
Abstract
Platelet-rich plasma (PRP) and mesenchymal stem cells (MSCs) are promising tools for muscular-skeletal regeneration. However, increasing evidence has demonstrated controversial effects of PRP on the tissue regeneration. To obtain optimum PRP concentrations for MSC expansion and to accurately control osteogenic, adipocytic, and chondrogenic differentiation, MSCs were exposed to PRP alone or in combination with induction medium. We found that PRPs with the platelet concentration beyond 1500 × 109 pl/L were preferable to promote MSC proliferation. In addition, PRPs ranging from 200 × 109 to 3000 × 109 pl/L were capable of augmenting MSC osteogenesis, and PRP with 1500 × 109 pl/L was most effective for MSC osteogenic differentiation. Furthermore, PRPs in low platelet concentration range just slightly promoted MSC adipogenesis, and only when the platelet concentration was beyond 1800 × 109 pl/L, the promoted effects were evident. Moreover, PRP range from 1000 to 3000 × 109 pl/L significantly enhanced chondrogenesis of MSCs in the absence and presence of chondrogenic induction medium, and PRP with 2000 × 109 pl/L was more effective for MSC chondrogenesis. Furthermore, we explored the mechanisms of PRP-induced MSC differentiation, showing that the growth factors played a major role in this process while other unknown factors may also be involved in it. At last, we measured the levels of cytokines to learn that PRP treatment suppressed the secretion of interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-α, but favored the production of IL-10 by MSCs. In summary, our findings demonstrated that PRPs with different concentrations of platelets exerted different effects on proliferation and differentiation of MSCs, which indicated that preparing appropriate PRPs may be a precise and efficient strategy for improving MSC-based tissue regeneration.
Impact Statement
In the current study, we screened the effects of different concentrations of platelet-rich plasma (PRP) on the mesenchymal stem cell (MSC) proliferation and differentiation. Our data demonstrated that concentrated PRP exerted different effects on cell proliferation and on the osteogenic, adipogenic, and chondrogenic differentiation of human bone marrow-derived MSCs. Meanwhile, we optimized PRP concentrations for each of these lineage differentiation and MSC expansion and, thus, suggested several promising clinical strategies for different kinds of diseases. These findings may help explain the controversial effects of PRP on MSCs and improve the progress in precise applications of PRP-based regenerative strategies.
Introduction
Platelet-rich plasma (PRP) is a volume of plasma that has a platelet count above baseline and is usually purified from autologous whole blood. 1 Accumulating proof has demonstrated that PRPs were composed of a cocktail of growth factors, such as basic fibroblast growth factor (bFGF), platelet-derived growth factor AB (PDGF-AB), insulin-like growth factor 1 (IGF1), and transforming growth factor β1 (TGF-β1), playing important roles in tissue regeneration.2,3 In addition, fibrins in PRP act as scaffolds for seed cell migration and differentiation.4,5 Furthermore, platelets store coagulation factors, antibacterial proteins, and chemokines. Therefore, PRP is defined as a potent approach in sports medicine and orthopedics, which has been widely used in the therapeutics of skeletal disorders, including arthritis, bone defects, cartilage injury, tendon, and muscle damage.6–9
Mesenchymal stem cells (MSCs) are originally identified from bone marrow and now have been successfully isolated from other connective tissues.10–12 MSCs and their progenies are important supporting cells in the hematopoietic stem cell niche.13,14 In addition, MSCs are able to suppress immune cell development and function in a dose-dependent manner by means of secretory factors and/or direct contact. High proliferation capacity is another vital characteristic of MSCs to generate a large quantity of cells for cell therapy. Most importantly, MSCs can differentiate into multiple tissue types, including bone, fat, and cartilage, under certain conditions.13–15 Therefore, exogenous MSC-based therapies for the treatment of diseases, especially for promoting skeletal regeneration, have been the focus of intensive efforts in the past decades.16–21 Alternatively, exciting strategies have been developed to target endogenous MSCs to reprogram their lineage commitment and improve their regeneration potentials.
Based on the increasing evidence of PRP trials and to improve MSC-based therapy, investigators tend to explore the modulatory effects of PRP on MSCs in regenerative applications. However, to date, the biological changes of MSCs post PRP stimulation, especially the changes of multiple differentiation of MSCs, still remain controversial.22–26 Chen et al. 27 demonstrated that human adipose derived stem cells (ADSCs) steadily differentiated into osteoblasts when these cells were embedded in activated PRP gel. In addition, Chieregato et al. 28 found that exposure to human PRP increased osteogenic differentiation of ADSCs. In contrast, Lohmann and colleagues 29 independently reported that osteogenic activity of MSCs decreased when exposed to PRP from an older donor or PRP with heparin at a high concentration. Similar to their controversial effects on osteogenesis, the modulatory properties of PRP on MSC adipogenic differentiation are also undefined. Chieregato et al. 28 observed increased adipogenic activity of MSCs cultured with PRP. However, Cervelli et al. 30 showed the promoting effects of MSCs in an insulin-dependent manner. In contrast, Reinisch et al. 31 and Lange et al. 32 observed impaired adipogenesis when human ADSCs or bone marrow MSCs were cultivated in PRP buffered medium. Articular cartilage has special nutritional requirements and is extremely difficult to repair spontaneously after injury because of lack of blood supply. Although PRP treatments have been demonstrated to potentiate cartilage repair in vivo and in vitro, little information about the appropriate PRP concentration for promoting MSC chondrogenesis is available. 33 Moreover, anti-inflammatory effects are important factors to be considered for MSC-based therapy. By secreting numerous cytokines, MSCs regulate the functions of most inflammatory cells and improve tissue regeneration.34,35 However, it remains unknown whether PRP influences the cytokine profile of MSCs.
Therefore, the objective of the current study was to explore the appropriate PRP concentration to guide MSC differentiation and proliferation to improve MSC-based tissue repair. We establish the hypothesis that concentrated PRP may exert different modulatory effects on the osteogenic, adipogenic, and chondrogenic differentiation of MSCs.
Materials and Methods
Ethics statement
This study was approved by the Institutional Ethics Review Board of the People's Liberation Army General Hospital, and informed consent was obtained from all donors (CAAE: 14878813.4.0000.5533).
Preparation of PRP with different platelet concentrations
Whole blood samples were obtained from eight donors, consisting of four men and four women with an average age of 39 years old (39.3 ± 5.8 years). All samples were used to prepare PRP individually instead of in a pooled manner. Briefly, 200 mL of whole blood was drawn from the antecubital vein of each donor into a disposable blood bag containing 10 mL ethylenediamine tetra-acetate (EDTA) as an anticoagulant by a licensed phlebotomist. A measure of 0.5 mL blood was drawn from each sample for a complete blood count, and the rest was divided equally in ten 20-mL sterile Vacutainers to prepare PRP by double centrifugation. Each volume (20 mL) was centrifuged at 300 g for 10 min, and then the whole upper phase and the top red layer (1–2 mm) were reserved for a second centrifugation at 1200 g for 10 min. Approximately 1–2 mL of volume at the bottom was reserved, while the upper 85% phase was discarded. The platelet concentration and the residual number of leukocytes in the PRP were determined with a cytoanalyzer. Different volumes of plasma in the upper phase were added to create a series of PRPs with concentrations of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109, 2700 × 109, and 3000 × 109 platelets (pl)/L. Serum was considered PRP with 0 × 109 pl/L. Centrifugation was performed at room temperature. All PRPs were activated with a mixture of 10% CaCl2 and bovine thrombin (250 U/mL) using the following protocol: 1 part of the mixture was added to 9 parts of PRP, and the resulting solution was incubated at 37°C for 1 h. Then, the formed gel was mashed and centrifuged at 3000 g for 10 min to remove debris and collect the supernatant (the activated PRP). The activated PRP was stored at −80°C.
