
Abstract
A method that realizes allogeneic islet transplantation into a subcutaneous site under no immunosuppressive treatment has been desired for the treatment of type 1 diabetes. In this study, two donor/recipient combinations were used: F344 rat donors (RT-1lv1) and ACI rat recipients (RT-1a); and Wistar rat donors (closed colony) and Lewis rat recipients (RT-1l). An agarose rod, which carried basic fibroblast growth factor (bFGF; 50 μg/rod, agarose-bFGF rod), was implanted into each of the two dorsal subcutaneous sites of a streptozotocin-induced diabetic rat for 7 days, and the space was formed by removal of the rod. One thousand five hundred allogeneic islets were transplanted into each of the subcutaneous spaces. No immunosuppressive medication was given before or after islet transplantation. The immunological analyses of the granulomatous tissue formed by agarose-bFGF rod were carried out with flow cytometry and gene expression to get some information for mechanism of acceptance of allogeneic islets. Subcutaneous spaces surrounded with highly vascularized granulomatous tissue were formed by agarose-bFGF rod implantation for 7 days. All recipients demonstrated long-term normoglycemia (>100 days) in both donor/recipient combinations. The percentages of regulatory T (Treg) cells/CD4 cells in the granulomatous tissue increased by the implantation of agarose-bFGF rod and the percentages remained high after islet transplantation. In addition, expression levels of genes associated with chemotaxis and induction of Treg cells increased in the granulomatous tissue by the implantation of agarose-bFGF rod. The agarose-bFGF rod implantation is a useful method for preparation of an immunotolerant site for successful allogeneic islet transplantation without immunosuppressive medication, and Treg cells might play a pivotal role in graft acceptance.
Impact Statement
Although transplantation of islets of Langerhans has been accepted as a fundamental treatment for insulin-dependent diabetes mellitus (IDDM), several problems are still remaining in order that it becomes the standard medical treatment of IDDM patients. In this study, using diabetic rat models, a subcutaneous space surrounded with highly vascularized granulomatous tissue was formed by agarose-bFGF rod implantation. Allogeneic islets transplanted into the space could survive and release insulin for a long period under no immunosuppressive medication. In conjunction with sufficient supply of islets from iPS/ES cells, our method can make islet transplantation the standard medical treatment of IDDM patients.
Introduction
P
Immune reactions against grafts depend on the animal species, the phenotype (i.e., high- and low-responder phenotypes), and the organ or tissue specificity. 7 In this study, we examined whether long-term graft function could be realized in an allogeneic islet transplantation without immunosuppressive medications. We studied two donor/recipient combinations: F344 rat donors (RT-1lv1) and ACI rat recipients (RT-1a); and Wistar rat donors (closed colony) and Lewis rat recipients (RT-1l). Furthermore, we carried out immunological studies to gain some insight into the mechanisms underlying graft rejection and acceptance.
Materials and Methods
Materials
Chemicals and a hemoglobin assay kit were purchased from local suppliers. Supplementary Table S1 lists the antibodies used in this study. We purchased basic fibroblast growth factor (bFGF, FIBLAST Spray 500) from KAKEN PHARMACEUTICAL CO., LTD (Tokyo, Japan), collagenase from Nitta Gelatin (Osaka, Japan), SeaKem GTG agarose from Lonza (Basel, Switzerland), and FITC-tomato lectin from Vector Laboratories, Inc. (Burlingame, CA).
Islet transplantation and analyses
We used two combinations of rat strains. One combination was Fisher 344 (F344) rat donors and ACI rat recipients; the other combination was Wistar rat donors with Lewis rat recipients. These strains were fully MHC incompatible in the donor/recipient setting. Diabetes was induced in recipient rats with a single intraperitoneal injection of streptozotocin (STZ) (60 mg/kg body weight). All animal experiments were approved by the Kyoto University Animal Care Committee and performed according to the guidelines.
The bFGF was used to form a blood vessel-rich tissue subcutaneously. Characteristics required for the device carrying bFGF used to make the space for pancreatic islet transplantation subcutaneously are (1) bFGF can be slowly released subcutaneously during 1–2 days and (2) the device is not adhered to the tissue and thus can be easily removed.
Hydrogels of agarose can be easily removed without adhesion with tissue after implantation. In addition, bFGF can be supported in the agarose gel only by dropping a bFGF aqueous solution onto the freeze-dried agarose gel. In this study, agarose was used as a material for device fabrication.
We prepared freeze-dried agarose scaffolds (rod-shaped, diameter 4 mm, length 25 mm) as previously described.5,6 We evenly applied two different bFGF solutions (10 μg or 50 μg bFGF in 100 μL buffer) into agarose rods.
Following the STZ injection, rats received agarose-bFGF rods implanted into two dorsal subcutaneous tissue sites to induce vascularization. Rods were removed 7 days after implantation. For a semiquantitative assessment of the vascularization level, we determined the amount of hemoglobin in the tissue adjacent to the agarose rods, as previously described. 1
Islets were isolated from donor F344 rats and Wistar rats with the collagenase digestion method. 8 Briefly, to isolate islets from donor F344 and Wistar rats, the pancreas was inflated with an 8 mL injection of collagenase solution (0.5 mg/mL in HBSS). Then, the pancreas was recovered and digested. Islets were collected with a Ficoll/Conray density gradient and cultured for 2 days in RPMI 1640 with 10% FBS before transplantation. With this method, 300–500 islets were isolated from each F344 or Wistar rat. On removing the agarose rod, 1500 isolated islets were transplanted into each prevascularized dorsal pocket in STZ rats. None of the recipients received immunosuppressive drugs. Blood samples were collected from tail veins of recipient rats under nonfasting conditions. Blood glucose levels were determined with a glucose sensor (GLUCOCARD MyDIA; Arkray, Inc., Kyoto, Japan). The day of graft rejection was defined as the first day of 2 consecutive days that blood glucose measurements exceeded 250 mg/dL.
