
Abstract
This study developed a xeno-free (XF) spherical cell cluster of human adipose-derived stem cells (hASCs) for the treatment of severe ischemic diseases. hASCs are an attractive cell source for therapeutic angiogenesis. Herein, we describe an innovative method of culturing a three-dimensional cell mass (3DCM) in the XF condition and demonstrate the therapeutic potential and safety of XF-3DCMs in ischemic mice and in SCID/BALB/c Slc-nu/nu nude mice, respectively. XF-3DCMs were generated by culturing hASCs on a maltose-binding protein-linked basic fibroblast growth factor (bFGF)-immobilized polystyrene surface. XF-3DCMs produced comparable amounts of angiogenic factors such as bFGF, vascular endothelial growth factor, and interleukin-8 compared to 3DCMs formed in media containing FBS (fetal bovine serum)-3DCMs. The majority of the mice injected with XF-3DCMs exhibited limb salvaging and displayed similar blood perfusion compared to normal limbs. To a similar degree, as for FBS-3DCMs, XF-3DCMs enhanced therapeutic efficacy by increasing blood vessel regeneration, attenuating muscle degeneration and fibrosis, and facilitating the implantation of injected cells into host vessels. Furthermore, the soft agar colony-forming assay and in vivo tumorigenicity analysis showed that XF-3DCMs did not transform into tumor cells. No side effects were observed, including hematological and immunological toxicity, in mice injected with XF-3DCMs. These results suggest that XF-3DCMs show angiogenic and therapeutic efficacy in critical limb ischemia in mice and eliminate the safety problem of animal-derived material contamination, which is a prerequisite for clinical applications.
Impact Statement
We describe an innovative method of culturing a three-dimensional cell mass (3DCM) of human adipose-derived stem cells (hASCs) in the xeno-free (XF) condition for the treatment of severe ischemic diseases. The majority of the mice injected with XF-3DCMs exhibited limb salvaging and displayed similar blood perfusion compared to normal limbs. In vivo tumorigenicity and toxicity analysis showed that XF-3DCMs did not transform into tumor cells and induce toxicity, respectively. Our results strongly suggest that XF-3DCMs can be effectively used for therapeutic applications and eliminate immunological reaction of animal-derived material contamination.
Introduction
C
Recent stem cell therapy has aimed to supply blood into damaged tissue with promising results in preventing limb amputation. Human mesenchymal stem cells (hMSCs) have been widely recognized as a tool for therapeutic applications for decades.8,9 In particular, human adipose-derived stem cells (hASCs) are an attractive and readily available cell source for regenerative medicine, owing to their ability to differentiate into multiple cell lineages and their immunoprivileged properties.1,10 It has been reported that transplanted hASCs induced angiogenesis in animal models by paracrine secretion of angiogenic, antiapoptotic, and anti-inflammatory factors.11,12 Thus, hASCs have been proposed as potential candidates for cell therapy of vascular disease and other degenerative diseases, and are currently being evaluated in multiple clinical trials.13,14 To date, eight clinical trials have been completed using hASCs, including diseases such as androgenic alopecia, Crohn's fistula, CLI, Romberg's disease, and sepsis. Among these, three trials used patient-specific or autologous hASCs (http://clinicaltrials.gov). Autologous hASCs are under examination in clinical case studies for treating breast cancer, chronic liver cirrhosis, chronic kidney disease, and so on (www.geron.com). In contrast, clinical trials using induced pluripotent stem cells and human embryonic stem cells are in their infancy (www.geron.com) and are limited by ethical considerations. 15
Multiple clinical trials are underway to evaluate the use of hASCs in various fields of regenerative medicine; limitations remain regarding clinical applications, predominantly because of their lower cell viability and safety. Several experimental strategies for improving the survival and therapeutic potential of stem cells in ischemic tissue have been developed, including transplantation in combination with growth factor delivery, genetic modification of stem cells, growing stem cells under hypoxic conditions, and growing cells in two-dimensional (2D) or three-dimensional (3D) spheroid cultures.16–19 In particular, 3D cell spheroid cultures improved cell survival, promoted cell stabilization, and prevented cell apoptosis after engraftment in ischemic tissue.19,20 Grafting cellular spheroids showed improved therapeutic efficacy together with enhanced cell survival and paracrine effects in ischemic models.21,22 Since cells within the 3D microenvironment are naturally exposed to mild hypoxia, they are naturally preconditioned to an ischemic environment. Therefore, spheroid grafting in ischemic tissues may render cells resistant to hypoxia compared to cells grown in monolayer cultures. Generally, cells form an in vitro 3D spherical aggregate by controlling the strength balance between cell–cell contact and cell–matrix adhesion. 23 We previously reported the mechanism and formation of 3D cell mass (3DCM) composed of hASCs on a recombinant maltose-binding protein-basic fibroblast growth factor (MBP-FGF2)-immobilized surface. 24 The 3DCM has been demonstrated to produce various angiogenic factors and enhance therapeutic potential in various degenerative diseases.20,22,24
To provide a sufficient number of cells in a timely manner, hASCs are often expanded in a 2D state using classical media, for example, Dulbecco's modified Eagle's medium/nutrient mixture F-12 in a 1:1 mixture (DMEM/F-12) and minimal essential medium, commonly supplemented with 10–20% fetal bovine serum (FBS).1,10 However, it is not recommended for clinical cell therapies because FBS may carry adventitious agents, risking viral infection or mycoplasma contamination, and other substances that can cause severe immunogenic reactions when transfused into a patient.25,26 Furthermore, human cells exposed to an animal-derived (xenogeneic) product may transfer xenogeneic proteins that are internalized or attached onto cell surfaces, with the possibility of infectious agent transmission.27,28 Thus, FBS-containing media is an unsuitable option in view of patient safety. The replacement of FBS with a xeno-free (XF) formulation has been reported to support equal or higher proliferation rates and multipotentiality in the culture of hASCs relative to using FBS.29–31
In this study, we develop XF-3DCMs on an MBP-FGF2-immobilized polystyrene (PS) substrate to evaluate their therapeutic benefits and safety in vitro and in vivo. We examined the transplantation of XF-3DCMs with regard to enhanced cell retention, tissue regeneration, angiogenesis, and limb survival in a mouse ischemic hindlimb model compared with transplantation of monolayer-cultured hASCs. Moreover, the therapeutic characteristics of XF-3DCMs are compared to those of hASC-3DCMs grown in a medium containing 10% FBS (FBS-3DCMs). Importantly, we demonstrate that XF-3DCMs do not induce tumorigenicity or any observable toxicological effects in the mouse model.
Materials and Methods
Preparation of a PS surface coated with MBP-FGF2
To prepare the MBP-FGF2-coated PS surface (PS-MBP-FGF2), the recombinant protein MBP-FGF2 was created using an MBP as a physical linker for the immobilization of FGF2 onto the hydrophobic PS surface as previously reported.20,22,24 In brief, 100 μL of MBP-FGF2 (10 μg/mL) was dissolved in phosphate-buffered saline (PBS; Sigma-Aldrich) and added to each well of the PS plates (nontissue culture-treated plates, 96-well, Falcon; Corning, Inc.). After 4 h at room temperature, nonadsorbed protein solutions were removed from the plates by washing thrice with PBS.
hASC culture and construction of XF-3DCMs
hASCs were purchased (CEFO Bio, South Korea) and subcultured in a basal medium of human-adipose-tissue-derived MSC growth medium (CEFOgro™ ADMSC [CB-ADMSC-GM]; CEFO Bio) with the provided supplementation. For each passage, the cells were plated at a density of 5 × 10 3 cells/cm 2 and grew by 70% confluence; then, a subculture was conducted using 0.25% trypsin-EDTA (Corning®). Fifth-passage (P5) cells were used for all the experiments after extensive washing with PBS. The morphology of adherent cells was observed using a phase-contrast microscope (Zeiss Axio Vet. A1, Germany). The scheme for forming XF-3DCMs of hASCs is presented in Figure 1. hASCs were suspended in an XF-hMSC culture medium (STEMPRO® MSC SFM; Life Technologies) or DMEM/F-12 (Welgene, South Korea) supplemented by 10% FBS (Welgene) and 100 U/mL penicillin and streptomycin (Invitrogen), seeded at a density of 1 × 10 5 cells/cm 2 in each well of the 96-well PS-MBP-FGF2 plate, and incubated for 24 h at 37°C and 5% CO2. 3DCM formation was observed using a phase-contrast microscope. The morphology of the 3DCMs was evaluated by phase-contrast microscopy and scanning electron microscopy (SEM, Inspect F50). hASCs suspended in XF media and cultured on a tissue culture plate (TCP, XF-hASCs) for 24 h served as the monolayer culture.
