
Abstract
The regeneration of functional osteochondral tissue is mired by the difficulty of recapitulating the zone-dependent differences in tissue composition and constituent cell phenotype. The objective of the current work is to study the ability of multidirectional zonal scaffolds to promote the differentiation of murine bone marrow stromal cells (BMSCs) and chondrocytes along the osteogenic and chondrogenic lineages, respectively, by analyzing dual or triple transgenic fluorescent reporter expression in vitro and in vivo. BMSCs containing fluorescent reporters for bone sialoprotein (BSP) and dentin matrix protein (DMP-1) were seeded on zonal scaffolds in vitro, where successful differentiation was determined from a continued progression of BSP to DMP1-exclusive fluorescence and was corroborated by gene expression and histological analyses. Zonal chondrogenesis was then separately studied using articular chondrocytes containing fluorescent reporters for types 1, 2, and 10 collagens. The majority of fluorescent cells (65.1%) displayed exclusive fluorescence of the type 2 collagen reporter, where a significant portion (27.7%) cofluoresced type 2 and type 10 collagen reporters, indicating a progression to hypertrophy in vitro. Zonal gene expression, histological, and immunohistochemical analyses revealed prehypertrophic chondrocytes embedded in zonally-organized matrix. Upon ectopic implantation, the prehypertrophic chondrocytes formed mineralized tissue through an endochondral ossification pathway. Fluorescent multireporter cells accurately monitored the differentiation behavior of BMSCs and articular chondrocytes on zonal scaffolds and revealed subtle differences in tissue and chondrocyte phenotype.
Impact Statement
This study represents significant advancement in the use of biomimetic scaffolds to direct zonal osteochondral tissue formation. We describe the use of a novel fluorescent reporter system that enables the real-time evaluation of cellular differentiation in a nondestructive manner. In this study, we use this tool to confirm the osteogenic and chondrogenic capabilities of our scaffold alongside control scaffolds, and use cryohistological methods to probe zone-specific differences in cell and tissue quality. We believe this approach can be widely adopted by others for a variety of biomaterial and cell systems in the development of tissue engineered therapeutics.
Introduction
A
A recent notable advancement in biomaterial design is the production of collagen-based multidirectional zonal (MZ) scaffolds that mimic both the fiber orientations and chemical compositions of the superficial, transition, calcified cartilage, and osseous zones of osteochondral tissue in a monolithic unit, as reported by our laboratory. 12 A 12 facile lyophilization bonding approach is used to join a unidirectionally-aligned superficial zone-mimicking scaffold (comprised of type 1 collagen and hyaluronic acid) with a lamellar osseous zone scaffold (comprised of type 1 collagen mineralized with carbonated hydroxyapatite) using a medial transition zone suspension (comprised of type 1 collagen and hyaluronic acid) (Fig. 1A). Resulting matrices contain a 200 μm-thick laterally-organized superficial zone, an isotropic transition zone, a calcified cartilage zone consisting of osseous zone lamellae interdigitating the overlying transition zone, and a vertically-oriented osseous zone consisting of lamellae of type 1 collagen mineralized with carbonated hydroxyapatite (Fig. 1B).
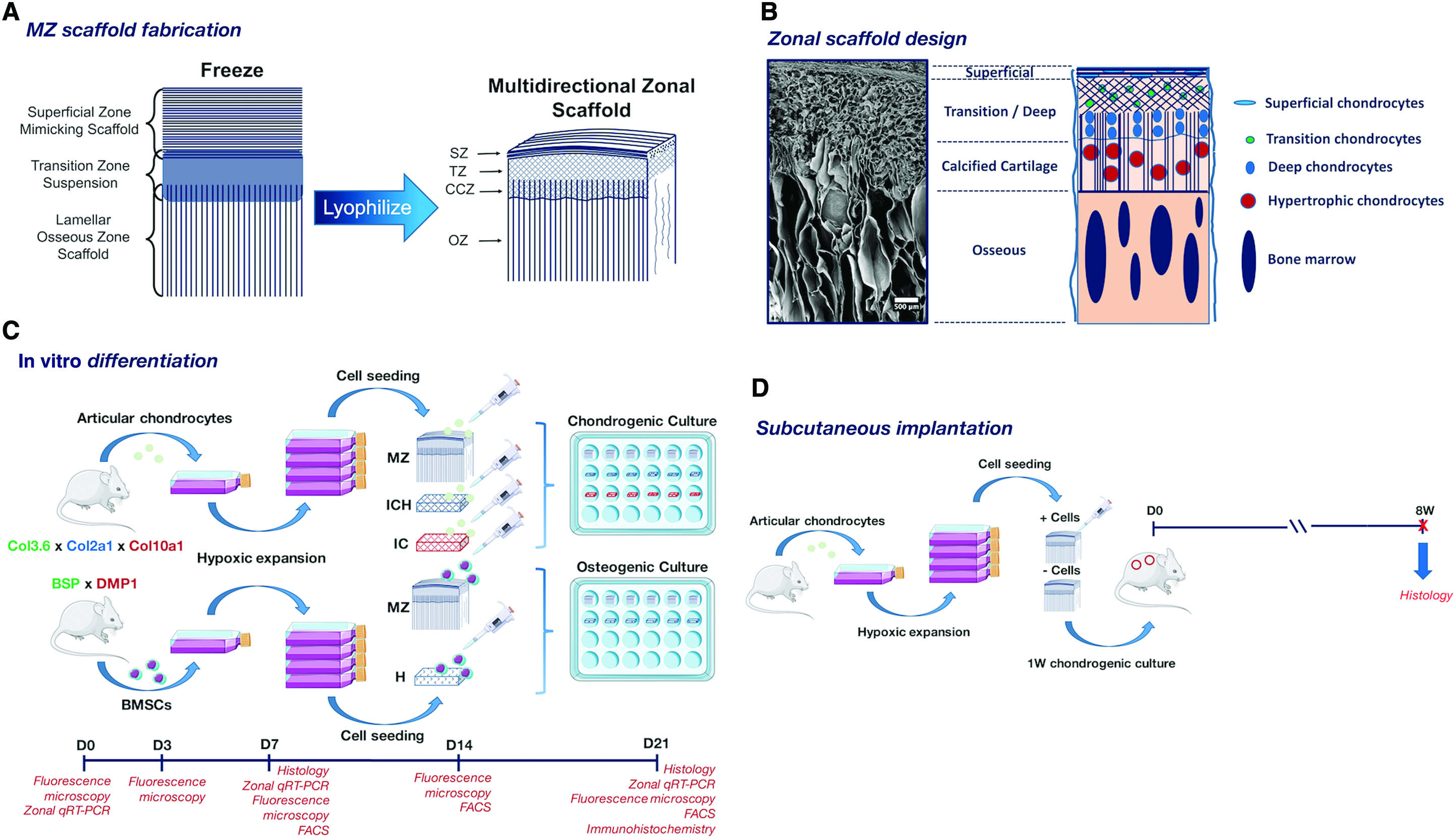
MZ scaffolds may provide the necessary chondrogenic (hyaluronic acid) and osteogenic (carbonated hydroxyapatite) chemical cues to promote the differentiation of chondrocytes and bone marrow stromal cells (BMSCs) along chondrogenic and osteogenic lineages, respectively. Furthermore, the zone-specific matrix alignment may guide the formation of new tissue that is aligned with the zonal pore structure of the material. Generated tissue may then recapitulate the native zonal properties of osteochondral tissue. The reader is referred to our previous study for a detailed description of the fabrication procedure and material properties. 12
To evaluate the ability of such biomaterials to support the differentiation of a given cell line, destructive characterization modes are often used. However, a need exists for a means to nondestructively monitor cell differentiation in real time. To address this need, we employ a novel fluorescent reporter gene approach to study cell differentiation on our MZ scaffolds. The Rowe laboratory has developed various lines of transgenic mice harboring different combinations of promoter-driven green fluorescent protein (GFP) reporters that activate at defined levels of cell differentiation. This system has been combined with a cryohistological protocol that preserves GFP expression and has been used to study various stages of osteoblastic differentiation in vivo,13,14 fracture repair, 15 changes in chondrocyte cell populations in response to load,16,17 and new progenitor cell populations through lineage tracing experiments.18,19 To date, however, this approach has seldom been applied to study cell differentiation on biomaterials in vitro.
The aim of this study is to investigate the ability of MZ scaffolds to support the differentiation of fluorescent reporter murine chondrocytes and BMSCs along chondrogenic and osteogenic lineages. For osteogenic differentiation, BMSCs derived from dual transgenic mice harboring reporters for bone sialoprotein (BSP-GFPtopaz) and dentin matrix protein (DMP1-RFPmCherry) will be used, where the progressive expression from BSP to DMP-1 (indicating preosteoblast to mature osteoblast/osteocyte maturation) will be analyzed. For chondrogenesis, articular chondrocytes from triple-transgenic mice (Col3.6-GFPtopaz, Col2a1-GFPcyan, Col10a1-RFPmCherry) harboring reporters for different chondrogenic fates (Col3.6–fibrochondrocytes, Col2a1–articular chondrocytes, Col10a1–hypertrophic chondrocytes) will be used. Furthermore, gene expression and cryohistological analyses will be used to investigate the zone-specific maturation of transgenic chondrocytes.
Methods
Scaffold fabrication
Zonal multidirectional scaffolds were fabricated by a lyophilization bonding procedure as previously published. 12 Superficial zone-mimicking and lamellar osseous zone scaffolds were first prepared. The former was fabricated by the unidirectional solidification of an aqueous suspension containing acid-soluble rat tail collagen (derived in-house according to Rajan et al) 20 and hyaluronic acid (hyaluronic acid sodium salt from Streptococcus equi; Sigma) at a 9:1 wt/wt ratio of collagen:hyaluronic acid to total 2% (wt/vol). The suspension was homogenized with a rotor–stator homogenizer (Omni TH, Omni-Inc) over ice, degassed by centrifugation, unidirectionally frozen from 4°C to −80°C at 4°C/min, and lyophilized at −25°C and 0.03 mBar for ≥3 days.
