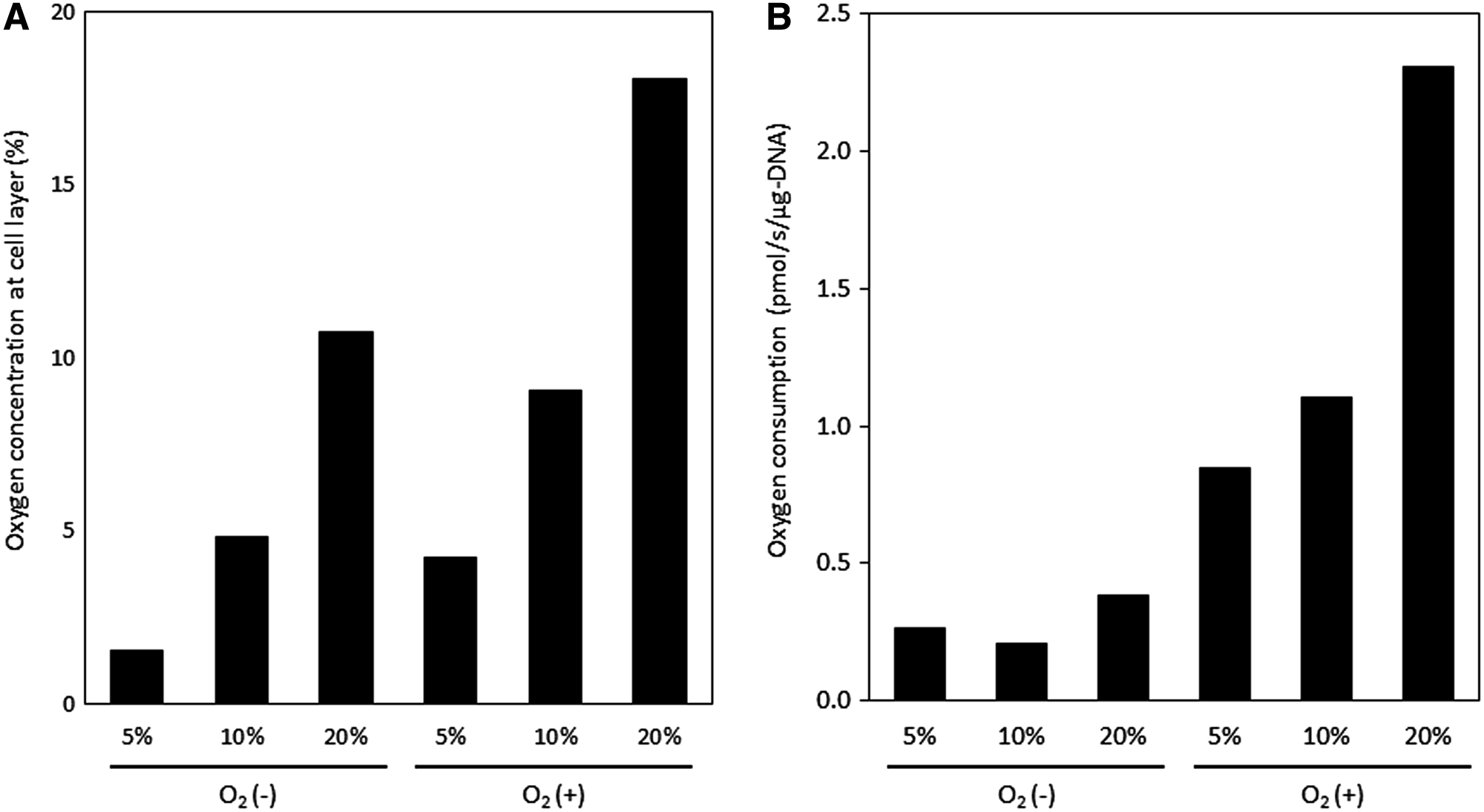
To investigate the effect of direct oxygenation in O2 (+) culture on iHep respiration, we measured the oxygen concentration above the cell layer on day 20 (Fig. 4A) and calculated the oxygen supply flux to the cell layer, which is the same value as the oxygen consumption rate (OCR), determined using the following equations [Eq. (1) for culture media, Eq. (2) for PDMS membrane]. The parameters used for the calculations are shown in Table 5.