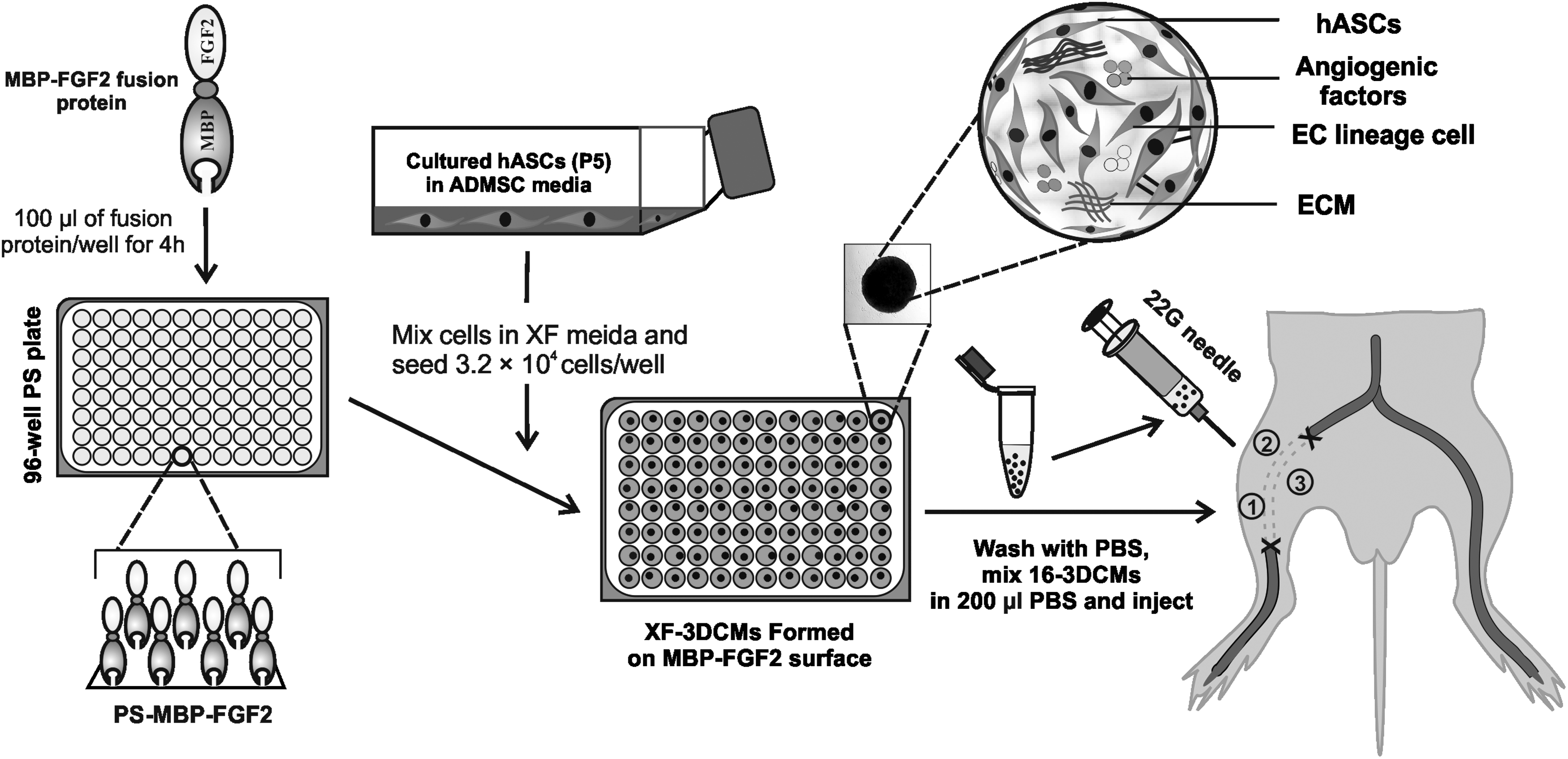
Schematic illustration of XF-3DCM generation for therapeutic angiogenesis. 3DCMs, three-dimensional cell mass; XF, xeno-free.
Cell viability assay
To assess cell survival, we experimented with a Live/Dead cell viability assay kit (BioVision) according to the manufacturer's instructions. Harvested 3DCMs were treated with Solution A (Live-Dye, green) and Solution B (PI, red) and incubated for 30 min in a CO2 incubator. The 3DCMs were washed thrice with PBS, fixed with 4% PFA for 30 min, embedded in an optimized cutting temperature compound (TISSUE-TEK® 4583), frozen, and cut into 10-μm-thick sections at −28°C. The sections were examined using confocal microscopy (Carl Zeiss, Germany). Freeze-thawed 3DCMs were used as control for dead cells.
Enzyme-linked immunosorbent assay
Vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF), and interleukin-8 (IL-8) production in the XF-3DCMs were determined using an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems) according to the manufacturer's protocols. Concentrations were expressed as the amount of human VEGF165, bFGF, and IL-8 per 10 5 cells at a given time.
The presence of residual bovine serum albumin (BSA) in hASC suspensions was calculated by a commercially available ELISA kit (Cygnus Technologies) according to the manufacturer's instructions. For ELISA sample preparation, hASCs suspended in XF media (300 μL containing 5 × 10 5 cells) were collected into a microcentrifuge tube and the medium (XF supernatant) was collected after centrifugation (at 2451 g for 5 min). The cell pellet was washed with PBS (300 μL) thrice; the PBS was collected separately after each wash for BSA analysis.
Mouse hindlimb ischemia model
Six-week-old male athymic mice (20–25 g body weight; Orient, South Korea) were anesthetized with xylazine (10 μg/kg) and ketamine (100 μg/kg). The femoral artery and its branches were ligated using a 5-0 silk suture (Ethicon). The femoral artery was excised from the proximal branch of the external iliac artery to the distal point where it bifurcates into the saphenous and popliteal arteries. All animal treatments and experimental procedures were approved by the Institutional Animal Care and Use Committee of Korea Institute of Science and Technology [2016-02015].
Treatment of limb ischemia
One day after ischemia induction surgery, the mice were divided randomly into four groups (n = 12 for each group and for each time point). The groups were as follows: a control group (PBS-injected mice, PBS group), XF-hASC group, XF-3DCM group, and FBS-3DCM group. XF-hASCs, XF-3DCMs, and FBS-3DCMs (5 × 10 5 cells suspended in 0.2 mL of PBS per mouse) were injected intramuscularly into the gracilis muscle at three locations (an approximately equal amount was used for each injection, Fig. 1) in the medial thigh of the ischemic limb as previously reported. 22 The physiological status of ischemic limbs was followed up until 28 days posttreatment. Every 7 days, cutaneous blood flow was observed by a laser Doppler perfusion imager system (PeriScan PIM 3 System; Perimed AB, Stockholm, Sweden). To minimize variables, including ambient light and temperature, perfusion was expressed as a ratio of values (the ischemic to nonischemic hind limb). The outcome was rated in four levels: limb salvage (similar limb integrity and morphology as in the normal limb control of the same animal), foot necrosis (damage of the foot tissue), limb necrosis (partial damage to the thigh tissues), or hind limb loss.
Immunofluorescence staining
Indirect immunofluorescence staining was performed using a standard procedure. In brief, six paraffin sections (from each group at 14 and 28 days) were randomly selected, deparaffinized, washed twice with PBS, and blocked with 4% BSA to avoid nonspecific binding. To detect transplanted human cells, the sections were immunofluorescent stained with anti-human nuclear antigen antibody (HNA, mouse; Millipore) overnight at 4°C. To stain endothelial cells and perivascular cells, the samples were stained with anti-CD31 (rabbit; Abcam) and anti-SMA (mouse; Abcam), respectively, overnight at 4°C. The samples were then washed with PBS and incubated for 1 h at room temperature with the corresponding fluorescence-conjugated secondary antibody (goat anti-mouse alexa 594; Invitrogen, goat anti-rabbit alexa 488; Invitrogen, and goat anti-mouse Alexa 488; Invitrogen). DAPI (4,6-diamidino-2-phenylindole; Vector Laboratories) was used for nuclear staining. Two images from each slide (200 × magnification) were evaluated in the ischemic regions to quantify the density of CD31-positive, αSMA-positive, and HNA-positive signals.