MSC isolation and expansion
Four milliliter of fresh human bone marrow aspirate was obtained from the eight donors. All bone marrow samples were used to culture and identify MSC individually instead of in a pooled manner. The samples were washed twice in phosphate-buffered saline (PBS) to remove residual blood, brought up to 6 mL with PBS, and then poured gently into a declivate 10-mL centrifuge tube containing 4 mL of lymphocyte separation medium, keeping the two phases clearly separate. The tube was spun at 900 g for 20 min at 4°C, separating the samples into three phases. The middle layer was removed and washed twice with PBS, and then, the pellet in the bottom was resuspended and transplanted in alpha-minimal essential medium (α-MEM) supplemented with 10% vol/vol fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin and incubated at 37°C in an atmosphere of 5% CO2/95% air. The medium was changed every 3 days, and cells were passaged and reseeded at a cell density of 5 × 104/cm2 after cells reached 80% confluency.
Flow cytometry analysis
The cell surface antigen profile of MSCs was analyzed by flow cytometry. Phycoerythrin (PE)-conjugated monoclonal antibodies against human CD29, CD31, CD34, and CD44 and Fluorescein isothiocyanate-conjugated monoclonal antibodies against human CD45, CD51, CD73, CD105, and HLA-DR were used in addition to allophycocyanin-conjugated antibodies against CD90. The purity and integrity of PRP were characterized with the markers of CD41a-PE and CD42b-FITC. All products were purchased from eBio-Science. Flow cytometry analysis was performed according to a previous protocol. Briefly, the MSCs and PRP were washed twice with PBS before incubation with or without antibodies for 20 min at 4°C in the dark. Then, each aliquot was washed twice with PBS and fixed with 0.4 mL of 1% paraformaldehyde for flow cytometry analysis. The data were analyzed with WinMDI 2.9 software.
Transmission electron microscopy (TEM) of PRP and MSCs
For the ultrastructural observations of transmission electron microscopy (TEM; HITACHI, Tokyo, Japan), 5 × 105 MSCs and 0.2 mL PRP were collected and fixed for 4 h at 4°C in 5% glutaraldehyde, washed twice in 0.1 mol/L PBS, postfixed for 2 h at 4°C in 2% osmium tetroxide, dehydrated in a graded series of ethanol solutions, embedded in Epon 812, cut into ultrathin sections (75 nm), and then stained with uranyl acetate and lead citrate. The sections were viewed and recorded with a HITACHI H-600 electron microscope at 80 kV.
Evaluation of the proliferation of MSCs
MSC proliferation was determined by Cell Counting Kit 8 (CCK-8; Dojindo) and carboxyfluorescein diacetate succinimidyl ester (CFSE) staining, according to the manufacturer's protocols. For the CCK-8 test, MSCs at passage 4 were seeded in 96-well plates (1 × 104/well, five wells in each group) and maintained in MSC culture medium supplemented with PRPs previously mentioned, and then, CCK-8 solution was added at a ratio of 100 μL/mL followed by incubation at 37°C for 1 h. Groups with PRP with 0 × 109 pl/L were considered the negative control. Absorbance was measured at a wavelength of 450 nm using a microplate reader. In the current study, the CCK-8 assays were performed at different time periods at day 1, 3, 6, 9, and 12.
For the CFSE-based assay, MSCs at passage 4 were loaded with CFSE as previously reported.35,36 In brief, MSCs were suspended at a concentration of 107 cells/mL in PBS containing 2% FBS before being incubated with 10 μM CFSE for 20 min in the dark, followed by the inhibition of CFSE incorporation by FBS. MSCs were then washed twice with PBS containing 2% FBS and reseeded at a density of 2 × 105/well in 6-well plates. Concentrated PRPs (0, 1500, and 3000 × 109 pl/L) were added into the culture system of CFSE-labeled MSCs at a 1:9 ratio of PRP:α-MEM. Groups with PRP with 0 × 109 pl/L were considered the negative control. At day 6, the MSCs from each group were harvested, and the dye dilution was assayed by FACSCalibur quantification. In some experiments, the growth factors present in the corresponding concentrated PRP alone were added into the MSC culture system to explore the potential mechanisms of PRP on MSC proliferation.
Evaluation of the multipotency of MSCs
To elucidate the controversial association between MSC differentiation and PRP exposure, osteogenic, adipogenic, and chondrogenic differentiation was assayed in the present study.
The previously reported protocols for MSC differentiation were used with minor revision in the current study.35,37 For osteogenic differentiation, MSCs at passage 4 were harvested and incubated in different concentrations of PRPs (0 × 109 (control group), 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109 pl/L, up to 2700 × 109 and 3000 × 109 pl/L) at a density of 5 × 103/well in 48-well cell culture dishes. To determine effects of PRP on MSC osteogenesis, cells at passage 4 were cultured in osteogenic induction medium (0.1 μM dexamethasone, 10 mM glycerol-2-phosphate, and 20 μM ascorbic acid) supplied by different concentrations of PRPs as described above. Groups with PRP of 0 × 109 pl/L were considered as the negative control. The osteogenic activity was assessed at day 14 using alkaline phosphatase (ALP) staining and at day 28 by Von Kossa staining, according to previous protocols, respectively.35,37
For adipogenic differentiation, MSCs were cultivated at 1 × 104 cells/well in 48-well cell culture plates. The PRPs described above were added into the culture medium to examine the direct effect of PRP on MSC adipogenesis. In addition, adipogenic induction medium (composed of 10−3 μM dexamethasone, 1 μM isobutylmethylxanthine [IBMX], and 10 ng/mL [wt/vol] insulin) was supplemented with different concentrations of PRP, as described before, and was added into the MSC culture system to detect the potential synergistic effects of PRP. Groups with PRP of 0 × 109 pl/L were considered as the negative control. The MSCs were maintained for 14 days before Oil-Red-O staining was performed by methods as we previously reported.35,37
For chondrogenic differentiation, MSCs were harvested and cultivated at a 1 × 104 cells/well in 48-well cell culture plates. The cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with different concentrations of PRP (0 × 109 [control group], 1000 × 109, 2000 × 109, and 3000 × 109 pl/L) to investigate the effects of PRP on MSC chondrogenesis. To detect potential synergistic effects, chondrogenic induction medium (10−4 μM dexamethasone, 1% [vol/vol] insulin-transferrin-sodium selenite [ITS], 50 μM ascorbate-2-phosphate, 1 mM sodium pyruvate, 50 μg/mL [wt/vol] proline, and 20 ng/mL [wt/vol] TGF-β3) supplemented with different concentrations of PRP was added into the MSC culture system. Groups with PRP with 0 × 109 pl/L were considered the negative control. After 14 days of induction, the expression of Sox-9 (SRY-type high-mobility group box-9) and Col-II (collagen type II) was detected by immunohistochemical assays as previously reported. 38 At 28 days of induction, Toluidine blue staining was performed to identify chondrocyte development and the secretion of cartilaginous matrix.35,37
The images were captured using an Omano OM159T biological trinocular microscope (Microscope Store, Roanoke, VA). The percentage of stained area within the total area, as well as the number of adipocytes and chondrocytes, was estimated using the computerized image analysis software Image-Pro Plus (IPP) 6.0.