At 40 and 90 days after islet transplantation, blood was collected from recipients to determine plasma insulin levels. For controls, blood samples were also collected from normal (untreated) rats and STZ ACI rats without islet grafts. Plasma insulin concentrations were determined with ELISA kits for rat insulin (Shibayagi Co., Ltd., Gunma, Japan). Recipient rats also underwent an IPGTT between 50 and 55 days and between 100 and 105 days after islet transplantation. Briefly, after 16 h of fasting, rats received intraperitoneal injections of a glucose solution (1 g glucose/kg body weight). Blood samples were collected at 0, 15, 30, 60, 90, and 120 min after glucose loading.
On predetermined days after transplantation, subcutaneous tissues that contained the islet grafts were collected and fixed in 4% paraformaldehyde in PBS. The tissue sections (4 μm) were stained with hematoxylin and eosin (H&E) or with immunofluorescent probes for detecting insulin. 5
We also examined the effects of injecting donor rat splenocytes on accepted islet grafts. Briefly, STZ-ACI rats with long-term functional F344 islet grafts were subjected to intraperitoneal injections of 107 F344 splenocytes at 30 (n = 3), 117 (n = 4), or 118 (n = 1) days after islet transplantation. STZ-Lewis rats with long-term functional Wistar islet grafts were also subjected to intraperitoneal splenocyte injections. The Wistar rat is an outbred stock. At the time islets were isolated from donor rats, splenocytes were also isolated, frozen, marked for donor identification, and stored until use. Thus, 107 Wistar splenocytes, isolated from the same rat that donated the islets, were given to the corresponding recipient rat at 100 (n = 2) days after islet transplantation. Blood glucose levels were determined every other day after splenocyte injection. At 5 days after splenocyte injections, subcutaneous tissues, which included the islet grafts, were recovered for immunological examinations.
Immunological environment of the subcutaneous tissue prevascularized with agarose-bFGF rods
Fresh lymphocytes were obtained from rat subcutaneous tissues and separated, as described previously. 6 Briefly, subcutaneous tissue was excised and cut into small pieces. Then, lymphocytes were purified with a Percoll gradient centrifugation. Erythrocytes were eliminated with ACK buffer (Sigma Aldrich, St. Louis, MO). For intracellular staining, cells were fixed and permeabilized with Cytofix/Cytoperm™ and Perm Wash™ (BD Pharmingen), according to the manufacturer's instructions. Flow cytometry was performed with an FACS Canto II (BD Biosciences), and cell sorting using the SH800 (Sony Co.,Tokyo, Japan). The sorted cells showed a cell purity of >96%, and were used in subsequent experiments.
We determined the TGF-β1 concentrations in wound exudates of subcutaneous tissues implanted with agarose-bFGF rods with the Emax ImmunoAssay System (Promega Co., Madison, WI).
Gene expression analyses of prevascularized subcutaneous tissue
From subcutaneous tissues treated with agarose-bFGF rods (n = 3) or control bFGF-free agarose rods (n = 3), we extracted RNA with the RNeasy Plus Kit. We then prepared biotinylated antisense RNA with the Express Kit Assay (Affymetrix). Microarray analyses were performed by Kurabo Industries Ltd. (Tokyo, Japan) with Affymetrix GeneChip Rat Genome 230 2.0 arrays. From microarray data uploaded at the National Center for Biotechnology Information Gene Expression Omnibus (GEO), we identified specific gene groups that comprised genes associated with Treg cells, the GO term, cytokine, or the GO term, chemokine. We then compared the gene scores (t statistics) of each gene group associated with the GO terms, cytokine or chemokine (but not the group associated with Treg cells), against the total gene score as reference.
Statistical analysis
Between two groups, comparisons were performed with the Student's t-test. Comparisons among three or more groups were performed using one-way analysis of variance (ANOVA) with the post hoc Tukey–Kramer test. Time course for different groups was analyzed by two-way ANOVA and Student's t-test at the same time point. A log-rank test was used to compare graft survival between experimental and control groups. p-values <0.05 were considered statistically significant.
Results
Islet transplantation
The bFGF was applied to freeze-dried agarose rods (50 μg/rod) to create agarose-bFGF rods. We created rat diabetes models by injecting STZ into rats (STZ rats; Fig. 1A). Under normal conditions, subcutaneous tissues of STZ rats displayed low levels of vascularization (Supplementary Fig. S1). We implanted agarose-bFGF rods into STZ rats in each of the two dorsal subcutaneous sites. After 7 days, we observed the formation of highly vascularized granulomatous tissue adjacent to the agarose-bFGF rods (Fig. 1B and Supplementary Fig. S1). The hemoglobin content of these tissues increased to 5.46 ± 1.70 mg/g (Fig. 1C). Numerous endothelial cells, visualized with an intravenous injection of FITC-lectin, indicated that many blood vessels existed in the granulomatous tissue (Fig. 1D and Supplementary Fig. S1). In subsequent experiments, the agarose-bFGF rods were removed on day 7, and islets were transplanted into the resulting subcutaneous spaces, surrounded by highly vascularized, granulomatous tissue (bFGF spaces).