Gene expression using the quantitative real-time polymerase chain reaction
The angiogenic gene expression of XF-3DCM-implanted mice hindlimb tissue was quantitatively analyzed by quantitative real-time polymerase chain reaction (qPCR) using SYBR Premix Ex Taq (TaKaRa) real-time PCR detection systems according to the manufacturer's instructions. The extracted RNA was dissolved in nuclease-free water, and the RNA concentration was quantified using a NanoDrop ND1000 spectrophotometer (Thermo Fisher Scientific). Complementary DNA synthesis was performed using Maxime RT PreMix (iNtRon, South Korea) following the manufacturer's instructions. All PCRs were performed using an ABI Prism 7500 (Applied Biosystems) and the relative gene expression was calculated by the comparative Ct method. All the primer sequences were received from Bioneer by special order. The reverse and forward primers of the target gene were as follows: mouse Pecam1: 5′-GAGCCCAATCACGTTTCAGTTT-3′ (forward) and 5′-TCCTTCCTGCTTCTTGCTAGCT-3′ (reverse); mouse αSMA: 5′-CTGACAGAGGCACCACTGAA-3′ (forward) and 5′-CATCTCCAGAGTCCAGCACA-3′ (reverse); and mouse GAPDH: 5′-CCAATGTGTCCGTCGTGGAT-3′ (forward) and 5′-TTGCTGTTGAAGTCGCAGGAG-3′ (reverse). Mouse GAPDH served as the internal control.
In vitro tumorigenicity assay
To evaluate tumorigenicity in vitro, anchorage-independent growth was assessed using soft agar colony formation assay with a CytoSelect™ 96-well transformation assay kit (Cell Biolabs) according to the manufacturer's instructions. HeLa cells (positive control) were suspended in RPMI-1640 media (Lonza) containing 10% FBS and plated at densities of 1 × 10 3 , 2.5 × 10 3 , 5 × 10 3 , and 1 × 10 4 cells per well. Test samples, XF-hASCs, and XF-3DCMs were seeded at the same density as for the HeLa cells. The negative controls were the culture media of each cell (HeLa cells, XF-hASCs, and XF-3DCMs). Plates were incubated at 37°C and 5% CO2 for 7 days, after which colony formation was observed under a microscope (Nikon, Japan). For quantitative analysis, the colonies were lysed and quantified with CyQuant using a fluorometer equipped with a 485/520 nm filter set (SPECTRAmax M3; Molecular Devices).
In vivo tumorigenicity and immunotoxicity analysis
Animals
Five-week-old female severe combined immunodeficiency (SCID) mice and 19-week-old male/female nude (BALB/c Slc-nu/nu) mice were purchased from Charles River Laboratories Japan, Inc. The mice were acclimatized for 4 days, and the healthy animals were assigned randomly to treatment groups using a toxicology study management software (PATH/TOX SYSTEM; Xybion Medical System Co.). Three animals were housed per polycarbonate filter-top cage throughout the study period. A pellet diet (PMI Nutrition International) and sterilized tap water were provided ad libitum. The animal room was maintained at 23°C ± 3°C, a humidity of 30–70%, air ventilation of 10–20 air changes per hour, and light intensity of 150–300 lux with a 12-h light/12-h dark cycle. All animal experiments were reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of the Korea Institute of Toxicology (KIT).
Experimental design
For the tumorigenicity study, healthy SCID mice were randomly divided into four groups (n = 6 for each group). Vehicle control (StemPro® MSC SFM basal medium for mesenchymal stem cells), positive control (HeLa cells, 1 × 10 6 ; a clinical dose of administration), and test groups, XF-3DCMs (1 × 10 6 , a low dose considered a clinical dose of administration or equal to a positive control clinical dose) and XF-3DCMs (5 × 10 6 , a high dose; five times higher than the low dose or five times higher than the positive control), were simultaneously injected subcutaneously into the lateral trunk region. After injection, mice were observed once a day for clinical signs and mortality, and twice weekly for the presence of tumors throughout the experiment period. Body weight was measured at the initiation of treatment and then once a week. Tumor growth was estimated by external measurement of the length and width of the tumors in two dimensions using a caliper as soon as tumors reached measurable size. The tumor volume was calculated using the formula volume = 1/2 × length (mm) × (width [mm]). 2 All animals, including those in control groups, were killed 13 weeks after injection and the following were harvested: the injection site area and organs (abnormal lesions, liver, mesenteric lymph node, and spleen). These tissues were embedded in paraffin blocks and were subjected to hematoxylin and eosin (H&E) and immunohistochemistry (IHC) staining.
A 4-week toxicity study was performed using BALB/c nude mice. The mice were divided into 3 groups of 12 (each group consisted of 6 males and 6 females). Vehicle control, XF-3DCMs (1 × 10 6 , a low dose considered a clinical dose of administration), and XF-3DCMs (5 × 10 6 , a high dose; five times higher than the low dose) were injected intramuscularly. After XF-3DCM injection, clinical signs and mortality were observed once per day. Body weights and food consumption were determined once per week. Hematology, serum biochemistry, and immunotoxicity (lymphocyte) evaluation were conducted during the last week of the experiment period. Gross findings, organ weights, and histopathological analysis were performed last; this included collecting the brain, liver (with gall bladder), spleen, heart, kidneys, adrenal glands, testes, epididymides, lung, thyroids (with parathyroids), uterus (with cervix), and ovaries. Weight and histopathological changes were observed macroscopically and microscopically for all organs from each group.
Hematology, serum biochemistry, and immunotoxicity
For hematology analysis, blood samples were collected in EDTA-2K tubes from the posterior vena cava. The following parameters were then measured by a hematological autoanalyzer (ADVIA120 Hematology System; Bayer): white blood cell (WBC), red blood cell (RBC), and hemoglobin concentrations, hematocrit, mean corpuscular volume, mean corpuscular hemoglobin (MCH), MCH concentration (MCHC), platelets, neutrophils, reticulocyte count, and WBC differential count.
For serum biochemistry analyses, blood samples were collected without an anticoagulant. The sample tubes were placed at room temperature over 90 min, and then serum samples were obtained by centrifugation at 2012 g for 10 min. Albumin, aspartate aminotransferase, alanine aminotransferase, alkaline phosphatase, albumin/globulin ratio, blood urea nitrogen, creatinine, creatine kinase, glucose, total cholesterol, total protein, total bilirubin, triglycerides, phospholipids, and gamma-glutamyl transferase (GGT) were analyzed using a clinical chemistry autoanalyzer (200FR NEO; Toshiba Co., Japan).
For immunotoxicity evaluation, the extracted spleens from each group were placed in a sterile Petri dish containing 10 mL of Earle's balanced salt solution (Sigma) and spleen cell suspensions were obtained by pressing each spleen with a syringe plunger. The cell suspension was treated with a lysis buffer (Lysing buffer; BD Biosciences), placed for 15 min, and then centrifuged at 234 g for 10 min, to remove RBCs. The cells were resuspended with staining buffer (BD Biosciences) and adjusted to 3 × 10 6 cells/mL. One hundred microliters of the adjusted suspension was reacted with 0.3 μg per sample of mouse BD Fc block (BD Biosciences) for 5 min at 4°C. Specific antibodies of cell membrane markers (Supplementary Table S1) were added to the suspension and the mixture was incubated for 30 min at 4°C and analyzed within 24 h using a flow cytometer (FACS Calibur; BD Biosciences). The percent content of the lymphocyte subset was calculated using Cell Quest Pro (version 5.2.1; BD Biosciences) or FlowJo (version 10.1; FlowJo, LLC). The phenotypes of the lymphocyte subset were total T cells (T), helper T cells (Th), cytotoxic T cells (Tc), B cells, and natural killer (NK) cells. The data were represented as relative values for the lymphocyte phenotypes rather than absolute values, as well as normalized values using spleen weights (corrected quantity). The corrected quantity was calculated from the formula (spleen weight × relative percentage/100). It was used to reduce variation due to difference in spleen weights since lymphocyte phenotypes in lymphoid organs might be affected with its weights.
Histology and IHC analysis
The injection site tissues and organs of animals from each group (SCID, BALB/c Slc-nu/nu) were fixed with 10% neutral buffered formalin. The paraffin-embedded tissues were sliced into 5-μm sections, mounted onto slides, and investigated by H&E staining and IHC staining. For IHC analysis, heat-induced epitope retrieval was applied using a citrate-based buffer (pH 6.0). Sections were incubated with an anti-human mitochondrial mouse monoclonal antibody (Abcam) for 2 h. For detection, diaminobenzidine chromogen was used and counterstained with hematoxylin. The slides were examined under a phase-contrast microscope.
Similarly, ischemic limb muscles harvested at 14 and 28 days posttreatment from each group were fixed in 4% (w/v) formaldehyde and embedded in paraffin. All samples were completely sectioned; six slides were randomly selected from each sample and stained with H&E to examine muscle degeneration and tissue inflammation. Masson's trichrome (MT) staining was performed to assess tissue fibrosis in the ischemic regions. The fibrotic area (% of total area) was measured using the ImageJ software analysis system.