Quantitative polymerase chain reaction
In the absence or presence of PRP, human bone marrow MSCs at passage 4 were maintained in osteogenic, adipogenic, and chondrogenic differentiation media at a density of 5 × 104 cells/well in 6-well cell culture plates for 10 days before being harvested for gene analysis. Total RNA was extracted from MSCs with TRIzol reagent (Fermentas) and reverse transcribed using the mRNA Selective PCR Kit (TaKaRa). Human RUNX2 (runt related transcription factor 2), OCN (osteocalcin), CEBP/α (CCAAT/enhancer binding protein alpha), PPARγ (peroxisome proliferator-activated receptor gamma), Sox-9, Col-I (collagen type I), and Col-II cDNA were amplified by real-time polymerase chain reaction (PCR) using a SYBR PCR Master Mix Kit (Sigma-Aldrich). The primers were synthesized by Invitrogen (Shanghai, China), and the sequences are shown in Supplementary Table S1.
Growth factor determination by enzyme-linked immunosorbent assay
Human TGF-β1, PDGF-AB, IGF1, and bFGF concentrations in primary PRPs from 8 donors and in concentrated PRPs (0 × 109 [serum], 1000 × 109, 1500 × 109, 1800 × 109, 2000 × 109, and 3000 × 109 pl/L) were determined according to the reagent protocols of the quantitative Enzyme-linked Immunosorbent Assay (ELISA) Kits (R&D Systems, Minneapolis). Optical density was determined at 450 nm as previously reported. 36
Investigation of growth factor effects alone on MSC proliferation and differentiation
The effects of growth factors alone (in the absence of platelets) at the concentrations seen in certain PRP preparations (for proliferation: 0 × 109, 1500 × 109, and 3000 × 109 pl/L; for osteogenesis: 0 × 109, 1500 × 109, and 3000 × 109 pl/L; for adipogenesis: 0 × 109, 1800 × 109, and 3000 × 109 pl/L; and for chondrogenesis: 0 × 109, 1000 × 109, 2000 × 109, and 3000 × 109 pl/L) on MSCs were evaluated. The growth factors (including bFGF, PDGF-AB, IGF1, and TGF-β1), corresponding to these platelet concentrations, were pooled and added into the MSC culture. Cell proliferation was also determined by CCK-8 (Dojindo) and CFSE staining as before, while osteogenic, adipogenic, and chondrogenic differentiation was evaluated by ALP, Von Kossa, Oil-Red-O, and Toluidine blue staining, as well as by quantitative PCR (qPCR) analysis, following the previous protocols used in the experiments for platelet concentration optimization. Groups with growth factor cocktails corresponding to PRP with 0 × 109 pl/L were considered the negative control.
Measurement of IL-1β, IL-6, IL-10, and TNF-α
MSCs at passage 4 were cultured at a density of 1 × 106 cells/well in 6-well cell culture plates and starved for at least 12 h in α-MEM without FBS supplementation. Different concentrations of PRP (0 × 109 [control group], 1500 × 109, and 3000 × 109 pl/L) were added into the culture system for another 24 h before the supernatants were collected to measure IL-1β, IL-6, IL-10, and TNF-α levels using specific ELISA Kits (R&D Systems) according to the manufacturer's protocol, respectively.
Statistical analysis
Data are presented as the mean ± standard error of the mean (SEM). One-way analysis of variance and the Bonferroni post hoc test were performed for statistical analysis using SPSS 18.0 software (SPSS, Inc., Chicago, IL). There were three independent experiments for each set of results. A value of p < 0.05 indicated statistical significance.
Results
PRP preparation and characterization
To prepare PRP from whole blood, each sample (20 mL) was divided into two phases after the first centrifugation at 300 g for 10 min. The lower phase was red blood cells; the upper one was platelet-poor plasma (PPP) and PRP layers (Fig. 1A, B). After discarding the lower volume (Fig. 1C), the upper PPP and PRP layers were subjected to a second centrifugation at 1200 g for 10 min to discard the PPP fraction (Fig. 1D, E). Finally, ∼2 mL PRP was achieved (Fig. 1F).

Preparation, ultrastructure, and immunophenotype of PRPs. A measure of 20 mL venous whole blood was placed in 20-mL sterile Vacutainer
The platelet number and the residual number of leukocytes in PRPs from each donor were measured using a cytoanalyzer (Tables 1 and 2). TEM images of platelets in the PRP showed that the cells were oval, and several pockets were in the cytoplasm surrounded by several fragments (Fig. 1G). These features indicated that the platelets in the PRP were both integrated and functional. PRP samples were positive for the markers CD41a-PE and CD42b-FITC (90.09% ± 2.66%), which showed a low positive rate in the control group, demonstrating the acquisition of qualified and functional PRP (Fig. 1H).
Platelet and Growth Factor Concentrations of Donors
b-FGF, basic fibroblast growth factor; F, female; IGF1, insulin-like growth factor 1; M, male; PDGF-AB, platelet-derived growth factor-AB; Pl, platelets; TGF-β1, transforming growth factor β1; MSCs, mesenchymal stem cells.
Leukocytes in Whole Blood and Platelet-Rich Plasma
Leukocyte concentration in PRP was significantly lower than that in whole blood (p < 0.001). Data are represented as mean ± SEM, n = 8.
p < 0.001.
PRP, platelet-rich plasma; SEM, standard error of the mean; WB, whole blood.
Isolation and definition of human bone marrow MSCs
After 1 week of initial MSC cultures, the cells were observed to have spindle-shaped morphology in a monolayer (Supplementary Fig. S1A). MSCs reached 80–90% confluency 2 weeks after initial seeding and were passaged at a 1:2 ratio every 2 days for the following passages. The MSC number harvested from each donor is shown in Table 1.
To examine the multidifferentiation capacity, several specific cytochemical assays, such as ALP, Von Kossa, and Oil-Red-O staining, were performed. As shown in Supplementary Figure S1B, strong ALP activity and mineralization were observed in the osteogenic cultures. In addition, obvious lipid droplets accumulated when MSCs were exposed to the adipogenic medium (Supplementary Fig. S1B). These results confirmed the multipotency of MSCs in the present study.
Furthermore, the results of flow cytometric analysis showed that these cells were homogeneously positive for mesenchymal cell markers, CD29 (91.8% ± 3.3%) and CD44 (92.6% ± 2.1%), and “stemness” associated markers, CD73 (80.35% ± 1.5%), CD105 (60.99% ± 3.2%), and CD90 (96.89% ± 1.5%), on the cell surface and negative for the endothelial or hematopoietic markers, CD51 (0.37% ± 0.05%), CD31 (0.39% ± 0.8%), CD45 (0.46% ± 0.1%), CD34 (0.15% ± 0.12%), and HLADR (0.24% ± 0.09%) (Supplementary Fig. S1C).37–39 The data validate the purity of MSCs for the subsequent experiments.
MSC proliferation in different concentrations of PRP
MSCs are distinguished from other seed cells for skeletal regeneration, partially due to their multidifferentiation potential and high proliferation capacity. To examine the effect of PRP on MSC proliferation, the CCK-8 test was performed in the present study. The results of the CCK-8-based cell growth assay demonstrated that PRP-treated MSCs showed enhanced cell growth compared to MSCs in the control groups only when the PRP concentration ranged from 200 × 109 to 1500 × 109 pl/L (Fig. 2A). It is noteworthy that the growth curves were smooth and nearly parallel when the PRP concentration ranged from 1500 × 109 to 3000 × 109 pl/L, which indicated that the promoting effects of PRP on MSC proliferation could not increase indefinitely with increasing PRP concentration, and the optimum concentration PRPs for MSC proliferation may be 1500 × 109 pl/L.

MSC proliferation in response to different concentrations of PRPs. CCK-8 test was performed to examine the effect of different concentrations of PRP (range from 200 × 109 to 3000 × 109 pl/L) on MSC proliferation at day 1, 3, 6, 9, and 12 as showed in
CFSE staining is a live fluorescence assay that reveals cell proliferation by evaluating fluorescence dilution caused by cell division. As shown in Figure 2B, PRPs with 1500 and 3000 × 109 pl/L markedly stimulated MSC proliferation, but no obvious differences were observed between these two PRP groups, which confirmed the results of the CCK-8 assay and suggested that PRP with 1500 × 109 pl/L may be economical and efficient for MSC expansion.