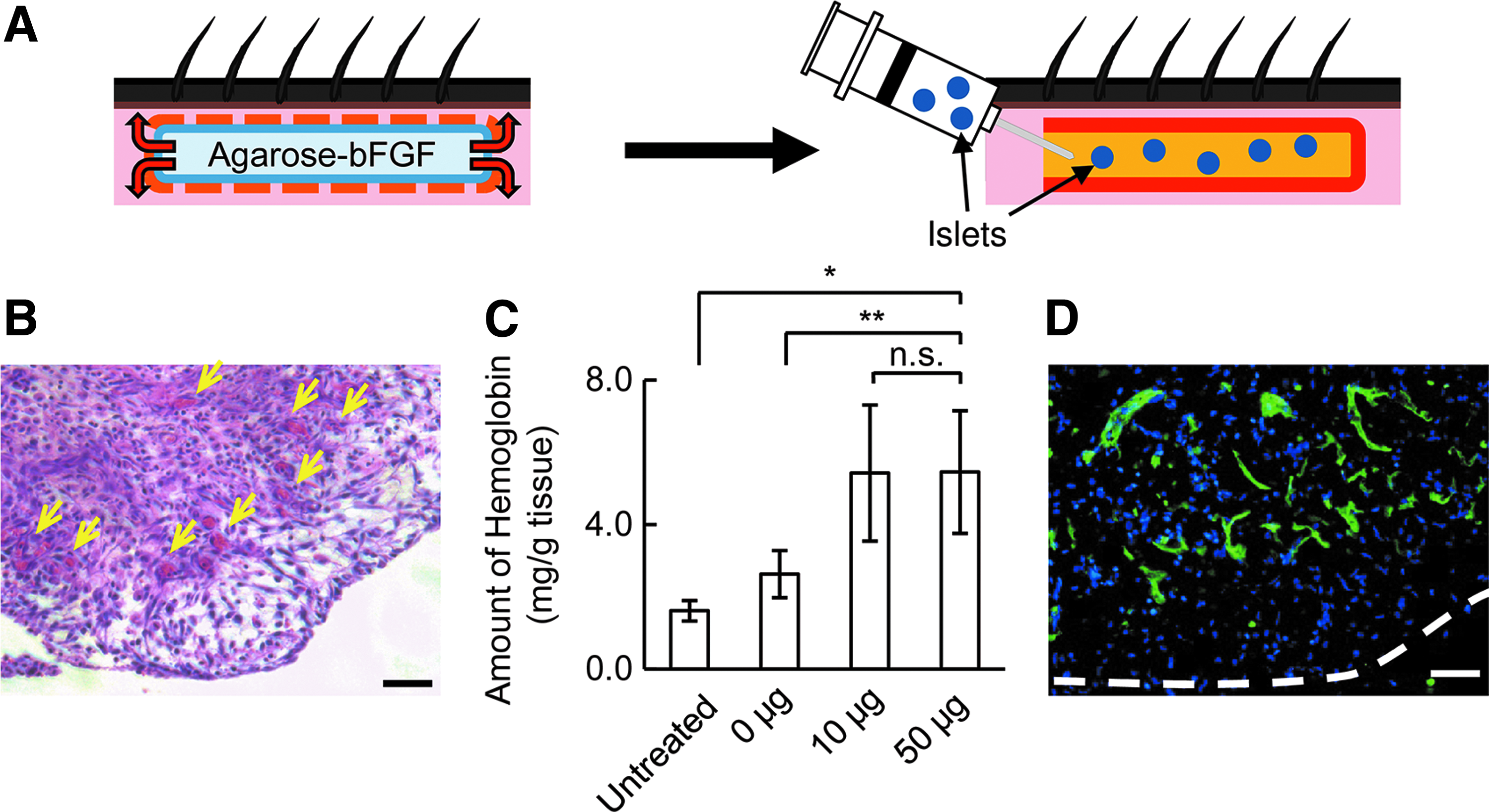
Preparation of a subcutaneous islet transplantation site.
Two models of donor/recipient combinations were used for transplantations. In one model, islets from donor F344 rats (RT-1lv1) were transplanted into recipient ACI rats (RT-1a); in the other model, islets from donor Wistar rats (closed colony) were transplanted into recipient Lewis rats (RT-1l). All recipient rats had STZ-induced diabetes (STZ-rats). After the agarose-bFGF rod treatments, a total of 1500 allogeneic islets were transplanted into each bFGF space for a total of 3000 islets per recipient rat. No immunosuppressive medications were given to recipient STZ rats, before or after transplantation. Blood glucose levels of the eight STZ-ACI rats and five STZ-Lewis rats are shown in Figure 2A and B, respectively. All recipients achieved normoglycemia more than 100 days after transplantation. The full normalization of blood glucose levels took longer in STZ-Lewis rats than in STZ-ACI rats. This difference might have been due to the low quality of islets isolated from Wistar rats. In control experiments, islets were transplanted into subcutaneous tissues that were either untreated or treated with a bFGF-free agarose rod. 6 No recipients exhibited normoglycemia (Fig. 2C, D; also our previous findings 6 ). Figure 2E and F shows graft survival during 100 days after transplantation.