Lamellar osseous zone scaffolds were fabricated by coprecipitation of collagen and hydroxyapatite at neutral pH in a modified simulated body fluid solution containing 92 mM NaCl, 3.1 mM K2HPO4 * 3H2O, 3.3 mM MgCl2, 50 mM HEPES, 8 mM CaCl2, 17.5 mM NaHCO3, and 2.25 mg/mL acid-soluble rat tail collagen.21–23 The resulting gel was subject to 15 min of unconfined self-compression, radically frozen at −25°C overnight, and lyophilized under the previous conditions. Lamellar osseous zone scaffolds were then crosslinked with 1% (wt/vol) 1-ethyl-3–(3-dimethylaminopropyl)carbodiimide hydrochloride (EDC) and 0.25% (wt/vol) N-hydroxysuccinimide (NHS) in 2-(N-morpholino)ethanesulfonic acid buffer at a pH of 5 for 4 h at room temperature, rinsed with 5% (wt/vol) glycine overnight, rinsed thrice with sterile deionized water, and lyophilized under the previous conditions.
Zonal multidirectional scaffolds were made by bonding underlying superficial zone mimicking scaffolds with overlying lamellar osseous zone scaffolds using 75 μL of an intermediate aqueous suspension of collagen–hyaluronic acid at an 8:2 ratio of collagen:hyaluronic acid to total 1.5% (wt/vol) (Fig. 1A). These constructs were then frozen at −25°C, lyophilized under previous conditions, and crosslinked with 1% (wt/vol) EDC containing 0.25% NHS in 95% ethanol for 4 hr at room temperature, rinsed with 5% glycine overnight, rinsed thrice with sterile deionized water, and lyophilized under the previous conditions.
Isotropic collagen–hyaluronic acid (ICH) and isotropic collagen (IC) scaffolds were fabricated to serve as scaffold controls for anisotropy (ICH), and both anisotropy and hyaluronic acid (IC). Aqueous suspensions of collagen–hyaluronic acid and collagen were prepared to total 1.5% (wt/vol). ICH scaffolds contained collagen:hyaluronic acid at an 8:2 ratio. Prepared suspensions were first homogenized over ice, degassed, isotropically frozen at −25°C, and lyophilized for ≥3 days. Scaffolds were then crosslinked with 1% EDC:NHS in 95% ethanol as before, then lyophilized for ≥3 days.
Transgenic primary articular chondrocyte and BMSC culture
Primary BMSCs were obtained by harvesting bone marrow from 3- to 4-week-old dual transgenic mice containing BSP-GFPtopaz and DMP1-RFPmCherry fluorescent reporter genes (Fig. 1C). Hind legs were harvested and tibias and femurs were collected in cold, sterile 1 × PBS. Epiphyseal bone was cut and bone marrow was flushed with 10 mL sterile expansion media using a 25 5/8-gauge syringe needle. Collected marrow was then passed through an 18 1/2–gauge needle to break up tissue pieces, plated at 10 × 10 6 cells/well in a six-well plate, and expanded in hypoxic incubation (37°C, 5% CO2, 5% O2) to accelerate growth of the progenitor cells. Media were replenished after 4 days of initial expansion to allow for hematopoietic cells to promote initial BMSC colony formation 22 and to remove nonadherent nonstromal cells. Expansion media were replenished every other day until confluency at passage 1.
Primary articular chondrocytes were harvested from 3- to 4-week-old triple transgenic mice containing Col3.6-GFPtopaz, Col2a1-GFPcyan, and Col10a1-RFPmCherry fluorescent reporter genes (Fig. 1C). Following CO2 asphyxiation, hind legs were collected and placed in cold, sterile 1 × PBS. Femoral condylar cartilage was isolated with a scalpel and placed in a digestion buffer containing 3 mg/mL collagenase D (Roche), 2 mg/mL dispase (GIBCO), and 1% (vol/vol) penicillin–streptomycin (GIBCO) in sterile Hank's Balanced Salt Solution (HBSS, GIBCO). Tissue was digested for three 40 min cycles at 37°C, with supernatant collected into sterile expansion media (high-glucose DMEM, GIBCO, containing 10% fetal bovine serum (FBS; Hyclone) and 1% penicillin–streptomycin) over ice. Tissue supernatant was then sterile-filtered over a 70 μm nylon membrane and plated at 65–80 × 10 6 viable cells per well in a six-well tissue culture-treated plate. Primary chondrocytes were expanded to confluency at passage 1 in hypoxic incubation with expansion media replacement every other day.
Scaffold seeding and in vitro culture
MZ scaffolds were cut to approximately 6 × 3 × 3 mm (0.054 cm 3 ) and Healos (a widely used commercial apatite/collagen scaffold to serve as a positive control for osteogenesis consisting of 20 wt% HA and 80 wt% type 1 collagen) (DePuy Spine), IC, and ICH scaffolds were cut to approximately 4 × 2 × 2 mm (0.016 cm 3 ) with sterile razor blades. Care was taken to remove closed-pore surfaces (artifacts of the lyophilization process). All scaffolds were disinfected with 70% ethanol for 1 h, then rinsed with sterile 1 × PBS thrice. Scaffolds were then soaked overnight in sterile DMEM at room temperature to promote protein adsorption. The next day, soaked scaffolds were briefly dried over sterile cloth to remove entrapped media, transferred to ultra-low attachment 24-well plates, and seeded with either 4.5 × 10 6 cells/cm 3 articular chondrocytes or 9 × 10 6 cells/cm 3 BMSCs. Cells were allowed to attach in hypoxic incubation for 2 h, with small media additions as needed to keep samples hydrated.
For articular chondrocytes, a full volume (1.5 mL) of defined chondrogenic media (high-glucose DMEM supplemented with 1% ITS [GIBCO], 500 μg/mL BSA [Sigma], 4.7 μg/mL linoleic acid [Sigma], 1 × 10–4 M ascorbic acid [Sigma], 4 × 10–4 M proline [Sigma], 1% penicillin–streptomycin, 10 ng/mL hTGFβ1 [R&D Systems, Inc.], and 10–7 M dexamethasone [Sigma]) was gently added. For BMSCs, 1.5 mL of defined osteogenic media (αMEM [GIBCO] supplemented with 10% FBS, 1% penicillin–streptomycin, 8 mM β-glycerophosphate [Sigma], and 50 μg/mL ascorbic acid) was added. For both groups, cell-seeded constructs were cultured for 21 days in either hypoxia (chondrocytes) or normoxia (20% O2, 5% CO2, 37°C, BMSCs) with media replacement every other day.
A positive control for osteogenesis was created by culturing BMSCs in six-well plates in DMEM, αMEM, and defined osteogenic media. Verification of osteogenesis was performed at day 21 by fluorescent imaging of reporter expression and von Kossa staining. A positive control for chondrogenesis was made by creating a pellet culture of transgenic articular chondrocytes (0.25 × 10 6 cells pelleted in a 15-mL conical tube) and cultured in 1 mL of defined chondrogenic media for 21 days, 24 where reporter expression of the pellet was checked at day 21.
In situ fluorescence microscopy
At days 0 (preseeding, data not shown), 3, 7, 14, and 21 of culture, reporter cell fluorescence (N = 2) was captured using the Zeiss Axio Observer Z.1 inverted fluorescence microscope. Fluorescence was captured by tiling images over the entire scaffold surface in ZEN software at an objective of 5 × . For osteogenic culture, exposure times of 100 ms and 500 ms were applied for GFP (BSP-GFPtopaz) and RFP (DMP1-RFPmCherry) channels, respectively. For chondrogenic culture, an exposure time of 100 ms was applied for GFP (Col3.6-GFPtopaz and CFP (Col2a1-GFPcyan) channels, whereas 300 ms was applied for RFP (Col10a1-RFPmCherry). Tiled images were then stitched together to create a mosaic. Fluorescence intensity of all images were then equalized by adjusting individual pixel intensity histograms in Adobe Photoshop so that the middle slider was positioned to the right base of the main intensity peak, exposing all present fluorescent signal at equal intensity. Scaffold autofluorescence was accounted for qualitatively by capturing fluorescent images of unseeded scaffolds in all relevant fluorescent channels. Despite noticeable autofluorescence in the cyan channel, Col2a1-GFPcyan reporter activity was always visible above scaffold background.
Fluorescence-activated cell sorting analysis
At days 7, 14, and 21, cultured cells were harvested from scaffolds and subject to fluorescence-activated cell sorting (FACS) analysis. At a given time point, scaffolds were digested in an enzyme buffer containing 2 mg/mL collagenase D (Roche), 2 mg/mL hyaluronidase (Sigma), 20% (vol/vol) 0.25% trypsin-EDTA (GIBCO), and 10% penicillin–streptomycin in HBSS. Four sequential digestions were carried out at 37°C while shaking for 30 min. After each cycle, the digest was collected into sterile expansion media over ice. The digestion solution was then spun down and added to sterile 1 × PBS containing 2% FBS, filtered over a 70 μm nylon membrane, and resuspended in 400 μL of fresh 1 × PBS with 2% FBS for FACS analysis.
Cell suspensions were subject to FACS analysis (Becton-Dickinson FACSAria II) with a 130 μm nozzle equipped with lasers of 405 nm at 50 mW for CFP, 488 nm at 100 mW for GFP, and 561 nm at 40 mW for RFP. At least 5000 events of ungated events were collected. Percentages of the fluorescent population were calculated by normalizing GFP-, RFP-, or CFP-positive events to the entire positive fluorescent events population.