Osteogenic differentiation of MSCs in different concentrations of PRP
To explore the effects of PRP on osteogenic differentiation, MSCs were treated with PRP at graded concentrations in the absence or presence of osteogenic agents. The results of ALP staining documented that PRP independently promoted MSC differentiation into osteoblasts in a dose-dependent manner when the PRP concentration ranged from 200 × 109 to 1500 × 109 pl/L (Fig. 3A). However, the promoting effect gradually decreased when the PRP concentration went above 1500 × 109 pl/L (Fig. 3A). The quantitative analysis of the ALP stained area showed a similar trend. As shown in Figure 3B, the ALP-positive area/total area ratio increased 2- to 12 fold in MSC cultures before falling steeply when the PRP concentration reached 1800 × 109 pl/L. In addition, the Von Kossa staining results demonstrated that PRP stimulated osteogenic mineralization of MSCs in a dose-dependent manner when the platelet concentration was below 1500 × 109 pl/L, and this stimulation decreased as the concentration went above 1800 × 109 pl/L (Fig. 3C). The quantitative analysis showed a similar tendency (Fig. 3D). These findings suggested that PRPs with different platelet concentrations have different capacities for MSC osteogenesis, and the optimum platelet concentration may be 1500 × 109 pl/L.
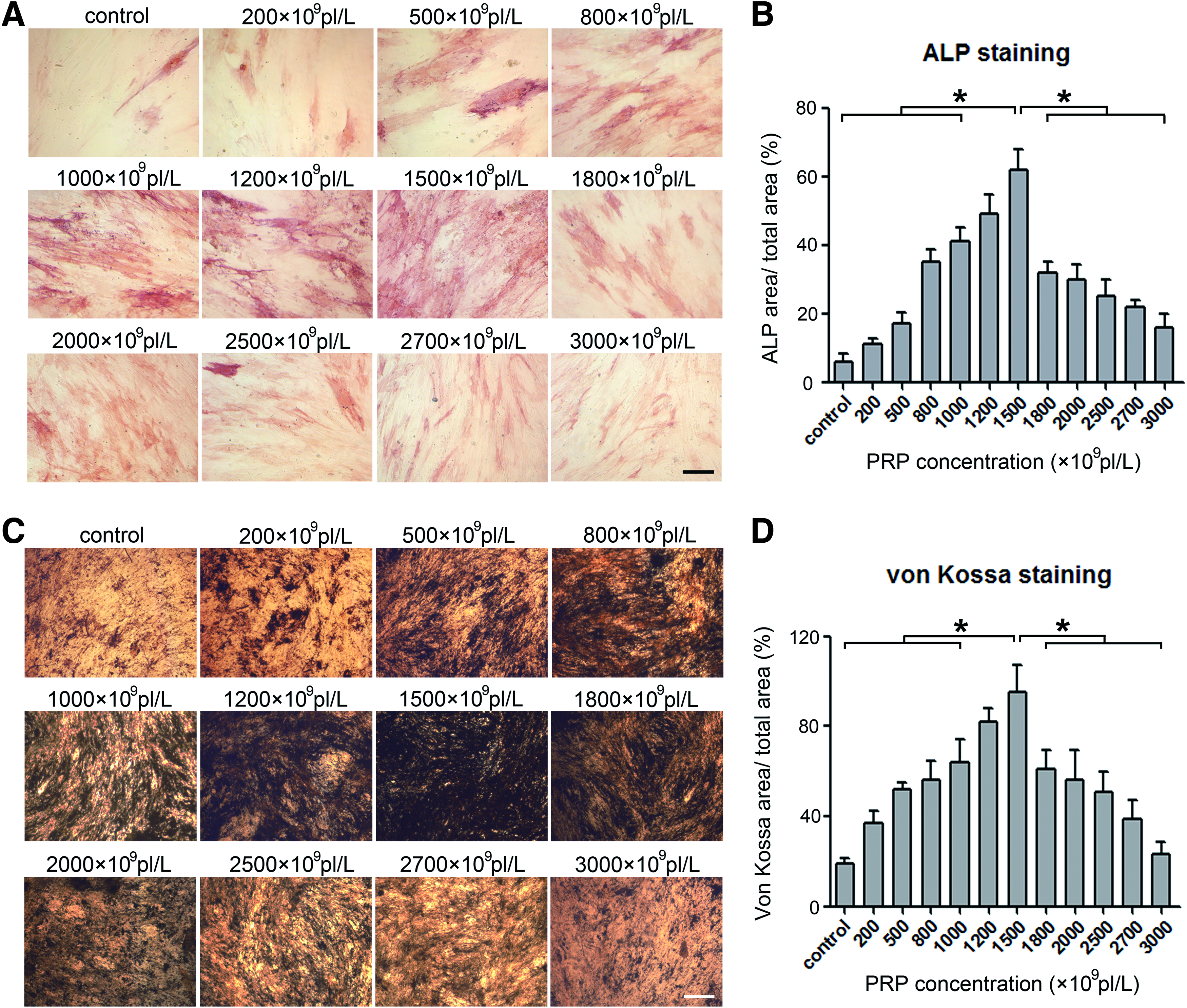
Evaluation of ALP activity and mineralization in MSC culture after being exposed in different concentrations of PRP. The results of ALP staining analysis revealed the promoting effects of different concentrations of PRP (range of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109, 2700 × 109, and 3000 × 109 pl/L) on osteogenesis
In the osteogenic medium, PRP supplementation significantly enhanced osteogenic differentiation when the concentrations of PRP were below 1500 × 109 pl/L. In addition, there was a steep decrease in ALP activity when the PRP concentration reached 1800 × 109 pl/L. However, ALP activity rose again when the PRP concentration further increased (Fig. 4A, B). Likewise, the Von Kossa staining of mineralization showed that PRP combined with inducing agents induced a similar osteogenic tendency as shown by ALP staining (Fig. 4C, D). These findings demonstrated that PRP supplemented with induction medium could yield two peaks of osteogenic activity, which may result from a synergistic effect of PRP and the induction medium, suggesting that even under osteogenic induction conditions, the optimum platelet concentration in PRP for MSC osteogenesis should be 1500 × 109 pl/L.

Evaluation of ALP activity and mineralization in MSC culture after being exposed in different concentrations of PRPs in combination with induction medium. ALP activity was measured to determine the promoting effects of different concentrations of PRP (range of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109, 2700 × 109, and 3000 × 109 pl/L) on osteogenesis
RUNX2 and osteocalcin (OCN) are important genes for differentiated osteoblasts. Complementing the histochemical analyses, MSCs were found to exhibit increased mRNA expression of RUNX2 and OCN after PRP exposure in the absence or presence of osteogenic agents (Fig. 5A–D). In addition, the expression levels of RUNX2 and OCN fluctuated over different concentrations of PRP, in accordance with the ALP and Von Kossa staining results (Fig. 5A–D), which further indicated that the platelet concentration played a pivotal role in controlling osteogenic activity at the transcriptional level and that PRP with 1500 × 109 pl/L might be the most effective for MSC osteogenic differentiation.