Statistical analysis
All the data were expressed as the mean ± standard deviation. Statistically significant difference was obtained from one-way analysis of variance (ANOVA) with a Tukey test using Prism software (GraphPad). Statistical significance was determined as *p < 0.05, **p < 0.01, and ***p < 0.001. For blood perfusion ratio analysis, one-way ANOVA with Tukey test was performed to compare the differences at each time point between the groups. Differences with p < 0.0001 were considered statistically significant. For safety data assessment, the variance homogeneity was examined by using Bartlett's test or the F-test. When the results showed nonsignificant deviation from variance homogeneity, the one-way ANOVA test was performed at α = 0.05. When there was significant deviation, Dunnett's t-test was performed. In the case of nonparametric comparison, the Kruskal–Wallis test or Dunn's rank sum test was conducted. Statistical analyses were examined using the Path/Tox System (Version 4.2.2; Xybion Co.). A value of p < 0.05 was considered to be significant.
Results
XF-hASC-3DCM formation and characterization
The morphological characteristics of XF-3DCMs were compared with those of FBS-3DCMs and XF-hASCs cultured on TCP. A monolayer of adherent hASCs cultured in XF media was observed within 4 h after seeding on PS-MBP-FGF2 by phase-contrast microscopy. The hASCs were observed to form cellular aggregates at 4 h after seeding and developed into a spherical form (XF-3DCM) within 24 h (Supplementary Fig. S1). XF-3DCMs were uniform in size and a nearly spherical shape was observed by phase-contrast microscopy and SEM (Fig. 2A). The morphologies of XF-3DCMs were similar to those of FBS-3DCMs (Fig. 2A), although XF-3DCMs were formed slightly earlier than FBS-3DCMs (Supplementary Fig. S1). XF-hASCs remained as a monolayer throughout the culture time (Supplementary Fig. S2A).
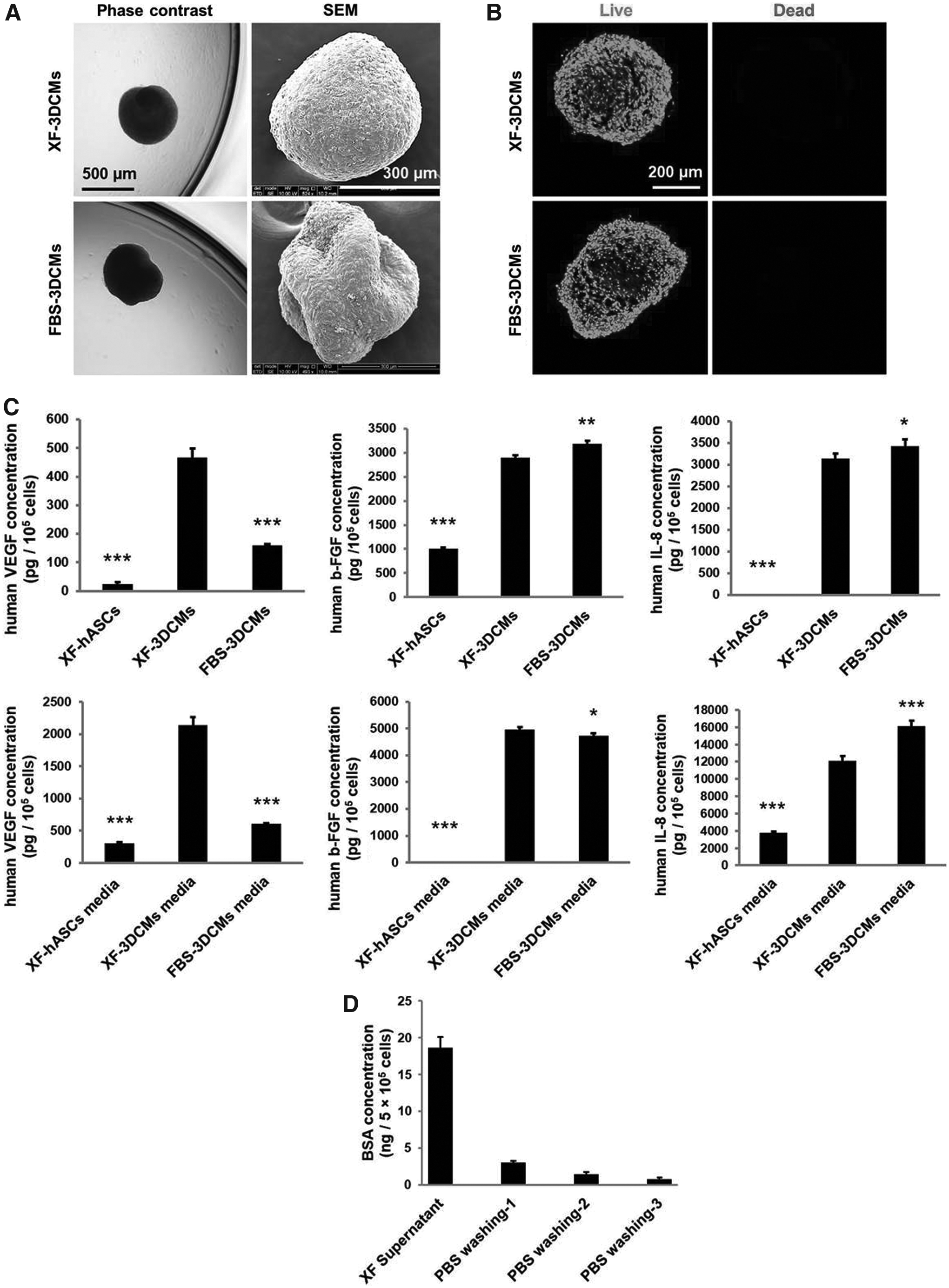
Characterization of XF-3DCMs of hASCs on a PS-MBP-FGF2 surface.
The cell viability of XF-3DCMs was qualitatively compared with that of FBS-3DCMs and XF-hASCs by live/dead assay. A microscopic image analysis revealed that no marked amount of cell death was observed in XF-3DCMs and FBS-3DCMs for 24 h (Fig. 2B). There were few dead cells observed in the XF-hASC culture (Supplementary Fig. 2B).
In our previous study, we found a correlation between increased expression of VEGF and upregulation of HIF1-α in FBS-3DCMs. 22 We speculated that similar characteristics may also exist in XF-3DCMs. Hence, we next analyzed the angiogenic factors in XF-3DCMs.
Angiogenic factor expression of XF-3DCMs
In our previous study, FBS-3DCMs produced angiogenic factors such as VEGF, bFGF, and IL-8.20,22,32 To investigate whether angiogenic factors produced by XF-3DCMs are comparable to those produced by FBS-3DCMs, the amount of angiogenic factors produced in XF-3DCMs and FBS-3DCMs was determined by ELISA. XF-3DCMs produced bFGF and IL-8 equivalent to FBS-3DCMs (Fig. 2C). Interestingly, XF-3DCMs showed threefold higher VEGF expression than FBS-3DCMs. The expression of the angiogenic factors in both XF-3DCMs and FBS-3DCMs was much greater compared with XF-hASCs (Fig. 2C). bFGF, IL-8, and VEGF secretion were found to be greater in the culture supernatant of XF-3DCMs and FBS-3DCMs than in their respective 3DCMs developed on MBP-FGF2 surfaces (Fig. 2C).
BSA quantification in hASC suspension
XF formulation exhibits significantly smaller immunological risk compared to FBS-supplemented media. When cells are used as therapeutic agents for human clinical applications, it is still necessary to check for the existence of any small traces of FBS before transplantation. BSA value in XF-3DCMs was estimated from the suspension of XF-hASCs that was used for the formation of 3DCMs. Since BSA is a major component of FBS, the existence of FBS in hASCs suspended in XF media for XF-3DCM formation was estimated by measuring the amount of BSA by ELISA. Residual BSA in the supernatant of the hASC XF suspension was ∼18.6 ng per 5 × 10 5 cells. Residual BSA concentrations were ∼3.0, 1.4, and 0.79 ng per 5 × 10 5 cells after the first, second, and third wash of the cells with PBS, respectively (Fig. 2D). BSA values after the third wash, that is, at the time of injection, were 26 times smaller than the WHO-set value of 21 ng/therapeutic dose. This result suggests that XF-3DCMs may be suitable for stem cell therapy according to WHO guidelines.