Real-time quantitative reverse transcription–polymerase chain reaction
At days 0, 7, and 21, mRNA from BMSCs and chondrocytes in superficial, transition/calcified cartilage, and osseous zones was extracted and subject to real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR). At a given time point, three (N = 3) scaffolds were removed from culture and separated into superficial, transition/calcified cartilage, and osseous scaffold zones under a stereomicroscope in a sterile hood. Scaffold zones were homogenized in 500 μL of TRIzol reagent (Life Technologies) over ice and frozen at −20°C until RNA extraction. Zonal RNA was extracted by phase separation using 20% (vol/vol) chloroform in TRIzol, followed by precipitation in isopropanol. The resulting total RNA pellet was rinsed with 75% ethanol and dissolved in DNAse digestion solution (New England Biolabs) to remove DNA contamination. The RNA was further purified by magnetic bead separation (RNAClean XP, Beckman Coulter) on a MiniMACS (Miltenyi Biotec) magnetic separator rack. The resulting nucleic acid yield was quantified using a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific) with purity and concentration determined from absorbance at 260 and 280 nm.
One hundred nanograms of total RNA was reverse-transcribed to cDNA using a MultiScribe Reverse Transcriptase with random primers (Applied Biosystems). qRT-PCR was performed on cDNA using the SYBR Green method (Bio-Rad). Forward and reverse primers used are listed in Table 1. Fold change of osteogenic markers (RUNX2, osteocalcin [OCN], alkaline phosphatase [ALP], BSP, and DMP-1) and chondrogenic markers (SOX9, aggrecan [AGG], type 1 collagen [COL1A1], type 2 collagen [COL2A1], and type 10 collagen [COL10A1]) relative to day 2 (osteogenic experiments) or day 0 (chondrogenic experiments) mRNA expression levels were calculated using the 2−ΔΔCT method, and are normalized to housekeeping gene GAPDH expression levels.
Cryohistology
At day 21, scaffold sections (N = 2) were fixed with 10% neutral-buffered formalin (Sigma) at 4°C overnight, rinsed with 1X PBS thrice, then soaked in 30% sucrose (Sigma) in deionized water at 4°C until samples became isotonic. Samples were then briefly patted dry and embedded in Optimal Cutting Temperature Compound (Tissue-Tek) by submersion in 2-methylbutane chilled with dry ice. Embedded samples were then cryosectioned to a thickness of 5 μm, adhered to cryotape (Cryofilm type II; Section-Lab, Co), and UV crosslinked onto glass slides.
For BMSC-seeded samples, sections were stained with Hematoxylin and Eosin for cell and tissue morphology, Vector Blue (Vector Blue Alkaline Phosphatase Substrate; Vector Laboratories) for ALP deposition, and von Kossa for calcium mineral deposits. For chondrocyte-seeded samples, sections were stained with Hematoxylin and Eosin, Vector Blue, von Kossa, and Safranin O–Fast Green for sulfated-proteoglycan deposition. Un-seeded scaffold sections were used as negative controls to test for background staining (data not shown), where minimal staining was observed. Sections were mounted with 30% glycerol in deionized water or 1 × PBS and color imaged with a Zeiss Axio Scan Z.1 automatic fluorescent slide scanning microscope with a 10 × objective.
Immunohistochemistry
Formalin-fixed chondrocyte-seeded scaffold sections (N = 2) from day 21 were subject to immunohistochemistry for protein deposition of the following markers: type 1 collagen, type 2 collagen, type 10 collagen, and Indian hedgehog homolog (Ihh). Tissue samples were first imaged to collect background fluorescence, then heated in antigen retrieval solution (2 mg/mL bovine serum albumin (BSA) in 1 × PBS) for 10 min at 42°C, rinsed in 1 × PBS, placed in 0.1% Triton X-100 in antigen retrieval solution for 10 min, rinsed in 1 × PBS, rinsed in 1 × Power Blocking Reagent (Power Block Universal Blocking Reagent; BioGenex) for 20 min, rinsed with 1 × PBS, then incubated with 1° antibody at 4°C overnight at optimized dilutions in antigen retrieval solution (goat polyclonal type 1 collagen, AB758, Millipore,1:100; mouse monoclonal type 2 collagen, II-II6B3, Developmental Studies Hybridoma Bank, 1:10; rabbit polyclonal type 10 collagen, AB58632, ABCAM, 1:100; rabbit polyclonal anti-Ihh, AB39634, ABCAM, 1:200; rabbit polyclonal anti-PRG4, NBP1-19048, Novus Biologicals, 1:50). Sections were then rinsed with 1 × PBS, covered with 1:200 dilution of Alexa Fluor 488-conjugated 2° antibody for 1 h at room temperature (Alexa Fluor 488 goat anti-rabbit IgG, A11008, Thermo Fisher Scientific; Alexa Fluor 488 donkey anti-goat IgG, A11055, Thermo Fisher Scientific; Alexa Fluor 488 goat anti-mouse IgG, A11001; Thermo Fisher Scientific), rinsed with 1 × PBS, then counterstained with 0.1% Hoechst (Hoechst 33342, trihydrochloride, trihydrate; Molecular Probes) in 50% glycerol in 1 × PBS. To check for nonspecific 2° antibody binding on scaffold materials, MZ sections were labeled with each variant of the 2° antibody (data not shown). Antibody fluorescence was imaged on a Zeiss Axio Scan Z.1 automatic fluorescent slide scanning microscope with a 10 × objective.
Subcutaneous implantation
Animal studies were conducted under approved protocols from the Institutional Animal Care and Use Committee. MZ scaffolds with and without articular chondrocytes were implanted subcutaneously in the backs of nontransgenic NSG mice for 8 weeks. First, zonal scaffolds cut to 6 × 3 × 3 mm (0.54 cm 3 ) were sterilized with ethylene oxide in a bench-top oven (AN74i Anprolene Sterilizer, Andersen Products) for 24 h, then vented overnight to remove residual ethylene oxide gas. Scaffolds were prepared for seeding as before (disinfected with 70% ethanol, soaked in expansion media), and seeded with 4.5 × 10 6 primary articular transgenic chondrocytes (passage 1) carrying Col3.6-GFPtopaz, Col2a1-GFPcyan, and Col10a1-RFPmCherry fluorescent reporter genes. Chondrocyte-seeded and unseeded (N = 2 per group) scaffolds were cultured for 1 week in vitro in defined chondrogenic media before implantation.
For surgery, mice were administered a preoperative intraperitoneal injection (5 μL/g of mouse) of ketamine and xylazine in 0.9% saline solution. Dorsal incisions 1 cm in length were made, where scaffolds were inserted in pouches. Dorsal pouches were sutured and mice were returned to their cages with a postoperative intraperitoneal injection (5 μL/g of mouse) of meloxicam at 1 mg/mL. Meloxicam was also administered 24 h following surgery. After 8 weeks, mice were sacrificed, and tissue was perfusion fixed with 10% NBF for histological analysis. Tissue was then subject to Hematoxylin and Eosin, Safranin O-Fast Green, Vector Blue, and von Kossa stains.
Statistical analysis
Statistical difference between three or more groups was calculated using a one-way ANOVA with a post-hoc Tukey test. Difference between two groups was calculated using a Student's t-test. Statistical significance was accepted at p < 0.05 (*). Analyses were performed in Minitab 16 software (Microsoft).
Results
BMSC osteogenesis
In situ fluorescence microscopy
Primary BMSCs carrying the fluorescent reporter combination BSP-GFPtopaz × DMP1-RFPmCherry were seeded atop both MZ and Healos scaffolds and cultured in osteogenic conditions for 21 days, where reporter fluorescence was evaluated in real-time at days 3, 7, 14, and 21 of culture. Reporter fluorescence is represented as intensity-adjusted data (Fig. 2A, B), which permits evaluation of all total fluorescence at each time point, and raw fluorescence data (Supplementary Fig. S1), which allows visualization of temporal changes in reporter activity (intensity).