The determination of the osteogenic gene expression in MSC culture. OCN and RUNX2 mRNA expression levels were examined after stimulation with different concentrations of PRP alone (range of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109 pl/L, up to 2700 × 109 and 3000 × 109 pl/L)
Adipogenic differentiation of MSCs in different concentrations of PRP
Although numerous studies have reported the effects of PRP on adipogenic differentiation, the data remain controversial. In the current study, human bone marrow-derived MSCs were treated with different concentrations of PRPs alone or in combination with adipogenic medium. As shown in Figure 6A, intracytoplasmic lipid droplet accumulation in MSCs slightly increased when the PRP concentrations were below 1800 × 109 pl/L, whereas they became much more evident as the platelet concentration increased above 1800 × 109 pl/L. Further quantitative analysis of adipocyte number (Fig. 6B) and the ratio of Oil-Red-O stained area/total area (Fig. 6C) in MSC cultures also supported that the effective PRP concentrations for MSC adipogenesis were above 1800 × 109 pl/L, which were considered as high in frequently used PRPs.

Evaluation of adipocyte formation in MSC culture after treatment with different concentrations of PRPs. The results of Oil-Red-O staining revealed the enhanced adipogenesis of MSCs treated with different concentrations of PRP (range of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109, 2700 × 109, and 3000 × 109 pl/L)
Moreover, even PRP with low platelet concentrations showed the ability to enhance the adipogenic activity of MSCs in adipogenic medium. As shown in Figure 7A, the intracytoplasmic lipid droplets in these MSCs were larger than those in PRP only stimulated MSCs. With increasing concentrations of PRP in adipogenic medium, the number of adipocytes in the PRP-treated groups was comparable to that in the control groups (Fig. 7B), while the ratio of Oil-Red-O-positive area/total area in the MSC cultures increased dramatically in a PRP dose-dependent manner (Fig. 7C).
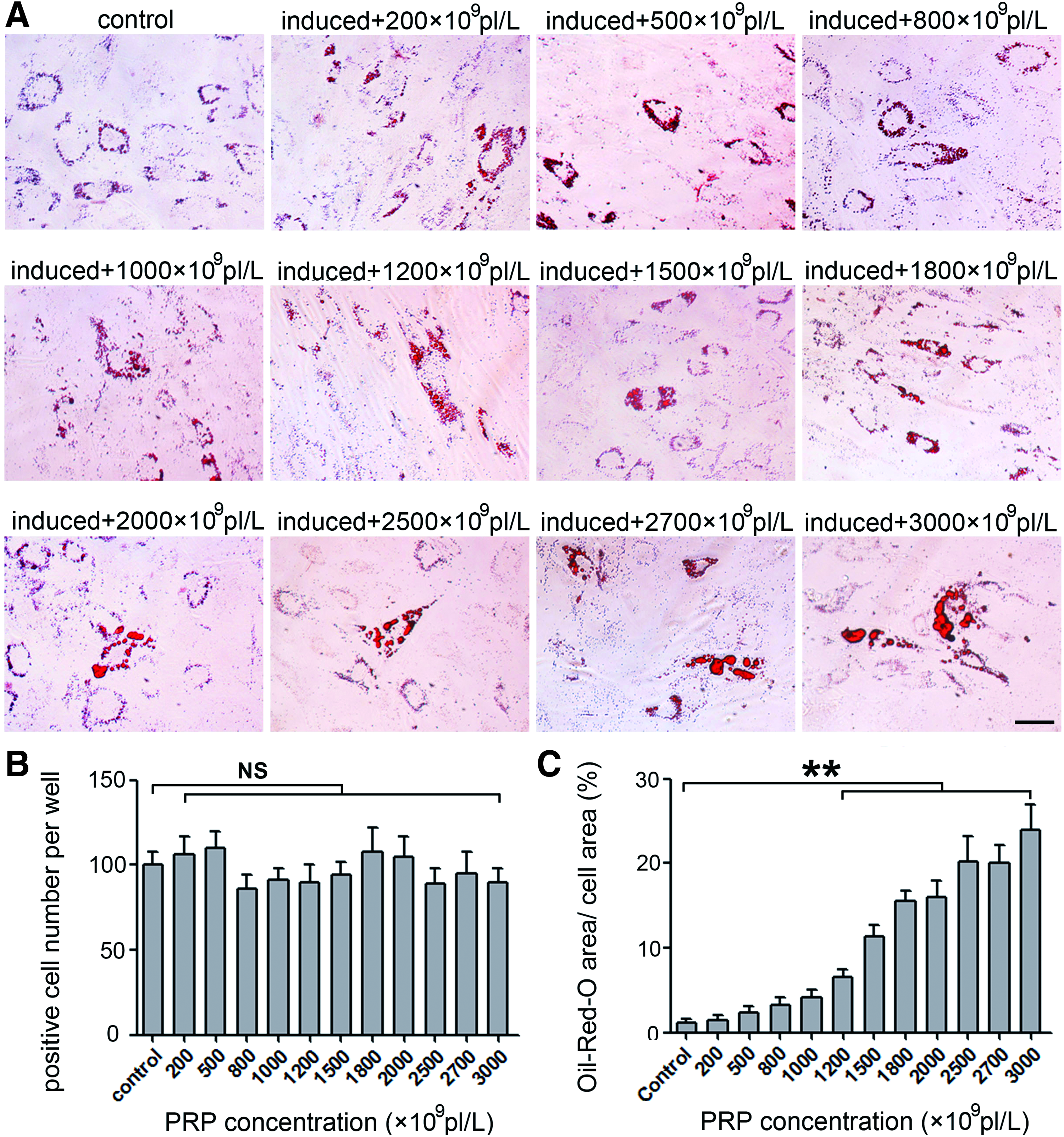
Adipogenesis in MSC culture exposed to adipogenic induction medium and different concentrations of PRP. The promoting effects of different concentrations of PRPs were evaluated through Oil-Red-O staining post 14 days cultivation in adipogenic medium combined with PRP (range of 200 × 109, 500 × 109, 800 × 109, 1000 × 109, 1200 × 109, 1500 × 109, 1800 × 109, 2000 × 109, 2500 × 109, 2700 × 109, and 3000 × 109 pl/L)
Complementary to the results of the histochemical analysis, those of the gene expression analysis showed that after exposure of MSCs to graded concentrations of PRP or PRP combined with adipogenic medium, the mRNA expression of peroxisome proliferator-activated receptor γ (PPARγ) and CEBP/α, which are molecular hallmarks of differentiated adipocytes, was detectable. As shown in Figure 8A and B, the mRNA expression levels of PPARγ were slightly elevated when the PRP concentrations were below 1800 × 109 pl/L, while an apparent increase was observed at concentrations above 1800 × 109 pl/L, which was consistent with the Oil-Red-O staining and further indicated that the effective PRP concentrations for MSC adipogenesis were above 1800 × 109 pl/L. The trends of CEBP/α mRNA expression levels were similar to those of PPARγ, providing additional evidence for the optimally effective platelet concentration (above 1800 × 109 pl/L) for MSC adipogenic differentiation (Fig. 8C, D).

The determination of the adipogenic gene expression in MSC culture. mRNA expression of PPARγ and CEBP/α, which are molecular hallmarks of differentiated adipocytes, documented enhanced adipogenesis of MSCs after exposure to graded concentrations of PRP or PRP combined with adipogenic medium
Chondrogenic differentiation of MSCs in different concentrations of PRP
To explore the appropriate PRP concentrations for MSC chondrogenic differentiation, the human bone marrow-derived MSCs were treated with different concentrations of PRPs (0, 1000, 2000, and 3000 × 109 pl/L) alone or in combination with chondrogenic agents. As indispensable regulators of chondrogenesis, the activities of Sox-9 and Col-II were detected. Compared with the cells in the control group, the number of Sox-9- and Col-II-positive cells increased when the platelet concentration in PRPs was increased (Fig. 9A, C). The results of immunohistochemistry showed that the expression of Sox-9 and Col-II in the 2000 × 109 pl/L group was markedly higher than that in the other groups. In addition, the results of Toluidine blue staining showed similar trends as the Sox-9 and Col-II activity, which suggested that PRP with 2000 × 109 pl/L may be preferable for the accumulation of proteoglycans (Fig. 9B, C). Complementing the histochemical analyses, the MSCs exhibited elevated mRNA expression of Sox-9 and Col-II as the concentration of PRP was increased (Fig. 9D, E). Furthermore, the mRNA expression of Col-II was increased compared to that of Col-I, which suggested that PRP favored the induction of hyaline cartilage instead of fibrocartilage (Supplementary Fig. S2A). Moreover, similar effects of PRP on the chondrogenesis of MSCs were observed in the presence of chondrogenic induction medium (Fig. 10A–E and Supplementary Fig. S2B). Taken together, the data above demonstrate that PRP with 2000 × 109 pl/L is effective and efficient for the chondrogenic differentiation of MSCs.