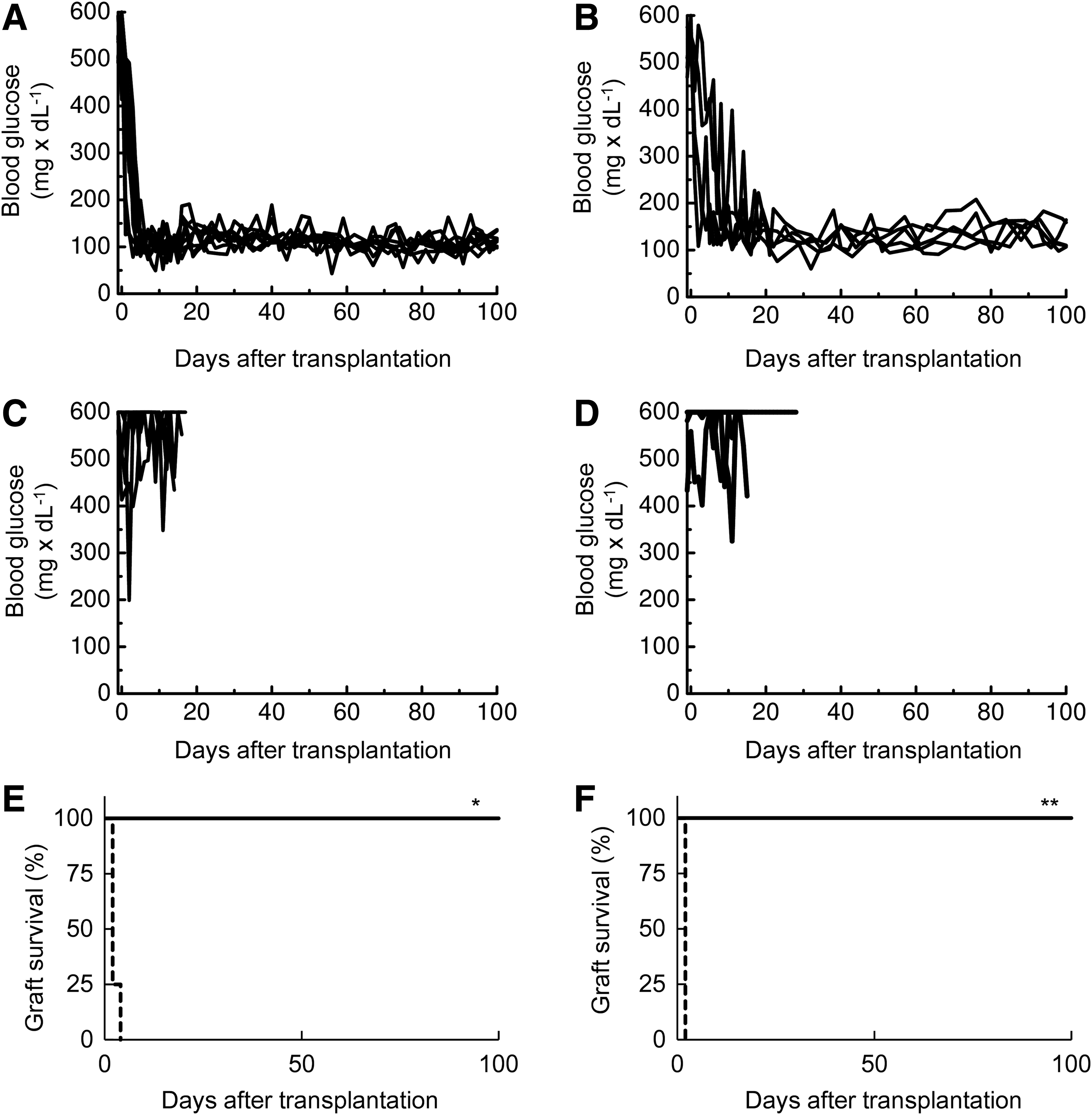
Normalization of blood glucose levels after islet transplantation into subcutaneous bFGF spaces. Rats received no immunosuppressive drugs, before or after islet transplantation. Blood glucose levels were measured after transplanting 1500 islets subcutaneously.
Islet graft function was characterized in ACI recipients by determining plasma insulin levels and performing intraperitoneal glucose tolerance tests (IPGTTs; Fig. 3A–C). The plasma insulin levels of the ACI recipients were higher than those of untreated STZ rats, at 40 and 90 days after transplantation (Fig. 3A). The levels in ACI recipients were nearly same as those observed in normal rats. Large differences were observed in the areas under the blood glucose curves (AUCs) of IPGTTs between untreated STZ rats and recipient STZ rats implanted with F344 islets at 50–55 days and 100–105 days after transplantation (Fig. 3B, C).
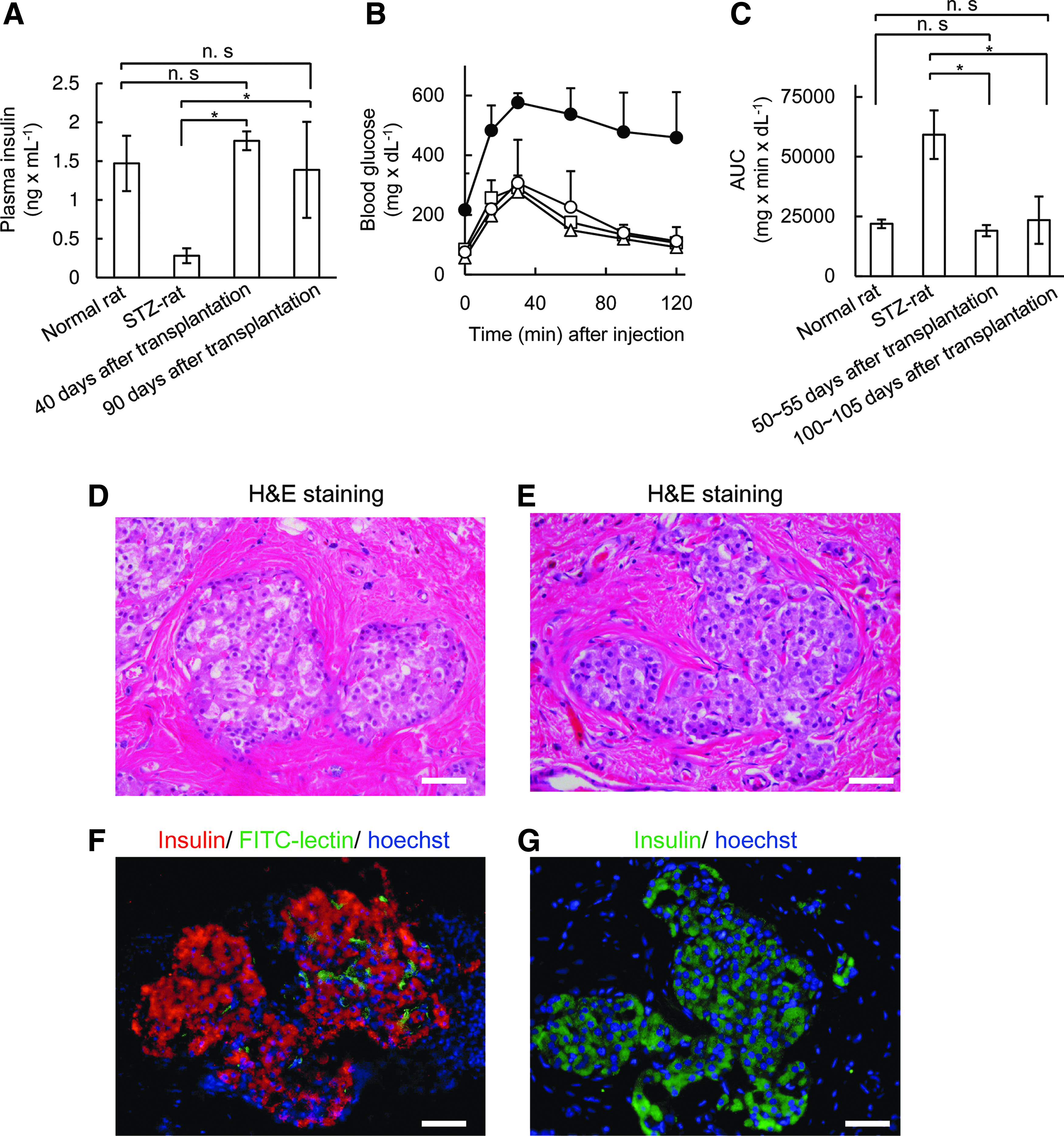
Functions and morphologies of islet grafts.