Improvements in limb salvage and blood perfusion by transplantation of XF-3DCMs
The therapeutic potential of implanted XF-3DCMs was examined by physiological observation of the ischemic limbs every 7 days until 4 weeks after treatment. We surgically generated a mouse model of hindlimb ischemia (by femoral artery ligation), which we verified by measuring blood perfusion on day 0 using a laser Doppler perfusion imaging system (Fig. 3B). Figure 3 shows the quantitative analysis of blood flow, blood perfusion ratios of the ischemic/normal hindlimb (Fig. 3C), and the quantification of limb salvage (3D). In the PBS group, rapid limb necrosis was observed on day 7 and complete limb loss by autoamputation was observed up to 28 days (Fig. 3A). All mice in this group underwent complete limb loss (92%) or severe limb necrosis (8%) without limb salvage (0%) (Fig. 3D). In the group transplanted with XF-hASCs, limb loss decreased after treatment compared to the PBS group; 75% and 25% of the mice ultimately exhibited limb necrosis and foot necrosis, respectively, within 28 days (Fig. 3D). On the other hand, we observed an increase in blood perfusion 7 days after transplanting in the XF-3DCM group compared with the XF-hASC group and PBS group (Fig. 3B). The blood perfusion continued to increase and was maintained in the XF-3DCM group until 28 days had passed. The rate of limb loss was reduced significantly in mice receiving XF-3DCMs compared to the XF-hASC group. In addition, 92% of the mice displayed normal hind limb appearance and no autoamputation. The physiological observation of the XF-3DCM group was similar to that of the FBS-3DCM group at all ischemic tissue time points after treatment (Fig. 3D). These results suggest that XF-3DCM therapy significantly increased angiogenesis and decreased limb necrosis of the ischemic tissue.
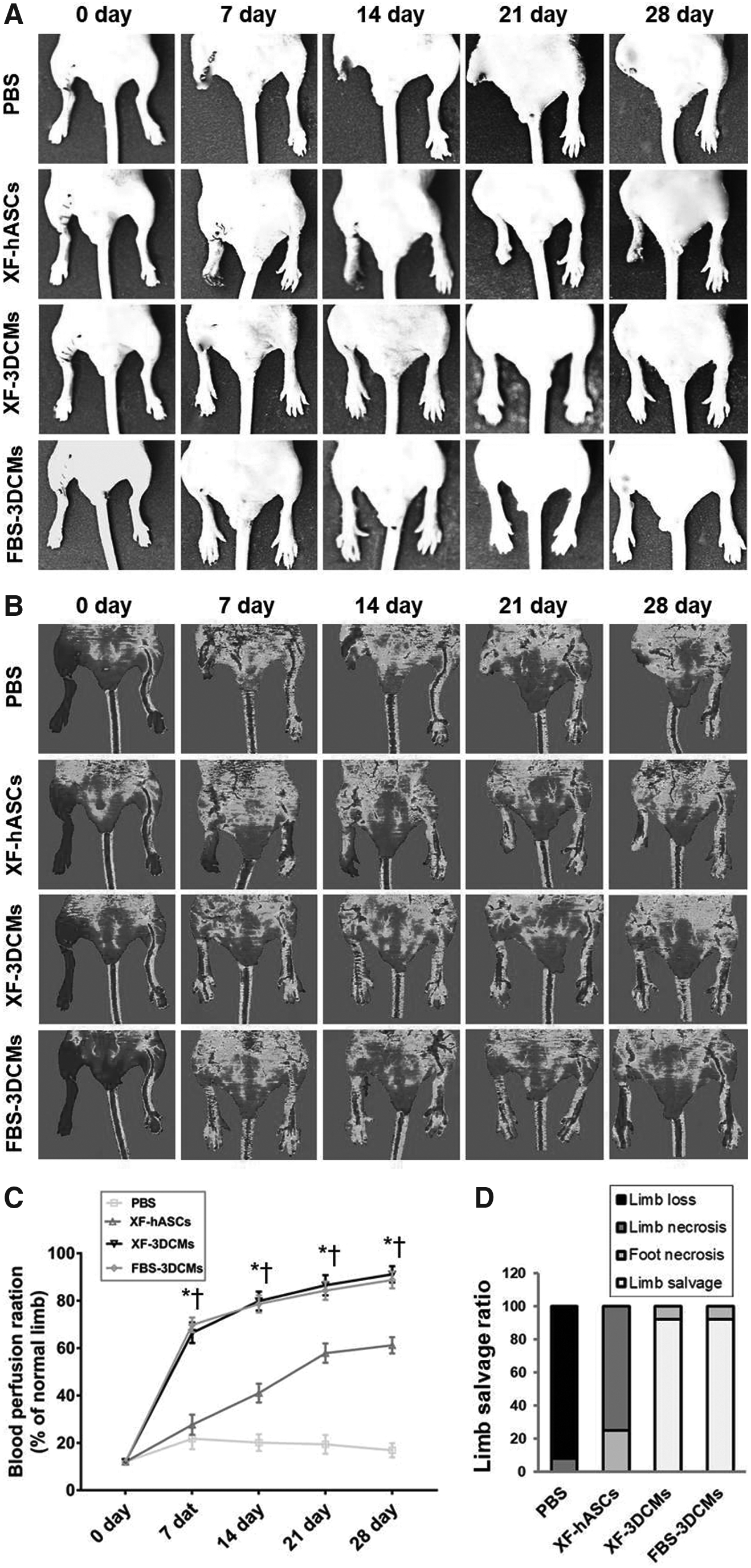
Limb morphology and blood perfusion.
Enhanced angiogenic effect of XF-3DCMs in ischemic limbs
To assess the angiogenic potential of XF-3DCM treatment, we measured the levels of CD31 and αSMA gene expression in the ischemic limbs at 14 and 28 days posttreatment using qPCR. Figure 4 shows the mRNA levels of CD31 and αSMA, which were quantified and normalized by the mRNA level in the normal (nonischemic) hindlimb. The mRNA expression of CD31 was highest in the XF-3DCM group at 14 days after treatment, showing ∼56% that of normal tissue (Fig. 4A). At 28 days after treatment, the mRNA level of CD31 in ischemic tissue transplanted with XF-3DCMs increased to ∼80% of normal tissue (Fig. 4B). As shown in Figure 4C, the mRNA expression of αSMA was also higher in the XF-3DCM group (77% of normal tissue) than in the XF-hASC group (23% of normal tissue) at 14 days after treatment. In Figure 4D, the mRNA level of αSMA in ischemic tissue transplanted with XF-3DCMs increased to ∼92% of normal tissue at 28 days after treatment. However, no significant differences were observed among the XF-3DCM group, FBS-3DCM group (86% that of the normal limb), and XF-hASC group (77% that of the normal limb) on day 28. It should be noted that immunofluorescence analysis at 28 days was not possible for the PBS group owing to autoamputation of the limbs.

Enhanced expression of angiogenic marker genes in ischemic limb tissue after XF-3DCM treatment. Real-time quantitative PCR analysis showing the expression of CD31
To verify the angiogenic potential and survival of XF-3DCM treatment, double immunofluorescence staining for CD31 and HNA was performed (n = 5 in each group) on days 14 and 28 (Fig. 5A). CD31-positive vessel-like structures (number/mm 2 ) and the distribution of HNA-positive cells were compared among groups (Fig. 5B). On day 14, the XF-3DCM group (27.7 ± 6.8 mm 2 ) and the FBS-3DCM group (26.1 ± 6.1 mm 2 ) showed higher levels of CD31-positive capillary-like structures than the XF-hASC group (15.1 ± 3.8 mm 2 ) and the PBS group (4.9.1 ± 2.21 mm 2 ), but statistically no difference was observed between the XF-3DCM group and the FBS-3DCM group (Fig. 5B-a). The number of capillary-like structures was observed to increase in all groups until day 28. However, the number of CD31-positive capillary-like structures in the XF-3DCM (39.7 ± 4.5 mm 2 ) group was nearly twice that of the XF-hASC group (20.1 ± 5.8 mm 2 ), but no statistical difference was observed compared to that of the FBS-3DCM group (38.8 ± 4.9 mm 2 ) (Fig. 5B-b). The HNA-positive cell density/field ratio was higher in the XF-3DCM and FBS-3DCM groups than in the XF-hASC group on days 14 and 28 (Fig. 5B-c and d). The HNA-positive cell density/field ratio of the XF-3DCM group (46.3 ± 9.1 mm 2 ) was nearly twice as high as that of the XF-hASC group (24.2 ± 6.6 mm 2 ), but did not show any statistical difference compared with that of the FBS-3DCM group (44.7 ± 9.3 mm 2 ) on day 14 (Fig. 5B-c). The number of transplanted cells on day 28 after transplantation decreased in all groups, but the reduction rate was lower in the XF-3DCM group (19.1 ± 4.7 mm 2 ) and the FBS-3DCM group (18.4 ± 4.3 mm 2 ) compared to the XF-hASC group (5.6 ± 1.8 mm 2 ) (Fig. 5B-d).

Survival and incorporation of the transplanted cells in ischemic limbs at 14 and 28 days after treatment.