Chondrogenesis in MSC culture after being exposed to different concentrations of PRPs. The MSCs were maintained in concentrated PRPs (1000 × 109, 2000 × 109, and 3000 × 109 pl/L). The results of immunohistochemistry showed the discrepant effects of PRPs for chondrogenesis of MSCs

Evaluation of chondrogenesis in MSC culture after being treated by different concentrations of PRPs and chondrogenic induction medium. Sox-9 and Col-II immunohistochemistry labelings determined the effects of different concentrations of PRPs on MSC chondrogenesis
Effects of growth factor cocktails on MSC proliferation and differentiation
Accumulating data have demonstrated that growth factors in PRP modulate the multipotency and tissue-regenerative effects of MSCs.40,41 Therefore, in the present study, the concentrations of PRP-derived growth factors (bFGF, PDGF-AB, IGF1, and TGF-β1) in concentrated PRPs (0, 1000, 1500, 1800, 2000, and 3000 × 109 pl/L) were determined (Table 3). Then, the growth factors were pooled together and added into the MSC proliferation and differentiation systems.
Growth Factor Cocktails
As shown in Figure 11A, growth factor cocktails corresponding to PRPs with 1500 and 3000 × 109 pl/L markedly stimulated MSC proliferation, but no obvious differences were observed between the 1500 and 3000 × 109 pl/L groups, indicating that the growth factor cocktail corresponding to PRP with 1500 × 109 pl/L is appropriate for supporting MSC proliferation. In addition, the results of the CFSE assay further supported that the growth factor cocktail corresponding to PRP with 1500 × 109 pl/L is most appropriate for promoting MSC proliferation (Fig. 11B).

The effects of the growth factor cocktails corresponding to PRPs with certain platelet concentrations on MSC proliferation. CCK-8 assays were performed to examine the effects of growth factor cocktails that corresponded to concentrated PRPs (1500 and 3000 × 109 pl/L) on MSC proliferation at days 1, 3, 6, 9, and 12
For MSC osteogenesis, the results of ALP and Von Kossa staining demonstrated that the growth factor cocktail corresponding to PRPs with 1500 × 109 pl/L exhibited higher promoting effects than that corresponding to PRP with 3000 × 109 pl/L (Fig. 12A). As shown in Figure 12B, the ratio of the ALP-positive area increased fourfold in the MSC cultures after treatment with the growth factor cocktail corresponding to PRP with 1500 × 109 pl/L. For MSC adipogenesis, the 1800 × 109 pl/L groups exhibited a slight increase in Oil-Red-O stained area, while the 3000 × 109 pl/L groups showed notable intracytoplasmic lipid droplet accumulation, indicating that the growth factor cocktail corresponding to 3000 × 109 pl/L was appropriate for inducing MSC adipogenesis (Fig. 12C, D). In addition, the qPCR analysis results showed that the changes of mRNA levels of osteogenic genes and adipogenic genes were consistent with the histochemical staining results (Fig. 12E–H).

The effects of the growth factor cocktails corresponding to PRPs of certain platelet concentrations on MSC osteogenic and adipogenic differentiation. ALP and Von Kossa staining revealed the promoting effects of growth factor cocktails corresponding to the PRPs of 1000, 1500, and 3000 × 109 pl/L
Effects of growth factor cocktails on MSC chondrogenic differentiation were similar to those of their corresponding PRPs. Sox-9 and Col-II immunohistochemistry labeling documented a dose-dependent increase in MSC chondrogenesis by growth factor cocktails when their corresponding PRPs ranged from 0 to 2000 × 109 pl/L. However, the enhancement decreased when the PRP concentration was 3000 × 109 pl/L (Fig. 13A). In addition, the results of Toluidine blue staining supported that the growth factor cocktail corresponding to PRP with 2000 × 109 pl/L was preferable for the accumulation of proteoglycans (Fig. 13B). Consistently, the Sox-9- or Col-II-positive cell ratios and the ratio of Toluidine blue-stained area/total area in the 2000 × 109 pl/L groups were higher than those in the other groups (Fig. 13C). Further gene expression analysis demonstrated that Sox-9 and Col-II gene expression levels in the 2000 × 109 pl/L groups were the highest among all groups (Fig. 13D, E). Moreover, the Col-II gene expression levels in MSCs were higher than the Col-I gene expression levels after exposure to the growth factor cocktails (Supplementary Fig. S2C).

The effects of the growth factor cocktails corresponding to PRPs of certain platelet concentrations on MSC chondrogenesis. The MSCs were treated by the growth factor cocktails corresponding to PRPs of 1000, 2000, and 3000 × 109 pl/L. Sox-9 and Col-II immunohistochemistry labelings exhibited modulatory effects by those growth factor cocktails
In summary, the results above showed that the growth factors alone (in the absence of platelets) were capable of modulating MSC proliferation and differentiation in a similar way as the corresponding PRPs, but the effects were milder than those of the PRPs.
Change in cytokine secretion by PRP-treated MSCs
To investigate the influence of PRP with different platelet concentrations on secretory factors of MSCs, the production of IL-1β, IL-6, IL-10, and TNF-α in MSC supernatant was detected by ELISA. The results showed a significant increase in IL-10 production by MSCs that were treated with concentrated PRPs (Fig. 14). In contrast, the levels of IL-1β, IL-6, and TNF-α in MSC supernatants decreased with the increase in platelet concentrations of PRPs (Fig. 14). Therefore, these data indicate that PRP enhanced the anti-inflammatory capacity of MSC.

Change of the secretory cytokines of the PRP-treated MSCs. The production levels of IL-1β, IL-6, IL-10, and TNF-α showed discrepant influences of PRP in graded concentrations for MSC inflammatory response. Data are represented as mean ± SEM, n = 3, *p < 0.05; **p < 0.01. IL, interleukin; TNF-α, tumor necrosis factor α.
Subcellular changes in MSCs
To explore the possible mechanisms by which PRPs with different platelet concentrations exert discrepant effects on MSC biological properties, subcellular change of PRP-treated MSCs was tested. Representative images observed by TEM of the control (Fig. 15A) and PRP-supplemented groups (Fig. 15B–D) revealed a significant influence of different concentrations of PRP on the MSC subcellular structure. Notably, there was more karyokinesis in MSCs in the PRP-treated groups (Fig. 15C, D) compared to MSC cultures in basal medium (Fig. 15A). In addition, more dividing nucleoli and larger organelles, such as dictyosomes, were observed in PRP-treated MSCs (1500 × 109 and 3000 × 109 pl/L) than in MSCs of the control and 500 × 109 pl/L groups (Fig. 15A–D). Moreover, greater numbers of coarser claws in cell membranes were found in cross-sections of PRP-treated MSCs (especially in the 1500 × 109 and 3000 × 109 pl/L groups) relative to untreated ones. However, there were no significant differences in cell size per section between the groups with and without PRP. These results indicated that PRPs with platelet concentrations above 1500 × 109 pl/L were capable of dramatically promoting the osteo-adipogenic differentiation and proliferation of MSCs probably by enabling self-replication and anabolism more efficiently than other platelet concentrations.