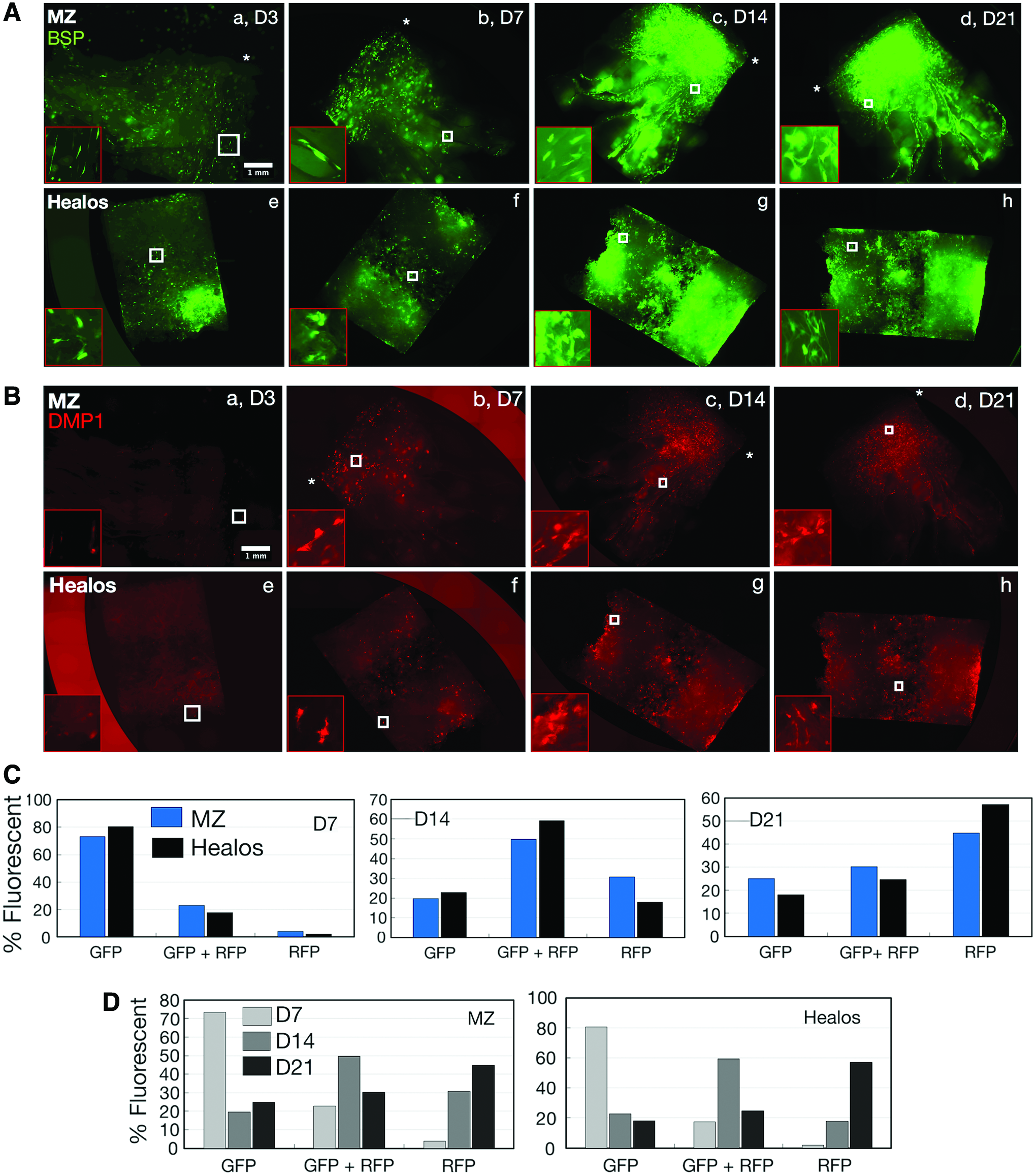
Intensity-normalized fluorescence of BSP-GFPtopaz
For both scaffolds, BSP-GFPtopaz fluorescence was present in few cells at day 3 (Fig. 2A, a, e), but dramatically increased up to day 21 (Fig. 2A, d, h). DMP1-RFPmCherry fluorescence was evident in just a few cells at day 3 for both materials (Fig. 2B, a, e), but became more prevalent at day 7 (Fig. 2B, b, f). Like BSP-GFPtopaz, the amount of cells expressing DMP1-RFPmCherry increased up to day 21, suggesting successful differentiation of BMSCs along the osteogenic lineage for both scaffolds. Notably, there was a large jump in reporter presence from day 7 to 14 for both reporters. From raw fluorescence data (Supplementary Fig. S1), the largest increase in reporter expression was observed from day 7 to 14, which corroborates the intensity-adjusted data. Concomitantly, the osteogenic capability of dual-reporter BMSCs was assessed by a 2D culture of BMSCs, and verified by the increased expression of DMP1-RFPmCherry at day 21 with corresponding mineral deposition (Supplementary Fig. S2A).
FACS analysis
To quantify the percentage of cells expressing a particular reporter gene, BMSCs were harvested at days 7, 14, and 21 and subject to FACS analysis (Fig. 2C, D). Percentages of GFP, RFP, or GFP+RFP-positive cells were quantified from the overall population of fluorescent cells. At day 7, the majority of fluorescence was attributed to GFP-positive cells, with smaller populations of GFP+RFP cells, and negligible RFP-only fluorescence. At day 14, a drastic shift in overall fluorescence occurred, with lowered GFP populations and increased GFP+RFP and RFP populations. At day 21, a shift toward increased RFP-only fluorescence was evident. Importantly, the RFP-only population continually increased to a maximum at day 21, whereas GFP+RFP populations peaked at day 14 and declined by day 21.
Real-time quantitative reverse transcription–polymerase chain reaction
To assay zone-specific gene expression, MZ scaffold zones were carefully sectioned for subsequent mRNA extraction. At day 7, ALP, BSP, DMP-1, and OCN were upregulated in all scaffold zones (Fig. 3A). Notably, BSP and OCN were significantly more upregulated for MZ scaffold zones compared with the other genes. Among scaffold zones at day 7, expression of ALP and OCN was significantly greater in the TZ/CZ and OZ than in the SZ. DMP-1 expression was significantly greater in the TZ/CZ compared with that in the SZ, whereas no significant increase in BSP expression was evident among scaffold zones. Healos was found to have significantly decreased expression of all markers compared with those in at least one zone at day 7. At day 21, expression of ALP, BSP, DMP-1, and OCN were upregulated for all scaffold zones, whereas just BSP, DMP-1, and OCN were upregulated for Healos. Cycle threshold (Ct) values are provided in Figure 3B, and give insight into the amount of the target gene used for fold-change quantification. A significant quantity of genes was measured, with a Ct range between 19.28 and 28.21.

Cryohistology
To assess matrix mineralization after osteogenic culture, MZ and Healos scaffold sections were subject to Vector Blue and von Kossa stains to assess ALP and calcium mineral deposition, respectively (Fig. 4). Both MZ and Healos scaffolds stained positively for ALP and calcium mineral, although the SZ of MZ scaffolds displayed significantly less ALP than all other zones. For all scaffolds, ALP deposition surrounded GFP and RFP signal. Von Kossa staining was even across all scaffold zones, and embedded with GFP+RFP BMSCs. A small quantity of RFP-only cells was present, highlighted by the white arrows in the CCZ. These data evince comparable matrix mineralization of BMSCs on MZ scaffolds to Healos.
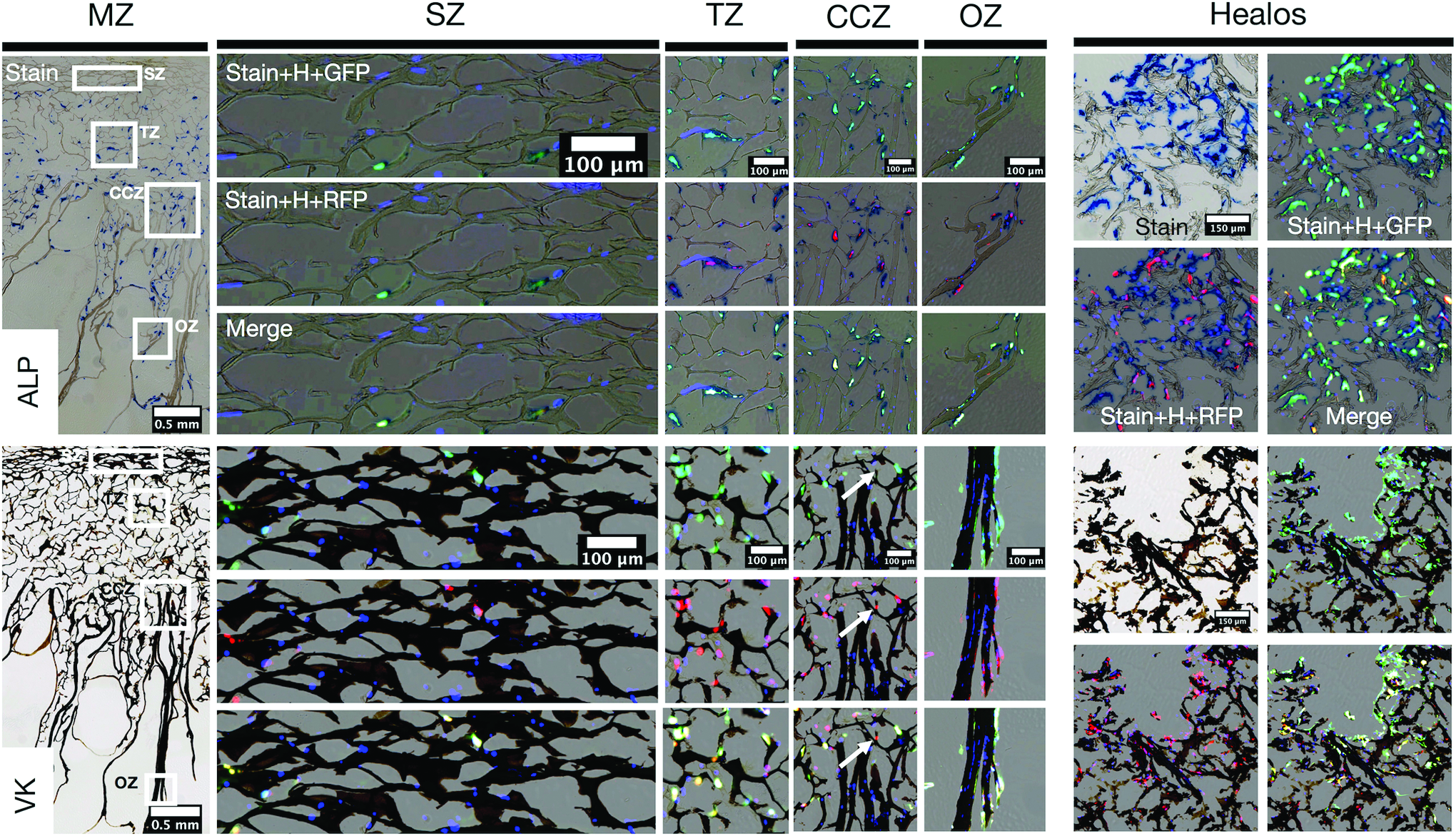
Vector Blue (ALP, upper panel) and von Kossa (VK, lower panel) staining of MZ and Healos scaffolds at day 21 of osteogenic culture. Stained MZ scaffold sections are presented alongside higher magnification images of the SZ, TZ, CCZ, and OZ. White squares indicate selected zonal regions. For each zone, the specified stain and Hoechst (H) are overlaid, with either GFP, RFP, or both (Merge). White arrows in VK staining of the CCZ indicate an RFP-only cell. Corresponding images of the Healos-positive control scaffold are presented on the right. ALP, alkaline phosphatase. Color images are available online.