In addition, the angiogenic effect of transplanted cells was confirmed by αSMA-positive staining (Fig. 5C) along with quantification of capillary density (Fig. 5D). The results revealed that the density of αSMA-positive microcapillaries was higher in the XF-3DCM group (9.7 ± 3.0 mm 2 ) and the FBS-3DCM group (9.4 ± 3.5 mm 2 ) at 14 days after transplantation than in the XF-hASC group (4.5 ± 1.9 mm 2 ) (Fig. 5D-a). The density of αSMA-positive microcapillaries on day 28 after transplantation increased in all groups, indicating that the density was still higher in the XF-3DCM group (16.4 ± 2.9 mm 2 ) and the FBS-3DCM group (15.9 ± 3.4 mm 2 ) than in the XF-hASC group (6.8 ± 2.7 mm 2 ). On day 28, for the ischemic area transplanted with XF-3DCMs, the density was 70–75% of normal mouse tissue (Fig. 5D-b). There was no statistical difference in capillary density between the XF-3DCM group and the FBS-3DCM group on days 14 and 28. These results are in agreement with the results of CD31-positive capillaries analysis.
Attenuation of muscle degeneration and fibrosis by XF-3DCMs
Histological analysis of ischemic limb tissues showed muscle degeneration (H&E staining, Fig. 6A) and tissue fibrosis (MT staining, Fig. 6B) induced by ischemic inflammation. Extensive muscle degeneration and severe fibrosis were observed in the limb tissue of PBS-injected mice. Ischemic tissue harvested from the XF-3DCM group depicted attenuated fibrosis and muscle degeneration at 14 and 28 days after treatment compared to the PBS group and the XF-hASC group. The fibrotic area of the ischemic tissue was quantified at 14 and 28 days from respective MT staining images (n = 5, Fig. 6C). The XF-3DCM group displayed significantly lower fibrosis (16% of total area) compared to the XF-hASC group (34% of the total area), and no significant histological or statistical differences were observed between the XF-3DCM group and the FBS-3DCM group (Fig. 6C-b). In particular, in the XF-3DCM group and the FBS-3DCM group, fibrotic area was decreased after 28 days compared to that after 14 days of treatment, whereas fibrotic area increased in the XF-hASC group after 28 days compared to that after 14 days of treatment (30% of the total area) (Fig. 6C-b). This result suggests that, although XF-hASCs do not prevent muscle degeneration, XF-3DCMs can prevent muscle degeneration.
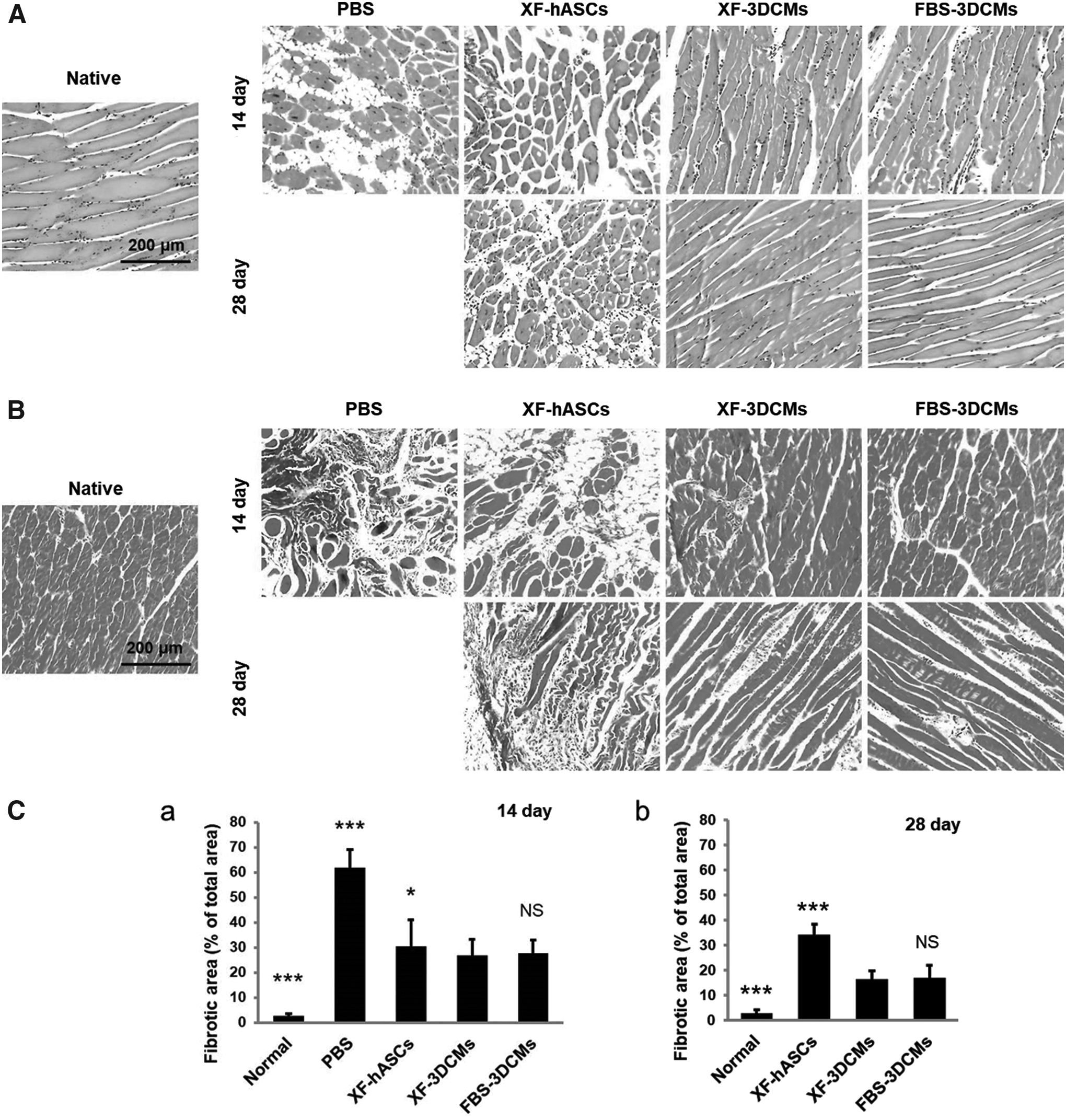
Histological analysis of ischemic hind limbs at 14 and 28 days after treatment.
In vitro tumorigenicity analysis
To compare the colony formation of XF-3DCMs with HeLa cells, we performed a soft agar colony assay using 1 × 10 3 , 2.5 × 10 3 , 5 × 10 3 , and 1 × 10 4 cells per well in a 96-well plate. HeLa cells cultured in soft agar showed progressive formation of colonies, and colony formation increased with increasing cell concentration, while XF-3DCMs and XF-hASCs did not form colonies (Fig. 7A). Furthermore, colony formation was quantitatively measured using a CyQuant assay in which the fluorescence intensity is proportional to the amount of colony formation or cell proliferation (Fig. 7A). No significant increase in the fluorescence level was observed in the XF-3DCMs and XF-hASCs, but fluorescence intensity was relatively increased for HeLa cells with respect to cell concentrations. These data suggest that both XF-3DCMs and XF-hASCs did not have tumorigenic properties.