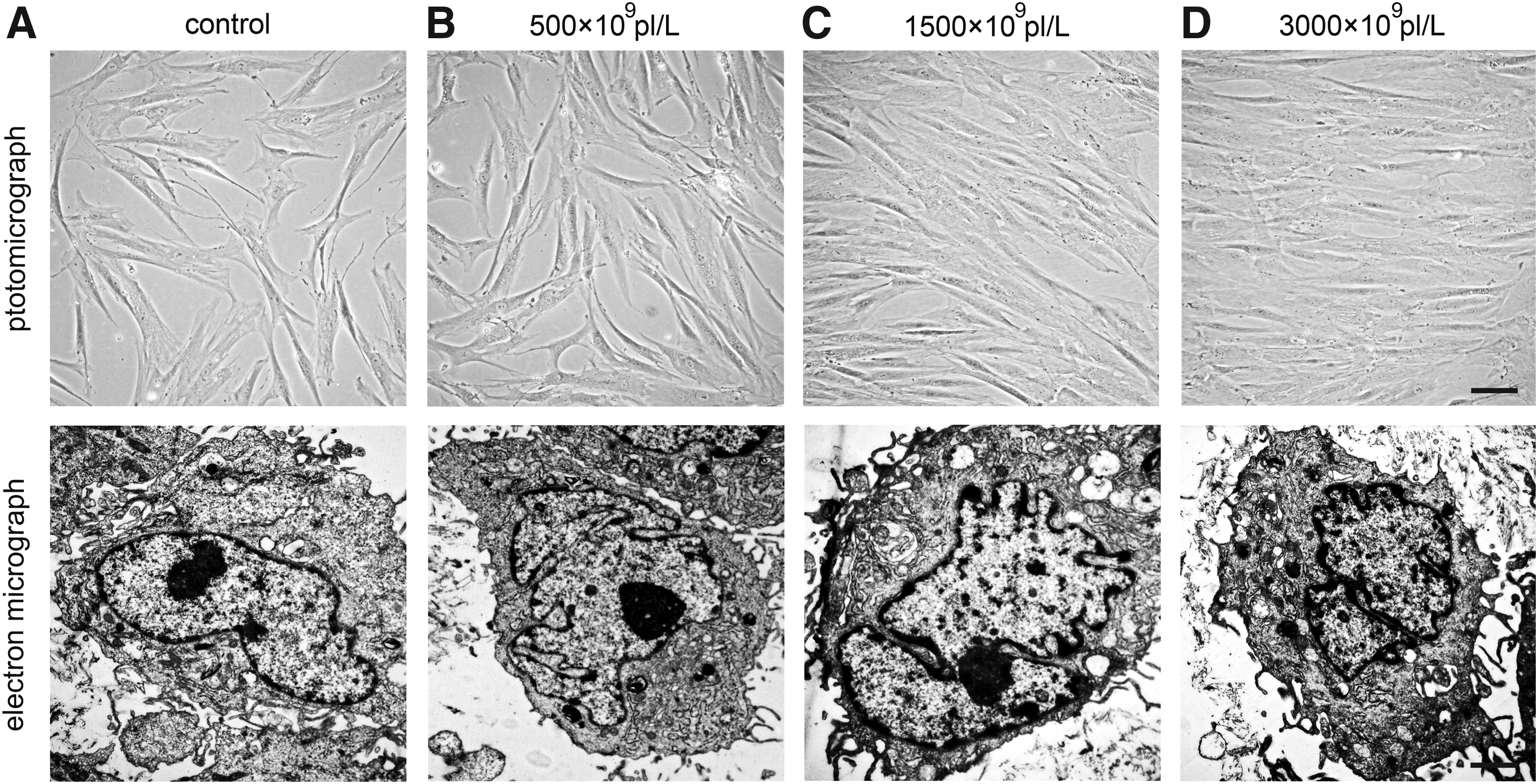
Subcellular changes of MSCs. MSCs
Discussion
In the current study, we screened the effects of PRP with different concentrations of platelets on MSC proliferation and differentiation. Our data demonstrated that concentrated PRPs exerted different modulatory effects on cell proliferation and the osteogenic, adipogenic, and chondrogenic differentiation of human bone marrow-derived MSCs. In addition, PRP-derived growth factors could mimic the effects of PRPs on the proliferation and differentiation of MSCs. Furthermore, PRP suppressed the production of inflammatory cytokines while promoting the secretion of the anti-inflammatory cytokine IL-10 by MSCs.
PRP has demonstrated favorable applications for wound healing, angiogenesis, and tissue remodeling, partially due to the high number of platelets and the many types of highly concentrated growth factors. 7 MSCs are attractive seed cells in regenerative medicine due to their multilineage differentiation capacity and easy expansion. However, the modulatory effects of PRP on MSC proliferation and multipotency are widely debated.31,40–42 The phenomenon could be explained by the variable quality of PRP in terms of platelet number and growth factor concentration. In addition, to our knowledge, there is insufficient information available about efficiently promoting MSC expansion and precisely directing MSC differentiation by optimizing PRP concentration.
In the present study, we first explored the appropriate concentration of PRPs for promoting the proliferation of MSCs. The PRPs we used contained different concentrations of platelets, ranging from 200 × 109 to 3000 × 109 pl/L. The concentrations covered most of the clinically used concentration range. 30 Our findings strongly suggest that PRP with 1500 × 109 pl/L is optimal for MSC proliferation. Obtaining sufficient MSC numbers for cell therapy requires efficient and effective in vitro expansion. The current methods for MSC expansion greatly depend on culture medium with FBS or serum-free medium. However, the risk of exon-antigen invasion and immune injection caused by FBS and the expensive cost of serum-free medium have hindered the large-scale expansion of clinically used MSCs. 33 The application of autologous blood-derived PRP overcomes these difficulties. Although several previous studies of MSC culture in PRP or platelet supernatant showed the enhancement of cell proliferation, the optimum strategy for the use of PRP for MSC expansion was lacking. 43 In the present study, we found that PRP with 1500 × 109 pl/L was preferable to that with other concentrations for MSC proliferation, which may be helpful for improving the PRP-based MSC expansion strategy.
Accumulating evidence has shown that PRP influences the multidifferentiation capacity of MSCs.40,42 However, the optimal PRP concentration that individually favors the osteogenesis, adipogenesis, or chondrogenesis of MSCs remained unexplored.
In the current study, we found that PRP alone increased MSC osteogenic differentiation in a dose-dependent manner when the platelet concentration ranged from 200 × 109/L to 1500 × 109/L, and then, the differentiation efficiency decreased smoothly from 1500 × 109/L to 3000 × 109/L. The findings indicated that the 1500 × 109/L may be preferable for the usage of PRP alone in promoting MSC osteogenesis. In addition, PRP was capable of synergistically promoting MSC osteogenesis in the presence of osteogenic induction medium. However, two peaks of osteogenic activity were observed in the presence of osteogenic agents. This phenomenon may lie with the different mechanisms between PRP-induced osteogenesis and osteogenic agent-induced osteogenesis. Among osteogenic agents, dexamethasone is a glucocorticoid hormone and an important modulator of bone differentiation from multipotential progenitors; β-glycerol phosphate is closely associated with mineralized bone development; and ascorbate-2-phosphate is vital for collagen formation and bone matrix mineralization.44–46 For PRP, regenerative factors are usually considered to be involved in the modulation of osteogenesis through specific signaling pathways.30,47,48 However, further investigations are needed to reveal the variety and the precise underlying mechanism of PRP-derived regenerative factors.