Parameters That Were Used for Oxygen Consumption Rate Calculation
PDMS, polydimethylsiloxane.
The calculated OCRs are shown in Figure 4B. Cells differentiating under O2 (−) conditions consumed <0.5 pmol-O2/s/μg DNA. In contrast, cells differentiating at O2 (+)-5%, O2 (+)-10%, or O2 (+)-20% consumed oxygen at rates 2.2, 2.9, and 6.0 times higher, respectively, than the rate in cells at O2 (−)-20%. Thus, cells in O2 (+) culture utilize more oxygen for their respiration, suggesting that these cells synthesize energy more efficiently.
Physiological oxygen supply enhanced the hepatic differentiation of hiPSCs
To investigate the hepatic maturity of iHeps under each condition, cells were differentiated to iHeps subsequently to endodermal differentiation. Based on immunostaining results, we observed many AFP-positive cells in O2 (−)-20% and O2 (+)-5%, O2 (+)-10%, and O2 (+)-20% condition. In contrast, we observed more cells that were positive for ALB under O2 (+) conditions than at O2 (−)-20% (Fig. 5 and Table 6), indicating that hepatic maturation was highly induced in O2 (+) conditions. Moreover, when we compared cells differentiating under each O2 (+) culture condition, more cells were positive for ALB when differentiating at physiological oxygen concentrations (5% or 10%). In contrast, under O2 (−) conditions, cells differentiating at lower oxygen concentrations [O2 (−)-5% and O2 (−)-10%] tended to exhibit reduced ALB expression according to immunostaining than those differentiating at O2 (−)-20%. In particular, we did not detect any ALB-positive cells at O2 (−)-5%. These cells were also negative for alpha fetoprotein (AFP), indicating that these cells did not differentiate into hepatocytes.
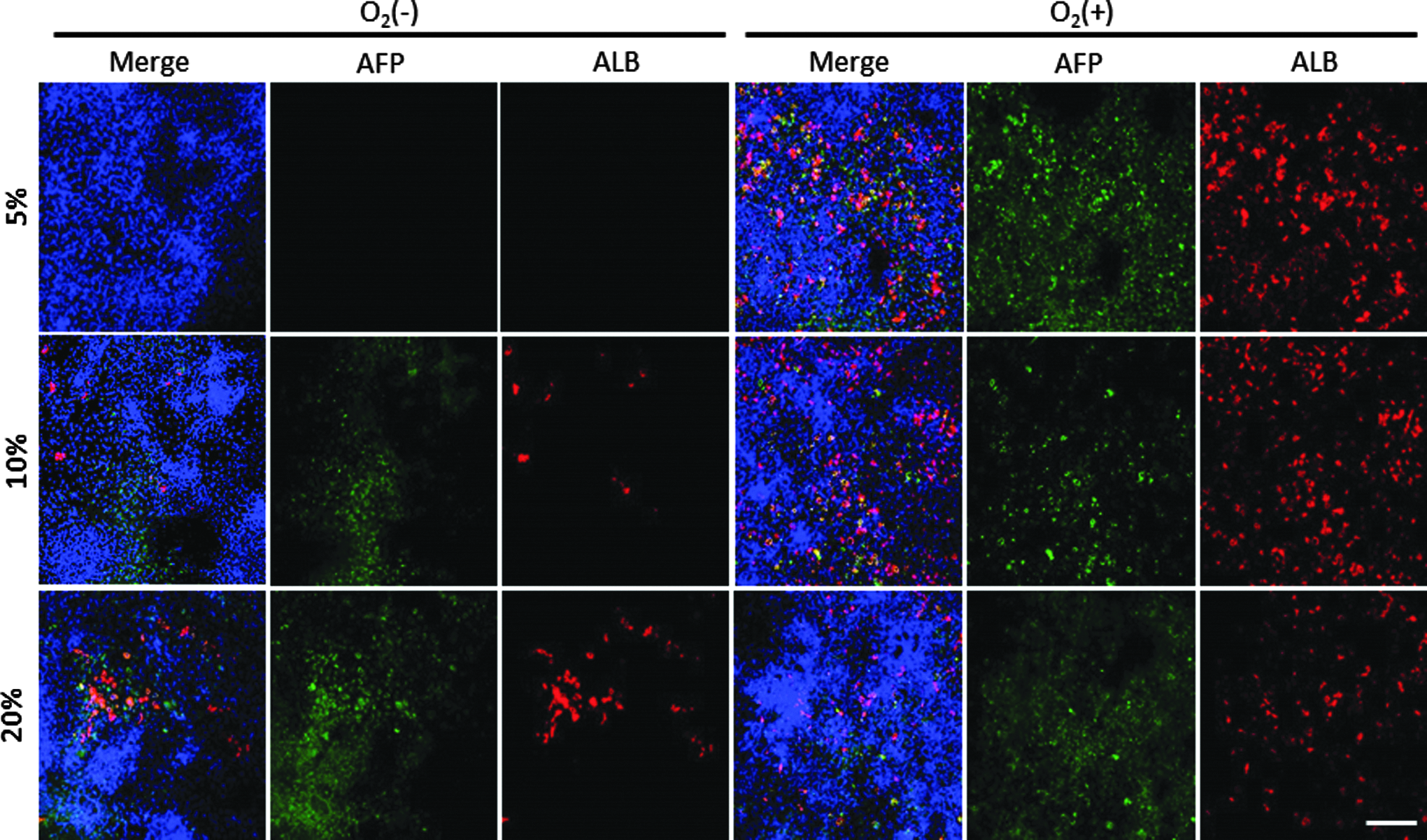
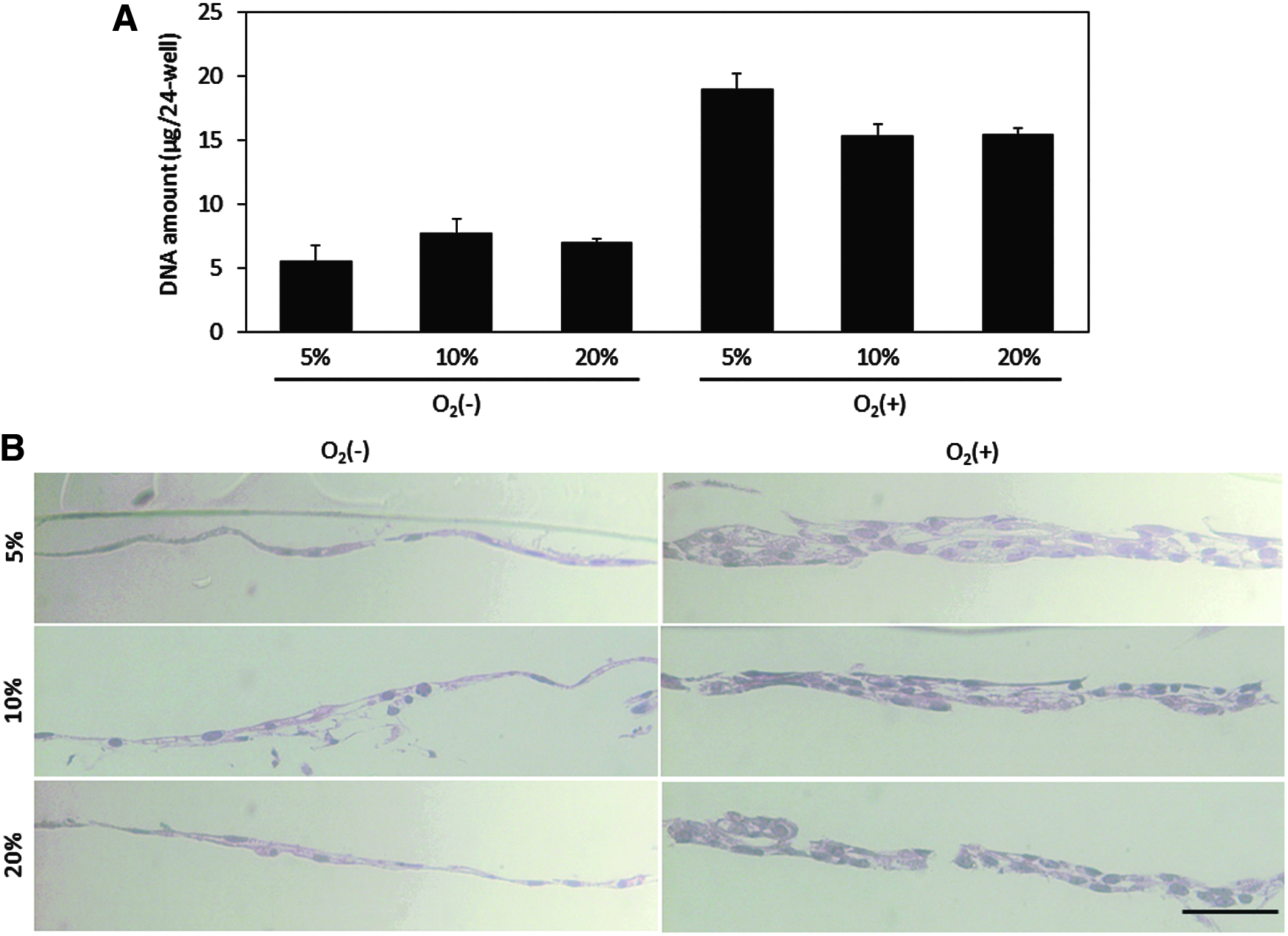
Immunohistochemical analysis of AFP and ALB in iHeps on day 20. Blue, DAPI; green, AFP; red, ALB. Scale bar represents 100 μm.
The Ratio of AFP-Positive or ALB-Positive Cells to DAPI-Positive Cells on Day 20
Next, we performed real-time qPCR to evaluate the gene expression levels of ALB (Fig. 6A), CYP3A4 (Fig. 6B), and CYP1A2 (Fig. 6C). Consistent with the immunostaining results, cells differentiating under O2 (+) culture conditions with lower atmospheric oxygen concentrations (5% O2) showed significantly higher expression of ALB. In contrast, cells differentiating under