The islet grafts in the left dorsal site of ACI recipients and in both sites of Lewis recipients were retrieved at 159 and 100 days after transplantation, respectively. The blood glucose levels of the Lewis recipients immediately increased after removing the islet grafts on both sides. This result indicated that blood glucose levels were maintained at normal levels by the islet graft. In contrast, the ACI recipients maintained normal blood glucose levels after the islet grafts were removed from one side. This phenomenon indicated that 1500 islets were sufficient to control blood glucose levels. The retrieved islet grafts were processed for histochemical examinations. The islet grafts were clearly visible in H&E-stained slices (Fig. 3D, E). Most islets were incorporated into the connective tissue. Few lymphocytes were found in and around the islet grafts. Many insulin-positive cells were observed in immunofluorescently stained slices (Fig. 3F, G). The vessels in islet grafts that were connected to host vessels of the recipients were visualized with an intravenous infusion of an FITC-lectin solution (Fig. 3F).
Immune characteristics of vascularized granulomatous tissue, before and after islet transplantation
Immune cells that infiltrated the granulomatous tissue before islet transplantation were isolated and analyzed with fluorescence-activated cell sorting (FACS; Fig. 4). The percentage of Foxp3+ cells among all CD4+ T cells (Treg%) increased in the presence of agarose-bFGF rods, reaching 25% at 2 days after implantation. The Treg% was maintained at levels >24% from 7 to 14 days after agarose-bFGF rod implantation (Fig. 4A). In contrast, the Treg% was <15%, when granulomatous tissue was implanted with bFGF-free agarose rods. Moreover, in all STZ rats that received agarose rods, with or without bFGF, less than 10% of Treg cells were detected in the draining lymph nodes, axial lymph nodes, and spleen on any day (Fig. 4B and Supplementary Fig. S2). When the Treg% in lymph nodes was compared with the Treg% in spleen, no difference was observed. These facts suggest that the agarose-bFGF