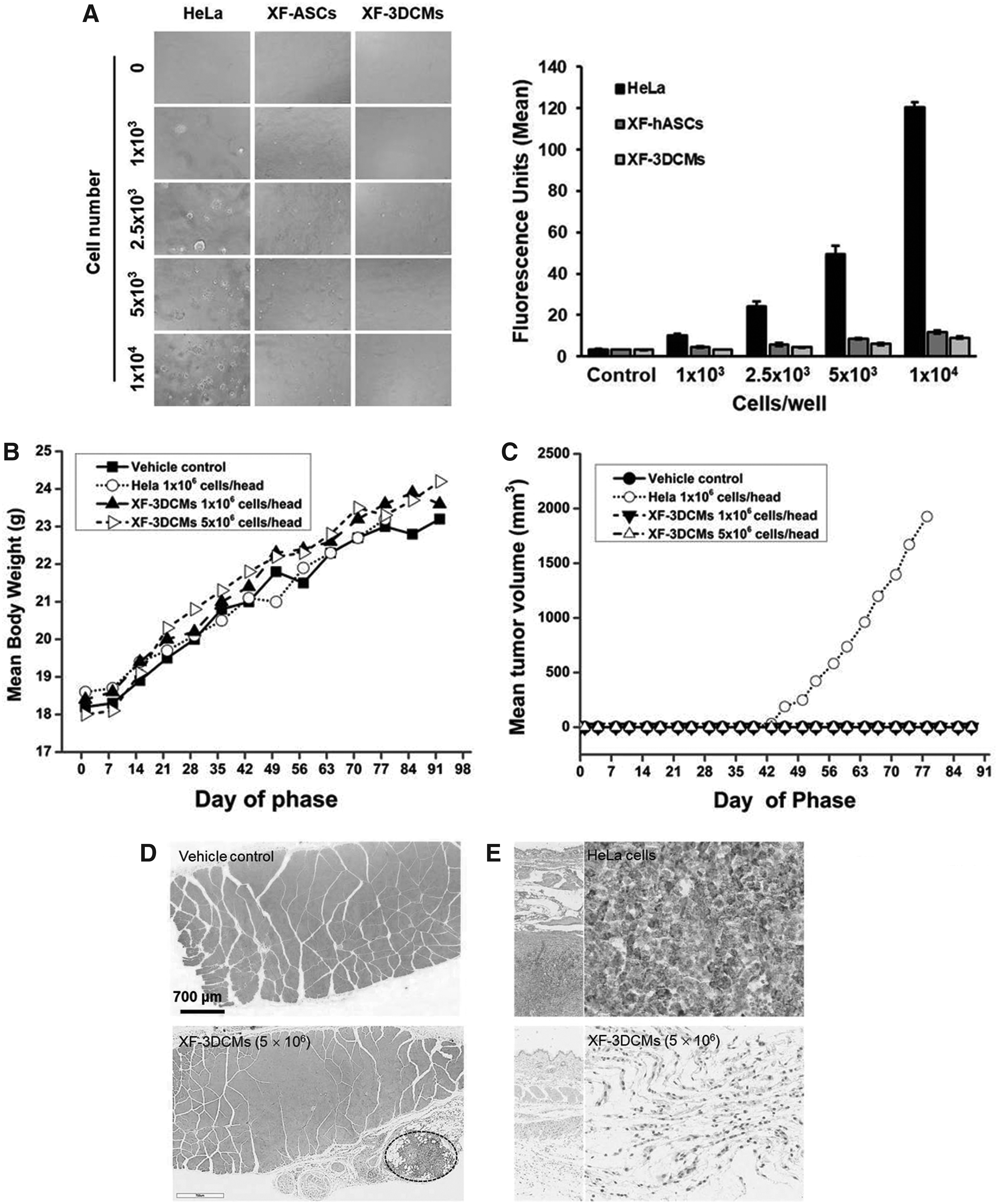
In vitro (7 day) and in vivo (13 week) analysis of tumorigenic property of XF-3DCMs.
In vivo tumorigenicity analysis
We first tried to evaluate the tumorigenic potential of XF-3DCMs (1 × 10 6 and 5 × 10 6 cells/head) by subcutaneously transplanting into SCID mice. SCID mice have been widely used for tumorigenic related findings of human cell transplantation studies. No mortality or significant body weight changes were noticed in any of the groups during the experimental period (Fig. 7B). Upon clinical observation, mice treated with a vehicle control and XF-3DCMs (low and high doses) had no clinical signs, but the HeLa cell-treated group exhibited swelling at the injection site (5/6), a palpable mass (6/6), and hard skin (6/6) with an increased tumor volume measured until necropsy (Fig. 7C). There was no treatment-related finding according to macroscopic observation in the vehicle control and XF-3DCM-treated mice during the period of observation. Microscopic examination also revealed no tumor formation in the vehicle control (Fig. 7D) or in either the XF-3DCMs with 1 × 10 6 dose (data not shown) or with 5 × 10 6 dose (Fig. 7D). The higher dose XF-3DCMs depicted only granulomatous inflammation at the site of injection (Fig. 7D, dotted elliptical curve). However, all mice injected with HeLa cells developed tumors at the injection site with four cases presenting hepatocyte hypertrophy/basophilia and extramedullary hemopoiesis in the liver and spleen, and three cases demonstrating extramedullary hemopoiesis in the mesenteric lymph node (data not shown). To confirm the origin of tumors in the mice, tissue sections were stained with anti-HNA antibodies (Fig. 7E). IHC images revealed no human mitochondrial antigen expression at the injection site in the vehicle control group (data not shown) and XF-3DCM group (Fig. 7E), while the HeLa cell-injected mice demonstrated high human mitochondrial antigen expression in tumors at the injection site (Fig. 7E).
In vivo toxicity analysis
Next, we attempted to determine the toxicity of XF-3DCMs in nude mice. Nude mice are frequently used for a variety of immunological and toxicity studies of cells expected to be used in human clinical applications. There was no treatment-related mortality or clinical signs observed for the control vehicle and XF-3DCM groups during the period of observation. No significant changes in body weight were found for XF-3DCM-treated male and female mice (Fig. 8A). Although statistically significant changes in mean daily food consumption were observed for the XF-3DCM group, these temporary changes were minimal, and they were not dose dependent. Upon hematological evaluation, the level of MCHC significantly increased and the level of EOS decreased in female mice (Table 1). However, individual variation was great and this was also not related to the microscopic observations. Hematological changes may have occurred accidentally owing to the small scale of the animals. Among serum biochemistry parameters, the level of GGT in females injected with 5 × 10 6 per animal of XF-3DCMs was significantly higher than that in the vehicle control (Table 2). However, the change was small and variation between individuals was great. This may not have any relationship with XF-3DCM treatment. The results of lymphocyte evaluation were not significantly changed in the XF-3DCM-treated male mice compared with the vehicle control (Table 3). In females treated with 5 × 10 6 per animal of XF-3DCMs, the percent values of NK cells were significantly decreased compared with the vehicle control. However, there were nonremarkable changes in the corrected quantity of immune cells.

In vivo toxicological analysis of the transplanted XF-3DCMs in BALB/c nude mice 4 weeks after treatment.
Hematological Data for Male and Female BALB/c-nu Mice Transplanted with Xeno-Free-Three-Dimensional Cell Mass in 13-Week Toxicity Study
Values are expressed as the mean ± standard deviation.
Significant difference compared to the control (p < 0.05).
Significant difference compared to the control (p < 0.01).
BAS, basophils; BASA, absolute basophils; 3DCMs, three-dimensional cell mass; EOS, eosinophils; EOSA, absolute eosinophils; HCT, hematocrit; HGB, hemoglobin; LUC, large unstained cells; LUCA, absolute large unstained cells; LYM, lymphocytes; LYMA, absolute lymphocytes; MCH, mean corpuscular hemoglobin; MCHC, MCH concentration; MCV, mean corpuscular volume; MON, monocytes; MONA, absolute monocytes; NEU, neutrophils; NEUA, absolute neutrophils; PLT, platelets; RBC, red blood cell; RET, reticulocytes; RETA, absolute reticulocytes; WBC, white blood cell; XF, xeno-free.
Serum Biochemistry for Male and Female BALB/c-nu Mice Transplanted with Xeno-Free-Three-Dimensional Cell Mass in 13-Week Toxicity Study
Values are expressed as the mean ± standard deviation.
Significant difference compared to the control (p < 0.01).
A/G, albumin/globulin; ALB, albumin; ALP, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; BUN, blood urea nitrogen; CK, creatine kinase; CREA, creatinine; GGT, gamma-glutamyl transferase; GLU, glucose; PL, phospholipids; TBIL, total bilirubin; TCHO, total cholesterol; TG, triglycerides; TP, total protein.
Lymphocyte Phenotyping for Male and Female BALB/c-nu Mice Transplanted with Xeno-Free-Three-Dimensional Cell Masss in 13-Week Toxicity Study
Significant difference compared to the control (p < 0.01).
Spleen weight × percentage/100.
B, B cells; NK, natural killer; T, T cells; Tc, cytotoxic T cells; Th, helper T cells.
Absolute and relative organ weight in the male and female mice injected with XF-3DCMs did not show any palpable difference compared with the vehicle controls. There were no toxicological findings according to macroscopic examinations in males or females, including vehicle controls. Histopathological analysis of sampled tissue from the site of injection also revealed no toxicological findings in the vehicle control and both the XF-3DCM groups with a 1 × 106 dose and a 5 × 106 dose (Fig. 8B). Even in this case, the lower dose XF-3DCM group showed mononuclear cell infiltration between muscle fibers and the higher dose XF-3DCM group depicted only granulomatous inflammation at the site of injection (Fig. 8B, dotted elliptical curve). This finding seemed to indicate infiltration of histiocytes and lymphocytes. Male mice treated with 5 × 106 cells per animal of XF-3DCMs demonstrated lymphoid hyperplasia in the spleen (data not shown). However, these symptoms were also noticed for female mice in the control group, suggesting that these were not toxicologically meaningful.
Discussion
Numerous studies have focused on promoting angiogenesis with stem cell therapy in several types of ischemic diseases. However, to reach a clinical grade of stem cell therapy, safety and standardization in the manufacturing process are important. Early studies at the laboratory level have suggested the possibility of stem cell therapy by confirming the therapeutic effect of stem cells proliferated or differentiated in a medium containing FBS. FBS-supplemented media were observed to be beneficial for cell proliferation or maintenance. However, when stem cells proceed to the clinical stage, xenogeneic substances have negative influences on safety such as immune rejection.