Chondrocyte chondrogenesis
In situ fluorescence microscopy
Triple transgenic articular chondrocytes harboring the reporter combination Col3.6-GFPtopaz × Col2a1-GFPcyan × Col10a1-RFPmCherry were cultured on MZ, ICH, and IC scaffolds for 21 days in defined chondrogenic media to evaluate the chondrogenic character of the materials. Col3.6-GFPtopaz fluorescence (Fig. 5A) was present on all scaffolds at D3 and continually declined to day 21. Raw fluorescence data for the reporter (Supplementary Fig. S3A) revealed no visible reporter presence at 300 ms exposure time. The chondrogenic ability of murine BMSCs was also assessed, but deemed inadequate as a pellet culture in chondrogenic conditions failed to produce cartilage matrix as judged by a negligible Safranin O staining at 21 days (Supplementary Fig. S2B).
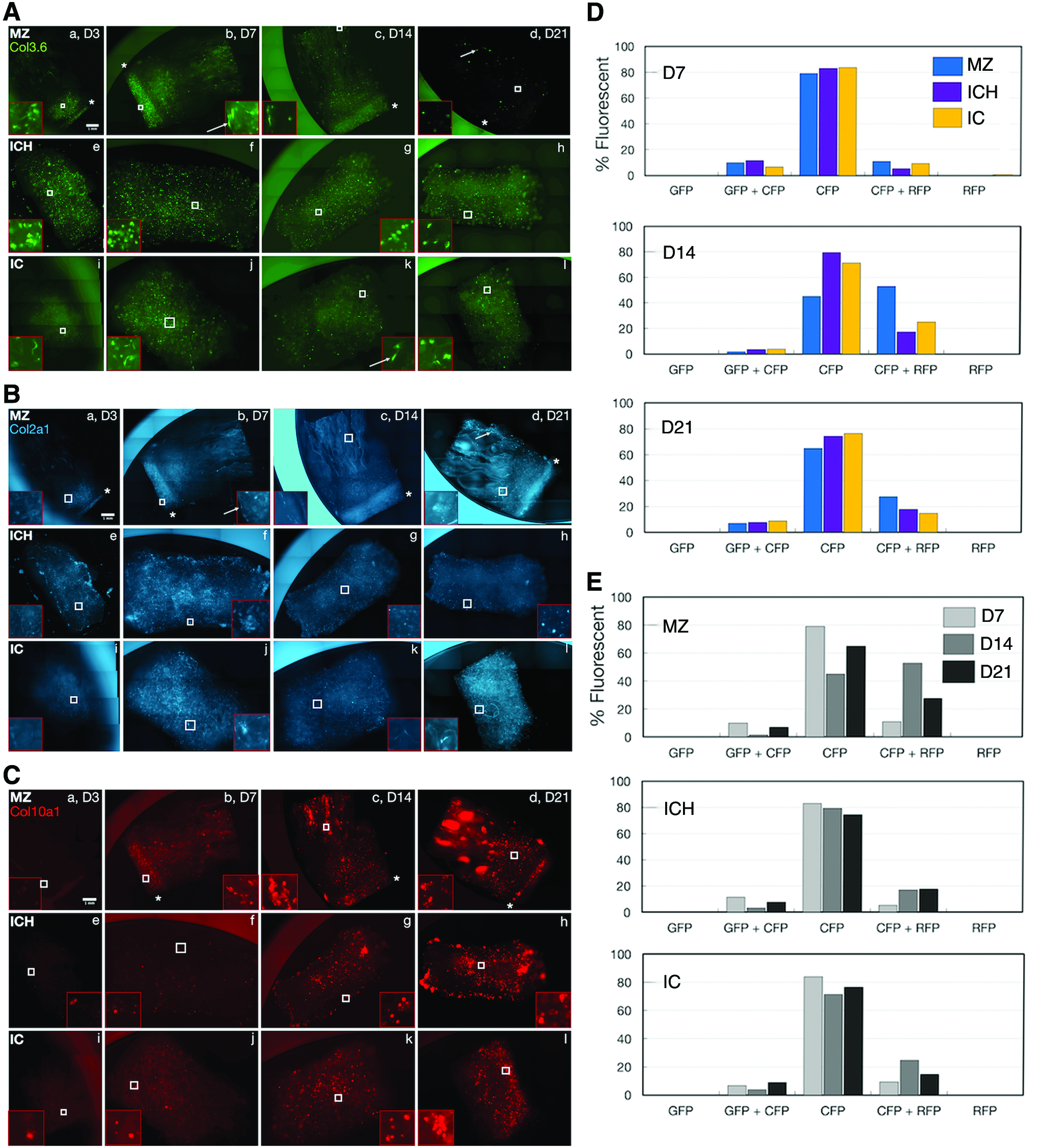
Intensity-normalized fluorescence of Col3.6-GFPtopaz
Unlike Col3.6-GFPtopaz fluorescence, Col2a1-GFPcyan fluorescence remained active for all scaffold types throughout the culture duration (Fig. 5B). Raw fluorescence data (Supplementary Fig. S3B) indicated that Col2a1-GFPcyan signal was stable for all scaffolds during culture.
Uniquely, Col10a1-RFPmCherry was absent at day 3 (Fig. 5C), but became active in all scaffolds at D7, where it was most widespread on MZ scaffolds (b). Col10a1-RFPmCherry presence steadily increased up to day 21 for all scaffolds, becoming most dense in the osseous zone of MZ scaffolds (d) and edges of ICH scaffolds (h). At a constant exposure time of 300 ms (Supplementary Fig. S3C), Col10a1-RFPmCherry continually increased for all scaffolds. The chondrogenic capability of triple-reporter chondrocytes was verified in pellet culture, where all three reporters were active at day 21 (Supplementary Fig. S4).
FACS analysis
Transgenic chondrocytes harvested at day 7 from MZ, ICH, and IC scaffolds (Fig. 5D, E) were nearly equally CFP positive (82% average), with small double-positive populations (averages of 9.4% and 8.5% for GFP+CFP and CFP+RFP populations, respectively). All scaffolds trended toward decreasing CFP in favor of increasing CFP+RFP populations, with negligible RFP-positive cells and minimal GFP-positive cells.
Real-time quantitative reverse transcription-polymerase chain reaction
Genetic markers for differentiation along the fibrocartilage (COL1A1), articular cartilage (COL2A1, AGG, and SOX9), and hypertrophic cartilage (COL10A1) lineages were analyzed at days 7 and 21 for MZ scaffold zones and IC and ICH scaffolds (Fig. 6A). At day 7, all markers, but SOX9, were upregulated for all materials. Expression of COL10A1 was significantly greater in the OZ compared with that in the SZ and TZ/CZ. Expression of hypertrophic marker COL10A1 was also significantly greater in the CZ/TZ and OZ than in IC, and IC and ICH scaffolds, respectively.
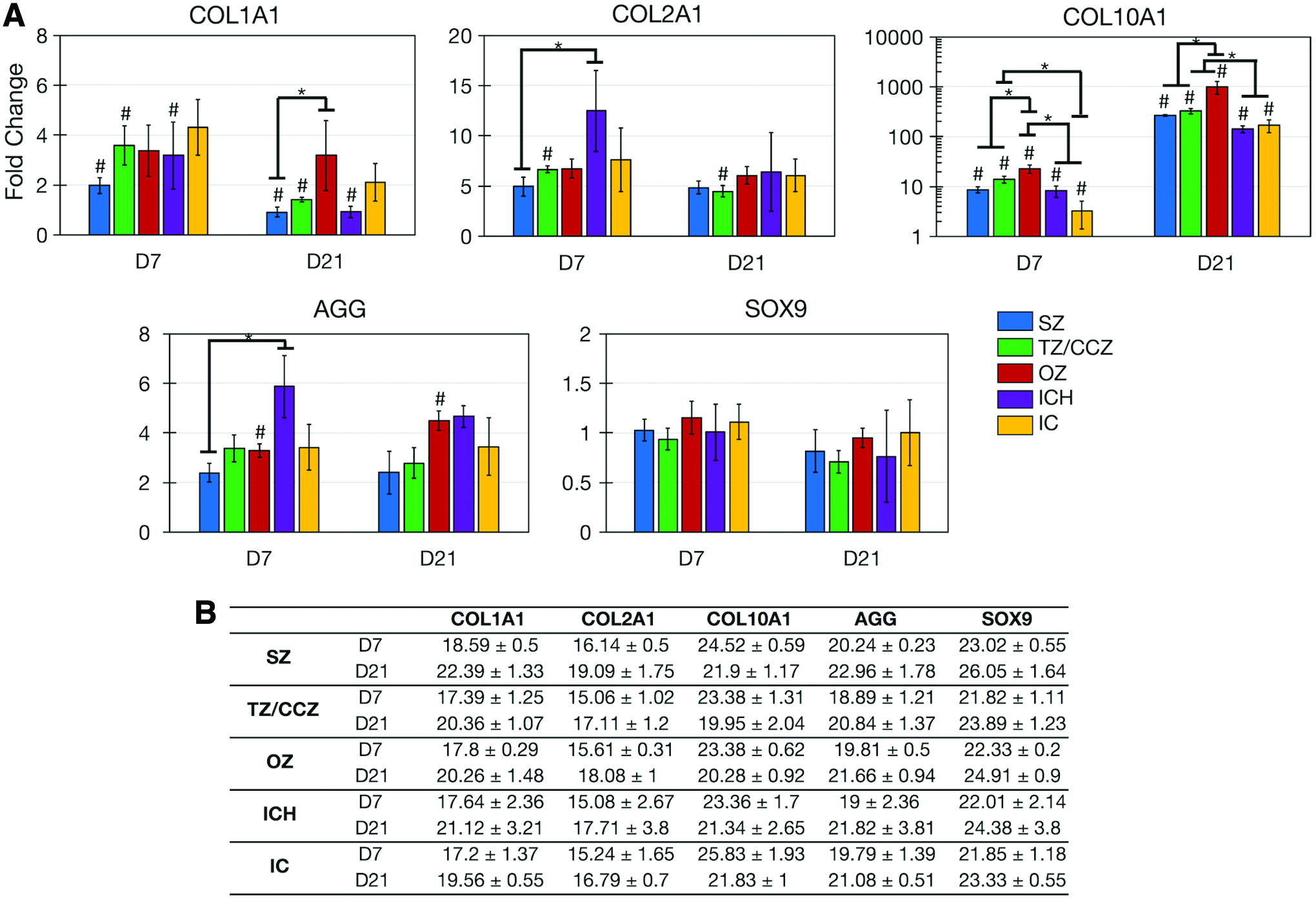
At day 21, COL2A1, COL10A1, and AGG were upregulated for all materials, whereas SOX9 remained stagnant. Most notably, COL10A1 expression had dramatically increased for all scaffolds by day 21. Expression was greatest in the OZ (reaching a nearly 1000-fold change by day 21) compared with all other scaffolds, and expression in the TZ/CZ was greater than that in IC and ICH scaffolds. Fold changes were determined from a significant amount of target gene, with Ct values between 15.06 and 26.05 (Fig. 6B).