rods evoked regional, but not systemic, modifications in the immunological environment. The CD8+ T/CD4+ T ratio was also determined. After agarose rods with/without bFGF were implanted, the CD8+ T/CD4+ T ratios of both groups increased from ∼35% to 50%–70% during the observation period (Fig. 4C). However, the CD8+ T/CD4+ T ratios slightly decreased, from 37% to 28%, in the draining lymph nodes (Fig. 4D). Implantation of agarose-bFGF rods did not produce a change in the CD8+ T/CD4+ T ratios in the axial lymph nodes or spleens of STZ rats (Supplementary Fig. S2). TGF-β1, which is reported to play a role in inducing Treg cells, 9 was maintained at higher concentrations in the tissue fluids of spaces implanted with agarose-bFGF rods than in spaces implanted with bFGF-free agarose rods (Fig. 4E).
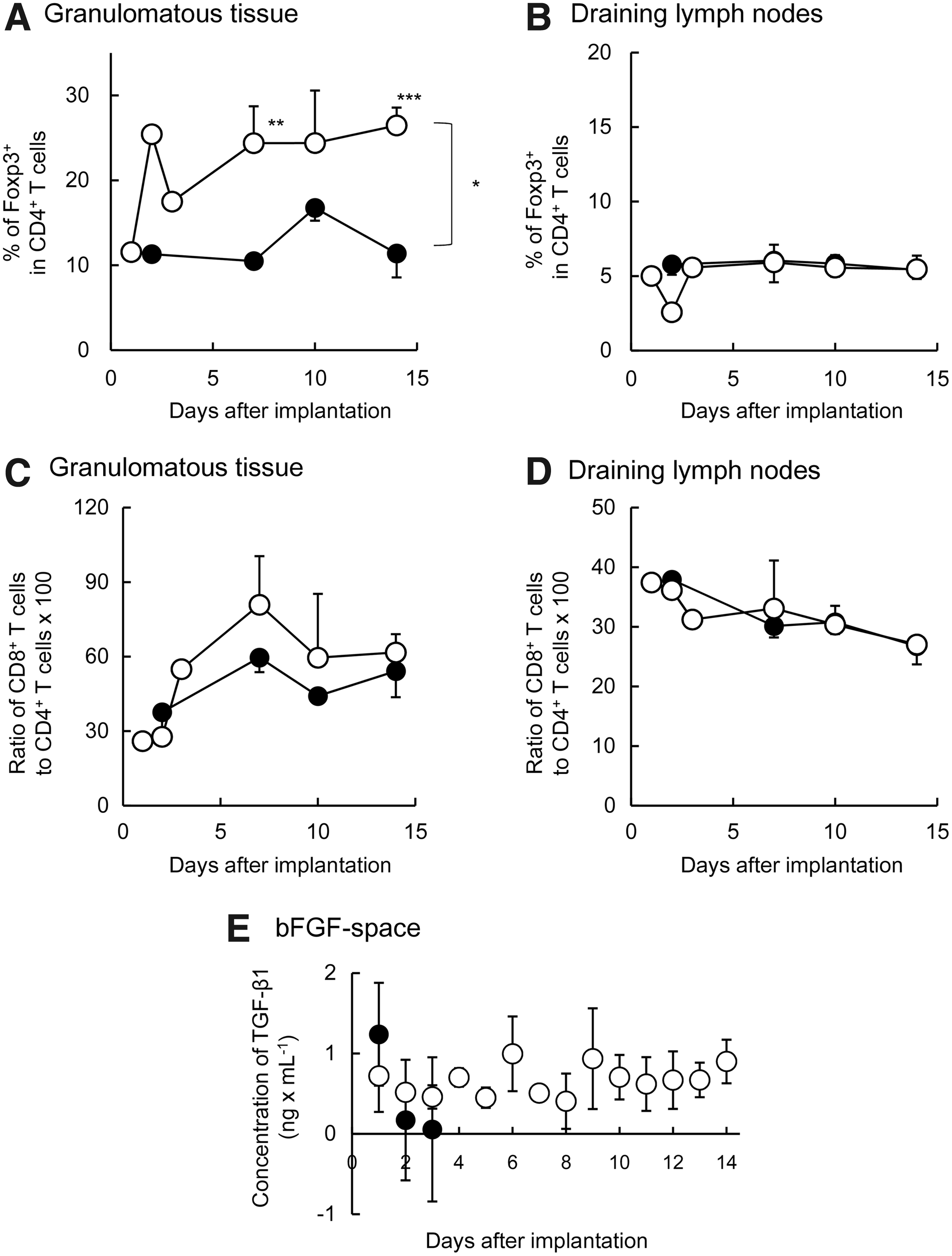
Recruitment of immune cells in granulomatous tissues formed at subcutaneous sites and in draining lymph nodes. Cells were measured for 14 days following implantation of agarose rods with or without bFGF.
The immunological characteristics of the islet graft environment played a pivotal role in the long-term survival of the graft and in its success in glycemic control. Immune cells that were in the graft and surrounding connective tissues were analyzed at 5 and 30 days after islet transplantation. Around the islet grafts, Treg cell levels >20% were maintained for 30 days after transplantation (Fig. 5A). In contrast, Treg cell levels were <10% in draining lymph nodes, in axillary lymph nodes, and in the spleen (Fig. 5B and Supplementary Fig. S3). Around the islet grafts, CD8+ T/CD4+ T ratios rose to slightly higher levels than observed pretransplantation, but later, these ratios decreased to preoperative levels (Fig. 5C). CD8+ T/CD4+ T ratios were <50% in draining lymph nodes, in axillary lymph nodes, and in the spleen, before and after islet transplantation (Fig. 5D and Supplementary Fig. S3).
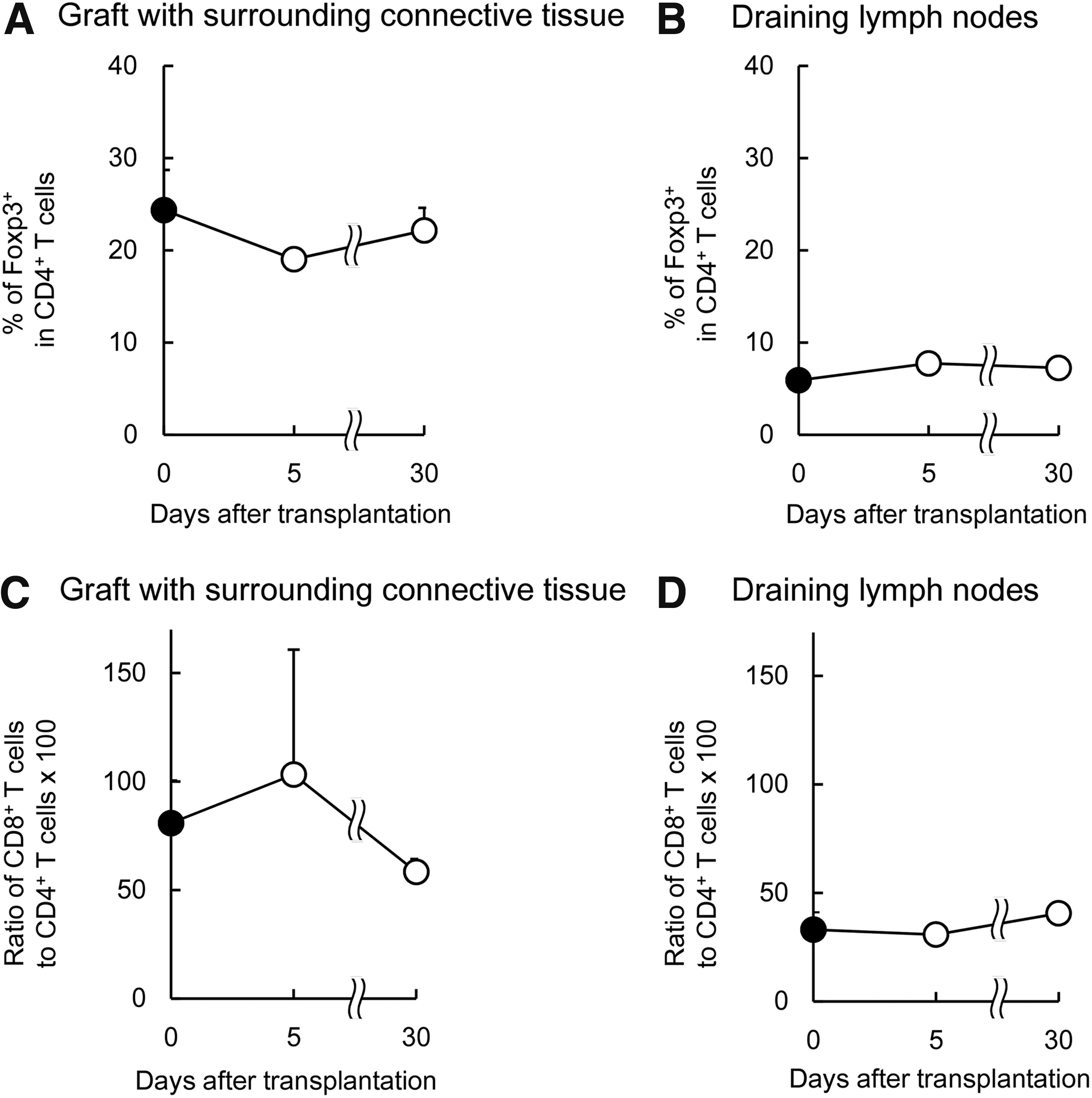
Recruitment of immune cells in STZ-ACI recipient rats measured at 5 and 30 days after F344 islet transplantation.