O2 (−)-5% conditions showed significantly lower expression of ALB. Similar to the ALB results, cells differentiating at O2 (+)-5% showed significantly higher expression of CYP3A4 and CYP1A2. For CYP3A4, cells differentiating at O2 (+)-10% also showed a significant increase in gene expression. However, cells differentiating at O2 (+)-20% were not significantly different from those differentiating at O2 (−)-20%. These results suggest that a sufficient oxygen supply with a physiological oxygen concentration induces hepatic maturation. However, these cells still expressed high levels of AFP, which indicates an immature hepatic phenotype (Fig. 6D), and the CYP3A4 activity was quite low in both O2 (+) and O2 (−) culture (data not shown).
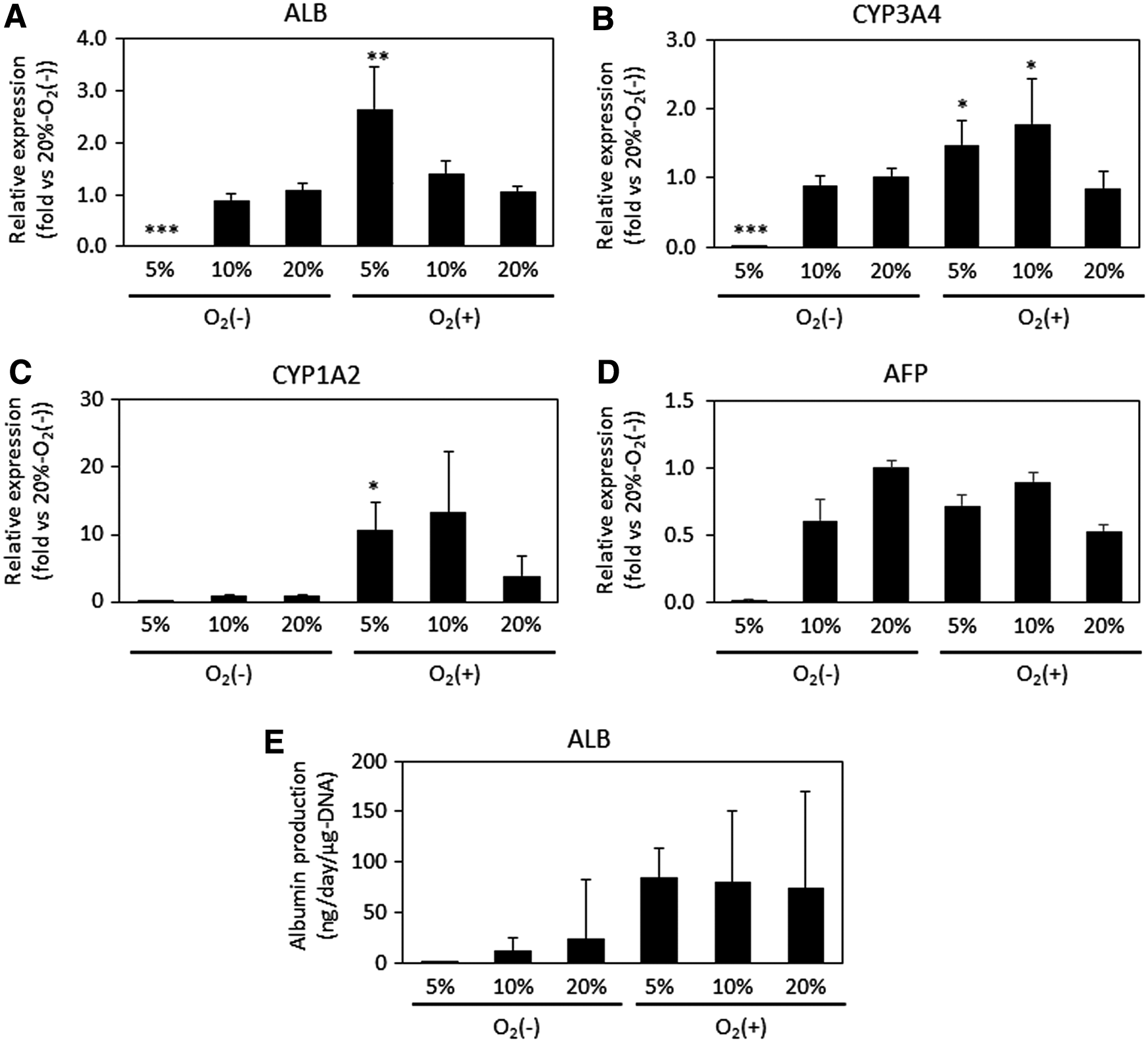
Relative gene expression of ALB
We also measured the cellular ALB secretion ability to confirm hepatic maturation (Fig. 6E). As a result, cells differentiating in O2 (+) culture tended to exhibit higher ALB production than those differentiating at O2 (−)-20%. In contrast, cells differentiating at O2 (−)-5% showed much lower ALB secretion. Thus, improving oxygen supply to the cell layer was effective in achieving hepatic functional activity.
Multilayered structure obtained with direct oxygenation
To evaluate cell proliferation during hepatic differentiation, we measured the DNA content on day 20 (Fig. 7A). Compared to the DNA content of the O2 (−)-20% condition, the O2 (+)-5%, O2 (+)-10%, and O2 (+)-20% cultures exhibited 2.7, 2.2, and 2.2 times the amount of DNA, respectively. In other words, we achieved cell numbers two to three times higher than the same culture surface area. Thus, these results indicated that O2 (+) culture was effective for not only hepatic maturation but also efficient generation of iHeps in per unit surface area. To further investigate this high-density culture, we prepared vertical cross-sections stained with hematoxylin and eosin (H&E) (Fig. 7B). Photographs of the H&E staining of vertical sections revealed that cells cultured under O2 (+) conditions showed a self-organized, thick, multilayered configuration. In contrast, cells cultured under O2 (−) conditions formed thin monolayers. Moreover, among the O2 (+) culture group, the low atmospheric oxygen condition (5% O2) showed the highest DNA content and the thickest multilayered formation.