Cryohistology
To assess the character of tissue that was deposited over chondrogenic culture, MZ (Fig. 7), IC (Supplementary Fig. S5), and ICH scaffolds (Supplementary Fig. S5) were cryosectioned at days 7 and 21 and subject to Hematoxylin and Eosin, Safranin O–Fast Green, Vector Blue, and von Kossa staining. Hematoxylin and Eosin staining showed a significant increase in both cellularity and matrix in all MZ scaffold zones, with new tissue aligned in the direction of the respective zone. Safranin O–Fast Green staining showed an increase in sulfated glycosaminoglycan deposition from day 7 to day 21 for all zones. At day 7, Safranin O staining was present in cellular aggregates in the TZ, CCZ, and OZ. At day 21, Safranin O staining was punctate in SZ and TZ, while it was denser and most abundant in the large cellular aggregates in the CCZ and OZ. This stain coincided with cells embedded in lacunae. All scaffold zones stained negative for ALP and calcium mineral at both time points.

Histological staining of MZ scaffold zones (SZ, TZ, CCZ, OZ) at days 7 and 21 of culture. H&E, Safranin O–Fast green, Vector Blue (ALP), and von Kossa stains were performed. H&E, Hematoxylin and Eosin. Color images are available online.
Immunohistochemistry
Zonal deposition of type 1 collagen, type 2 collagen, type 10 collagen, and Ihh at day 21 of culture was analyzed by immunofluorescence imaging. In addition, nuclei (Hoechst) and chondrocyte reporter fluorescence (Col2a1-GFPcyan and Col10a1-RFPmCherry) was overlaid with the immunofluorescence signal. No Col3.6-GFPtopaz signal was present across MZ scaffolds (Supplementary Fig. S6). Type 1 collagen (Fig. 8A) was present in all scaffold zones, but was most intense in the SZ compared with the TZ, CCZ, and OZ. Col2a1-GFPcyan-positive chondrocytes were mostly embedded inside this matrix, where Col10a1-RFPmCherry activity was coexpressed in hypertrophic cells.
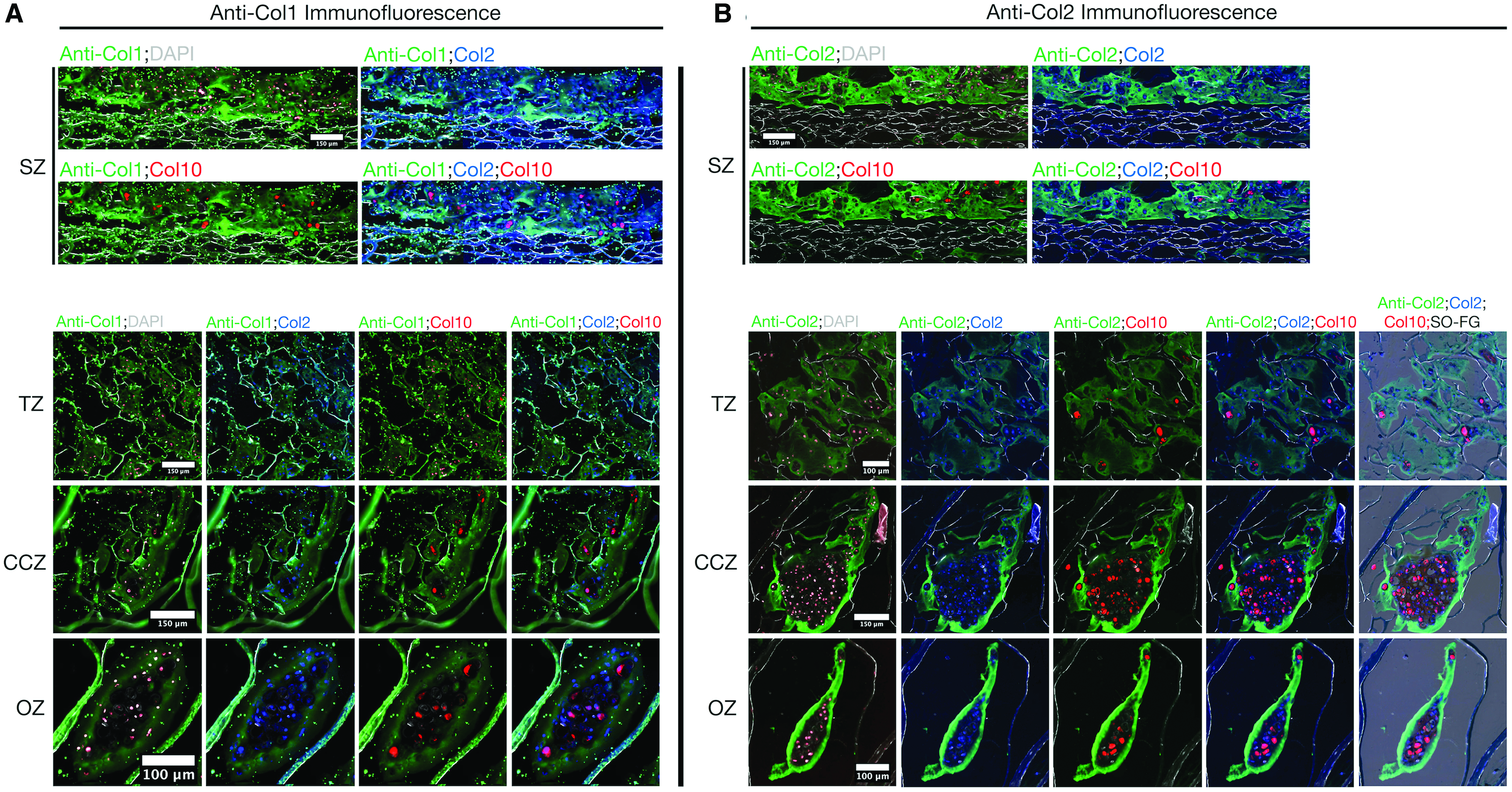
Immunofluorescence for zonal deposition of type 1 collagen
Type 2 collagen (Fig. 8B) filled the pore volume of all zones, where it was most prominent in the SZ, CCZ, and OZ. An abundance of type 2 collagen was aligned atop the SZ and embedded with mostly Col2a1-GFPcyan-positive chondrocytes. This same matrix composition was evenly deposited in the pores of the TZ. In the CCZ and OZ, aggregates of articular and hypertrophic chondrocytes were surrounded by a rich layer of type 2 collagen and embedded in a matrix abundant in sulfated glycosaminoglycans, as evident from the Safranin O overlay.
Hypertrophic markers, type 10 collagen and Ihh, were also assayed for in each zone (Fig. 9). Both proteins were evident in all zones, although at different intensities. Type 10 collagen matrix (Fig. 9A) was strongest in the OZ, where it encapsulated aggregates of Col2a1-GFPcyan and Co10a1-RFPmCherry-positive cells. Ihh was most prominent in cellular aggregates of the CCZ and OZ, where the greatest amount of Col10a1-RFPmCherry signal was also present. Ihh was localized with all Col2a1-GFPcyan and Col10a1-RFPmCherry fluorescence. In both IC (Supplementary Fig. S7A) and ICH (Supplementary Fig. S7B) scaffolds, deposited matrix was also a composite of types 1, 2, and 10 collagens and Ihh.