Gene expression in granulomatous tissue
We also analyzed gene expression in granulomatous tissue formed after implanting agarose rods, with or without bFGF (Fig. 6). First we examined the gene group defined in gene ontology (GO) terms as “chemokines and cytokines.” The expression of those genes, excluding the Treg cell-associated genes, was significantly reduced in the bFGF group compared with the bFGF-free group (Fig. 6A). In contrast, expression levels of Treg cell-associated chemokines, cxcl12 and the tgf signaling-associated gene (grem1), increased significantly in the bFGF group compared with the bFGF-free group (Fig. 6B, C). The expression levels of grem1 might play an important role in recruiting or inducing Treg cells. 10 These data suggested that the agarose-bFGF rod implantation stimulated the expression of some chemokines, cytokines, and receptors associated with chemotaxis and induction of Treg cells. Gene expression analyses also suggested that Treg cells contributed to the long-term acceptance of allogeneic islet grafts.
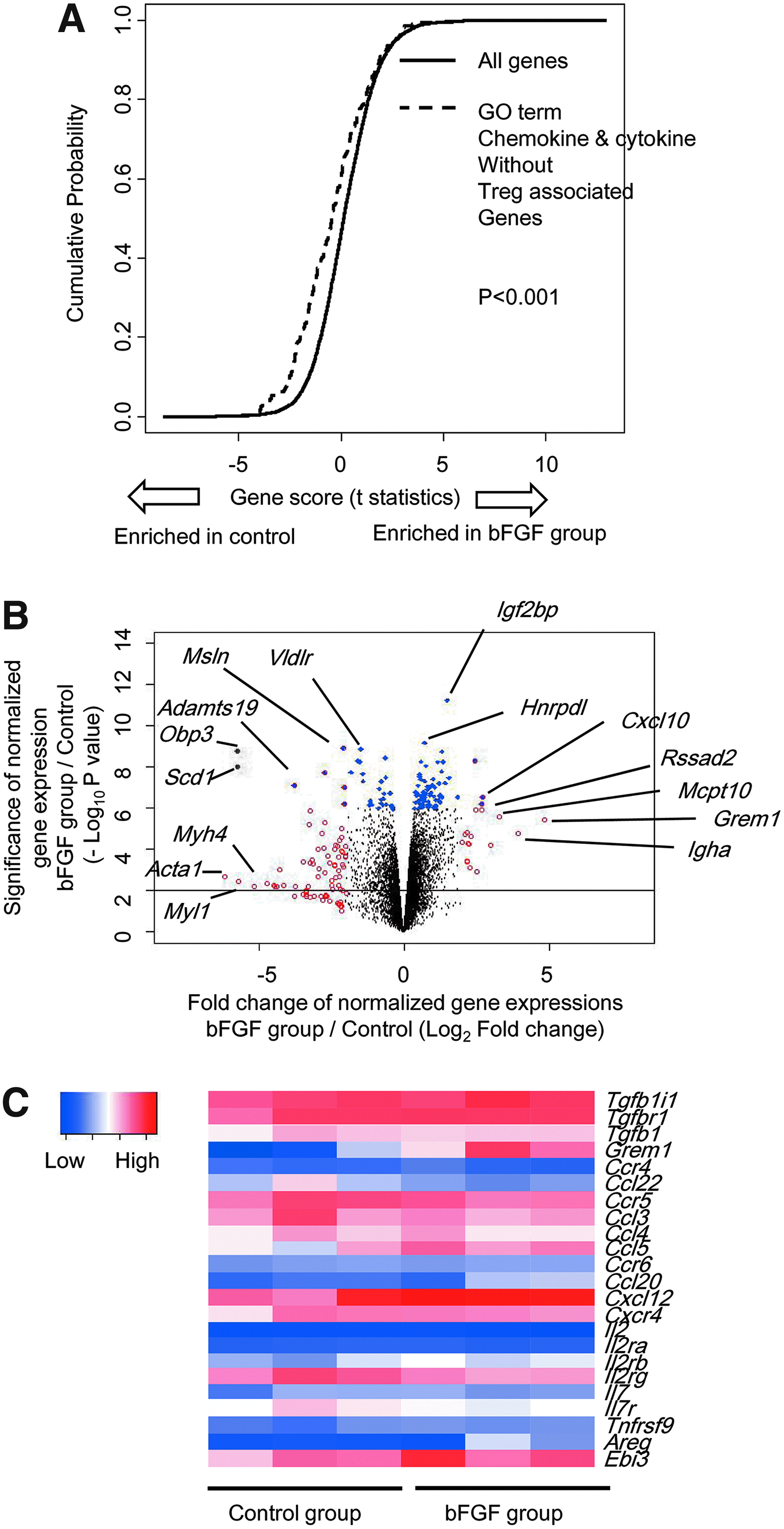
Gene expression changes in granulomatous tissue formed by implanting agarose rods with or without bFGF. Whole gene expression was analyzed in granulomatous tissue newly formed at 7 days after STZ-ACI rats were implanted with rods with or without (control) bFGF (n = 3, each group).