For a long time, the modulatory properties of PRP on MSC adipogenic differentiation remained controversial. 30 In the current study, we found that PRP concentration below 1800 × 109 pl/L only displayed a mild enhancement of MSC adipogenesis in the absence or presence of adipogenic induction medium. Importantly, adipogenic activity was significantly improved in the range of 2000 × 109 to 3000 × 109 pl/L. Notably, PRP with 3000 × 109 pl/L was most effective for adipogenesis but less effective for osteogenesis, which indicated that PRP with 3000 × 109 pl/L could be used to promote adipose tissue regeneration with less risk of heterotopic ossification. Moreover, the combined application of adipogenic agents and PRP yielded a synergistic effect in promoting MSC adipogenic differentiation. During differentiation, insulin enhances the conversion of glucose to CO2 and lipids. 37 IBMX increases not only the number of insulin receptors but also the specific insulin binding activity to its receptors. 37 Promisingly, this phenomenon is consistent with that reported in a previous study showing that PRP, in association with insulin, greatly potentiates adipogenesis in human ADSCs. 30 Therefore, our findings suggest that a higher concentration of PRP is superior to PRP with fewer platelets for promoting adipose tissue repair; moreover, adipogenic agents can strengthen the effects. Interestingly, there was a difference in the adipogenic change in cells grown with or without induction medium, contrary to what was observed in osteogenic media. The underlying mechanisms of this phenomenon need to be further clarified in future studies.
Many applications of PRP have been tried to restore hyaline cartilage lost in osteoarthritis and other cartilage defects by regulating endogenous or exogenous progenitor cells.49,50 Thus, seeking the appropriate PRP concentration for MSC chondrogenesis was important for cartilage repair. Our data showed that PRP with platelet concentrations of 1000 × 109 to 3000 × 109 pl/L supported MSC chondrogenesis. However, the promoting effect of PRP with 2000 × 109 pl/L was the most obvious in the current study. Sox-9 is one of the key transcription factors that acts very early in the chondrogenic lineage.51,52 In the present study, the expression of Sox-9 was upregulated when MSCs were treated with concentrated PRPs, which suggests that a Sox-9-related signal pathway may be involved in PRP-induced chondrogenesis. Most importantly, Sox-9 expression declined when the platelet concentration in PRP reached 3000 × 109 pl/L. This finding suggests that there might be a feedback mechanism by which chondrogenic activity in MSCs responds to higher concentrations of PRP. Col-II formation and proteoglycan accumulation are indispensable for the reconstruction of cartilage tissue structure.51,52 Our data in the current study showed that PRP with platelet concentration of 2000 × 109 pl/L was appropriate for both Col-II and proteoglycan expression, indicating that precise PRP treatment also influences the later phase of chondrogenesis in MSCs.
Collectively, we may further draw conclusions that 1500 × 109 pl/L, which is simultaneously optimum for MSC proliferation, most effective for MSC osteogenesis, and weak for adipogenesis, is the optimal concentration that could be used for osteogenesis-related injuries, such as avascular necrosis, atrophic nonunion, and osteoporosis. However, in the field of cosmetology, which needs more adipocytes and fewer osteoblasts, the optimal PRP concentration could be 3000 × 109 pl/L for the simultaneous enhancement of MSC proliferation and adipogenesis but the weak promotion of MSC osteogenesis. For cartilage regeneration, PRP of 2000 × 109 pl/L may be preferable to provide strong proliferation and preferable chondrogenesis but weak adipogenesis and osteogenesis. Furthermore, induction media, such as dexamethasone, can be used when the exorbitant PRP triggers undesirable results in osteogenesis-dependent therapies.
PRP contains multiple growth factors, including vascular endothelial growth factor, epidermal growth factor, bFGF, PDGF-AB, TGF-β1, and IGF-1. Previous reports demonstrated that these factors stimulate Col-II expression, increase matrix synthesis, and promote cell cycle progression to accelerate wound healing.53–55 To understand the underlying mechanisms by which PRP with different numbers of platelets exerts discrepant effects on MSC proliferation and differentiation, the stimulatory effects of these factors together on MSC differentiation and proliferation were investigated in our study. We found that certain levels of bFGF, IGF-1, PDGF-AB, and TGF-β1 in the 1500 × 109, 2000 × 109, and 3000 × 109 pl/L groups potently stimulated the osteogenic, chondrogenic, and adipogenic differentiation of MSCs, but these growth factors alone in the absence of platelets exerted milder effects than did PRPs, indicating that there are unknown factors in addition to the growth factors (bFGF, PDGF-AB, IGF1, and TGF-β1) in PRPs that are involved in promoting MSC-based tissue regeneration.
Although previous investigations were highly concerned with changes in the proliferation and differentiation of MSCs after PRP stimulation, little is known about the changes in the ultrastructure of these cells after treatment. In the current study, we found many visible changes in subcellular organelle structure of MSCs. More karyokinesis increased in a dose-dependent manner in PRP-treated MSCs, which partly accounts for the discrepant proliferative phenomena described above. In addition, the results of TEM clearly confirmed the greater increases in the number and size of organelles, such as dictyosomes, Golgi apparatus, and endoplasmic reticulum, in PRP-treated MSCs, indicating a marked increase in MSC metabolism, especially protein synthesis.
Recent studies showed that PRP modulates immune responses to promote tissue regeneration. van Buul et al. reported that PRP release diminished many of the inflammatory IL-1β-mediated effects on human osteoarthritic chondrocytes by inactivating the NF-κB pathway. 56 In addition, Osterman et al. found that treatment with PRP significantly decreased the expression of inflammatory genes in the cartilage and synovium from osteoarthritis patients. 57 However, it remains unknown whether PRP influences the immunological properties of MSCs. In the current study, we found that PRP treatment suppressed the secretion of the pro-inflammatory cytokines IL-1β, IL-6, and TNF-α, but promoted the production of the anti-inflammatory cytokines IL-10 by MSCs. Moreover, the anti-inflammatory effect is an important factor to be considered for MSC-based therapy. By secreting numerous cytokines, MSCs regulate the functions of most inflammatory cells and improve tissue regeneration. The findings reveal a novel role of PRP in MSC-based regeneration.
However, we are aware that the findings are based on an in vitro study, and the in vivo effects of concentrated PRPs on MSCs remain unknown. In addition, the source of MSCs affecting the repair of specific tissues probably emanates from the host MSCs rather than from grafted stem cells. Therefore, further in vivo investigations are needed in future studies to evaluate the effects of concentrated PRPs on tissue regeneration.
Conclusions
We report that PRPs with different platelet concentrations have different effects on human bone marrow MSC proliferation, as well as the osteogenic, adipogenic, and chondrogenic differentiation of MSCs. In addition, we determined the appropriate PRP concentrations for each lineage and MSC expansion. These findings may suggest promising clinical strategies for different diseases. However, further studies of clinical applications and in vivo and in vitro mechanistic investigations will be required to reinforce the findings of this study.
Footnotes
Acknowledgments
The authors thank Bin Wang, Qian Wang, Yimeng Yang, and Chunhui Liu (Chinese PLA General Hospital) for assistance and helpful discussions. The authors also thank Haibao Wang (Chinese PLA General Hospital) and Hongtao Yang (College of Engineering, Peking University) for preparing the whole blood samples. This study was supported by the Specialized Research Fund for Military Medicine Innovation (Grant No. 13CXZ030), the National Natural Science Foundation of China (Grant No. 81572159, 81871771 and 81371945), and the Beijing Natural Sciences Foundation (Grant No. 7182123).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.