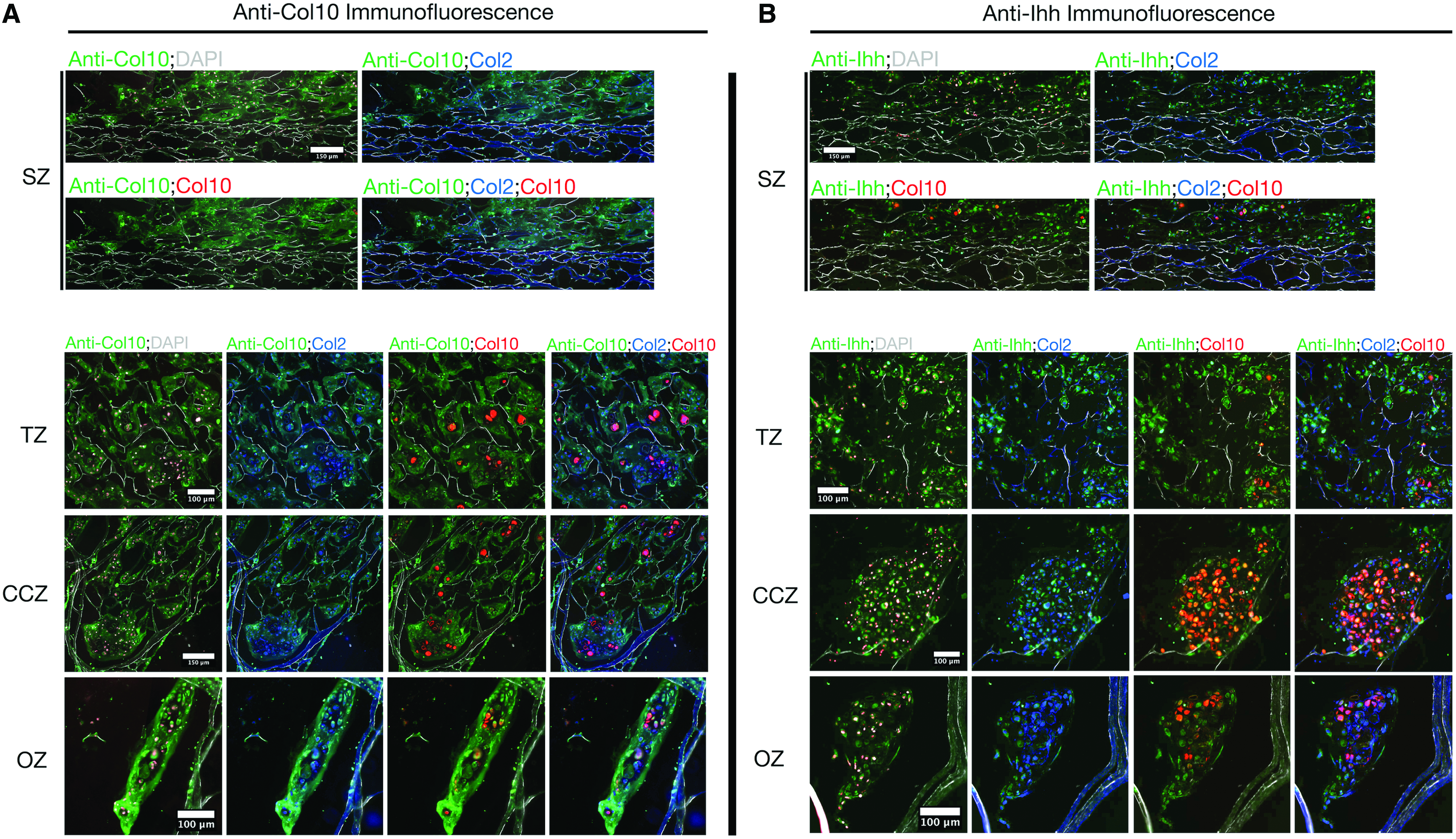
Immunofluorescence for zonal deposition of type 10 collagen
Subcutaneous implantation
Finally, we sought to evaluate the fate of chondrocyte-seeded MZ constructs in a vascularized environment in vivo (Fig. 10A). After 8 weeks of subcutaneous implantation, scaffolds were infiltrated with a fibrous-like tissue. Pockets of cellular aggregates rich in sulfated proteoglycan were present, and were comprised of cells harboring Col3.6-GFPtopaz, Col2a1-GFPcyan, and Col10a1-RFPmCherry reporters. Notably, some Col10a1-RFPmCherry fluorescence became independent of the other reporters in these aggregates.

Histological staining of chondrocyte seeded
Unlike the in vitro culture, harvested scaffolds stained positively for both ALP and calcium mineral. Both ALP and mineral nodules consisted of Col10a1-RFPmCherry fluorescence surrounded by Col3.6-GFPtopaz and Col2a1-GFPcyan fluorescence. Unseeded MZ scaffolds precultured in defined chondrogenic media for 1 week and implanted alongside the seeded scaffolds (Fig. 10B) displayed significant host cell infiltration and fibrous tissue infill, but lacked Safranin O, ALP, or mineral staining, indicating donor cells were required for proteoglycan and mineral formation. All observations were consistent between both scaffolds implanted per test group.
Discussion
In vitro cellular differentiation on biomaterials is conventionally assessed by destructive characterization modes, including gene expression analyses and histological staining. Such methods are not only destructive, but fail to provide spatiotemporal information of cellular maturity along a specific lineage. To overcome these limitations, we turn to the use of fluorescent multireporter cells that provide real-time readouts of cellular identity.
The Rowe laboratory has generated various lines of transgenic mice harboring GFP reporters to study bone and cartilage biology and pathology. To date, only one study has been published that describes the use of transgenic reporters to assess cellular differentiation on biomaterials; the COL1A1 2.3 kb fragment-driven GFP reporter was used as a readout of osteoblastic differentiation of murine BMSCs on carbonated hydroxyapatite. However, this reporter becomes active during late-stage osteoblast mineralization, failing to reveal early commitment to the osteoblastic lineage or other lineages. 25
As a first step to evaluating the regenerative potential of our MZ scaffolds, we have chosen to separately study osteogenesis and chondrogenesis of transgenic murine BMSCs and chondrocytes, respectively, by primarily monitoring the fluorescence of multiple transgenic reporters. For osteogenesis, BMSCs from mice containing both BSP-GFPtopaz and DMP1-RFPmCherry reporters were harvested. BSP, a noncollagenous matrix protein that nucleates hydroxyapatite growth, is expressed in early differentiated osteoblasts, 26 whereas DMP-1 is a phosphoprotein present in mineralized mouse, rat, and chicken tissues.27,28 For successful osteoblastic differentiation, we expect to see a transition from BSP-GFPtopaz to DMP1-RFPmCherry fluorescence. The combination of these BSP and DMP-1-promoter-driven GFPs, therefore, permits the progressive evaluation of maturity along the osteoblast lineage.
For chondrogenesis, we chose the triple reporter combination of Col3.6-GFPtopaz x Col2a1-GFPcyan x Col10a1-RFPmCherry to provide readouts of multilineage chondrocyte progression. The Col3.6-GFPtopaz transgene contains a 3.6 kb DNA fragment of the COL1A1 promoter that activates at basal levels in preosteoblasts and intensifies during osteoblastic differentiation. It has also been shown to activate during differentiation into tendon, skin, and lung tissues. It was also active in dermal fibroblasts and has been used to label fibrochondrocytes in the murine temporomandibular joint.14,16,17,29 The Col2a1-GFPcyan transgene activates CFP upon Col2a1 promoter activation and has been shown to be expressed in articular cartilage.16,29,30 Lastly, the Col10a1-RFPmCherry transgene produces an RFP upon Col10a1 promoter activation and has been used to label hypertrophic chondrocytes in the temporomandibular joint.17,29 For chondrogenic differentiation, we would expect decreasing Col3.6-GFPtopaz fluorescence as chondrocyte dedifferentiation diminishes, alongside a transition from Col2a1-GFPcyan to Col10a1-RFPmCherry as chondrocytes mature. Hence, this multireporter system is suited for validating the chondrocyte phenotype on our MZ scaffolds under chondrogenic conditions.
Guided primarily by the use of these reporters, we have concluded the following about our MZ scaffolds: (1) MZ scaffolds were able to support the differentiation of both BMSCs and chondrocytes in vitro, and (2) subtle differences in zone-specific cellular phenotype and tissue were identified.
Indeed, MZ scaffolds promoted the differentiation of BMSCs to a degree similar to Healos, as evidenced at the reporter (continued progression from BSP-GFPtopaz to DMP1-RFPmCherry fluorescence, Fig. 2), genetic (upregulation of BSP and DMP-1, Fig. 3), and protein (zonal deposition of ALP and calcium mineral, Fig. 4) levels. As expected, the chondral layers did not outright prohibit osteogenic differentiation. However, zones containing hydroxyapatite demonstrated the greatest osteogenic ability, as judged by the enhanced expression of ALP, BSP, DMP-1, RUNX2, and OCN relative to other scaffold zones and Healos (Fig. 3). This effect is presumably influenced by PO43− ion release and subsequent A2b adenosine receptor activation. 31 Dissolution of Ca2+ ions may also promote osteogenic differentiation through intake through L-type calcium channels. 32
Importantly, the use of the reporters was able to uniquely identify the phenotype of seeded BMSCs unlike gene expression or histological analyses. At the end of culture, roughly half of fluorescent cells had progressed to sole DMP1-RFPmCherry fluorescence, with depletion in BSP-GFPtopaz and double-positive fluorescent populations. This observation suggests that BMSCs continually matured along the osteoblastic lineage throughout the culture period. At the protein level, DMP-1 is processed into C-terminal and N-terminal fragments, where the former mediates osteocyte maturation and mineralization, whereas the latter controls phosphate homeostasis. 33 It is important to note that DMP1 is also present in the nuclear compartment of undifferentiated/early stage osteoblasts 34 ; it is possible that DMP1 fluorescence is contributed by these cells. However, we believe that we are mostly achieving a late osteoblast/early osteocyte phenotype from a large population of cells from the progressive loss in BSP-GFPtopaz fluorescent population and increase in DMP1-RFPmCherry fluorescence (FACS, Fig. 2C, D), along with the appearance of robust ALP and von Kossa staining at day 21 of culture (Fig. 4). Furthermore, previous studies have demonstrated weak expression of the utilized DMP1-RFPmCherry reporter in osteoblasts and pronounced expression in osteocytes.35,36
MZ scaffolds supported the maturation of triple-reporter articular chondrocytes to a hypertrophic-like phenotype over the 21-day culture period. This progression to hypertrophy was not unique to our MZ scaffolds, but also present on IC and ICH scaffolds, as well as pellet culture. These data indicate that hypertrophic induction is a function of the culture conditions rather than the chosen substrate. Under standard chondrogenic conditioning with TGF-β, MSCs express hypertrophic markers, such as type 10 collagen, ALP, and MMP13.37,38 Furthermore, the presence of TGF-β1 and dexamethasone may not totally prohibit terminal differentiation, but slow its progression.37,38 In our experiments, we did not see histological deposition of ALP, or matrix mineralization (Fig. 7), which is a unique phenotype reminiscent of costal cartilage chondrocytes.37,39 In these cells, type 10 collagen is solely expressed, where ALP activates during puberty and initiates matrix mineralization. Mineralization will not occur without the presence of additional systemic proteins, such as insulin-like growth factor or triiodothyronine; despite the expression of type 10 collagen, costal cartilage is still of hyaline character. 40 Therefore, we believe that some of our cultured chondrocytes (27.7% of reporter cells, Fig. 5E) have reached a prehypertrophic stage of maturity, but not terminal maturation. These chondrocytes are stable at day 21 and still retain an articular-like phenotype, as judged by continued upregulation of COL2A1 (Fig. 7A) and significant deposition of type 2 collagen matrix throughout the scaffold (Fig. 8). Further, in support of this conclusion is the finding that the majority (>65%) of total fluorescent chondrocytes was solely Col2a1-GFPcyan positive for all scaffolds. There was a negligible percentage of cells found to be solely Col10a1-RFPmCherry positive, indicating that terminal maturation was not achieved. These data highlight that our MZ scaffolds may largely support an articular-like phenotype during in vitro culture.