Effect of donor splenocyte injections on graft survival
The STZ-ACI rats with stable F344 islet grafts received an intraperitoneal injection of F344 rat splenocytes at 30, 117, and 118 days after transplantation. At 106 days post-transplantation, the left islet graft of each ACI recipient rat was retrieved for histological examination. Blood glucose was maintained at normal levels by the remaining right islet graft. Blood glucose levels tended to increase a few days after splenocyte injections (Fig. 7A). Two STZ-Lewis rats with Wistar islet grafts also received splenocytes from the donor Wistar rats at 100 days (Fig. 7B). Again, blood glucose levels tended to increase. A similar increase in blood glucose response was observed when a recipient underwent surgical removal of the grafts on both sides (dashed line). Among STZ-ACI rats that received splenocytes 30 days after transplantation, we analyzed the immunological characteristics of the connective tissue in the graft environment, before and after the splenocyte injection. The Treg% levels in and around islet grafts were 23.4% ± 5.2% before donor antigen injection, and they slightly decreased to 20.7% ± 3.1% at 5 days after the injection of splenocytes (Fig. 7C). In contrast, the ratio of CD8+ to CD4+ T cells increased from 59.6% ± 8.2% to 131.8% ± 41.8% at 5 days after the injection (Fig. 7D).

Rejection of established subcutaneous allogeneic islet grafts caused by infusing splenocytes from islet donors into the peritoneal cavities of recipient rats.
Discussion
Various methods have been investigated for transplanting islets without immunosuppressive medications, including the following: (1) transplanting into immune privileged (or tolerant) sites; (2) creating a bioartificial pancreas; (3) reducing islet immunogenicity; and (4) transplanting islets plus suppressive immune cells. Although these methods looked promising in small animals, each has proven difficult to apply in humans. For example, immune privileged sites, such as the cerebral ventricle, the anterior eye chamber, and the testicle,11–13 were difficult to apply clinically because they were too small for islet transplantation. Alternatively, a bioartificial pancreas or islets encapsulated in a semipermeable membrane 14 were limited in clinical application in humans, due to the large size or volume of the devices. Other methods investigated involved modulating islet immunogenicity, such as UV light irradiation or culturing at low temperatures.15–17 To our knowledge, none of those methods was successful in clinical applications. Other groups and ours have reported promising results by transplanting coaggregates that comprised islets and immunosuppressive cells, such as Treg cells,18,19 Sertoli cells,20,21 and mesenchymal stem cells. 22 However, it is difficult to prepare coaggregates of human islet cells and Treg cells in large quantities for use in human patients.
Our approach to protecting islet grafts from the host immune system was to transplant islets into a subcutaneous site formed by implanting agarose rods applied with bFGF. We found that the agarose-bFGF rod evoked regional, but not systemic, immunological reactions. This feature was suggested by the increase observed in the percentage of Foxp3+ (Treg) cells among CD4+ T cells in granulomatous tissue, but not in the lymph nodes or spleen. Finally, the allogeneic islet grafts survived in subcutaneous bFGF spaces for long periods, in the absence of immunosuppressive medications.
Presently, clinical islet transplantation is performed with the intraportal route. It is reported that nearly 60% of transplanted islets are lost by instant blood-mediated inflammatory reaction (IBMIR). 23 In subcutaneous transplantation, transplanted islets are not directly contacted by blood, so it is expected that IBMIR does not occur, and it is another advantage of subcutaneous transplantation. 4 It is an important remaining study to evaluate the number of islets before and after transplantation to claim the additional utility of subcutaneous transplantation.
As shown in Figure 7, islet grafts that had functioned for long periods of time were rejected when donor lymphocytes were injected into the peritoneal cavities of recipients. This fact was interesting from an immunological standpoint, and also from a safety standpoint, which is important for clinical islet transplantations. Islet shortage is considered an obstacle that must be overcome, but this problem will likely be resolved in upcoming years. Several groups have successfully regenerated quasi-islets from human pluripotent stem cells (ES/iPS cells). They reported that transplantation of quasi-islets in a murine diabetes model could normalize hyperglycemia.24,25 However, it is important to ensure the safety of transplantation. The major safety concerns with those methods are teratoma formation by residual ES/iPS cells and tumor formation by transformed cells, which might arise in the process of differentiation. 26 When a teratoma/tumor forms in a subcutaneous site, it can be readily removed surgically, due to easy access in subcutaneous tissue. Moreover, residual malignant cells can be killed by administering antigen prepared from the original ES/iPS cells. Thus, cell therapy safety can be increased by using functional cells derived from ES/iPS cells.
In the present study, we analyzed T cells that had infiltrated into the granulomatous tissue to gain some information about the immunological environment. The populations of Treg cells were increased before islet transplantation (Fig. 4A). Our results suggested that Treg cells played a pivotal role in graft acceptance. However, in addition to Treg cells, various other cells, including regulatory B cells27,28 and M2 macrophages,29,30 have been known to suppress immune reactions. Notably, immunological reactions to grafts highly depend on the response phenotype, the type of organ or tissue, and the animal species involved. Future studies should conduct islet transplantations in larger animal models, such as pigs and macaque monkeys, and perform more detailed immunological analyses in mouse models.
Conclusion
We can prepare immunotolerant pockets surrounded with blood vessel-rich tissue under skin of diabetic rats. Allogeneic islets transplanted into the pockets can survive and normalize blood glucose levels more than 100 days even under no immune suppressive treatment. We believe that the minimally invasive procedure and the lack of need for immunosuppressive drugs motivate researchers of islet transplantation and diabetologists to begin research to provide our method to diabetic patients.
Footnotes
Acknowledgments
This study was supported, in part, by a Grant-in-Aid for Scientific Research on Innovative Areas (No. 23107008), by Scientific Research (A) (No. 24240078), by Challenging Exploratory Research (No. 26670577), by Scientific Research (A) (No. 17H01579), by JSPS Research Fellow (No. 16J06573) from Japan Society for the Promotion of Science, by the Cooperative Research Program of Institute for Frontier Medical Sciences, Kyoto University, Japan, and by the Center of Innovation Program from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Disclosure Statement
The authors declare no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.