Discussion
To use iHeps for engineering in vitro liver models, further maturation of iHeps is essential. The improvement of the culture environment is one of the keys to the hepatic differentiation of hiPSCs. While many approaches have focused on the composition of the culture medium10–13 or co-culture with nonparenchymal cells,1,15 few reports have focused on oxygen conditions of the culture system during hepatic differentiation, which is an important factor for human development 19 and hepatocyte culture. 30 Although Si-Tayeb et al. applied different oxygen pressures during differentiation, the advantages of the different oxygen concentrations were not discussed.10,20 In this study, we used PDMS membranes as the bottom of the culture plates to supply oxygen directly to the cell layer, and we evaluated the effect of oxygen supply on the hepatic differentiation of hiPSCs. When compared to cells differentiating in O2 (−) culture, we observed higher expression of GATA4 in endoderm cells differentiating in O2 (+) culture, as well as significant improvements in the expression of several mature hepatic marker genes (ALB, CYP1A2, and CYP3A4) and ALB production in iHeps generated in O2 (+) culture with physiological oxygen concentration (5% or 10% O2). Moreover, oxygen flux measurements revealed that iHeps generated in O2 (+) culture exhibited a higher consumption of oxygen. Under such sufficient oxygen supply conditions, cells spontaneously formed multilayered structures, and thus, the efficient generation of iHeps was achieved.
Oxygen plays an essential role in embryo development and liver organogenesis. 20 In vivo, oxygen is supplied by blood flow. During liver organogenesis, early liver development occurs under low oxygen supply conditions because the blood vessels are not yet developed. 19 In contrast, as vascular formation occurs along with liver bud formation, 30 in the latter stage of liver organogenesis, 31 the amount of oxygen supplied to the cell markedly increases and cells utilize more oxygen. In our study, oxygen supply to the cell layer was improved by using an oxygen-permeable membrane as the bottom of culture plate. Oxygen supply to the cell layer is therefore determined by the concentration gradient between the atmospheric and pericellular oxygen concentrations and can be expressed using Fick's Law. 24 While in the conventional TCPS plate, oxygen supply to the cell layer depends only on the diffusion of oxygen through the layer of culture medium, in the PDMS plate, it is supplied through both the culture medium and the PDMS membrane. Based on the results of the oxygen concentration measurements, we observed similar oxygen concentrations at the cell layers in the O2 (−)-20% (10.77%) and O2 (+)-10% (9.08%) cultures and in the O2 (−)-10% (4.81%) and O2 (+)-5% (4.24%) cultures. However, the supplied oxygen flux to the cell in O2 (+) was about two to four times higher than that in O2 (−) culture. A comparison of the permeability coefficient of oxygen through both the culture medium and PDMS membrane revealed that PDMS has a permeability coefficient that is about seven times higher (PPDMS: 1.72 × 10−1 pmol·cm/s·cm2·mmHg; Pmedium (Dmedium × Kmedium): 2.38 × 10−2 pmol·cm/s·cm2·mmHg). Thus, PDMS contributes considerably to improving the oxygen supply and cellular oxygen consumption. Moreover, considering that the oxygen concentration in the liver roble is between 4% and 11%, 32 this implies that the improved maturation observed in O2 (+) culture with a 5% or 10% oxygen concentration occurred through the replication of both an in vivo-like oxygen concentration and an in vivo-like oxygen supply.
In most hepatic differentiation protocol, many expensive cytokines were added to induce differentiation. Thus, the cost-effective differentiation methods would be attractive. 33 In sufficient oxygen-supplied condition using PDMS membrane, we revealed cells spontaneously formed multilayered configuration. Especially, in O2 (+)-5% or O2 (+)-10% condition, we achieved about 2.5 times higher DNA content than that in O2 (−)-20% condition. Moreover, we observed about three times higher ratio of ALB-positive cells in these conditions than that in O2 (−)-20% condition. Based on these results, we achieved about 7.5 times higher per unit surface area productivity of iHeps. Thus, O2 (+) culture under physiological oxygen concentration was effective for efficient generation of iHeps. Furthermore, it is suggested the formation of multilayered configuration is also effective on hepatic maturation. Many researchers show the importance of 3D culture compared to 2D culture16,17 for hepatic differentiation of hiPSCs, especially, the regeneration of cell–cell junction is critical. Although cells formed a thin monolayer in the O2 (−) condition, cells spontaneously formed 3D configuration in O2 (+) culture. Thus, in addition to the replication of in vivo-like oxygen concentration and oxygen supply, replication of in vivo-like cell–cell interaction by direct oxygenation is also effective for hepatic differentiation of hiPSCs.