To pin down the phenotype of the Col2a1-GFPcyan+Col10a1-RFPmCherry cells at the end of the culture period, we performed immunohistochemistry for Ihh, an indicator of prehypertrophic chondrocytes. Indeed, we saw wide-spread cytoplasmic Ihh. In prehypertrophic chondrocytes, Ihh inhibits the onset of endochondral ossification by activating endogenous parathyroid hormone-related protein expression, which in turn suppresses Ihh activity in a negative-feedback loop. 38 In fact, it has been shown that TGF-β's function to suppress hypertrophy is mediated through endogenous Ihh activity; TGF-β signaling induces phosphorylation of SMAD2/3, which binds to SMAD4, then induces Ihh transcription. Paracrine/autocrine Ihh signals subsequently induce SOX9 expression. Concomitantly, endogenous TGF-β3 and BMP-2 activity are induced by TGF-β1, which cause further SMAD2/3 phosphorylation, Ihh transcription, SOX9 expression, inducing Col10a1 expression. 38 Thus, it is likely that TGFβ1 may be acting to maintain the prehypertrophic phenotype through Ihh signaling.
Our next step was to study the stability of our chondrocyte-seeded MZ scaffolds in a vascularized environment in vivo, with particular interest in studying the fate of the prehypertrophic chondrocyte population. Chondrocyte-seeded MZ scaffolds were cultured in chondrogenic media for 1 week before subcutaneous implantation to establish an initial prehypertrophic chondrocyte population (Fig. 5C, b). We had hypothesized that prehypertrophic chondrocytes would follow an endochondral ossification regime upon exposure to signaling molecules, such as insulin-like growth factor and thyroid hormone, present in the vasculature. Indeed, after 8 weeks of implantation, MZ tissues stained positively for ALP and calcium mineral (Fig. 10). A telling sign of terminal maturation was the observation of exclusive Col10a1-RFPmCherry fluorescence, present in the center of cellular aggregates (Fig. 10, white arrows). The presence of significant Col3.6-GFPtopaz reporter activity localizing with mineralized nodules and ALP deposits may be indicative of osteoblastic differentiation, as it is unlikely for fibrochondrocytes to localize with osseous structures. This subcutaneous implantation follows an endochondral ossification-like process, where chondroprogenitors aggregate, differentiate into hypertrophic chondrocytes, and mineralize to provide a template for bone formation.6,39 Furthermore, the subcutaneous environment has been validated as a suitable model for studying endochondral ossification.41,42 Ultimately, this progression along endochondral ossification may be leveraged as an in vivo template for construction of subchondral bone in the OZ.
Our second conclusion from the study was that zonal scaffold properties lead to subtle differences in cellular phenotype and tissue. First, significant differences in the extent of gene or protein production were found in each zone. At both the fluorescence and RNA level, Col10a1 was maximal in the OZ compared with other scaffold zones or reference IC and ICH scaffolds. The OZ deliberately contains the most hydroxyapatite (25 wt%) in the scaffold. 12 Hydroxyapatite has been found to promote expression of type 10 collagen of chondrocytes and MSCs in a dose-dependent manner, although the mechanism for which is unclear and mandates mechanistic understanding.43–45
Intensity of Safranin O staining also varied with zone and was greatest in the CCZ and OZ compared with the SZ and TZ. The large pore areas within the CCZ and OZ supported the formation of large cellular aggregates rich in sulfated glycosaminoglycans. These aggregates are indicative of ongoing chondrogenesis, where cell–cell interactions are favored. 6 Although aggregates were found in both the CCZ and OZ, they were larger and more spherical in the CCZ, surrounded directly by type 1 collagen–hyaluronic acid scaffold matrix. The negatively charged carboxyl groups of hyaluronic promote a spherical morphology of seeded chondrocytes, which may facilitate aggregation of chondrocytes into spherical aggregates shown.9,46 In contrast, aggregates in the OZ are elongated with the direction of the mineralized collagen scaffold lamellae, where cell–matrix interactions are favored in the absence of hyaluronic acid. All aggregates were colocalized with abundant Col10a1-RFPmCherry signal (Figs. 7–9), indicating that these aggregates were undergoing hypertrophic differentiation. The high proteoglycan content may serve as binding sites for hydroxyapatite nucleation, ultimately leading to mineralization as observed in the in vivo data. 47
Lastly, direction of neotissue was templated by the scaffold pores, as shown in immunohistochemical data (Figs. 8 and 9), and formed zone-specific alignment. The ability of the tissue to adapt the surrounding pore morphology is a powerful tool to ensure the reconstruction of new tissue that recapitulates native zonal organization. It is to be expected then that this orientation shall remain once the scaffolding biodegrades in vivo, as we have shown in our previous studies.48,49
However, chondrogenic culture of triple-reporter chondrocytes led to the formation of a composite matrix of type 1, type 2, and type 10 collagen in all scaffolds, regardless of zonal matrix orientation or composition (Figs. 8 and 9, Supplementary Fig. S7A, B). It is our understanding that the major components of the produced matrix are type 2 and type 10 collagens, produced from the majority of chondrocytes that were either only Col2a1-GFPcyan or Col2a1-GFPcyan+Col10a1-RFPmCherry positive. These data highlight that soluble factors present in the differentiation media are the most critical mediators for cellular phenotype and tissue formation. It will be important in future studies to quantify the amount of protein deposited in each zone to further distinguish zonal tissue composition.
Several limitations of the current study exist that warrant addressing. First is the discrepancy between day 21 osteogenic gene expression data and fluorescence/histological data. As observed in Figure 3, expression of all bone markers declined from day 7 to day 21. This is in direct conflict with the observation that both BSP-GFPtopaz and DMP1-RFPmCherry reporters increased in both number and intensity continually throughout culture. It is likely that osteogenic cells were buried under a heavily mineralized matrix (Fig. 3), making RNA retrieval difficult. This points to the limitations of qRT-PCR and the advantages of reporter fluorescent evaluation; in a nonquantitative manner, it may replace qRT-PCR analyses. Second, the SZ fostered limited cellular infiltration and subsequent matrix deposition. Instead, laterally-aligned tissue was formed on the surface of the SZ in an area with less-compressed scaffold pores, which remains from the scaffold fabrication process. The lack of matrix formation in this zone is most likely attributed to cell seeding, as cells were seeded on both surfaces of the scaffold; cell and tissue presence relies on cellular migration to the scaffold interior. Furthermore, when implanted in vivo, an abundance of laterally-aligned fibrous tissue was formed inside the SZ by host cells (Fig. 10). Additional work will call upon an improved seeding procedure to ensure adequate infiltration of seeded chondrocytes in this layer, while increasing sample size for in vivo experiments.
The fluorescent reporter culture system described in this work holds potential as a powerful diagnostic tool for scaffold and/or technique development. In our work, for example, we may seek to optimize culture conditions and/or scaffold chemistry that may arrest chondrocyte hypertrophy in the SZ and TZ by screening for Col2a1-GFPcyan fluorescence exclusive of Col10a1-RFPmCherry fluorescence over the culture period. Moreover, the nondestructive and versatile nature of this reporter system can be extended to optimize and evaluate the performance of a plethora of biomaterial systems both in vitro and in vivo.
Conclusions
Altogether, the utilization of reporter cells endows us with a powerful tool to nondestructively evaluate cellular phenotype in situ. Furthermore, this GFP activity remains stable postculture. Using cryohistological protocols, reporter activity may be overlaid with histological or immunochemical analysis, as evinced in this study. In our work, we have shown successful BMSC osteogenic differentiation on our MZ scaffolds from the progressive activation of exclusive BSP-GFPtopaz to DMP1-RFPmCherry reporters. Chondrocyte maturity was verified by observing a continual transition from Col2a1-GFPcyan exclusivity to Col2a1-GFPcyan+Col10a10-RFPmCherry fluorescence, suggesting a population of cells became prehypertrophic. This was confirmed by histological and immunohistochemical analyses. Subtle differences in cellular phenotype and zonal tissue were identified. The combination of these reporters with MZ scaffolds may guide their continued development for the reconstruction of functional osteochondral tissue.
Footnotes
Acknowledgments
The authors would like to acknowledge NSF Grants (CBET-1133883, CBET-1347130, and Grant #1639914) for their support.
The authors would also like to acknowledge Ms. Theresa Samuels for rat tail donation, Mr. Andrew Nguyen and Mr. Simon Du for assistance with scaffold fabrication, Ms. Li Chen for assistance with histological procedures, Ms. Lisa Wu for management of mice used in the study, Mr. Tapan Dalal for assistance with qRT-PCR, Dr. Evan Jellison for assistance with FACS analysis, and Dr. Mark Kronenberg for insightful discussions.
Disclosure Statement
Dr. Wei has an ownership interest in OrteoPoniX LLC, which is developing and commercializing biomaterials for orthopedic applications.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.