Currently, 3D spheroid culture technology has been given research attention, and studies for developing a therapeutic agent using 3D spheroids have increased in number. In 3D cultures, when the cells are cultured in a xenogeneic medium, it is difficult to remove xenogeneic residues trapped in the 3D structure, whereas in 2D cultures, the cultured cells are extensively washed with a solution to remove xenogeneic remnants before being used in therapeutic applications. Therefore, to develop therapeutically safe 3D cultures, cells need to be cultured in a medium that is not contaminated with xenogeneic materials. In previous work, we also developed 3DCMs using an MBP-FGF2-immobilized surface under an FBS-containing culture condition and demonstrated the resulting therapeutic effect in CLI, 32 spinal cord injury, 33 and cardiac infarct animal models. 20 In this study, to advance 3DCMs to the clinical phase, XF-3DCMs were cultured under an XF condition and compared with 3DCMs established in the presence of FBS. Furthermore, the therapeutic efficacy equivalence of XF-3DCMs was assessed in a CLI animal model and its nonclinical safety testing was performed. hASCs in XF-3DCMs secreted three times more VEGF than FBS-3DCMs (Fig. 2C). However, bFGF and IL-8 expression were not observed to be very different between XF-3DCMs and FBS-3DCMs. The difference in the amount of angiogenic factor expressed in 3D cultures depending on the composition of the medium may help to develop a culture medium capable of achieving the optimal therapeutic effect. This is especially true in cases where the kind and amount of angiogenic factor can be correlated with the therapeutic effect such as in the case of angiogenesis.
The transplanted XF-3DCMs of hASCs exhibited increased blood flow (Fig. 3B) and improved therapeutic effect (limb salvage in the ischemic region) (Fig. 3A, C), substantially reduced fibrosis (Fig. 6B), and attenuated muscle degeneration (Fig. 6A) equal to that in the FBS-3DCM-transplanted group, indicating that hASCs might directly and actively promote angiogenesis in ischemic mice. However, it remains unclear whether the implanted hASCs can be directly differentiated into endothelial cells to form part of the newly formed vessels. As shown in Figure 5A, CD31-positive cells found in the blood vessels of the ischemic area in XF-3DCM-transplanted mice were also positive for HNA (white arrowheads in the inset). This indicates that the implanted hASCs may be differentiated into endothelial cells and incorporated into the host vasculature. Furthermore, considerable numbers of implanted cells were found in the vicinity of the vasculature in the ischemic region, suggesting that the cells promote angiogenesis through paracrine factor secretion. The enhanced survival of cells in the XF-3DCM group compared to the XF-hASC group may be due to prevention of anoikis together with increased expression of proangiogenic factors such as VEGF and bFGF. Overall, XF-3DCM transplantation appeared to contribute to angiogenesis and muscle regeneration in ischemic mice as seen by immunofluorescence, histological and physiological improvement in ischemic limbs treated with XF-3DCMs. It was previously reported that hMSCs cultured in animal serum-free media might provide additional advantages. For example, XF media might increase the clonogenic potential 34 and enhance the anti-inflammatory properties of hMSCs. 35 Recent studies demonstrated that hMSCs exhibited enhanced anti-inflammatory properties, as evidenced by elevated levels of IL-24 or TSG-6 when being cultured as spheroids.36,37 hASC spheroids also showed enhanced therapeutic angiogenesis in various ischemic diseases owing to enhancement of cell survival and retention.19,20,32 The existence of hypoxic conditions may precondition the hASCs so that they can survive in the harsh hypoxic conditions of the target ischemic tissue. In recent studies, it has been reported that hypoxic preconditioning results in improved therapeutic potential of hMSCs. 18 Several studies have reported that hASCs formed as spheroids show upregulation of hypoxia-adaptive signals, antiapoptotic and anti-inflammatory molecules, and preservation of extracellular matrix compared with monolayer cultures.19,21,38 Taken together, these studies suggest that the anti-inflammatory properties, higher survival rate and differentiation potential, and therapeutic efficacy of hASCs may be further improved in XF formulations supporting cultures of cells as 3D spheroids, compared to those developed in media containing FBS. We believe that XF-3DCMs, which have excellent therapeutic effects in CLI, will be available for clinical application if safety can be secured against toxicity such as tumorigenicity or immunogenicity.
While clinical applications are moving forward, there are concerns that implantation of stem cells might lead to the formation of tumors in the recipient. It has been reported that the initiation of tumor formation can be influenced by the microenvironment at the transplantation site or the ultimate homing site of the host and depends on the cell types involved. 39 However, the possible risk of hASCs in tumor progress and growth is controversial. According to research of MacIsaac et al. and Kucerova et al., no evidence was found for tumorigenicity in immunocompromised animals injected with human adipose-tissue-derived MSCs.40,41 In contrast, there is evidence that indicates that hASCs promote tumor initiation and accelerate tumor growth through IL-6 production. 42 These controversies highlight the necessity of investigating carefully the transformation of hASCs when being prepared for human clinical use. In this study, we evaluated the tumorigenicity of XF-3DCMs in vitro and in vivo through soft agar colony formation assay and transplantation into SCID mice, respectively. Soft agar colony formation assay is used to measure cell anchorage-independent proliferation potential. Anchorage-independent growth is a hallmark of transformation and the most accurate in vitro indication of tumorigenicity. 43 Our results indicate that XF-3DCMs do not possess anchorage-independent proliferation potential (Fig. 7A). Furthermore, in our 13-week tumorigenicity study, in which XF-3DCMs were transplanted into female SCID mice, there were no tumor formation-related findings in terms of body weight, abnormal clinical signs, or macroscopic and microscopic examination. However, all HeLa cell-injected mice show increased tumor volume (Fig. 7C). These findings correlate with their clinical observations and microscopic examinations (data not shown). In addition, IHC images revealed no existence of HNA expression in the XF-3DCM-injected groups, indicating a lack of human cell persistence (Fig. 7E). However, the tumors detected in the HeLa cell-injected mice likely originated from the transplanted HeLa cells based on the detection of HNA expression (Fig. 7E). Our results obtained from in vivo and in vitro tumorigenicity studies consistently demonstrated that XF-3DCMs did not induce tumor formation. These data are consistent with a previous study, which reported no evidence of tumor development in mice transplanted with human adipose tissue-derived MSCs. 44
To examine the toxicity of XF-3DCMs, we injected XF-3DCMs at two different concentrations (1 × 106 and 5 × 106 cells/animal) into male and female BALB/c nude mice and evaluated their toxicity potential for 4 weeks. No mortality, clinical signs, body weight, organ weight, or microscopic and macroscopic changes were found. Although few changes were observed in hematological (Table 1), serum biochemistry (Table 2), and lymphocyte analysis (Table 3), variation between individuals was large, and animals did not show any changes likely to be toxicologically relevant. Based on these results, we concluded that there is no toxicologically relevant effect and no observed adverse effects for XF-3DCMs.
For clinical trial, stem cells need to be generated in an XF environment. In this study, we prepared 3DCMs of hASCs using an XF medium and confirmed their angiogenic factor secretion and cell viability in vitro. In addition, we demonstrated that XF-3DCMs have therapeutic potential and did not induce any toxicological effects in vivo. XF-3DCMs can be manufactured as a clinical grade and is available as a novel treatment for therapeutic angiogenesis. For the clinical and practical use of XF-3DCMs, the next task will be to establish a standardized manufacturing process for quality control such as size, shape, survival rate, and efficacy analysis of XF-3DCMs.
Conclusion
We have shown that hASCs formed 3DCMs in XF conditions on an MBP-FGF2-immobilized substrate. The majority of hASCs in XF-3DCMs exhibited similar viability and almost equal or enhanced secretion of proangiogenic factors compared to those in 3DCMs formed in FBS-containing media. Transplanted XF-3DCMs augmented host neovascularization and the regeneration of ischemic tissue in the hindlimb ischemia model similar to 3DCMs generated in FBS-containing media. The transplanted cells remained in the ischemic region for 28 days and few cells were directly incorporated into newly formed microvessels. Moreover, our data showed no signs of tumorigenicity or toxicological side effects for XF-3DCMs in vitro and in vivo, providing evidence of the safety of XF-3DCMs for clinical applications. Based on in vitro and in vivo therapeutic efficacy and safety of XF-3DCMs, we conclude that 3DCMs of hASCs will be applicable to therapeutic angiogenesis in clinical trials of peripheral artery disease in the future.
Footnotes
Acknowledgment
This work was supported, in part, by a grant (HI16C2234) from the Korea Health Industry Development Institute, Ministry of Health and Welfare, Republic of Korea.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.