Our findings also revealed that direct oxygenation through PDMS is effective for endodermal differentiation, even though vascularization has not yet occurred during early liver organogenesis. Although one advantage of direct oxygenation through the PDMS membrane is enabling cells to utilize sufficient oxygen for respiration, in this study, we found that endodermal cells differentiating under both O2 (−) and O2 (+) culture conditions performed anaerobic respiration (Fig. 3). However, we also found that HIF-1α, which is highly expressed under hypoxic conditions, was downregulated by PDMS culture. Several reports have revealed that Wnt signaling plays an important role during early liver organogenesis,34–37 and thus, some hepatic differentiation protocols add Wnt3A cytokines to the culture medium to promote endoderm differentiation.12,38 The relationship between Wnt signaling and HIF-1α expression is widely known, and Wnt signaling is activated by the inhibition of HIF-1α gene expression. 39 This suggests that the inhibition of HIF-1α using the PDMS membrane contributes to the effective differentiation of hiPSCs toward endodermal cells.
Although iHeps differentiating under O2 (+) culture conditions with physiological oxygen concentrations showed significantly higher expression of several mature hepatic markers and increased the production of ALB compared to cells differentiating under O2 (−) conditions, they still exhibited some fetal hepatocyte phenotypes, such as the high expression of AFP, whose expression is downregulated in PHHs, or the quite low metabolic activity of CYP3A4. Several promising approaches have been reported for the further maturation of iHeps, including co-culture with nonparenchymal cells 15 and 3D culture,16–18 in addition to the optimization of soluble factors added to the culture medium during differentiation protocols. However, these approaches have not yet resulted in fully mature iHeps. In these studies, oxygen supply to the cell layer has not been considered. As a synergistic approach of co-cultures and oxygen supply improvements, Xiao et al. reported that co-culture of rat hepatocytes with nonparenchymal cells under PDMS membrane-based direct oxygenation enabled in vivo-like hierarchical double-layered microstructure on the PDMS membranes and maintained higher levels of hepatic function for prolonged time. 25 In the near future, through integration of various advanced culture methods such as 3D culture or co-culture with the improved oxygenation as evidenced in this study, better physiologically relevant in vitro liver tissue models with prolonged duration can be developed for efficacy/toxicity screenings of drugs or liver disease models. Therefore, co-cultures with various nonparenchymal cells with sufficient oxygen supply through PDMS membranes are expected to enhance further maturation of iHeps in vitro.
In summary, we have shown sufficient oxygen supply to the cell layer through the use of a PDMS membrane to be effective in the hepatic maturation of iHeps. We demonstrated the effect of direct oxygenation through the PDMS membrane by performing hepatic differentiation of hiPSCs under several different oxygen concentrations with varying methods of supplying oxygen. The combination of the PDMS membrane and a physiological oxygen concentration contributes to an improvement in cellular oxygen consumption. Hepatic differentiation under a sufficient oxygen supply can promote the further maturation of iHeps. Moreover, direct oxygenation through the PDMS membrane also promotes the self-organization of a 3D configuration during hepatic differentiation, leading to an increase in per area-based productivity. Thus, direct oxygenation through a PDMS membrane, rather than a conventional TCPS plate, is a promising fundamental culture method for the maturation of iHeps.
Footnotes
Acknowledgments
This work was supported by the Japan Agency for Medical Research and Development (AMED, “Development of Cell Production and Processing Systems for Commercialization of Regenerative Medicine”). We thank Vecell, Inc. for supplying the permeability coefficient of the PDMS membrane. K.K. is grateful for the financial support from SEUT (Graduate School of Engineering, The University of Tokyo, Doctoral Student Special Incentives Program).
Disclosure Statement
No competing financial interests exist.